Классификация и филогения / Курс (original) (raw)
Классификация и филогения
Любое научное исследование начинается с идентификации объектов. Для этого их классифицируют по свойствам (признакам) и строят систему объектов, руководствуясь определёнными целями. При ограниченном наборе свойств обычно пользуются комбинативными системами, в которых всем выделенным признакам (например, А и а) соответствуют все возможные их комбинации:
(А + а) (А + а) = АА + 2Аа + аа, или в виде матрицы
По положению объекта в системе можно заранее предсказывать его свойства, но если число свойств и объектов особенно велико, то построение комбинативных систем становится невозможным: например, для объектов, различающихся по 100 признакам, требуется уже 10 000 комбинаций.
Классификация бесчисленного множества насекомых (более миллиона видов), различающихся многими признаками, возможна лишь при использовании других подходов. Объединяя отдельные виды в роды, роды в семейства, семейства в отряды, отряды в классы и так далее, мы получаем иерархическую систему соподчинённых групп (таксонов), характеристики которых обобщают все свойства включённых в эти группы объектов. При этом пользуются правилом конгрегации, суть которого в том, что всякий вид одного рода ближе к своему сородичу, нежели к любому виду другого рода. Эти системы используют не только в энтомологии, они стали стандартом для всех органических форм.
Система животного царства есть объективное описание многообразия животных, то есть сходств и отличий между ними, в первую очередь между видами и далее соответственно между родами, семействами и всеми остальными таксонами. Таким образом, система базируется на огромном числе фактов. Как обобщение всего поддающегося анализу материала о животных она представляет собой итог и показатель уровня развития зоологии и вместе с тем является основой для всех, более специальных исследований, поскольку изучение любого животного начинается с определения его систематической принадлежности.
Вряд ли могут быть сомнения в том, что система животных в той мере, в какой она отражает реальное распределение таксонов и приближается к естественной системе, является результатом исторического развития животного мира, его эволюции. Однако при трактовке проблемы «система и эволюция» возникают особые сложности: ведь система строится на основе непосредственно наблюдаемых фактов (чаще всего морфологических), тогда как все наши суждения об эволюции имеют характер более или менее веских предположений. Эти суждения могли бы основываться на реальных фактах палеонтологии, но фрагментарность и неполнота геологической летописи не позволяют установить ни общего предка, ни переходных форм с необходимой достоверностью. При этом обычно пытаются на основании реальных сходств и отличий между современными (рецентными) видами (и другими таксонами) реконструировать филогенетическое древо: ось времени представляют вертикалью, а по горизонтали выстраивают сходства и отличия форм. Для демонстрации этой процедуры (рис. 199) воспользуемся простейшим случаем и в соответствии с уже известным нам правилом конгрегации распределим виды (в виде отдельных точек), сравниваемые по двум количественным признакам, например числу глаз или ног.

Рисунок 199. Система, построенная всего по двум признакам:
N1 и N2 — признаки, выраженные в численной форме; 1 — границы родов; 2 — границы семейств; 3 — границы отрядов; _точки характеризуют виды
Группы точек (видов), ближайшие друг к другу и очерченные на рисунке общим контуром, формируют роды, включённые, в свою очередь, в контуры семейств, и так далее. Здесь сходства и различия видов по признакам откладываются по двум осям координат. Разумеется, чтобы получить не столь упрощённую, а более полную систему, надо учитывать многие признаки и строить модели со многими координатами, располагая виды (точки) в многомерном пространстве. При этом многообразие всех существующих сегодня видов было бы приведено в систему, близкую к естественной. Вполне понятно, что для каждого геологического периода следовало бы строить сходные системы, чему, однако, препятствуют и неполнота геологической летописи, и трудности с критериями вида в палеонтологии. Тем не менее такие построения необходимы.
Располагая системы разных геологических периодов во времени, по вертикали, мы видим, что они являются сечениями филогенетического древа (рис. 200). Здесь каждый ствол или пучок стволов пересекается на разных уровнях, и плоскости сечений дают системы взаимно соподчинённых таксонов.
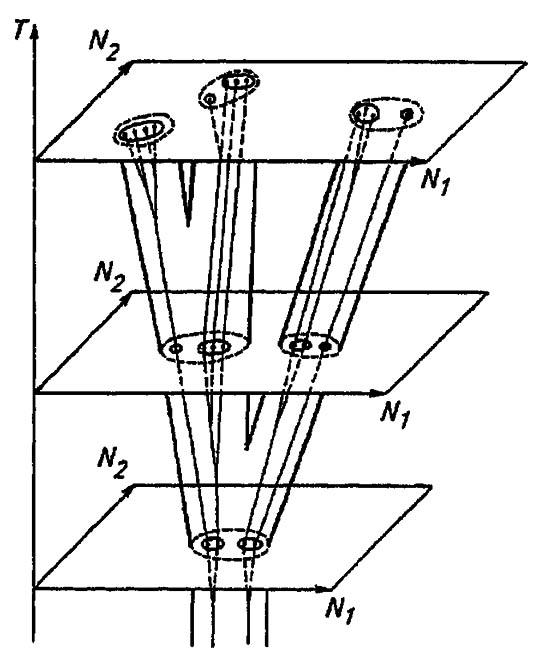
Рисунок 200. Эволюционное древо и системы для разных геологических эпох как его сечения:
Т — ось времени. N1 и N2 — признаки, выраженные в численной форме; 1 — границы родов; 2 — границы семейств; 3 — границы отрядов; _точки характеризуют виды
Обращаясь к построенному древу, необходимо сделать методологически очень важное замечание. Работая с системами современных и ископаемых форм и восстанавливая ход эволюции, мы не должны сообразовываться с существующими гипотезами и представлениями о ходе эволюции. Дело в том, что, имея какие-либо предварительные суждения в этом плане, мы рискуем «подогнать под ответ» наши выводы, внося поправки в системы, которые мы строим. Между тем многие со времён Чарльза Дарвина убеждены, что в каждое суждение необходимо вводить эволюционный аспект, полагая, что сходство видов является синонимом родства, а общий прототип — есть общий предок.
Многие, говоря о естественной системе, подразумевают только ту, которая построена на основании уже известной эволюционной истории форм, но дело обстоит иначе — эволюционная история известна очень плохо: мы судим об эволюции по системе, а не о системе по эволюции.
Известно, что не все формы животных сохраняются в ископаемом состоянии (например, паразиты); ни для одного из современных типов животных предки не установлены, и палеонтология не даёт материалов об их происхождении; до сих пор мы не имеем прямых свидетельств о происхождении насекомых, восполняя их отсутствие разного рода гипотезами. Между тем, располагая системой современных форм, можно с большей или меньшей определённостью судить об исходной примитивности или же, наоборот, производности тех или иных видов и общих тенденциях их эволюции. Очевидно, что примитивность и производность различаются по числу эволюционных этапов, реализованных видом: чем меньше их, тем он более примитивен. Рассматривая и комментируя положение насекомых в системе органических форм, мы отмечали примеры примитивности членистоногих разных групп, такие, как исходно водный образ жизни, однообразие сегментов тела и их придатков, несовершенство локомоции и т. п. При этом общими тенденциями их эволюции являются стабилизация и сокращение числа сегментов тела, выделение сегментарных комплексов — тагм и пополнение переднего из них — головы — за счёт прилежащих сегментов туловища, редукция и преобразование конечностей брюшного отдела и тому подобного. В соответствии с этим кольчатые черви примитивнее многоножек, которые в этом отношении примитивнее насекомых, из которых наиболее совершенны высшие двукрылые и перепончатокрылые.
Если расположить современные виды в соответствии с их примитивностью (числом реализованных этапов), то одна из осей на сечении древа (см. рис. 200) станет осью примитивности, вторая же — осью их различий и сходств. Далее следует повернуть нашу схему так, чтобы ось примитивности совпадала с осью времени (рис. 201, Б), и признать, что примитивные формы скорее всего и являются более древними. Они сохранили те черты организации, которые ещё не были превзойдены иными формами, когда ни одна группа животных ещё не прошла ни одного из высших этапов (рис. 201, А). Это заключение — единственное основание реконструкции эволюционного древа при отсутствии палеонтологического материала.

Рисунок 201. Построение эволюционного древа, исходя из системы организмов:
А — связь между древностью таксонов и их примитивностью (по Фотгу, 1975). По вертикальной оси отложены время и (одновременно) уровни организации, по горизонтальной — примитивность современных форм. Таксоны изображены дужками. Виды каждого таксона могут вымереть (1), перейти на следующий уровень организации (2) или сохраниться до современности без изменений (3). Уровни организации на оси времени — суть этапы эволюции, отличающиеся все меньшей примитивностью; Б — схема преобразования оси примитивности в ось времени, при котором графически изображённая система переходит в эволюционное древо; Т — ось времени; Р — ось примитивности; V — ось сходства между видами. Уровни примитивности показаны линиями, параллельными оси V.
Между тем выявление всех свойств и признаков вида связано с многочисленными трудностями и с неизбежным субъективизмом при их сравнении. Для объективной оценки сходства и различия используют разные приёмы. Один из них назван нумерической таксономией и связан с кодированием признаков (и их сочетаний) на матрицах для последующей компьютерной обработки. Принимая все признаки равными друг другу, это направление исследований не учитывает их реальной разнокачественное, хотя в некоторых случаях может оказаться полезным. Более соответствует реальности таксономический анализ, использующий математический приём «взвешивания» признаков по их оригинальности. Однако столь оригинальный признак, как отсутствие антенн у бессяжковых и у личинок мух, искусственно сближает эти далёкие друг от друга формы, противопоставляя их всем другим насекомым.
Так или иначе, но формализация традиционных приёмов работы специалиста-систематика обычно основывается на признании тождественности сходства объектов и их родства. Однако следует отметить, что не всякое сходство определяется родством и может служить его мерой и критерием. Например, развитие хватательных ног у богомолов и некоторых сетчатокрылых или прыгательных бёдер у прямокрылых и блох происходило в каждой группе насекомых независимо и не свидетельствует об их родстве. Сходство этих структур определяется сходством выполняемых ими функций, то есть является конвергентным. Ещё более нагляден пример конвергентного сходства в деталях строения центральной нервной системы насекомых и позвоночных животных, общим прототипом которых могли быть только примитивные кишечнополостные животные (или их личинки) с диффузным сплетением нервных клеток. Впрочем, можно предполагать, что отмеченный параллелизм в строении ганглиев и нервных стволов, рефлекторных дуг, синапсов и рецепторов у насекомых и позвоночных выражает общие тенденции эволюции нервной системы от общей примитивности к специализации. Однако однажды описанный отпечаток крыла ископаемого насекомого, который при более внимательном рассмотрении оказался отпечатком крыловидного придатка семени растения, показывает ещё одну возможную категорию сходств, названных системными. Примеры такого рода изоморфизма и их анализ составляют предмет общей теории систем, интенсивно разрабатываемой философами, математиками и биологами.
Открытие Д. И. Менделеевым периодической системы химических элементов побудило многих биологов к построению периодических (параметрических) систем организмов, но предпринятые попытки не привели к ожидаемым результатам. Лишь ряды гомологичной изменчивости растений Н. И. Вавилова можно рассматривать как явное достижение в этом плане.
Наряду с идентификацией объектов систематика сталкивается с проблемой различения (дискриминацией) близких видов. Эта проблема особенно актуальна в энтомологии, так как среди насекомых имеется много видов-двойников, неотличимых друг от друга по явным признакам. Например, у постоянно используемых в генетических исследованиях представителей вида Drosophila pseudoobscura Fr. было обнаружено неодновременное наступление половой зрелости. По этому признаку внешне неотличимые друг от друга формы были условно названы расой А и расой В, но последующие наблюдения и эксперименты выявили ещё более значительные различия между ними. В результате были признаны разные виды: D. pseudoobscura Fr. и D. persimilis Dobzh.
Учение о виде (эйдология) как важнейший раздел биологической систематики в связи с многообразием насекомых приобрело в энтомологии особое значение. Обычно используют следующее определение этого понятия: вид — это совокупность сходных особей, имеющих определённый ареал и дающих при скрещивании плодовитое потомство, сохраняющее сходство с родителями. Однако четырём перечисленным свойствам вида противопоставляются свойства видов-двойников и другие свойства, требующие специального обсуждения.
Определяя понятие «вид», мы основываемся на том, что те или иные его свойства не совпадают со свойствами других видов. Например, Drosophila pseudoobscura Fr. отличается от D. persimilis Dobzh. размерами крыльев (варьирование крылового индекса: от 45,7 до 62,8 у первого и от 68,8 до 76,2 у второго вида). Таким образом, для видов-двойников важны не столько различия между ними, сколько их обособленность друг от друга, не плодовитость потомков, а реальная изолированность родителей.
Виды-двойники отмечены как у двукрылых, так и у чешуекрылых. Зерновая моль (Tinea granella L.) имеет четыре вида, из которых только два вредоносны; листовёртка Choristoneura fimiferana Hbn. по характеру пищевой специализации подразделяется на два вида. Среди жесткокрылых пример вида-двойника — рисовый долгоносик (Calandra oryzae L.): один из них питается кукурузой, второй — пшеницей Виды-двойники зафиксированы у прямокрылых, некоторых муравьёв, тлей и ногохвосток.
Большинство видов насекомых все же различаются по достаточно явным, специфическим признакам, но вместе с тем они далеко не всегда представлены вполне однородными особями. Приведённые выше примеры полиморфизма выражают разнокачественность популяций. Многие хорошо известные виды имеют свою собственную внутреннюю структуру, проявляющуюся с большей или меньшей определённостью в системе внутривидовых форм.
Например, перелётная саранча (Locusta migratoria L.) имеет весьма обширный ареал и в соответствии с разнообразием условий существования в его отдельных зонах дифференцируется на подвиды. Основной подвид — азиатская перелётная саранча (Locusta migratoria aziatica L.) — населяет умеренную зону Евразии; в Западной Европе его сменяет подвид L. migratoria gallica Rem., в Нечернозёмной зоне европейской части в б. СССР обитает среднерусская саранча (L. migratoria rossica Uv. et Zol.), в Юго-Восточной Азии — восточная перелётная саранча (L. migratoria manilensis Мс.), а в тропической Африке — тропическая перелётная саранча (L. migratoria migratorioides Reich, et Fairm.). В данном случае подвиды представляют собой географические расы, различающиеся между собой устойчивыми, но подчас нерезкими особенностями морфологии и экологии.
Наряду с подвидами система внутривидовых форм нередко классифицируется более дробно — на отдельные экологические расы, или экотипы. Обычно экотипы не имеют стойких морфологических особенностей, не разобщены в пространстве. Например, ивовая и берёзовая расы ивового листоеда (Lochmaea capreae L.) различаются по кормовым растениям и по требованиям к микроклимату: особи берёзовой расы предпочитают относительно сухие и хорошо освещённые местообитания, ивовой расы — более влажные и тенистые. Те и другие способны скрещиваться между собой, хотя в реальных условиях существования вероятность встреч половых партнёров, обитающих в одном биотопе, значительно выше.
Очевидно, что существование внутривидовых форм свидетельствует о динамичности структуры видов, которая выражает их приспособляемость к переменчивым условиям жизни.
Весьма важные характеристики вида — его уникальность и уникальность занимаемой им экологической ниши. Выявление всех свойств и признаков вида сопряжено со многими трудностями и с неизбежным субъективизмом в оценках и сравнениях.
Обилие и разнородность объектов, их соответствие условиям существования и постоянное воспроизводство в цепи сменяющих друг друга поколений придают биологической систематике особую сложность и значение. Систематика — это начало и венец всякого биологического исследования; ее конечной целью может стать лишь естественная система организмов.
Кропотливая работа энтомологов-систематиков привела к созданию вариантов классификации насекомых. Различаясь в деталях, они имеют более или менее непротиворечивую и стабильную основу, что позволяет сделать вывод о прогрессе, достигнутом в исследованиях, и о близости их результатов к тому порядку, который реально существует в природе, то есть к естественной системе организмов.
В исторической ретроспективе первоначально можно отметить обобщения Аристотеля (384–322 годы до нашей эры), который разделил все известное ему многообразие насекомых на три группы.
1. Крылатые — Pterota (современные отряды: Coleoptera — часть, Orthoptera, Heteroptera, Lepidoptera, Neuroptera, Hymenoptera, кроме муравьёв, и Diptera).
2. Крылатые и подобные им бескрылые — Pterota simuli et Aptera (муравьи и жуки-светляки).
3. Бескрылые — Aptera (все остальные членистоногие).
Наиболее радикальные преобразования в архаичную классификацию Аристотеля были внесены Я. Сваммердамом (1637–1685), впервые предложившим разделять насекомых по способу превращения на Metabolia и Ametabolia, соответствующих современным Holometabola и Hemimetabola.
В трудах Карла Линнея мы находим уже семь отрядов, выделенных по структуре крыльев: Coleoptera (современные Coleoptera и Orthoptera), Hemiptera, Lepidoptera, Hymenoptera, Diptera, Neuroptera (разнородные отряды) и Aptera (бескрылые насекомые наряду с многоножками, паукообразными и некоторыми ракообразными). Вскоре после Линнея Ф. Брауер показал гетерогенность группы Aptera и противопоставил первичнобескрылым насекомым (Collembola и Thysanura) все формы, утратившие крылья вторично (вши, блохи, пухоеды).
Таким образом, к началу нашего столетия сформировались общие принципы и подходы к классификации насекомых, в которую были введены признаки метаморфоза, наличия или отсутствия крыльев и их структуры, а также признаки строения ротовых органов.
Основы современной классификации были заложены в более поздних исследованиях А. Гандлирша, Ф. Сильвестри, Г. Вебера и А. В. Мартынова.
Заслуга А. Гандлирша состоит в выделении из сборного отряда прямокрылых — уховёрток и тараканов, а из ложносетчатокрылых — стрекоз, подёнок, веснянок и трипсов. Доведя общее число отрядов насекомых до 30, этот исследователь свёл их в надотрядные комплексы, введя комплекс панорпоидных насекомых, объединяющий, с одной стороны, двукрылых и блох, а с другой — ручейников и чешуекрылых.
Разносторонние исследования Ф. Сильвестри привели к открытию двух новых отрядов: Protura и Zoraptera, а проведённая Г. Вебером ревизия системы насекомых в целом — к выделению четырёх подклассов: Collembola, Protura, Thysanura и Pterygota, причём подкласс Thysanura был подразделён на два отряда — Thysanura Ectotrophi и Thysanura Entotrophi, в зависимости от положения челюстей снаружи или внутри головной капсулы. В группу Entotrophi некоторые авторы включали также Collembola, Protura и Diplura.
Система крылатых насекомых А. В. Мартынова в редакции 1928 года имела следующий вид.
Отдел I. Palaeoptera: Odonata, Ephemeroptera
Отдел II. Neoptera:
подотдел Polyneoptera:
- Blattopteroidea: Blattodea, Mantodea, Isoptera
- Orthopteroidea: Orthoptera, Phasmodea, Plecoptera, Embioidea
- Dermapteroidea: Dermatoptera, Hemimeroidea
- Thysanoptera: Thysanoptera
подотдел Paraneoptera:
- Corrodentia: Psocoptera, Zoraptera, Mallophaga, Anoplura
- Rhynchota: Homoptera, Heteroptera
подотдел Oligoneoptera:
- Coleoptera, Strepsiptera, Megaloptera, Neuroptera, Rhaphidioptera, Hymenoptera
- Mecoptera, Diptera, Aphaniptera, Trichoptera, Lepidoptera
В более поздней редакции 1938 года А. В. Мартынов перевёл комплекс Thysanoptera в подотдел Paraneoptera, что было положительно воспринято современными исследователями.
Такая классификация насекомых сохранилась до наших дней без существенных видоизменений. Приведём один из наиболее удачных её вариантов — систему Г. Я. Бей-Биенко, в которой зафиксированы все современные отряды и их комплексы.
I. Подкласс низшие, или первичнобескрылые, — Apteiygota
А. Инфракласс энтогнатные — Entognatha
- Отряд протуры, или бессяжковые, — Protura
- Отряд подуры, или ногохвостки, — Podura
- Отряд диплуры, или двухвостки, — Diplura
Б. Инфракласс тизануровые — Thysanurata
4. Отряд тизануры, или щетинохвостки, — Thysanura П.
Подкласс высшие, или крылатые, — Pteiygota
А. Инфракласс древнекрылые — Palaeoptera
5. Отряд подёнки — Ephemeroptera
6. Отряд стрекозы — Odonatoptera
Б. Инфракласс новокрылые — Neoptera
Отдел с неполным превращением — Hemimetabola
Надотряд ортоптероидные — Orthopteroidea
7. Отряд таракановые — Blattoptera
8. Отряд богомоловые — Mantoptera
9. Отряд термиты — Isoptera
10. Отряд веснянки — Plecoptera
11. Отряд эмбии — Embioptera
12. Отряд гриллоблаттиды — Giylloblattida
13. Отряд палочники — Phasmoptera
14. Отряд прямокрылые — Orthoptera
15. Отряд гемимериды — Hemimerida
16. Отряд кожистокрылые — Dermaptera
17. Отряд зораптеры — Zoraptera
Надотряд гемиптероидные — Hemipteroidea
18. Отряд сеноеды — Psocoptera
19. Отряд пухоеды — Mallophaga
20. Отряд вши — Anoplura
21. Отряд равнокрылые — Homoptera
22. Отряд клопы — Hemiptera
23. Отряд трипсы — Thysanoptera
Отдел с полным превращением — Holometabola
Надотряд колеоптероидные — Coleopteroidea
24. Отряд жуки — Coleoptera
25. Отряд веерокрылые—Strepsiptera
Надотряд нейроптероидные — Neuropteroidea
26. Отряд сетчатокрылые — Neuroptera
27. Отряд верблюдки — Raphidioptera
28. Отряд большекрылые — Megaloptera
Надотряд мекоптероидные — Mecopteroidea
29. Отряд скорпионовые мухи — Mecoptera
30. Отряд ручейники — Trichoptera
31. Отряд бабочки — Lepidoptera
32. Отряд перепончатокрылые — Hymenoptera
33. Отряд блохи — Aphaniptera
34. Отряд двукрылые — Diptera
Этот вариант системы вошёл во многие учебники и руководства по энтомологии. Однако некоторые его положения подвергают критике. В частности, введение в класс насекомых отрядов Protura, Diplura и Collembola (Podura) оправдывается только стремлением к сохранению традиций. Ведь по многим признакам эти организмы сближаются с многоножками и как противопоставление Thysanura — истинным первичнобескрылым насекомым — заслуживают выделения в самостоятельные классы. Имеются веские аргументы против объединения подёнок и стрекоз в отдел древнекрылых. Весьма далёкие друг от друга, они не являются древнекрылыми в прямом смысле слова, так как древнейшие крылатые насекомые имели признаки новокрылых. Трудно также признать перепончатокрылых представителями надотряда мекоптероидных. Более частные возражения касаются самостоятельности отряда веерокрылых, вводимого на правах подотряда в отряд жуков.
Наиболее радикальные преобразования системы А. В. Мартынова предлагают палеоэнтомологи. Основываясь на строго датированных отпечатках ископаемых форм, можно проследить, как от самых примитивных крылатых насекомых — Protoptera — отошли две эволюционные ветви: Gryllones и Scarabaeones. Первые вернулись в крупные скважины субстратов под пологом леса, испытали эмбрионизацию развития, приобрели способность складывать крылья плоско на брюшке и в конечном счёте дали начало подотделу Polyneoptera системы А. В. Мартынова. Вторые, сохраняя долгое время повадки и морфологию Protoptera, питающихся спорангиями голосеменных, впоследствии дали начало предкам современных подёнок, затем — предкам современных стрекоз и наконец общему предку — Paraneoptera и Oligoneoptera.
Работа по систематике насекомых продолжается. Современная система насекомых совершенствуется на основе сопоставления разных фактов, мнений исследователей.
Отряды насекомых мы рассмотрим, не претендуя на исчерпывающие характеристики, стремясь лишь к оценке своеобразия каждого из них. Однако прежде всего обратимся к многоножкам — низшим Atelocerata, которые рассматриваются как ближайшие предшественники насекомых.