Genetic Alterations of the Retinoblastoma-Related Gene RB2/p130 Identify Different Pathogenetic Mechanisms in and among Burkitt’s Lymphoma Subtypes (original) (raw)
Abstract
Alterations of cell cycle-associated genes probably contribute to the pathogenesis of Burkitt’s Lymphoma (BL), in addition to c-myc translocation. Mutations disrupting the nuclear localization signal of the retinoblastoma-related gene RB2/p130 have been documented recently in BL cell lines and primary tumors. Given the importance of the RB2/p130 gene in controlling cell growth, mutations of this gene may result in uncontrolled cell proliferation. We tested the expression and genomic organization of the RB2/p130 gene in relation to the proliferative features of a series of BL samples collected from the endemic and sporadic regions, regardless of whether the samples were acquired immune deficiency syndrome (AIDS)-related. The expression of the Rb2/p130, p107, and cell proliferation-related proteins (cyclin A and B) was determined by immunohistochemistry. The structures of exons 19 through 22 of the RB2/p130 gene, encoding for the B domain and C terminus, were analyzed by polymerase chain reaction (PCR) analysis and single-strand conformation polymorphism (SSCP) technique. The direct PCR products were sequenced to identify the actual mutations. Our results suggest that BL is composed of a mixture of molecular types with distinct genetic and phenotypic patterns, probably resulting from different pathogenetic mechanisms. In endemic BL, the RB2/p130 gene is mutated in most of the cases, and the protein is restricted to the cytoplasm. In AIDS-related BL, high levels of nuclear expression of the wild-type pRb2/p130, p107, and cell proliferation-related proteins were detected. This finding is in line with the molecular mechanisms observed in virus-linked oncogenesis. Sporadic BLs were mainly characterized by the low nuclear values of the wild-type pRb2/p130 and, conversely, the high values of p107. The increased cell proliferation due to different alterations of cell growth control by Rb-related proteins may be the first step in lymphomagenesis, during which additional genetic changes, including missense mutations of c-myc, may subsequently occur.
In 1958, Burkitt first described a disorder associated with jaw tumors in African children. 1 In 1961, the neoplasm was identified as a form of malignant lymphoma, and what had emerged as a clinical syndrome became a pathological entity called Burkitt’s lymphoma (BL). 2 Through clinical-epidemiological studies, 3 it became evident that BL can occur as an Epstein-Barr virus (EBV)-associated malignancy, with endemic BL (eBL) occurrence among children in the malaria belt of equatorial Africa and sporadic BL (sBL) occurrence in other geographical areas, where it also occurs among adults. 4,5 The tumor as seen in endemic areas most often involves facial bones, 5 whereas the terminal ileum and lymph nodes are the more common sites in sBL. 6,7 The morphological similarity between eBL and sBL probably relates to their similar origin from germinal-center B cells. 8 In the Revised European-American Lymphoma (“REAL”) classification of lymphoid neoplasms, 10 eBL and sBL are collectively reported as BL because of their isomorphism, although differences between the two are acknowledged in terms of EBV incidence, 6 type of translocation involving the c-myc proto-oncogene, 9 and cell kinetics. 11 Furthermore, with the spread of the acquired immune deficiency syndrome (AIDS) epidemic, BL has frequently been reported as a common neoplasm in HIV-infected patients. 12 These lymphomas, which may be better termed “AIDS-related BL,” 13 usually display an activation of c-myc by chromosome translocations that show structural similarities to those found in patients with sBL. 14 Still the vast majority of AIDS-related BL in Western countries are EBV-negative, 14 whereas in Africa they are strongly associated with EBV. 15
The above listed differences between eBL, sBL, and AIDS-related BL do not mean that each of these subtypes of lymphoma may represent a perfectly homogenous entity, nor is there reason to believe that sBL does not also occur in endemic regions. 6,13 Some not very strict associations between JH and DH recombinations at 14q32 with distant 5′ c-myc recombinations in the African cases of BL versus Sμ and Sα recombinations at 14q32 with nearby 5′ or intronic c-myc recombinations at 8q24 in sporadic cases have been identified. 16 Thus, it seems probable that BL is composed of a mixture of molecular types and that the incidence of each subtype may depend on environmental factors. 6,17 The African type and the non-African type of BL may represent the opposite ends of this spectrum.
In addition, it should also be considered that the c-myc translocation is not the only genetic lesion found in BL 6 and that myc_-transformed cells are usually characterized by the loss of expression of several genes. 18 Cooperating alterations of cell cycle-associated genes probably contribute to the pathogenesis of BL. 6,19 p53 mutations have been found in 30% to 40% of BL samples, 20 whereas the majority of these lymphomas with the wild-type p53 might have lesions in other growth suppressor genes. 21 Genetic alterations disrupting the nuclear localization of the retinoblastoma-related gene RB2/p130 have recently been documented in BL cell lines as well as in primary tumors. 22 These mutations may impair the ability of the respective gene product to function as a growth suppressor nuclear protein, pRb2/p130 being exclusively cytoplasmic._ pRb2/p130 is a member of the Rb family that includes pRb/p105 and p107_._ 23 Whereas pRb/p105 is found in both quiescent and proliferating cells, pRb2/p130 and p107 expression is peculiar during the cell cycle. 24 These proteins act in a cell cycle-dependent manner to regulate the activity of a number of important cellular transcription factors, such as E2FS. 25,26 In this paper, we investigated the expression of pRb2/p130 and p107 in relation to the proliferative features of a series of BL, collected from endemic and sporadic areas, to assess whether alteration of the control mechanisms by these proteins may contribute to the pathogenesis of BL.
Materials and Methods
Selection of Cases and Conventional Histology
Most tumor biopsy specimens in paraffin wax blocks were obtained from 24 patients with BL observed at the Nairobi Hospital, Kenya, from 1995 to 1998. These cases were selected because they came from hot, moist woodland and savanna country in the Lake Victoria region and from coastal lowlands with holoendemic malaria. Five additional cases of BL were retrieved from the Institute of Pathological Anatomy and Histology, University of Siena, Italy.
The stains used for the qualitative histological evaluation included hematoxylin and eosin, Giemsa, periodic acid/Schiff reagent, and Gomori’s silver impregnation. The diagnoses were established, with knowledge of the immunohistochemical results, after an independent evaluation by and consensus among three pathologists, based on the REAL Classification. 10 Information on the age, sex, and site of biopsies was obtained. Clinical records, including the clinical stage, treatment, and follow-up, were not available in most of the African cases, because they had been sent for a histological diagnosis to the Department of Pathology at the Nairobi Hospital from other districts. Clinicopathological data, including EBV and human immunodeficiency virus (HIV) infection status, are summarized in Table 1 ▶ . The HIV status, assessed by an enzyme-linked immunoabsorbant assay on patient serum, was available in four cases; the HIV status was also tested by polymerase chain reaction (PCR) in all of the cases.
Table 1.
Clinical-Epidemiological Characteristics of BL Summarized in Relation to Molecular and Immunohistochemical Data
| Case | Origin† | Age | Sex | Site‡ | EBV | HIV | RB2/p130 exon no.§ | % Nuclear staining¶ | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 19 | 20 | 21 | 22 | pRb2/p130 | p107 | |||||||
| eBL | ||||||||||||
| 1 | A | 2 | M | Jaw | + | − | Wt | Wt | Wt | Wt | 10% | 60% |
| 2 | A | 4 | M | Mandible | + | − | Mut | Wt | Wt | Wt | NV | NV |
| 3 | A | 5 | M | Jaw | + | − | Wt | Wt | Mut | Wt | Neg | 35% |
| 4 | A | 5 | F | Mandible | + | − | Wt | Wt | Mut | Wt | Neg | 25% |
| 5 | A | 6 | F | Jaw | + | − | Wt | Wt | Mut | Wt | Neg | 40% |
| 6 | A | 6 | M | Mandible | + | − | Wt | Wt | Mut | Wt | Neg | 15% |
| 7 | A | 8 | M | Orbit | − | − | Mut | Wt | Mut | Wt | Neg | 12% |
| 8 | A | 9 | M | Mandible | + | − | Wt | Wt | Mut | Wt | Neg | 23% |
| 9 | A | 9 | F | Mandible | + | − | Wt | Wt | Wt | Wt | 5% | 55% |
| 10 | A | 10 | M | Jaw | + | − | Wt | Mut | Mut | Wt | Neg | 5% |
| 11 | A | 11 | M | Mandible | + | − | Wt | Mut | Mut | Wt | Neg | 30% |
| 12 | A | 13 | F | Femur | + | − | Mut | Wt | Mut | Wt | Neg | 10% |
| 13 | A | 13 | M | Mandible | − | − | Wt | Wt | Mut | Wt | Neg | 18% |
| sBL in endemic regions | ||||||||||||
| 14 | A | 3 | M | Ileum | + | − | Wt | Wt | Wt | Wt | 15% | 65% |
| 15 | A | 5 | F | Ileum | + | − | Wt | Wt | Wt | Wt | 35% | 70% |
| 16 | A | 16 | M | LN | + | − | Wt | Wt | Wt | Wt | 5% | 65% |
| 17 | A | 18 | M | LN | − | − | Wt | Wt | Wt | Wt | 20% | 70% |
| 18 | A | 20 | F | LN | − | − | Wt | Wt | Wt | Mut | Neg* | 30% |
| 19 | A | 32 | M | Ileum | − | − | Wt | Wt | Mut | Wt | Neg | 5% |
| sBL | ||||||||||||
| 20 | I | 16 | M | LN | − | − | Wt | Wt | Wt | Wt | 7% | 95% |
| 21 | I | 23 | F | Ovary | + | − | Wt | Wt | Wt | Wt | 20% | 60% |
| 22 | I | 27 | M | Cecum | − | − | Wt | Wt | Wt | Wt | 5% | 70% |
| 23 | I | 36 | M | Ileum | − | − | Wt | Wt | Wt | Wt | 10% | 75% |
| AIDS-related BL | ||||||||||||
| 24 | A | 23 | M | Ileum | + | + | Wt | Wt | Wt | Wt | 80% | 70% |
| 25 | A | 33 | F | Ileum | + | + | Wt | Wt | Wt | Wt | 90% | 70% |
| 26 | A | 23 | M | LN | + | + | Wt | Wt | Wt | Wt | 85% | 90% |
| 27 | A | 26 | M | Ileum | + | + | Wt | Wt | Wt | Wt | 75% | 95% |
| 28 | A | 56 | M | Ileum | + | + | Wt | Wt | Wt | Wt | 70% | 80% |
| 29 | I | 33 | M | LN | − | + | Wt | Wt | Wt | Wt | 60% | 70% |
Immunohistochemistry
The immunophenotyping on paraffin sections was performed using a large panel of antibodies to detect formalin-resistant epitopes of malignant lymphoma cells and by the Envision +/HPR method (Dako, Milan, Italy). 27 For antigen retrieval, deparaffinized 3- to 4-μm sections were treated with microwaves or by pressure cooking in 1 mmol/L ethylenediaminetetraacetic acid, pH8.0, for 5 minutes, followed by cooling at room temperature before incubation with the antibodies. 28 The negative controls were obtained by replacing the primary antibodies with normal mouse serum. Normal human tonsils served as positive controls.
Immunostaining of the cell cycle regulator proteins examined in this study was carried out as described above. Anti-cyclin B-1, AB-1 (clone V152), and anti-cyclin A (clone 6E6) were obtained from Neomarkers (Fremont, CA). Anti-Rb2 monoclonal antibody (clone 10) was obtained from Transduction Laboratories (Lexington, KY). Anti-Rb2 and anti-p107 polyclonal antibodies were generated in one author’s (A. G.’s) laboratory and used as previously described. 29,30
In all sections, the cells exhibiting a positive immune reaction to a given antibody were counted in a randomly chosen high-power field (see below) of lymphoma tissues, and the results were expressed as percentages of all of the neoplastic cells in those areas. Intra- and interobserver reproducibility of the counts was ≈95%. The intensity of the immune reactions and the nuclear and cytoplasmic localization of the stain were estimated qualitatively.
Molecular Analysis
DNA was extracted from the deparaffinized tissue by using a salting-out procedure and was shown to be suitable for PCR by the amplification of a β-globin DNA region. 31
Mutational Screening
The structure of exons 19 through 22 of the RB2/p130 gene, encoding for the B domain and C terminus were analyzed by PCR SSCP analyses. The PCR mix (50 μl) contained genomic DNA at the final concentration of 4 ng/μl, 0.2 mmol/L of each of the four deoxynucleotide triphosphates, 2 U of Klen _Taq_I (Ab Peptides, St. Louis, MO) and the intron primer panels (exon 19: 5′-AGGTCCTATCACCAAGGGTGT-3′; exon 19 rev: 5′-GCTTAGTTACTTCTTCAAGGC-3′; exon 20: 5′-GAGAAAGTTAATATCCTAGCTG-3′; exon 20 rev: 5′-GTGAATGGTCCATATATAAATCA-3′; exon 21: 5′-TGGTTTAGCACACCTCTTCAC-3′; exon 21 rev: 5′-GCTTAGCACAAACCCTGTTTC-3′; exon 22: 5′-CTGAGCTATGTGCATTTGCA-3′; exon 22 rev: 5′-AAGGCTGCTGCTAAACAGAT-3′) at the final concentration of 0.4 μmol/L each. Thirty-five cycles of denaturation (95°C, 1 minute), annealing, (55°C, 1 minute), and extension (72°C, 1 minute) linked to one cycle at 72°C for 7 minutes were carried out in a thermal cycler (Perkin-Elmer, Norwalk, CT).
For the SSCP analysis, a mutation detection enhancement (MDE) gel solution (FMC BioProducts, Rockland, ME) supplied as 2× liquid concentrate was used. PCR products (2 μl) were denatured and run through the MDE gel at 8 W of constant power for 8 hours at 15°C, in 0.6× running buffer (90 mmol/L Tris, 64.6 mmol/L boric acid, 2.5 mmol/L ethylenediaminetetraacetic acid, pH 8.3) as described by Cinti et al 22 The bands were visualized using a silver staining kit (PlusOne DNA Silver Staining Kit, Pharmacia Biotech, Uppsala, Sweden).
The PCR products were resolved on a 1.5% ethidium bromide-stained agarose gel. Bands were cut from the gels. The DNA was purified using the QUIAquick gel extraction kit (Qiagen, Santa Clarita, CA) and used for automated DNA sequencing using dideoxyterminator reaction chemistry for sequence analysis on the Applied Biosystem model 373A DNA sequencer.
Detection of HIV Infection
HIV DNA was amplified by nested PCR using the lentivirus universal primer pair UNIPOL1/2 32 as outer primers (25 cycles) and the degenerate primers UNIPOL3 (5′GAAACAGGAMRRGAGACAGC 3′) and UNIPOL4 (5′ TTCATDGMTTCCACTACTCCTTG 3′) as inner primers (30 cycles). This nested primer set, when used at a low-stringency annealing, specifically amplifies all HIV1, HIV2, and HIV0 pol sequences known to date. The PCR products were visualized by ethidium bromide staining of agarose gels. The specificity of the products was first assessed by direct sequencing and confirmed in each run by the appropriate positive and negative controls. Sensitivity was determined to be 1 target copy for the nested HIV PCR, as assessed by quantitative experiments with titrated cloned targets.
In Situ Hybridization to Show EBV Infection
For in situ hybridization, 5-μm-thick sections were mounted and dewaxed. Six hundred microliters of proteinase K (Roche Diagnostic, Milano, Italy), diluted in 50 nmol/L Tris-HCl, pH 7.6, were added for 30 minutes at 37°C. The slides were washed in distilled water, dehydrated in ethanol, series air-dried, and then incubated with 30 μl of EBV-oligonucleotides/fluorescein isothiocyanate complementary to the two nuclear EBER RNAs encoded by EBV (Dako) for 2 hours at 37°C. They were washed and incubated with 150 μl of rabbit F(ab′) anti-fluorescein isothiocyanate/AP for 30 minutes. After being washed and buffered, they were incubated with 10 ml of substrate solution containing 32 μl of 5-bromo-4-chloro-3-indolyl-phosphate (50 mg/ml) and 64 μl of nitroblue tetrazolium (50 mg/ml) for 2 hours at 37°C, and finally counterstained with fuchsine. EBV-positive sections from cases of Hodgkin’s disease were used as the positive controls, whereas EBV-negative lymphoid tissue was used as the negative control. Slides that were negative for EBV RNA control cases were tested for the viability of total RNA using a poly d(T) probe.
Registration of Mitotic and Apoptotic Index
We used an oil-immersion objective (100×) and first established the cellularity in 20 randomly chosen high-power fields (56,000 μm 2 per field) and then assessed the mitotic index (MI) and apoptotic index (AI) by registering the percentages of neoplastic cells in mitosis and the number of apoptotic cells/bodies per 100 neoplastic cells in an entire section per case (≈100 high-power fields). 31 With this approach, the relative number of apoptotic cells/bodies corresponded well with the percentage of cells showing in situ end labeling of the DNA strand breaks. 32 The intra- and interobserver reproducibility of the counts was ≈95%.
Statistical Analysis
To investigate the existence of different expression patterns on the basis of the variables p107 and pRb2/p130 and to determine their cut-off values, probability density functions were estimated using the kernel method. 35 This is a nonparametric method that does not impose any presupposed functional forms on the probabilities, such as the hypothesis of normality for example, and estimates probability density functions directly from the sample data. One great advantage of such a method is that, when a multimodal probability density function is found, the cut-off values between different populations are unequivocally identified to correspond at the distribution minima.
A one-way univariate analysis of variance was used to compare the identified groups.
Results
Clinical-Epidemiological Features
The clinical and epidemiological characteristics of all of the BL cases studied are summarized in Table 1 ▶ . The cases are grouped according to age, the localization of the disease, and presence of the HIV infection, and they correspond to the three forms of BL: eBL, sBL, and AIDS-related BL.
In the African cases, the ages ranged from 2 to 56 years (median 11 years). The maxilla and mandibula were the most common sites of tumors in patients under 16 years of age. However, 11 cases showed tumors in the cervical nodes or in the terminal ileum; 8 of them were over the age of 18. These cases clearly do not conform to the classical syndrome described by Dennis Burkitt and probably represent sBL occurring in endemic regions. 13 In addition, five cases of BL occurring in adults were associated with HIV and are therefore better classified as AIDS-related BL. 13 HIV sequences were not detected in BL in any of the patients under 18 years of age. EBV was detected in 19 cases and was associated with patients who were under 16 years of age or HIV-positive.
The five BL cases from Italy showed a median age of 26 years (range, 16–36). Two had involvement of the cervical lymph nodes, one had a tumor of the ovary, and two had a tumor of the terminal ileum; EBV was present in one case and HIV in one case.
Expression of pRb2/p130 and p107 and Mutational Screening
The results of the immunohistochemical and molecular analysis by PCR and SSCP are summarized in Table 1 ▶ .
The cytoplasmic localization of pRb2/p130 was found in 12 cases of BL using the monoclonal antibody that recognizes the amino-terminal region of the protein. In all of these samples except for one (case number 18, marked with an asterisk in Table 1 ▶ ), no staining of pRb2/p130 was revealed using the polyclonal antibody, which recognizes the carboxy-terminal region of the protein. The remaining cases showed nuclear localization of pRb2/p130, ranging from 5% to 90% of the registered positive cells, using either monoclonal or polyclonal antibodies. The molecular analysis by SSCP revealed mutations of the RB2/p130 gene in all of the samples with the cytoplasmic staining of the protein. Figure 1 ▶ illustrates the DNA-sequencing analysis of these mutants. Eleven cases showed either a guanosine or a cytosine insertion at exon 21, which caused the loss of the bipartite nuclear localization signal (NLS) resulting from a shift in the coding frame with a consequent stop codon upstream of the NLS. This mutant protein loses the amino acid at the carboxy-terminal region necessary for polyclonal antibody recognition. One additional case presented two point mutations at exon 22, inside the NLS, which do not cause amino acid loss at the end of the protein but, nevertheless, disrupt the nuclear signal. In this case, cytoplasmic staining was observed also with the polyclonal antibody. Other heterozygous point mutations were detectable at exons 19 and 20 but did not involve the nuclear localization signal.
Figure 1.
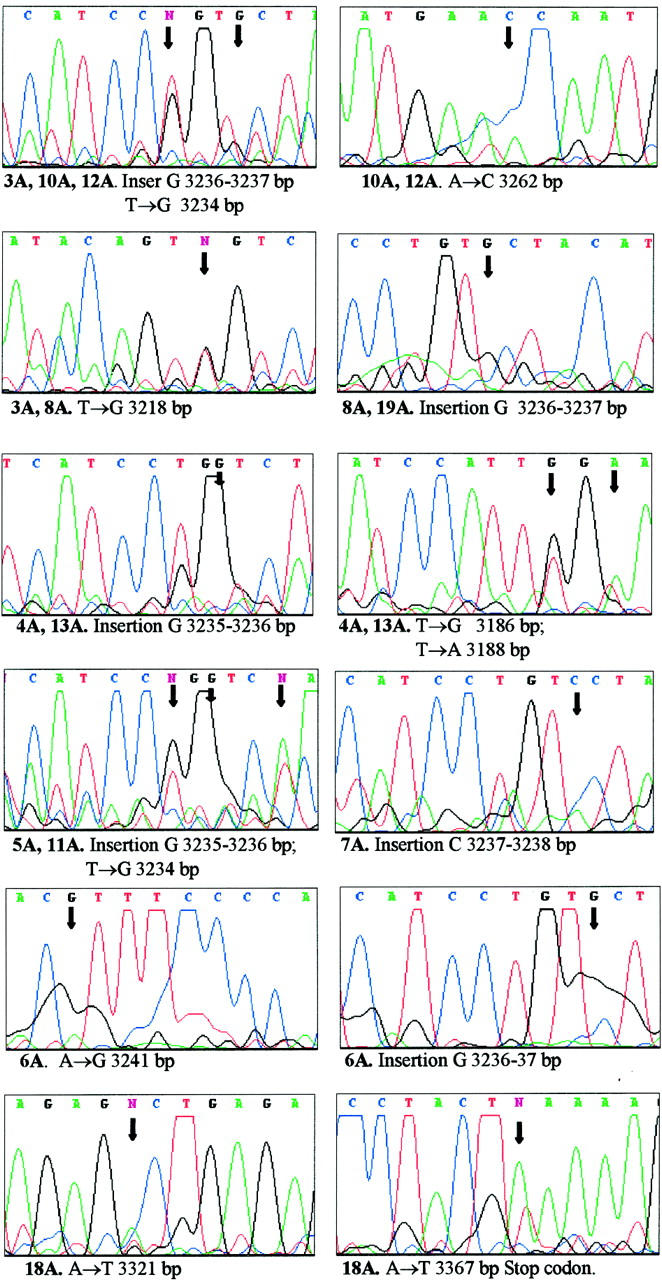
Insertions and point mutation found in RB2/p130 gene sequences of BLs, which affect a bipartite NLS. The nucleotide localization is derived from the numbering scheme used by Mayol et al. 55
Because p107 maps to a chromosome region that is not commonly found to be cytogenetically altered in the human neoplasia, 36 we used only immunohistochemistry to determine the expression of p107; this was characterized by a nuclear stain in all of the cases. Values ranging from 5% to 90% of p107-positive cells were recorded.
The Correlation of pRb2/p130 and p107 Expression with Clinical Data and HIV and EBV Infection
On the basis of the estimated probability density function of pRb2/p130 and p107 expression (Figure 2) ▶ , the cut-off values were determined, and the three groups of cases could be distinguished: 1) Cases with cytoplasmic staining of pRb2/p130 and the corresponding low values of p107 <45%; these cases showed mutations of the RB2/p130_ gene and were defined pRb2-/p107-/RB2 mutated (Figure 3, a and b) ▶ ; 2) Cases with nuclear values of pRb2/p130 <45% but values of p107>45%; these cases did not show mutations of the RB2 gene and were defined pRb2-/p107+/RB2_ wild type (Figure 3, c and d) ▶ ; 3) Cases with nuclear values of both pRb2/p130 and p107 > 45%; these cases did not show mutations of the RB2 gene and were defined pRb2+/p107+/RB2 wild-type (Figure 3, e and f) ▶ .
Figure 2.

Comparison of percentages of nuclear staining of pRB2/p130 and p107 for each case. Top and right side of the plot report the probability density functions (p.d.f.) of the variables for which the cut-off values were determined.
Figure 3.
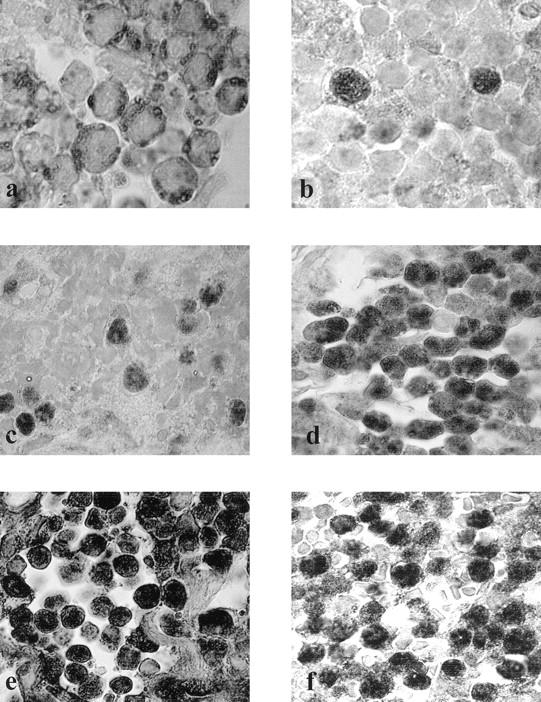
Immunostaining of pRb2/p130 and p107 in BL. a and b: Cases with cytoplasmic staining of pRb2/p130 (a) and corresponding values of p107 <45% (b**), pRb2_-/p107_- /RB2_mutated; c and d: cases with nuclear values of pRb2/p130 <45% (**c**) but values of p107>45% (d), pRb2_-/p107_+_ /RB2 wild type; e and f: cases with nuclear values of both pRb2/p130 (e) and p107 (f) > 45%, pRb2_+_ /p107+/RB2 wild type. ENVISION+/HRP method (see text); original magnifications, ×970 (a and b); ×850 (b–f**).
Table 2 ▶ summarizes the distribution of the BL cases among the groups with distinct pRb2/p130 and p107 expression. The first group (pRb2-/p107-/RB2 mutated) consisted mainly of classical eBLs with tumors involving the facial bones, in African children. The second group (pRb2-/p107+/RB2 wild type) predominantly included cases with the clinical features of sBL, characterized by terminal ileum and lymph node involvement, occurring in children or adults from both Africa and Europe. However, this association had some exceptions. The first group included one case with localization in the ileum (32-year-old man) and one case with lymph node involvement (20-year-old woman), whereas two cases of classical eBL were found in the second group. It is interesting that the third group of cases (pRb2+/p107_+_ /RB2 wild type) was made up exclusively of AIDS-related BLs.
Table 2.
Distribution of BL Cases among Groups with Distinct pRb2/p130 and p107 Expression
| BL type | pRb2−/p107−/RB2 mutated | pRb2−/p107+/RB2 wild type | pRb2+/p107+/RB2 wild type |
|---|---|---|---|
| eBL | 10 /13 | 2 /13 | 0 /13 |
| sBL* | 2 /10 | 8 /10 | 0 /10 |
| AIDS-related BL | 0 /6 | 0 /6 | 6 /6 |
No correlation was found between EBV infection and pRb2/p130 and p107 expression.
Proliferative Features of BL with Different pRb2/p130 and p107 Expression
A comparison of proliferative features (mean and SD) among the BL groups with different pRb2/p130 and p107 expression is presented in Table 3 ▶ .
Table 3.
Proliferative Features (Mean and SD) in the Groups of BLs with Different pRb2/p130 and p107 Expression
| Feature | pRb2−/p107−/RB2 mutated | pRb2−/p107+/RB2 wild type | pRb2+/p107+/RB2 wild type |
|---|---|---|---|
| MI % | 0.9 ± 0.35 | 0.8 ± 0.49 | 0.4 ± 0.09** |
| AI % | 0.65 ± 0.13 | 1.0 ± 0.59 | 0.6 ± 0.13* |
| MI/AI | 1.6 ± 0.69 | 0.78 ± 0.06 | 0.65 ± 0.09** |
| Cyclin A % | 77 ± 20.9 | 88 ± 10.6 | 82.5 ± 9.35 (NS) |
| Cyclin B % | 84 ± 7.76 | 80 ± 10 | 85 ± 5.85 (NS) |
The pRb2+/p107_+_ /RB2 wild-type group of cases, corresponding to the AIDS-related BLs, had a lower value of MI than the other two groups (P < 0.02), whereas AI was higher (P < 0.05) in the pRb2-/p107_+_ /RB2 wild-type cases. It is interesting that the MI/AI ratio was higher (P < 0.01) in pRb2-/p107_-_ _/RB2-_mutated cases than in the other two groups.
The expression of cyclins A and B was determined by immunohistochemistry. Cyclin A showed a nuclear stain whereas cyclin B exhibited a cytoplasmic and/or nuclear localization. All cases were characterized by high values of both cell cycle-associated proteins without any statistically significant difference among the groups with distinct pRb2/p130 and p107 expression.
Discussion
In this study, we used immunohistochemistry and molecular analysis to assess the expression of pRb2/p130 and p107 in a series of BLs collected from endemic and nonendemic regions. Most of the patients from Africa corresponded to the clinical syndrome described by Burkitt 1 ; however, a few cases stood out because they had tumors of the terminal ileum or lymph nodes, and most of them occurred in young adults; these cases may well represent forms of sBL occurring in endemic regions. 6,13 The cases collected from Italy showed clinical features typical of sBL. 10 Additionally, six patients were associated with HIV infection and are therefore better classified as AIDS-related BL. 13 In summary, our series well reflects the clinical and epidemiological heterogeneity of BL.
Three patterns of pRb2/p130 and p107 expression were detected and a correlation, although not very close, was found between the distinct pRb2/p130 and p107 expression and clinical-epidemiological features of the BL cases. Most of the classical eBLs were characterized by cytoplasmic staining of pRb2/p130 and the corresponding low values of p107 (pRb2-/p107_-_ /RB2 mutated). The cases with clinical features of sBL were mainly characterized by low nuclear values of pRb2/p130 but, conversely, high values of p107(pRb2-/p107_+_ /RB2 wild type). However, some discrepancies were noted, as two patients with clinical features of sBL were included among pRb2-/p107_-/RB2_-mutated cases and two patients with the clinical features of eBL among pRb2-/p107+/RB2 wild-type cases. Interestingly, the AIDS-related BLs showed values of both pRb2/p130 and p107 up to 80% to 90% (pRb2+/p107+ /RB2 wild type).
Based on the experimental data, a model for a pRb2/p130 and p107 control during cell proliferation has previously been proposed. 24 In quiescent G0 cells, the nuclear E2F-pRb2/p130 complex is responsible for the active repression of a number of cellular promoters 37_._ After its release into the cell cycle, pRb2/p130 is phosphorylated by G1 cyclin-dependent kinases (CDKs) and subsequently degraded through a proteosome-dependent mechanism, 38 resulting in the derepression of a variety of genes, including p107. 24 The accumulated p107 protein is then able to interact with E2F4 and E2F5, which have been released from pRb2/p130 and associate with cyclinA/CDK2. 29,39 Thus, in an ideally controlled proliferating cell population with identical cell cycle and cycle phase times, during which a given protein can be detected in the nucleus by the corresponding antibody, the percentages of cells expressing pRb2/p130 and p107 should be inversely correlated. This mechanism is probably maintained in the group of cases characterized by low wild-type pRb2/p130 and high p107 nuclear values, as observed in other types of malignant lymphomas. 30 In the tumors where the RB2/p130 gene is mutated and the protein is restricted to the cytoplasm, the interaction between individual proteins and E2F family members and the timing of the formation of the particular protein/E2F complexes during the cell cycle can be deregulated. 40 In particular, mutations of RB2 can also result in a loss of the ability to activate p107. 41 Yet, a significant portion of E2F4 can exist as free E2F 42 and/or can be bound to an as yet unidentified protein, 43 thus driving cells to proliferation. In AIDS-related BL, the absence of mutations in the RB2/p130 gene and the unusually high levels of nuclear expression of both pRb2/p130 and p107 suggest that other mechanisms similar to the one observed in the virus-linked oncogenesis could be involved. 44,47 All members of the retinoblastoma gene family share the ability to physically interact with certain oncoproteins of the DNA viruses. 46 These viral oncoproteins compete with the E2F family of transcription factors for binding to the Rb family of proteins. 48 The release of the E2F family members from the pocket structure leads to the induction of genes needed to proceed through the cell cycle, including p107. 25
Our results suggest that BL is composed of a mixture of molecular types with distinct genetic and phenotypic patterns, probably resulting from different pathogenetic mechanisms. It seems likely that different genetic lesions may be able to cooperate with c-myc deregulation in the genesis of BL. 6,19 The Rb-related proteins may bind to the transactivation domain of the c-myc proto-oncogene, 49 and the cells in which there is an impaired production of Rb-related proteins may be more susceptible to transformation by a deregulated c-myc gene. 50,51 Our finding of an altered expression of pRb2/p130 in BL, due either to mutations or to still unidentified virus-linked mechanisms, supports this hypothesis. In particular, mutations of the RB2/p130 gene involving an insertion at exon 21, which has occurred upstream of the putative NLS, or point mutations inside the NLS may predispose the cells to a malignant transformation. 47 Mutations of the p107 gene are unlikely, given that p107 maps to a chromosome region not commonly found to be cytogenetically altered in human neoplasia. 36 Furthermore, alterations of the p107 gene have recently been documented in only a restricted number of human hematological malignancies 52 (ie, diffuse large B-cell lymphomas).
It remains to be proven whether such molecular alterations result in different growth patterns among BL cases, because Rb-related proteins are also involved in the regulation of an elusive switch point between the cell cycle, differentiation, and apoptosis. 53 Of some interest in this context was the disparity between the MI/AI ratio in pRb2_-/p107_- _/RB2_-mutated cases, which mainly corresponded to the endemic form of BL, and the other two groups. This finding must be viewed against the background of cell birth and cell deletion kinetics, and it is in line with a previous observation that eBLs grow markedly faster than sBLs. 11 Burkitt mentioned that the tumor volume doubling time of eBL may be less than 1 day, the shortest known in malignant lymphomas. 3
Increased cell proliferation due to the alterations of the control mechanism by Rb-related proteins may represent a first step in lymphomagenesis in BL, because it has been demonstrated that increased cell proliferation is sufficient to account entirely for the tumor prevalence. 54 Additional genetic changes may occur subsequently, and missense mutations of c-myc may be selected during lymphomagenesis. 50
Footnotes
Address reprint requests to Prof. Lorenzo Leoncini, Institute of Pathologic Anatomy and Histology, University of Siena, Strada delle Scotte 6, 53100 Siena, Italy. E-mail: leoncinil@unisi.it.
Supported by Ministero Università e Ricerca Scientifica (Murst), IOR Ricerca Corrente, Funds for Selected Research Topics from the University of Bologna, Sbarro Institute for Cancer Research and Molecular Medicine, NIH grants RO1 CA 60999–01A1 and PO1 NS 36466, and The Council for Tobacco Research (to A. G.). P. P. C. is the recipient of a fellowship from the Associazione Leonardo Di Capua.
C. Cinti and L. Leoncini contributed equally to this work.
References
- 1.Burkitt DP: Sarcoma involving jaws in African children. Br J Surg 1958, 46:218-223 [DOI] [PubMed] [Google Scholar]
- 2.O’Conor GT: Malignant lymphoma in African children. II. A pathological entity. Cancer 1961, 14:270-283 [DOI] [PubMed] [Google Scholar]
- 3.Burkitt DP: The discovery of Burkitt’ s lymphoma. Cancer 1983, 51:1777-1786 [DOI] [PubMed] [Google Scholar]
- 4.O’Conor GT, Rappaport H, Smith EB: Childhood lymphomas resembling Burkitt’s tumor in the United States. Cancer 1965, 18:411-417 [DOI] [PubMed] [Google Scholar]
- 5.Wright DH: Gross distribution and morphology in Burkitt’s lymphoma. Edited by DP Burkitt, DH Wright. Edinburg and London, E and S Livingstone, 1970, pp 64–81
- 6.Magrath I: The pathogenesis of Burkitt’s lymphoma: advances of cancer research. Edited by GF Van de Woude, G Klein. London, Academic Press, 1990, pp 133–270 [DOI] [PubMed]
- 7.Wright D, Maclever P, Carter R: Childhood non Hodgkin’s in the United Kingdom: findings from the UK children’s cancer study group. J Clin Pathol 1997, 50:128-134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tamaru J, Hummel M, Marafiotti T, Kalvelage B, Leoncini L, Minacci C, Tosi P, Wright D, Stein H: Burkitt’s lymphomas express VH genes with a moderate number of antigen-selected somatic mutations. Am J Pathol 1995, 147:1398-1407 [PMC free article] [PubMed] [Google Scholar]
- 9.Pellicci PG, Knowles DM, Magrath I, dalla Favera R: Chromosomal breakpoints and structural alteration of the c-myc locus differ in endemic and sporadic forms of Burkitt’s lymphoma. Proc Natl Acad Sci USA Med Sci 1986, 83:2984-2988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Harris NL, Jaffe ES, Stein H, Banks PM, Chan JKC, Clearly ML, Delsol G, De Wolf-Peeters C, Falini B, Gatter KC, Grogan TM, Isaacson PG, Knowles DM, Mason DY, Muller-Hermenelik HK, Pileri SA, Piris MA, Ralfkiaer E, Warnke RA: A revised European-American classification of lymphoid neoplasms: a proposal from the International Lymphoma Study Group. Blood 1994, 84:1361-1392 [PubMed] [Google Scholar]
- 11.Spina D, Leoncini L, Megha T, Gallorini M, Disanto A, Tosi P, Abinya O, Nyong’o A, Pilei S, Kraft R, Laissue JA, Cottier H: Cellular kinetic and phenotypic heterogeneity in and among Burkitt’s and Burkitt-like lymphomas. J Pathol 1997, 182:145-150 [DOI] [PubMed] [Google Scholar]
- 12.Carbone A, Gloghini A, Gaidano G, Cilia AM, Basi P, Polito P, Vaccher E, Saglio G, Tirelli U: AIDS-related Burkitt’s lymphoma: morphologic and immunophenotypic study of biopsy specimens. Am J Clin Pathol 1995, 103:561-567 [DOI] [PubMed] [Google Scholar]
- 13.Wright D: What is Burkitt’s lymphoma? J Pathol 1997, 182:125-127 [DOI] [PubMed] [Google Scholar]
- 14.Subar M, Neri A, Inghirami G, Knowles DM, Dalla-Favera R: Frequent c-myc oncogene activation and infrequent presence of Epstein-Barr virus genome in AIDS-associated lymphoma. Blood 1988, 72:667-671 [PubMed] [Google Scholar]
- 15.Lazzi S, Ferrari F, Nyong’o A, Palummo N, De Milito A, Zazzi M, Leoncini L, Luzi P, Tosi P: HIV-associated malignant lymphomas in Kenya (equatorial Africa). Hum Pathol 1998, 29:1285-1289 [DOI] [PubMed] [Google Scholar]
- 16.Neri A, Barriga F, Knowles DM, Magrath IT, dalla Favera R: Different regions of the immunoglobulin heavy-chain locus are involved in chromosomal translocations in distinct pathogenetic forms of Burkitt lymphomas. Proc Natl Acad Sci USA 1988, 85:2784-2792 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shiramizu B, Barriga F, Neequaye J, Jafri A, Dalla-Favera R, Neri A, Gutierrez M, Levine P, Magrath I: Patterns of chromosomal breakpoint location on Burkitt’s lymphoma: relevance to geography and Epstein-Barr virus association. Blood 1991, 77:1516-1526 [PubMed] [Google Scholar]
- 18.Peukert K, Staller P, Schneider A, Casrmichael G, Hanel F, Eilers M: An alternative pathway for gene regulation by myc. EMBO J 1997, 16:5672-5686 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hangaishi A, Ogawa S, Imamura N, Miywaki S, Miura Y, Uike N, Shimazaki C, Emi N, Takeyama K, Hirosawa S, Kamada N, Kobayashi Y, Takemoto Y, Kitani T, Toyama K, Ohtake S, Yazaki Y, Ueda R, Hirai H: Inactivation of multiple tumor-suppressor genes involved in negative regulation of the cell cycle, MTS1/p16INK4A/CDKN2,MTS2/p15INK4B, p53, and Rb genes in primary lymphoid malignancies. Blood 1996, 87:4949-4958 [PubMed] [Google Scholar]
- 20.Preudhomme C, Dervite I, Wattel E: Clinical significance of p53 mutation in newly diagnosed Burkitt’s lymphoma and acute lymphoblastic leukemia: a report of 48 cases. J Clin Oncol 1995, 13:812-820 [DOI] [PubMed] [Google Scholar]
- 21.Bhaitia K, Fan S, Spangler G, Weintraub M, O’Connor PM, Judde JG, Magrath I: A mutant 21 cyclin-dependent kinase inhibitor from a Burkitt’s lymphoma. Cancer Res 1995, 55:1431-1435 [PubMed] [Google Scholar]
- 22.Cinti C, Claudio PP, Howard CM, Neri ML, Fu Y, Leoncini L, Tosi GM, Maraldi NM, Giordano A: Genetic alterations disrupting the nuclear localization of the retinoblastoma related gene RB2/p130 in human cell lines and primary tumors. Cancer Res 2000, 60:383-389 [PubMed] [Google Scholar]
- 23.De Luca A, Esposito V, Baldi A, Giordano A: The retinoblastoma gene family and its role in proliferation differentiation and development. Histol Histopathol 1996, 11:1029-1034 [PubMed] [Google Scholar]
- 24.Smith EJ, Leone G, Nevins JR: Distinct mechanisms control the accumulation of the Rb-related p107 and p130 proteins during cell growth. Cell Growth Diff 1998, 9:297-303 [PubMed] [Google Scholar]
- 25.Howard C, Giordano A: Neoplastic transformation: oncogenes, tumor suppressors, cyclins, and CDKs. Hormones and Cancer. Edited by W Vedeckis, Birkäuser, 1996, pp 3–41
- 26.Giordano A, Lee JH, Scheppler JA: Cell cycle regulation of histone H1 Kinase activity associated with the adenoviral protein E1A. Science 1991, 253:1271-1275 [DOI] [PubMed] [Google Scholar]
- 27.Sabattini E, Bisgaard K, Ascani S, Poggi S, Picioli M, Ceccarelli C, Pieri F, Fraternali-Orcioni G, Pileri SA: The En Vision+ system: a new immunohistochemical method for diagnostics and research. Critical comparison with the APAAP, ChemMate, CSA, LABC, and SABC techniques. J Clin Pathol 1998, 51:506-511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pileri S, Roncador G, Ceccarelli C, Piccioli M, Briskomatis A, Sabattini E, Ascani S, Santini D, Piccaluga PP, Leone O, Damiani S, Ercolessi C, Sandri F, Pieri F, Leoncini L, Falini B: Antigen retrieval techniques in immunohistochemistry: comparison of different methods. J Pathol 1997, 183:116-123 [DOI] [PubMed] [Google Scholar]
- 29.Claudio PP, DeLuca A, Howard CM, Baldi A, Firpo EJ, Koff A, Paggi MG, Giordano A: Functional analysis of pRb2/p130 interaction with cyclins. Cancer Res 1996, 56:2003-2008 [PubMed] [Google Scholar]
- 30.Leoncini L, Bellan C, Cossu A, Claudio PP, Lazzi S, Cinti C, Cevenini G, Megha T, Laurini L, Luzi P, Fraternali Orcioni G, Piccioli M, Pileri S, Tosi P, Giordano A: Retinoblastoma related p107 and pRb2/p130 proteins in malignant lymphomas: distinct mechanisms of cell growth control. Clin Cancer Res 1999, 5:4065–4072 [PubMed]
- 31.Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn GT, Mullis KB, Erlich HA: Primer directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 1988, 239:487-491 [DOI] [PubMed] [Google Scholar]
- 32.Miura T, Sakuragi J, Kawamura M: Establishment of a philogenetic survey system for AIDS-related lentiviruses and demonstration of a new HIV-2 subgroup. AIDS 1990, 4:1257-1261 [DOI] [PubMed] [Google Scholar]
- 33.Leoncini L, Del Vecchio MT, Megha T, Barbini P, Galieni P, Pileri S, Sabattini E, Gherlinzoni F, Tosi P, Kraft R, Cottier H: Correlations between apoptotic and proliferative indices in malignant non-Hodgkin’s lymphomas. Am J Pathol 1993, 142:755-763 [PMC free article] [PubMed] [Google Scholar]
- 34.Spina D, Leoncini L, Del Vecchio MT, Megha T, Minacci C, Poggi SA, Pileri S, Tosi P, Kraft R, Laissue JA, Cottier H: Low versus high cell turnover in diffusely growing non-Hodgkin’s lymphomas. J Pathol 1995, 177:335-341 [DOI] [PubMed] [Google Scholar]
- 35.Krzanowski WJ, Marriot FHC: Multivariate analysis (part 2). 1995:pp 26-27 Wiley & Sons New York, Toronto:
- 36.Ewen ME, Xing Y, Lawrence JB, Livingston DM: Molecular cloning, chromosomal mapping, and expression of the cDNA for p107, a retinoblastoma gene product-related protein. Cell 1991, 66:1155-1164 [DOI] [PubMed] [Google Scholar]
- 37.Stiegler P, Kasten M, Giordano A: The RB family of cell cycle regulatory factors. J Cell Biochem Suppl 1998, 30/31:30-36 [PubMed] [Google Scholar]
- 38.Hateboer G, Kerhoven RM, Shvarts A, Bernards R, Beijersbergen R: Degradation of E2F by the ubiquitin-proteasome pathway: regulation by retinoblastoma family proteins and adenovirus transforming protein. Genes Dev 1996, 10:2960-2970 [DOI] [PubMed] [Google Scholar]
- 39.Shirodkar S, Ewen M, De Caprio JA, Morgan JA, Livingston DM, Chittenden T: The transcription factor E2F interacts with the retinoblastoma product and a p107-cyclin A complex in a cell cycle-regulated manner. Cell 1992, 68:157-166 [DOI] [PubMed] [Google Scholar]
- 40.Johnson DG, Schneider-Broussard R: Role of E2F in cell cycle control and cancer. Front Biosci 1998, 3:d447-458 [DOI] [PubMed] [Google Scholar]
- 41.Kim YW, Otterson GA, Kratzke RA, Coxon AB, Kaye FJ: Differential specificity for binding of the retinoblastoma binding protein 2 to RB, p107, and TATA-binding protein. Mol Cell Biol 1994, 14:7256-7264 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Moberg K, Starz MA, Lees JA: E2F-4 switches from p130 to p107, and pRB in response to cell cycle reentry. Mol Cell Biol 1996, 16:1436-1449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mulligan GJ, Wong J, Jacks T: p130 is dispensable in perpheral T lymphocytes: evidence for functional compensation by p107 and pRb. Mol Cell Biol 1998, 18:206-220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Desjardins P, Pilon AA, Hassell JA, Mes-Masson AM: Polyomavirus large T-antigen binds the “pRb related” protein p130 through sequences in conserved region 2. Virus Res 1997, 47:85-90 [DOI] [PubMed] [Google Scholar]
- 45.Lee BH, Liu M, Mathews MB: Regulation of the human proliferating cell nuclear antigen promoter by the adenovirus E1A-associated protein p107. J Virol 1998, 72:1138-1145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.De Luca A, Baldi A, Esposito V, Howard CM, Bagella L, Rizzo P, Caputi M, Pass HI, Giordano GG, Baldi F, Carbone M, Giordano A: The retinoblastoma gene family pRb/p105, p107, pRb2/p130 and simian virus-40 large T-antigen in human mesotheliomas. Nat Med 1997, 3:913-916 [DOI] [PubMed] [Google Scholar]
- 47.Howard CM, Claudio PP, Gallia GL, Gordon J, Giordano GG, Hauck WW, Khalili K, Giordano A: Retinoblastoma-related protein pRb2/p130 and suppression of tumor growth in vivo. J Natl Cancer Inst 1998, 90:1451-1460 [DOI] [PubMed] [Google Scholar]
- 48.Fattaey AR, Harlow E, Helin K: Independent regions of adenovirus E1A are required for binding to and dissociation of E2F-protein complexes. Mol Cell Biol 1993, 13:7267-7277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gu W, Bathia K, Magrath IT, Dang CV, Dalla-Favera R: Binding and suppression of the myc transcriptional activation domain by p107. Science 1994, 264:251-254 [DOI] [PubMed] [Google Scholar]
- 50.Hoang AT, Lutterbach B, Lewis BC, Yano T, Chou TY, Barret JF, Raffeld M, Hann SR, Dang CV: A link between increased trasforming activity of lymphoma-derived MYC mutant alleles, their defective regulation by p107, and altered phosphorylationb of the c-myc transactivation domain. Mol Cell Biol 1995, 15:4031-4042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Alevizopoulos K, Vlach J, Hennecke S, Amanti B: Cyclin E and c-myc promote cell proliferation on the presence of p16INK4a and hypophosphorylated retinoblastoma family proteins. EMBO J 1997, 16:5322-5333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Takimoto H, Tsukuda K, Ichimura K, Hanafusa H, Nakamura A, Oda M, Harada M, Shimizu K: Genetic alteration in the retinoblastoma protein-related p107 gene in human hematologic malignancies. Biochem Biophys Res Commun 1998, 251:264-268 [DOI] [PubMed] [Google Scholar]
- 53.Harwig S, Strauss M: The retinoblastoma protein: a regulator of cell cycle differentiation and apoptosis. Eur J Biochem 1997, 246:581-601 [DOI] [PubMed] [Google Scholar]
- 54.Ellwein LB, Purtillo DT: Cellular proliferation and genetic events involved in the genesis of Burkitt Lymphoma (BL) in immune compromised patients. Cancer Genet Cytogenet 1992, 64:42-48 [DOI] [PubMed] [Google Scholar]
- 55.Mayol X, Grana X, Baldi A, Sang N, Hu Q, Giordano A: Cloning of a new member of the retinoblastoma gene family (pRb2) which binds to the E1A transforming domain. Oncogene 1993, 8:2561-2566 [PubMed] [Google Scholar]