Establishment of the Dorso-ventral Axis in Xenopus Embryos Is Presaged by Early Asymmetries in β-Catenin That Are Modulated by the Wnt Signaling Pathway (original) (raw)
Abstract
Eggs of Xenopus laevis undergo a postfertilization cortical rotation that specifies the position of the dorso-ventral axis and activates a transplantable dorsal-determining activity in dorsal blastomeres by the 32-cell stage. There have heretofore been no reported dorso-ventral asymmetries in endogenous signaling proteins that may be involved in this dorsal-determining activity during early cleavage stages. We focused on β-catenin as a candidate for an asymmetrically localized dorsal-determining factor since it is both necessary and sufficient for dorsal axis formation. We report that β-catenin displays greater cytoplasmic accumulation on the future dorsal side of the Xenopus embryo by the two-cell stage. This asymmetry persists and increases through early cleavage stages, with β-catenin accumulating in dorsal but not ventral nuclei by the 16- to 32cell stages. We then investigated which potential signaling factors and pathways are capable of modulating the steady-state levels of endogenous β-catenin. Steadystate levels and nuclear accumulation of β-catenin increased in response to ectopic Xenopus Wnt-8 (Xwnt_-8) and to the inhibition of glycogen synthase kinase-3, whereas neither Xwnt_-5A, BVg1, nor noggin increased β-catenin levels before the mid-blastula stage. As greater levels and nuclear accumulation of β-catenin on the future dorsal side of the embryo correlate with the induction of specific dorsal genes, our data suggest that early asymmetries in β-catenin presage and may specify dorso-ventral differences in gene expression and cell fate. Our data further support the hypothesis that these dorso-ventral differences in β-catenin arise in response to the postfertilization activation of a signaling pathway that involves Xenopus glycogen synthase kinase-3**.**
Our understanding of how the embryonic axes are specified during the early development of vertebrates is progressing towards an understanding at the cellular and biochemical level. To a large degree, the discussion now centers on establishing the interplay between specific biochemical pathways and cellular events. The rate of these theoretical advances was stimulated by the discovery that ectopic expression of a member of the Wnt family in the ventral marginal zone of Xenopus embryos is sufficient to elicit a complete duplication of the embryonic axes (McMahon and Moon, 1989). This observation raised the possibility that an endogenous Wnt pathway might normally be involved in axis formation, but the observation that the unrelated signaling factors noggin (Smith and Harland, 1992) and Vg1 (Dale et al., 1993; Thomsen and Melton, 1993) can also induce complete axes suggests that further research is needed to distinguish which, if any of these factors, are normally involved in axis formation.
Any attempt to evaluate secreted factors that may be involved in specifying the dorso-ventral axis in Xenopus embryos should take into account known observations regarding the cellular basis for axis specification. Specifically, the postfertilization cortical rotation of Xenopus is important in determining the position of the future dorsal axis (for reviews see Gerhart et al., 1989; Larabell et al., 1996). Suggesting that dorsal-determining information is present in the vegetal pole before cortical rotation, removal of this region blocks axis formation (Sakai, 1996), and injection of vegetal pole cytoplasm into host embryos has the capacity to induce an ectopic axis (Fujisue et al., 1993; Holowacz and Elinson, 1993). After cortical rotation, this dorsal-determining activity is displaced to the future dorsal side of the embryo, and transplantation of dorsal cells or cytoplasm to the ventral side of a host embryo elicits formation of a secondary dorsal axis (Gimlich, 1986; Kageura, 1990; Yuge et al., 1990; Fujisue et al., 1993). While there are presently no data showing dorso-ventral differences in the localization or activity of endogenous secreted factors that correlate with this dorsal-determining activity of the egg and early embryo, candidate molecules include Wnts (for reviews see Cui et al., 1995; Torres et al., 1996), Vg1 (Dale et al., 1993; Thomsen and Melton, 1993), and noggin (Smith and Harland, 1992).
Given the lack of evidence of a dorsal enrichment in expression or activity of any of these secreted factors, it is likely that a greater understanding of the signal transduction cascades stimulated by these factors would contribute to an understanding of which of these signaling pathways, if any, are actually used by Xenopus embryos to initiate formation of the endogenous axis. With regard to candidate cytoplasmic signaling factors, attention justifiably should focus on β-catenin, a multifunctional protein that is involved in cell adhesion at adherens junctions and in cytoplasmic and nuclear signal transduction events (for review see Miller and Moon, 1996). β-Catenin meets a number of reasonable criteria for playing a role in specification of the dorso-ventral axis in vertebrate embryos. β-Catenin is maternally expressed at the RNA and protein level (DeMarais and Moon, 1992), and when ectopically expressed, it is sufficient to mimic the endogenous dorsal-determining activity by inducing the formation of complete secondary axes in Xenopus (Funayama et al., 1995; Guger and Gumbiner, 1995) and in zebrafish (Kelly et al., 1995). Moreover, depletion of maternal β-catenin transcripts from Xenopus oocytes prevents formation of the endogenous axis (Heasman et al., 1994) and disruption of the gene in mice prevents mesoderm formation (Haegel et al., 1995). It is likely that the ability of β-catenin to alter gene expression and cell fate involves its interaction with architectural HMG box transcription factors (Behrens et al., 1996; Molenaar et al., 1996). Importantly, injection of a mutant form of one of these factors, XTcf-3, into the dorsal side of Xenopus embryos blocks formation of the endogenous dorsal axis and blocks the ability of ectopic β-catenin to induce a secondary axis (Molenaar et al., 1996). These data collectively support the hypothesis that dorsal β-catenin interacts with architectural transcription factors to regulate the expression of dorsal genes required for dorso-ventral axis specification.
While these reports indicate a role for β-catenin in dorso-ventral axis formation, the following two key issues need to be resolved: (a) Is endogenous β-catenin expressed appropriately to play a role in axis specification? If β-catenin were involved in dorso-ventral axis specification, it should be dorsally enriched and/or have greater dorsal activity by the 32-cell stage. However, previous investigations of β-catenin localization have reported either no dorso-ventral differences in embryos (Fagotto and Gumbiner, 1994) or dorso-ventral differences after the mid-blastula transition (Schneider et al., 1996)—too late to play a role in the dorsal-determining activity defined by cytoplasm and blastomere transplantation assays using cleavage stage embryos. (b) Can any or all of the secreted factors that induce an ectopic axis also modulate β-catenin levels? Since β-catenin is required for the formation of the endogenous axes (Heasman et al., 1994), and since β-catenin steady-state levels and nuclear accumulation rise in response to modulation of cytoplasmic components of the Wnt signaling pathway in the gastrula (Yost et al., 1996), it is important to directly determine whether any of the axisinducing secreted factors modulate levels of β-catenin in the cleavage stage embryo. Such assays would determine whether all of the axis-inducing factors operate through the same intracellular signaling pathway and which are candidates for normally playing a role in modulating β-catenin levels.
The present study resolves both of these issues in a manner consistent with the embryological data on dorso-ventral axis formation. Using optimized immunocytochemistry and Western blot methods, we demonstrate that endogenous β-catenin is enriched in dorsal regions of the early embryo, becoming pronounced by the 16- to 32-cell stages, when it accumulates in dorsal nuclei and cytoplasm. We then show that the Wnt signaling pathway, but not ectopic noggin or Vg1, regulates steady-state β-catenin levels in the early embryo. Our concluding model for specification of the embryonic dorso-ventral axis takes into account these data, as well as the possibility that endogenous β-catenin is regulated in a Wnt-independent manner.
Materials and Methods
cDNA Constructs and Embryo Microinjection
cDNAs encoding Xenopus Wnt-8 (Xwnt)1, Xwnt-5A, BVg1, noggin, prolactin, and β-galactosidase were obtained and transcribed in vitro as described (Torres et al., 1996), as were wild-type and point mutant Xenopus β-catenin tagged with a c-myc epitope (Yost et al., 1996), wild-type and kinase-dead Xenopus glycogen synthase kinase-3 (Xgsk-3) (Pierce and Kimelman, 1995), and Xwnt-II (Ku and Melton, 1993). Embryos were injected with these RNAs and cultured as described (Torres et al., 1996; Yost et al., 1996) with further details in the figure legends.
Confocal Microscopic Localization of β-Catenin
For Figs. 1, 2, 3, and 8 A, whole-mount confocal immunocytochemistry was carried out on untreated embryos, and embryos were irradiated at the vegetal pole with UV light to prevent cortical rotation (e.g., Gerhart et al., 1989; Yost et al., 1996). Embryos were fixed overnight in 4% paraformaldehyde, 0.1% gluteraldehyde, 100 mM KCl, 3 mM MgCl2, 10 mM Hepes, 150 mM sucrose, and 0.1% Triton X-100, pH 7.6. Fertilization envelopes were manually removed from all embryos and blocking of nonspecific binding was carried out using 0.1% Triton X-100 in Super Block (Pierce, Rockford, IL). Embryos were incubated overnight with an antibody raised in rabbit (Yost et al., 1996) against a bacterial fusion protein derived from the amino-terminal region of Xenopus β-catenin (kindly provided by P. McCrea, M.D. Anderson Cancer Center, Houston, TX). Control antibodies (see Fig. 3) were polyclonal antibodies against membrane skeleton protein 4.1 (Spencer et al., 1990), polyclonal antibodies against α-spectrin (Giebelhaus et al., 1987), or a commercial pan cytokeratin mouse monoclonal antibody (No. C2931; Sigma Chemical Co., St. Louis, MO). Embryos were rinsed for 1 h in Super Block plus 0.1% Triton X-100 with three changes and then incubated overnight in TRITC-conjugated antirabbit or anti-mouse (Sigma Chemical Co.) secondary antibody, with rotation, followed by three additional rinses in Super Block. Embryos were bisected relative to pigmentation and/or the sperm entry point under a dissecting microscope and cleared in a 1:2 mixture of benzyl alcohol/benzyl benzoate.
Figure 1.

Immunolocalization of β-catenin in early Xenopus embryos. Whole-mount staining with antibody to β-catenin demonstrates greater staining on the dorsal side of embryos (to the right in all panels) at the 8- (A and B) and 16-cell (C and D) stages. Immunolabeled embryos were cut along the equator separating the animal (A and C) from vegetal (B and D) blastomeres and examined with a confocal microscope. Optical sections through these blastomeres demonstrate an arc of cytoplasmic β-catenin in the periphery of the animal dorsal blastomeres (A and C) and vegetal dorsal blastomeres (B and D). Ventral blastomeres stain for β-catenin at the cell surface but not in the cytoplasm.
Figure 2.

Spatial and temporal distribution of β-catenin during early development of the Xenopus embryo. Embryos were bisected along the dorso-ventral axis after immunolabeling so that each half contained dorsal and ventral blastomeres. Each half was imaged from the bisected surface in the confocal microscope. Embryos are oriented with dorsal on the right, ventral on the left, animal hemisphere at the top, and vegetal hemisphere at the bottom, in A–F. Immunocytochemistry reveals an increase in cytoplasmic β-catenin (orange) on the dorsal side of the embryo from the two-cell (A), to four-cell (B), to eight-cell (C) stages. At the 16-cell stage, intense β-catenin is observed in the cytoplasm (D, arrow denotes nucleus) as well as in the nucleus (G, arrow) of the dorsal vegetal blastomeres. At the 32-cell stage, β-catenin is enriched in the cytoplasm (E, arrows denote nuclei) and nuclei (I, arrow) of all dorsal blastomeres, while ventral blastomeres have lower cytoplasmic staining (E) but neglible nuclear staining (H, arrow). Overall, 93% of the embryos analyzed (n = 120) from the 2–32-cell stages displayed this dorsal enrichment of β-catenin. At the blastula stage, β-catenin is detected in the cytoplasm of both dorsal and ventral cells (F), as well as in ventral (J) and dorsal (L) nuclei, though the dorsal signal was stronger in both cellular compartments. Primarily the outer layers of blastomeres demonstrate intense cytoplasmic β-catenin staining, even when blastula were bisected before immunolabeling to assure access of the antibodies to the inner blastomeres. Specific β-catenin staining is detectable in these inner vegetal blastomeres (K) when fluorescence sensitivity is increased to the point where the dorsal signal becomes saturated.
Figure 3.

Immunolocalization of proteins unrelated to β-catenin. To assess whether the patterns of β-catenin observed in the eight-cell embryo (Fig. 1) were unique or were shared by other proteins, we undertook a comparable analysis of eight-cell embryos stained with other antibodies, optically sectioned near the equator, and viewed at low magnification to monitor dorsoventral patterns (A–C) or viewed at the membrane at high magnification (D–F). Dorsal is to the right for A, B, and C as in Fig. 1. While eight-cell embryos show a clear dorsal enrichment of β-catenin (Fig. 1, A and B), membrane skeleton protein 4.1 (A), α-spectrin (B), and cytokeratin (C) antibodies show no dorsal enrichment. The α-spectrin image in B is somewhat more pigmented in the animal hemisphere than the other images, hence the ventral blastomeres appear somewhat darker than the dorsal blastomeres (right) owing to pigmentation differences. The cytokeratin image is yellow because of the extensive overlap of the green autofluorescence signal with the red cytoplasmic staining of the antibody (see F for cytoplasmic stain at higher magnification). While β-catenin staining extends extensively from the membrane through the cytoplasm at the eight-cell stage (Fig. 1, A and B), both protein 4.1 (D) and α-spectrin (E) are restricted to the plasma membrane, which is consistent with their known functions in the membrane skeleton, while the cytokeratin staining extends into the cytoplasm (F) as filaments as previously noted (Klymkowsky et al., 1987), though our fixation methods also retain nonpolymeric protein and increase nonfilamentous cytokeratin staining.
Figure 8.
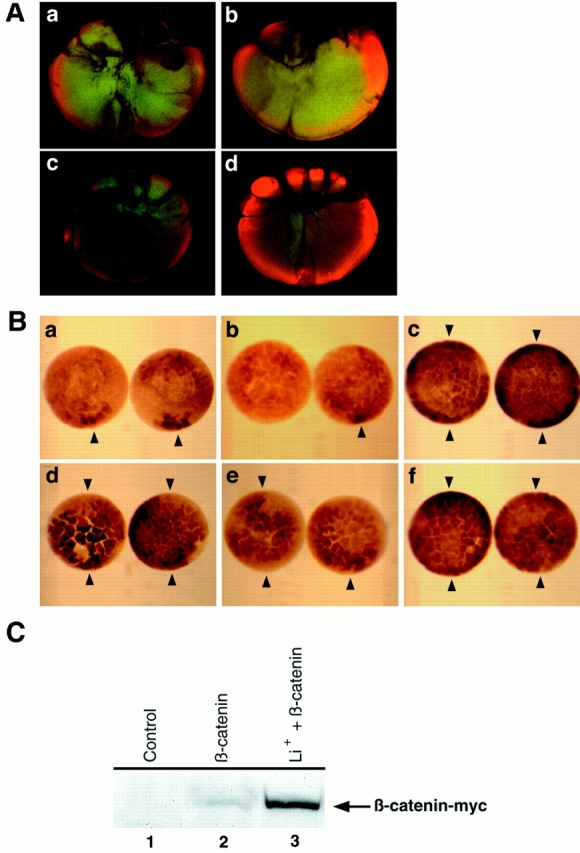
Effects of Xgsk_-3 and LiCl on the spatial accumulation of β-catenin. (A) The dorso–ventral asymmetry in β-catenin requires the postfertilization cortical rotation and is regulated by Xgsk_-3. When the postfertilization cortical rotation is prevented by UV-irradiation of the vegetal pole during the first cell cycle (a), 32-cell embryos do not display dorsal enrichments in β-catenin observed in control embryos (b), and some show ventral increases. Injection of 4 ng of RNA encoding Xgsk-3 into the dorsal marginal zone of four- to eight-cell embryos also blocks the dorsal increase in β-catenin staining at the 32-cell stage (c). Conversely, injection of 4 ng of RNA encoding a dominant negative Xgsk-3 into the ventral marginal zone promotes accumulation of β-catenin throughout the 32-cell embryo (d). (B) Injection of a mixture of prolactin and β_-catenin–myc RNAs leads to the accumulation of β-catenin–_myc on the future dorsal side of the embryo (a), though the RNAs are injected and expressed throughout the embryo. Decreased staining (left embryo) or staining for β-catenin–myc similar to controls (right embryo) is evident when _Xgsk_-3 RNA is injected with the β_-catenin– myc RNA (b). Inhibition of endogenous Xgsk-3 by injecting dnXgsk_-3 RNA increases and expands the accumulation of ectopic β-catenin– myc to the entire embryonic marginal zone (c). Treatment with lithium also expands the domain of β-catenin–_myc accumulation in embryos overexpressing prolactin (d) or Xgsk-3 (e). Expression of ptβ-catenin–_myc (which encodes a form of β-catenin that is not phosphorylated by Xgsk-3; Yost et al., 1996) with control prolactin results in the accumulation of ptβ-catenin–myc everywhere in the embryo (f). In these experiments 1 ng of _β_-catenin–myc or _ptβ_-catenin–myc RNA mixed with 3 ng of prolactin, Xgsk_-3, or dnXgsk_-3 RNA was injected into the marginal zone of each blastomere of fourcell embryos. Arrowheads denote dorsal (single arrowhead) or dorsal and ventral (two arrowheads) staining. (C) Lithium treatment increases steady-state levels of β-catenin–_myc expressed from injected RNA. Embryos were injected and treated with LiCl as in Materials and Methods. At stage 6.5–7, embryo extracts were prepared and probed for the c-myc epitope–tagged β-catenin by Western blot analysis. Lane 1, Uninjected controls do not express β-catenin–_myc; lane 2, embryos injected with _β_-catenin–myc RNA express the encoded protein; lane 3, injection of β_-catenin–myc RNA as in lane 2 followed by treatment with LiCl leads to greater accumulation of β-catenin–_myc.
Embryos were viewed using a confocal laser scanning microscope (model 1024; BioRad Labs, Hercules, CA) using rhodamine filters to collect the β-catenin signal (orange) superimposed on the blastomeres, which were imaged using fluorescein filters to excite the inherent autofluorescence (green). The threshold of fluorescence was set so that the greatest fluorescence was not saturated, with one consequence being that the images shown do not show the β-catenin, which is evident at plasma membranes, albeit at lower levels. The small, dorsal blastomeres were easily distinguished from the larger, ventral blastomeres. Confirmation of dorsoventral polarity was also achieved by monitoring blastomere pigmentation, and in one- to four-cell embryos, by following the sperm entry point. Secondary antibody alone did not stain embryos (not shown), and preblocking the primary antibody with the fusion protein used as immunogen blocks all fluorescent staining (Yost et al., 1996). These fixation and visualization methods differ substantially from those that heretofore did not reveal dorso-ventral differences in β-catenin in cleavage stage embryos (Fagotto and Gumbiner, 1994; Schneider et al., 1996).
To determine the effects of _Xwnt_-8 or _Xwnt_-5A on endogenous β-catenin localization in the nucleus (see Fig. 7), embryos were fertilized and injected in the animal poles at the two-cell stage with 5–10 ng of RNA encoding Xwnt-8 (Christian et al., 1992), Xwnt-5A (Moon et al., 1993), or bovine prolactin. Animal caps were cut from stage 6.5–7 embryos and fixed in 4% paraformaldehyde-PBS overnight at 4°C. Animal caps were dehydrated by washing once in PBT (PBS, 0.2% Triton X-100), once in 75% methanol (MeOH)/75% PBT, and twice in 100% MeOH. Animal caps were then bleached overnight in 50% MeOH/30% H202/20% DMSO at room temperature. Explants were rehydrated by washing once in 75% PBT/25% MeOH and twice in PBT, followed by incubation overnight at 4°C in anti–β-catenin antibody (Yost et al., 1996; 1:500 in PBT with 10% normal goat serum). After three washes in PBT, animal caps were incubated in CY-3–conjugated antibody (1:250 in PBT, with 10% normal goat serum; Jackson Laboratories, Bar Harbor, ME). After washing three times in PBT, animal caps were mounted under coverslips in Vectashield (Vector Laboratories, Burlingame, CA) and viewed under a confocal microscope (model MRC; BioRad Labs). Nuclear counts were performed on stacked z-series, and images shown in Fig. 7 are single optical sections.
Figure 7.

_Xwnt_-8, but not _Xwnt_-5A, increases the accumulation of endogenous β-catenin in nuclei. The animal pole of each blastomere at the two-cell stage was injected with RNA encoding prolactin (A), Xwnt-8 (B), or Xwnt-5A (C). At stage 6.5–7 animal cap explants were isolated, fixed, and processed for immunolocalization of endogenous β-catenin.
Enzyme-linked Whole-Mount Immunolocalization of β-Catenin
Where indicated, embryos were processed for whole-mount immunolocalization of c-myc–_tagged ectopic β-catenin using previously described methods for localizing c-myc (Christian and Moon, 1993). Where indicated, embryos were injected with various RNAs before localization of β-catenin–_myc.
Western Blot Analyses
To compare potential dorso-ventral differences in the steady-state levels of endogenous β-catenin on protein blots, control embryos and embryos vegetally irradiated with UV light (e.g., as in Gerhart et al., 1989; Yost et al., 1996) were fixed in 3.7% formaldehyde in 1× modified Barth's solution (MBS; 88 mM NaCl, 1 mM KCl, 2.4 mM NaHCO3, 10 mM Hepes, pH 7.5, 0.82 mM MgSO4·7H2O, 0.33 mM Ca(NO3)2·4H2O, CaCl2·6H2O) for 18 min at room temperature, followed by manual dissection as diagrammed in Fig. 4 A. Protein was extracted in lysis buffer (10 mM Tris, 150 mM NaCl, 2 mM EDTA, 2 mM EGTA, 1% NP-40, 4 mM PMSF, pH 7.5) and 1,1,2-trichlorotrifluoroethane (Sigma Chemical Co.). Protein samples equivalent to five dissected portions were analyzed by SDSPAGE, and immunoblots were probed with a rabbit anti–GST-β-catenin polyclonal antibody (Yost et al., 1996), a rabbit anti-α-spectrin polyclonal antibody (Giebelhaus et al., 1987), or a rabbit anti–chicken tubulin antibody, which was generated as in Giebelhaus et al. (1987) by immunizing rabbits with polyacrylamide gel slices containing chicken erythroid cell tubulin. Immunoblots were then probed with an HRP-conjugated goat anti– rabbit secondary antiserum (BioRad Labs). The HRP signal was visualized by enhanced chemiluminescence (Amersham, Corp., Arlington Heights, IL). Signals for β-catenin, α-spectrin, and tubulin were quantitated by use of densitometry (model UA-5 Absorbance/Fluorescence detector; Isco, Lincoln, NE). Dorsal and ventral β-catenin levels were compared after normalization for α-spectrin or tubulin, to control for variability in protein content. Dorsal β-catenin values were set at 100% and ventral β-catenin levels were expressed as a ratio: (ventral β-catenin/ dorsal β-catenin) × 100%.
Figure 4.
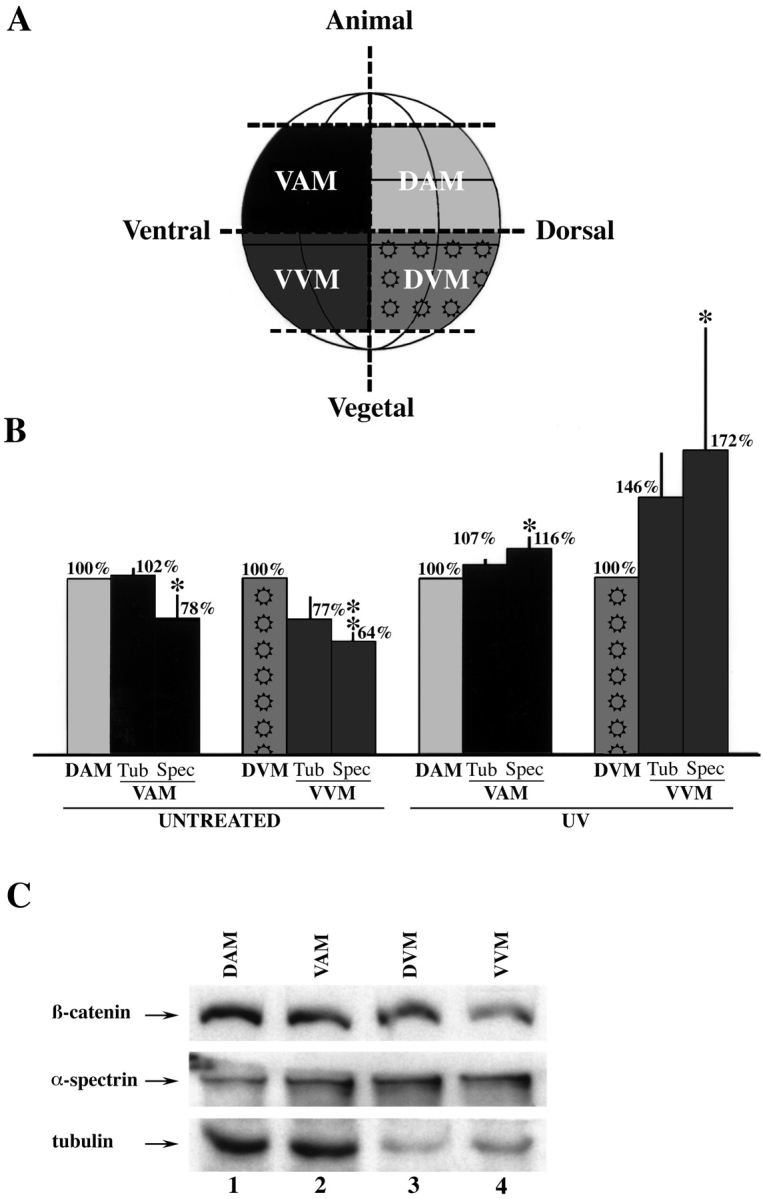
Western blot analysis of microdissected 32-cell Xenopus embryos reveals a dorso-vegetal enrichment of β-catenin. (A) Schematic diagram of how embryos were dissected into dorsal animal marginal (DAM; light gray), ventral animal marginal (VAM; black), dorsal vegetal marginal (DVM; pattern), and ventral vegetal marginal (VVM; dark gray) portions. (B) Dorsal steady-state levels of endogenous β-catenin are greater than ventral levels in untreated but not in UV-ventralized embryos. On average, the VVM portions from untreated embryos contained 77% (normalized to tubulin, three experiments) and 64% (normalized to spectrin, five experiments) as much β-catenin as the DVM zones. The VVM portions dissected from UV-irradiated embryos contained more β-catenin relative to their dorsal counterparts; 146% (normalized to tubulin, four experiments) and 172% (normalized to spectrin, four experiments). Spectrin-normalized steady-state levels of β-catenin in the VAM zone were also lower than the DAM region in untreated embryos (78%, five experiments). These differences were eliminated with UV treatment (four experiments for both tubulin and spectrin normalization). The asterisks indicate that the difference in β-catenin content between the dorsal and ventral marginal zones is statistically significant, as determined by Student's _t_-test (P < 0.05 for a single asterisk, P < 0.005 for a double asterisk). The error bars represent the standard error. (C) Representative Western blot detecting endogenous β-catenin, α-spectrin, and tubulin. Ventral levels of β-catenin are lower in the dissected VAM (lane 2) relative to the DAM (lane 1) regions, and in the VVM (lane 4) relative to DVM (lane 3) regions. It is worth noting that the dissected quadrants of the embryo do not precisely correspond to the areas of maximal β-catenin staining determined by confocal microscopy. Moreover, the thresholds of fluorescence chosen for Figs. 1 and 2 were set high, so as not to saturate the dorsal-ventral differences, and as a result these images do not show the detectable lower levels of cytoplasmic and membrane-associated β-catenin, which nevertheless would contribute to the signals on Western blots. Thus, one cannot quantitatively compare the dorso-ventral differences monitored by the dissections vs. confocal microscopy.
To monitor endogenous and ectopic β-catenin in injected and LiCltreated embryos, protein was extracted as described above 2–3 h after RNA injection (stage 5–6) and analyzed by Western blot using anti–β-catenin or anti–c-myc antibodies, respectively (above and Yost et al., 1996). Endogenous β-catenin signals were normalized to α-spectrin and tubulin bands from the same Western blot to control for protein content and gel loading (above). All Westerns were subsequently processed as described above for endogenous β-catenin quantitation or as described (Yost et al., 1996) for ectopic β-catenin–myc quantitation. β-catenin levels present in control samples were set at 100% and experimental samples were expressed as a ratio: (experimental β-catenin/control β-catenin) × 100%. Where described, eight-cell embryos were incubated for 12 min at room temperature in 0.3M LiCl in 0.1× MMR (0.1 M NaCl, 2 mM KCl, 1 mM MgSO4·7H2O, 2 mM CaCl2, 5 mM Hepes, pH 7.8) immediately after RNA microinjection.
Results
Dorso-ventral Asymmetries in β-Catenin in Cleavage Stage Embryos
We first conducted a confocal microscopical analysis of the early Xenopus embryo to investigate whether there were asymmetries in β-catenin along the prospective dorso-ventral axis. Optical equatorial sections of eight-cell embryos demonstrate distinct peripheral cytoplasmic staining on the dorsal side of the embryo, monitored in both the animal (Fig. 1 A) and vegetal hemispheres (Fig. 1 B). Cytoplasmic β-catenin is also more pronounced on the dorsal side of 16-cell embryos, in both the animal (Fig. 1 C) and vegetal (Fig. 1 D) hemispheres.
To determine the earliest time at which asymmetries in endogenous β-catenin could be detected, and to determine whether it persisted through to the 32-cell stage when dorsal vegetal blastomeres can be transplanted and exhibit axis-inducing activity (Gerhart et al., 1989, Kageura, 1990), we analyzed embryos beginning at the two-cell stage. To allow simultaneous comparison of animal and vegetal regions, we undertook the analysis using optical sectioning along the dorso-ventral, rather than equatorial, axes. Twocell embryos exhibit an asymmetry in β-catenin in the peripheral cytoplasm (94%, n = 17 embryos; Fig. 2 A), with the elevated β-catenin predominantly forming opposite the visible sperm entry point (not shown), and thus corresponding to the prospective dorsal side. This asymmetry is also evident through the 4- (Fig. 2 B), 8- (C), 16- (D), and 32-cell (E) stages. By the blastula stage, β-catenin is still elevated on the dorsal relative to ventral side with greatest staining of outer ectodermal layers (Fig. 2 F), as previously reported (Schneider et al., 1996). However, internal blastomeres also stain for β-catenin (Fig. 2 K) when fluorescence sensitivity is increased to levels that obscure the dorso-ventral differences.
Staining of eight-cell embryos with control antibodies for membrane skeleton protein 4.1 (Fig. 3 A), α-spectrin (Fig. 3 B), or cytokeratin (Fig. 3 C) did not reveal any dorsal enrichment for these proteins, demonstrating that the dorsal enrichment of β-catenin is not simply a reflection of the patterns of expression of structural proteins. Moreover, α-spectrin strongly stained plasma membranes between blastomeres and at the periphery of the embryo (Fig. 3 B), demonstrating that these internal regions were accessible to all reagents for immunolocalization.
Strikingly, by the 16-cell stage β-catenin is no longer restricted to the cytoplasm but is also observed in nuclei of dorsal vegetal blastomeres (Fig. 2 G). At the 32-cell stage there is a pronounced enrichment of nuclear β-catenin in the dorsal (Fig. 2 I) relative to ventral (Fig. 2 H) blastomeres. At the blastula stage, nuclei stain for β-catenin on both dorsal (Fig. 2 L) and ventral (Fig. 2 J) sides, though with greater staining on the dorsal side. Previously, Schneider et al. (1996) observed nuclear β-catenin only in dorsal regions of blastulas, but the discrepancy is likely due to the greater sensitivity of confocal microscopy. The structures examined at the 32-cell stage were confirmed to be nuclei by using the nuclear stain Bo-Pro-3 as in Molenaar et al. (1996), which labeled all nuclei, each of which resembled the β-catenin–stained nucleus in Fig. 2 I (data not shown).
To test whether the apparent differences in the spatial distribution of endogenous β-catenin in the early Xenopus embryo reflect dorso-ventral differences in steady-state levels of β-catenin, we extracted proteins from the dorsal and ventral regions of 32-cell stage embryos (Fig. 4 A) and performed immunoblot analyses as in Fig. 4 C. We found that the ventral vegetal region (VVM) of the embryo contained a statistically significant average of 64% of the β-catenin present in the dorsal vegetal region (DVM) (n = 5 experiments, P < 0.005 by Student's t test; Fig. 4 B) after normalizing β-catenin to the levels of the membrane skeleton protein α-spectrin (the same antibody as used in Fig. 3). Consistent with these results, we found the VVM region contained an average of 77% of the β-catenin present in the DVM zone after normalization to tubulin (n = 3 experiments). Smaller dorso-ventral differences were detected in the animal hemisphere (Fig. 4 B), where the ventral animal region (VAM) contained 78% of the β-catenin present in the dorsal animal region (DAM) (n = 5 experiments, P < 0.05 by Student's t test) after normalization to α-spectrin. The comparable dorso-ventral distribution of β-catenin relative to tubulin in the animal hemisphere suggests that the dorsal enrichment of β-catenin observed by confocal microscopy is shared by tubulin, while in the vegetal hemisphere β-catenin is dorsally enriched compared to both α-spectrin and tubulin. Although the α-spectrin and β-catenin antibodies were also used in immunolocalization studies (tubulin could not be used owing to high background), the extent of the dorso-ventral differences measured by confocal microscopy and Western blots cannot be compared quantitatively (see Fig. 4 legend).
Since UV irradiation of the vegetal pole during the first cell cycle blocks formation of the dorsal axis (Gerhart et al., 1989), we tested its effects on the dorso-ventral steadystate levels of β-catenin. Prospective dorsal and ventral sides were assigned before dissection with reference to blastomere size (ventral cells are larger) and pigmentation differences (ventral cells are darker). As shown in Fig. 4 B, UV-irradiated embryos dissected at the 32-cell stage no longer show a dorsal enrichment of β-catenin in either the animal or vegetal hemispheres, and instead displayed greater relative ventral levels, especially in the VVM region. We emphasize that the higher bar heights in VVM and VAM in UV-treated embryos are due to normalization to dorsal DVM and DAM and do not imply increases in absolute levels above those seen dorsally in controls. Indeed, Western blot analysis of entire embryos shows that total amounts of β-catenin remain relatively unchanged after UV irradiation sufficient to ventralize embryos; further UV irradiation reduces β-catenin levels nonspecifically (not shown). We do not know why the greater level of β-catenin on the ventral side of UV-irradiated embryos does not cause this side to become dorsal, since ectopic expression of β-catenin in UV-irradiated embryos restores axis formation (Gruger and Gumbiner, 1995; Yost et al., 1996). It is possible that the absolute level of β-catenin on the ventral side of UV-irradiated embryos is too low to promote axis formation or that the UV irradiation leads to subsequent damage to some β-catenin such that it is no longer functional.
Wnt Signaling, but Not BVg1 or Noggin, Stabilizes β-Catenin in Early Embryos
We next tested whether we could detect dorso-ventral asymmetries in the accumulation of ectopically expressed β-catenin as a prelude to asking which signaling factors might modulate this accumulation. We injected a mixture of c-myc_-epitope–tagged β-catenin and control prolactin RNAs into the marginal zone of each blastomere at the fourcell stage, followed by anti–_c-myc immunostaining of the embryos at different stages of development. Control Western blots show this leads to three- to fourfold increases in total β-catenin levels relative to uninjected embryos (data not shown). In these control embryos, the β-catenin–myc staining is evident on the dorsal side when stained either before mid-blastula transition (MBT) (Fig. 5 C, relative to uninjected embryos, A) or after MBT (Fig. 5 D, relative to uninjected embryos, B). When embryos were cultured to stage 10 (not shown), the enrichment of anti–c-myc staining colocalized with the dorsal lip in 100% of the embryos analyzed (n = 38), indicating that the accumulation of ectopic β-catenin occurs on the prospective dorsal half of the developing embryo. These results establish that the ectopic tracer β-catenin–myc accumulates in a manner similar to the spatial pattern of endogenous β-catenin (Fig. 2). Although β-catenin–myc does not accumulate on the ventral side of embryos, proteins from injected RNAs are indeed expressed, as shown by ventral injection and expression of green fluorescent protein RNA and β-galactosidase RNA (not shown). The localized accumulation of β-catenin–myc on the prospective dorsal side, despite the RNA being injected on both dorsal and ventral sides, is consistent with the proposed greater activity of Xgsk-3 on the ventral side (Pierce and Kimelman, 1995), which targets β-catenin for degradation (Yost et al., 1996).
Figure 5.
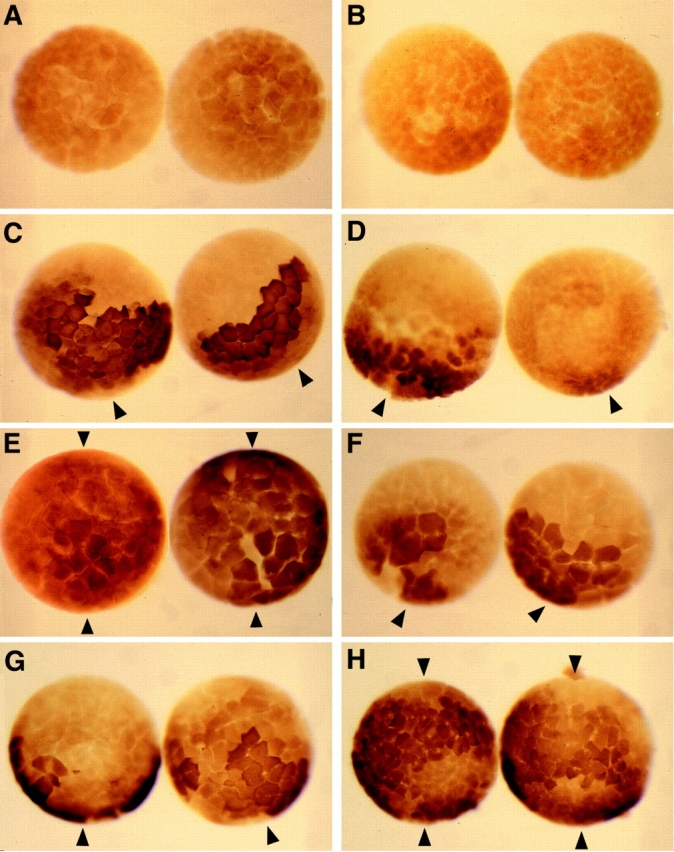
Wnts, but not BVg1 or noggin, are able to modulate ectopic β-catenin–myc accumulation both before and after MBT, as assayed by anti–c-myc immunostaining. Uninjected controls do not stain for c-myc when assayed before (A) or after (B) MBT. Embryos expressing β-catenin–myc and prolactin display a dorsal accumulation of ectopic β-catenin–myc both before (C) and after (D) MBT, while the ventral side shows negligible staining. (E) Xwnt_-8 RNA expands the domain of accumulation of β-catenin–_myc when assayed before MBT (shown, 100 pg Xwnt_-8 RNA was injected for the embryo on the left, and 3 ng RNA for the embryo on the right) or after MBT (see Table I). (F) Neither BVg1 (pre-MBT, shown; and post-MBT, Table I) nor noggin RNA (see Table I) are able to elevate β-catenin–_myc accumulation on the ventral side, while β-catenin–myc accumulates on the dorsal side similarly to control embryos. (G) Xwnt_-5A RNA does not alter the pattern of β-catenin–_myc accumulation during the early cleavage stages, but causes β-catenin–myc to accumulate around the entire marginal zone after MBT (H). In these experiments, 1 ng of _β_-catenin RNA and 3 ng of prolactin, _Xwnt_-8 (100 pg in left embryo of panel E), Xwnt_-5A, BVg1, or noggin RNA were injected into the marginal zone of each blastomere of four cell stage embryos, followed by anti–_c-myc immunostaining before and after MBT. Arrowheads denote dorsal (single arrowhead) or dorsal and ventral (double arrowhead) staining.
Since both endogenous and ectopic β-catenin accumulate on the prospective dorsal side of the embryo, we then asked whether overexpression of Wnts (for review see Du et al., 1995), BVg1 (a modified version of Vg1 which is efficiently processed; Thompsen and Melton, 1993), or noggin (Smith and Harland, 1992), all of which induce ectopic dorsal axes, would stabilize ectopic β-catenin on the ventral sides of embryos. We injected β_-catenin–myc RNA with Xwnt_-8, Xwnt_-5A, BVg1, or noggin RNA into the marginal zone of each blastomere at the four-cell stage and assayed for ectopic β-catenin accumulation by anti–_c-myc immunostaining. We found that neither ectopic BVg1 (Fig. 5 F; Table I) nor noggin (Table I) altered the dorso-ventral accumulation of β-catenin–_myc before MBT when compared to the pattern in control-injected embryos (Fig 5 C), nor did they promote increased accumulation of β-catenin–_myc after MBT (Table I). In contrast, _Xwnt_-8 consistently expanded the domain of β-catenin accumulation to span the entire marginal zone, when assayed either before MBT (Fig. 5 E, relative to control, C) or after MBT (Table I). Pointing to differences between distinct Xwnts, overexpression of _Xwnt_-5A before MBT did not alter the dorsally enriched pattern of ectopic β-catenin accumulation (Fig. 5 G, relative to control, C; Table I), which is consistent with the inability of ectopic Xwnt_-5A to induce dorsal structures in developing embryos (Moon et al., 1993; Du et al., 1995). However, anti–_c-myc immunostaining after MBT revealed that _Xwnt_-5A RNA injected at the fourcell stage is capable of stabilizing ectopic β-catenin after the onset of zygotic transcription (Fig. 5 H, relative to control, D; Table I), suggesting that at certain periods in development, _Xwnt_-8 and _Xwnt_-5A might act in part through a common pathway.
Table I.
Modulation of β-catenin–myc Accumulation
| RNAs injected* | D > V | D = V | Indistinguishable‡ | n |
|---|---|---|---|---|
| % | % | % | ||
| At 32–cell stage | ||||
| Uninjected | 63 | |||
| prolactin + β-catenin–myc | 64 | 10 | 26 | 86 |
| Before stage 7 | ||||
| Uninjected | 183 | |||
| prolactin + β-catenin–myc | 66 | 18 | 16 | 167 |
| _Xwnt_-8 (10 pg) + β-catenin–myc | 59 | 24 | 17 | 46 |
| _Xwnt_-8 (100 pg) + β-catenin–myc | 54 | 32 | 14 | 82 |
| _Xwnt_-8 (3 ng) + β-catenin–myc | 40 | 47 | 13 | 118 |
| _Xwnt_-5A (100 pg) + β-catenin–myc | 62 | 5 | 33 | 39 |
| _Xwnt_-5A (3 ng) + β-catenin–myc | 65 | 21 | 14 | 113 |
| BVg1 + β-catenin–myc | 67 | 6 | 27 | 82 |
| noggin + β-catenin–myc | 83 | 2 | 15 | 47 |
| _Xgsk_-3 + β-catenin–myc | 65 | 6 | 29 | 63 |
| prolactin + β-catenin–myc + Lithium | 30 | 47 | 23 | 89 |
| Xgsk-3 + β-catenin–myc + Lithium | 48 | 34 | 18 | 113 |
| After stage 7 | ||||
| Uninjected | 264 | |||
| prolactin + β-catenin–myc | 68 | 9 | 23 | 225 |
| _Xwnt_-8 + β-catenin–myc | 32 | 53 | 15 | 136 |
| _Xwnt_-5A + β-catenin–myc | 36 | 57 | 7 | 108 |
| BVg1 + β-catenin–myc | 58 | 14 | 28 | 123 |
| noggin + β-catenin–myc | 62 | 22 | 16 | 147 |
| prolactin + ptβ-catenin–myc | 20 | 67 | 13 | 122 |
| _dnXgsk_-3 + β-catenin-myc | 35 | 56 | 9 | 124 |
| _Xgsk_-3 + β-catenin–myc | 53 | 4 | 43 | 117 |
To further investigate whether Wnts are capable of modulating β-catenin accumulation, we injected embryos with RNAs encoding β-catenin–myc along with RNAs encoding various secreted factors and then analyzed total embryo extracts by Western blots. The blots were probed with anti–c-myc and anti–β-catenin antibodies to monitor the steady state levels of ectopic and endogenous β-catenin, respectively (Yost et al., 1996). We found that Xwnt_-8 (Fig. 6 A, lane 4) is able to increase the steady-state level of ectopic β-catenin–_myc an average of 231% when compared to embryos injected with _β_-galactosidase (Fig. 6 A, lane 2; n = 4 experiments, SE = 42%, P < 0.05 by Student's t test). Similarly, _Xwnt_-8b, a maternal Wnt gene with axis-inducing activity (Cui et al., 1995), also increased steady-state levels of ectopic β-catenin (data not shown), while Xwnt_-11 (functionally similar to Xwnt_-5A; Du et al., 1995; Torres et al., 1996) was much less effective at stabilizing ectopic β-catenin (Fig. 6 A, lane 3 relative to 4). Neither BVg1 (Fig. 6 A, lane 7 relative to control, lane 6) nor noggin (Fig. 6 A, lane 8) had a reproducible effect on β-catenin–_myc accumulation, consistent with their lack of effect on β-catenin–_myc staining in intact embryos (Fig. 5).
Figure 6.

_Xwnt_-8, but not BVg1 or noggin, is able to increase the accumulation of both ectopic and endogenous β-catenin as assayed by Western blots. (A) In multiple independent experiments (see text), probing Western blots for c-myc, microinjection of 100 pg of _β_-catenin–myc RNA with 200 pg of _β_-galactosidase (lane 2), _Xwnt_-11 (lane 3), or _Xwnt_-8 (lane 4) reveals that only Xwnt_-8 is able to increase β-catenin levels (compare lanes 2 and 4). Uninjected embryos (lane 1) do not express the c-myc epitope. In separate experiments (lanes 5–8), neither BVg1 (lane 7) nor noggin (lane 8) increases the accumulation of β-catenin above control-injected levels (lane 6), while uninjected embryos do not express β-catenin–_myc (lane 5). (B) Microinjection of 5–10 ng of control prolactin (lane 1), _Xwnt_-5A (lane 2), _Xwnt_-8 (lane 3), BVg1 (lane 4), or noggin (lane 5) RNA demonstrates that _Xwnt_-8 increases the steady state levels of endogenous β-catenin (compare lanes 1 and 3), as shown above for ectopic β-catenin. In these experiments, the animal poles of two-cell embryos were injected with RNA, followed by the extraction of protein before stage 7. To control for protein content and even loading, all endogenous β-catenin bands were normalized to both α-spectrin and tubulin signals from the same Western blot.
Turning to the effects of these secreted factors on endogenous β-catenin, Western blots of embryos injected with _Xwnt_-8 versus prolactin RNA reveal that _Xwnt_-8 elevated endogenous β-catenin (Fig. 6 B, lane 3 relative to control lane 1) an average of 254% relative to prolactin controls (n = 6 experiments, SE = 66%, P < 0.1 by Student's t test), similar to its effects on ectopic β-catenin– myc (Fig. 6 A). In these same experiments, _Xwnt_-5A, BVg1, and noggin RNAs did not reproducibly elevate endogenous β-catenin after normalization of protein levels to α-spectrin or tubulin (Fig. 6 B). We conclude that Wnts, but not BVg1 or noggin, reproducibly increase the steadystate levels of both endogenous and ectopic β-catenin.
We have previously shown that increasing the steadystate levels of endogenous β-catenin by inhibiting Xgsk-3 activity in Xenopus embryos results in a greater nuclear accumulation of β-catenin (Yost et al., 1996). This led us to test whether overexpression of _Xwnt_-8 would also lead to the nuclear accumulation of endogenous β-catenin, which would be consistent with the idea that an axis-inducing Wnt can suppress the activity of endogenous Xgsk-3 and thereby promote the nuclear accumulation of β-catenin. We injected embryos with _Xwnt_-8, _Xwnt_-5A, or prolactin RNA and stained animal cap explants of pre-MBT embryos for endogenous β-catenin. Since pre-MBT explants were used, this is a time when _Xwnt_-8 but not _Xwnt_5A stabilizes ectopic β-catenin (Fig. 5). Control prolactinexpressing cells had nuclear staining of β-catenin in a low percentage of cells (Fig. 7 A; 15% of cells scored, n = 324 cells), while ectopic _Xwnt_-8 led to strong nuclear staining of β-catenin (Fig. 7 B; 50% of cells scored, n = 537 cells). Embryos injected with _Xwnt_-5A RNA, which does not induce a secondary axis (Du et al., 1995; Torres et al., 1996), did not display elevated nuclear β-catenin (Fig. 7 C; 2% of cells; n = 423 cells).
Implication of Localized Xgsk-3 Activity in Generating Asymmetry in β-Catenin
If the observed dorso-ventral differences in β-catenin were important in the known ability of transplanted dorsal vegetal blastomeres (Gerhart et al., 1989; Kageura, 1990) and cytoplasm (Holowacz and Elinson, 1993; Fujisue et al., 1993) to induce a complete embryonic axis, then treatments of the embryo that block formation of this axis- inducing activity should affect the spatial pattern of β-catenin. We irradiated embryos with UV light at the vegetal pole in the first cell cycle to block both cortical rotation and the formation of the axis-inducing activity (Gerhart et al., 1989) and then analyzed the spatial pattern of β-catenin by confocal microscopy at the 32-cell stage. UVirradiated embryos (100%, n = 12) do not display dorsal enrichment of β-catenin (Fig. 8 A, a) relative to control embryos (Fig. 8 A, b). Some UV-irradiated embryos showed suppressed levels of β-catenin as we observed on Western blots with high levels of irradiation, and some irradiated embryos showed an increase in ventral relative to dorsal β-catenin as observed analyzing populations of embryos on Western blots (Fig. 4). These data establish a linkage between the postfertilization cortical rotation and the formation of the dorsal enrichment in β-catenin in cleavage stage embryos.
Recent data have demonstrated a postfertilization mechanism for regulating the stability of β-catenin in Xenopus embryos, as well as its accumulation in nuclei. According to this model, in the absence of a Wnt signal, Xgsk-3 directly phosphorylates β-catenin, targeting it for rapid degradation in a pathway that also involves the tumor suppressor protein adenomatosis polyposis coli (for review see Yost et al., 1996). Inhibition of Xgsk-3, which may occur in response to a Wnt signal or which theoretically could occur through a Wnt-independent mechanism, increases the stability of β-catenin and allows the accumulation of β-catenin in nuclei (Yost et al., 1996). Thus, if an endogenous Wnt signaling pathway were a candidate for establishing the observed dorso-ventral differences in levels and spatial distribution of β-catenin in 2–32-cell embryos, then one would predict that active Xgsk-3 should antagonize this Wnt pathway and reduce dorsal β-catenin levels to those observed ventrally (Yost et al., 1996). Conversely, a dominant negative (kinase-dead) Xgsk-3 (dn Xgsk-3) should mimic a Wnt signal and increase β-catenin staining when overexpressed ventrally. Consistent with these predictions, active Xgsk-3 reduces endogenous dorsal β-catenin staining in 32-cell embryos (Fig. 8 A, c, compared to controls, Fig. 2 E), while a dnXgsk-3 increases ventral β-catenin staining (Fig. 8 A, d).
Xgsk-3 activity also alters the spatial pattern of accumulation of ectopic β-catenin by the blastula stage, both before and after MBT. Injection of β_-catenin–_myc and Xgsk-3 RNA into each blastomere at the four-cell stage results in patterns of c-myc immunostaining (right embryo in Fig. 8 B, b; Table I) that resemble prolactin controls (Fig. 8 B, a) or results in background levels of staining for c-myc (Fig. 8 B, b, left embryo). As many of the embryos injected with Xgsk-3 had background levels of staining, this RNA produced the highest percentage of embryos classified as having no dorso-ventral polarity in β-catenin–myc (Table I, Indistinguishable). Consistent with its effects on endogenous β-catenin at the 32-cell stage (Fig. 8 A), expressing a dn_Xgsk-3_ causes ectopic β-catenin to accumulate to high levels everywhere at the blastula stage (Fig. 8 B, c, compared to B, a; Table I).
As treatment of embryos with lithium mimics Wnt signaling (for review see Christian and Moon, 1993), in part by inhibiting the activity of endogenous Xgsk-3 (Klein and Melton, 1996), we tested whether lithium treatment could modulate ectopic β-catenin accumulation in a manner similar to Xwnt_-8 and dn_Xgsk-3. Consistent with our expectations, lithium treatment expanded the domain of β-catenin–myc accumulation in both prolactin (Fig. 8 B, d; compared to control B, a; Table I) and Xgsk-3_–treated embryos (Fig. 8 B, e, compared to Xgsk-3 alone, in B, b). Lithium also increased the accumulation of β-catenin–_myc when analyzed by Western blots (Fig. 8 C, lane 3 relative to 2). Finally, expression of point mutation (pt)β-catenin– myc, which lacks the NH2-terminal conserved Xgsk-3 phosphorylation site required for efficient degradation of β-catenin (Yost et al., 1996), resulted in the accumulation of ptβ-catenin–myc everywhere in the embryo (Fig. 8 B, f; Table I). These data are consistent with a requirement for active Xgsk-3 in establishing the observed dorso-ventral asymmetry in endogenous and ectopic β-catenin.
Discussion
Establishment of the embryonic axes of Xenopus laevis begins with the postfertilization cortical rotation and is elaborated during the cleavage stages, as evidenced by the ability of transplanted dorsal cytoplasm and dorsal cells to induce formation of a new dorsal axis in recipient embryos (reviewed in introduction). The multifunctional protein β-catenin is required for formation of the endogenous dorsal axis in Xenopus (Heasman et al., 1994; for review see Miller and Moon, 1996), strongly suggesting that if a secreted signaling factor or downstream elements of its signal transduction cascade were involved in axis formation, then these pathways should impinge upon β-catenin. The present study investigated two heretofore unresolved issues: whether endogenous β-catenin is enriched in dorsal blastomeres by the 32-cell stage, when blastomere and cytoplasm transfer data demonstrate a dorsal determining activity is present in these cells, and whether any of the secreted signaling factors that can mimic this dorsal-determining activity can also modulate β-catenin levels. We report that endogenous β-catenin is indeed expressed at higher levels in dorsal relative to ventral blastomeres of cleavage stage embryos and that it accumulates in dorsal nuclei by the 16- to 32-cell stages. These results provide the first demonstration that an endogenous protein with a dorsaldetermining activity is asymmetrically localized along the dorso-ventral axis in the cleavage stage of a vertebrate embryo. Our data further show that ectopic Wnt ligands and Xgsk-3, a serine/threonine kinase known to function in the Wnt pathway, can modulate the dorso-ventral levels of β-catenin, while Vg1 and noggin do not.
β-Catenin, Axis Specification, and the Definition of the Nieuwkoop Center Activity
There is a remarkable agreement between our observed patterns of enrichment of β-catenin on the dorsal side of cleavage stage embryos and the ability of transplanted dorsal blastomeres or cytoplasm to initiate formation of a new axis. Specifically, cytoplasm and blastomere transfer experiments show that both dorsal vegetal and dorsal animal cells of 32-cell stage embryos, but not ventral cells, can induce a new axis when transplanted to the ventral side of a host embryo (reviewed in introduction). Strikingly, we show that β-catenin is enriched in both the cytoplasm and nuclei of both dorsal vegetal and dorsal animal cells by the 32-cell stage, and this persists through the blastula stage when zygotic transcription commences. This timing and localization of β-catenin expression is consistent with a likely role in the Nieuwkoop Center activity of the embryo.
The Nieuwkoop Center activity of Xenopus embryos has been operationally defined as the ability of transplanted cells or injected factors to elicit formation of a gastrula organizer in a non–cell autonomous manner (e.g., Smith and Harland, 1992). Thus, when noggin, BVg1, some Wnts, and _β_-catenin RNAs are injected into prospective ventral vegetal cells, which will physically become part of the endoderm, the overlying cells develop with properties of the gastrula organizer, leading to a new axis with an ectopic notochord, somites, and neural structures. These observations have led some in the field to consider that the Nieuwkoop Center activity is a discrete physical entity and/or that it is present only in the dorsal vegetal region of embryos. However, this is not consistent with blastomere transplantation experiments showing an axis-inducing activity extending into the animal hemisphere (see introduction), nor does it readily explain the mesoderm-forming ability of isolated or rotated eight-cell animal pole cells (Cardellini, 1988; Grunz, 1994; Li et al., 1996). To account for these data, we have previously suggested (Kimelman et al., 1992; Moon and Christian, 1992) that the Nieuwkoop Center comprises a large region of the dorsal side of the embryo, spreading from the dorsal animal to the dorsal vegetal cells, with maximum activity in the dorsal vegetal cells. This is the same localization as we report here for β-catenin during the cleavage stages.
The Wnt Pathway, but Not Vg1 or Noggin, Modulate β-Catenin
Given the data presented here indicating that there are greater levels of β-catenin on the dorsal side relative to the ventral side by the 32-cell stage, it is important to ask how this asymmetry arises. As a theoretical device for allowing rational discussion, one can divide the possibilities into mechanisms that require a secreted signaling factor and mechanisms that do not.
If a secreted signaling factor were involved in the mechanism that establishes the observed dorso-ventral asymmetry in β-catenin, then this signaling factor should be able to mimic the Nieuwkoop Center activity. Secreted factors meeting these criteria include a subset of the Xwnt gene family, specifically _Xwnt_-1, _Xwnt_-3A, _Xwnt_-8, and _Xwnt_-8b (for reviews see by Du et al., 1995; Torres et al., 1996), as well as the unrelated factors noggin (Smith and Harland, 1992) and Vg1 (Dale et al., 1993; Thomsen and Melton, 1993). Of the Xwnts mentioned, only _Xwnt_-8b is maternally expressed and thus a candidate for an endogenous regulator of β-catenin in cleaving embryos (Cui et al., 1995; Du et al., 1995). In the present study, we found that ectopic expression of _Xwnt_-8, and the highly related maternal _Xwnt_-8b, but not noggin (Smith and Harland, 1992) or BVg1 (Thomsen and Melton, 1993), consistently promotes the accumulation of β-catenin in embryos, as monitored by immunocytochemistry and Western blot analyses. Thus, of the small set of candidates proposed to be initiators of dorsal development, only Xwnts were able to regulate steady-state levels of β-catenin in embryos. However, there are no loss of function data showing that any Xwnt is actually required for axis specification. Since ectopic expression of a dominant negative _Xwnt_-8 blocks formation of ectopic but not endogenous axes (Hoppler et al., 1996) and since overexpression of _Xwnt_-5A blocks ectopic axis formation by _Xwnt_-8 and _Xwnt_-8b but not the endogenous axes (Torres et al., 1996), available data support the conclusion that even if Wnt ligands are involved in endogenous axis formation, they are not strictly required.
Somewhat unexpectedly, we found that overexpression of _Xwnt_-5A does not promote the stabilization of newly synthesized β-catenin assayed before the onset of zygotic expression, but when sibling embryos are cultured further and assayed for β-catenin levels after MBT, they now demonstrate much higher levels of newly synthesized β-catenin relative to control embryos. We speculate that this might be explained by the expression of new Wnt receptors (Bhanot et al., 1996; Yang-Snyder et al., 1996) after the midblastula stage, with these new receptors allowing the _Xwnt_-5A to activate an intracellular pathway that stabilizes β-catenin. Despite the interesting effects of _Xwnt_-5A on β-catenin levels after MBT, our data suggest that neither _Xwnt_-5A nor the functionally similar _Xwnt_-11 (Du et al., 1995; Torres et al., 1996) are involved in stabilizing β-catenin levels during the cleavage stages when cytoplasm and blastomere transplantation experiments have demonstrated the presence of a localized dorsal-determining activity.
Do any data discriminate between greater synthesis versus stability of β-catenin on the dorsal side? While we have not investigated regional synthesis of β-catenin owing to technical difficulties in such studies, experiments reported here and elsewhere (Domingues et al., 1995; He et al., 1995; Pierce and Kimelman, 1995; Yost et al., 1996) strongly argue for Xgsk-3 regulating the stability of newly synthesized β-catenin such that it is degraded on the prospective ventral side and stable on the dorsal side. Specifically, we injected RNA encoding c-myc_-epitope–tagged β-catenin to allow us to follow newly synthesized β-catenin. This injection protocol leads to the RNA being distributed throughout the entire marginal zone on the dorsal and ventral sides. Despite the widespread distribution of the RNA (and uniform synthesis of unrelated proteins such as β-galactosidase and green fluorescent protein, data not shown), we see the accumulation of the c-myc epitope only on the future dorsal side. This almost certainly involves synthesis of β-catenin throughout the embryo, followed by selective degradation of β-catenin on the ventral but not the dorsal side. We have presented three results in further support of this conclusion. First, coinjection of a dn_Xgsk-3 results in the accumulation of β-catenin on the ventral as well as dorsal side. Second, treating embryos with LiCl, which inhibits Xgsk-3 (Klein and Melton, 1996), similarly promotes the ventral accumulation of epitope-tagged β-catenin, as well as increasing its steady-state levels. Third, mutating the amino-terminal site of β-catenin required for phosphorylation by Xgsk-3 (Yost et al., 1996) overrides the endogenous mechanism that promotes the dorsal accumulation of β-catenin, such that the mutant β-catenin accumulates on both the ventral and dorsal sides.
We propose that the dorsal–ventral asymmetry in β-catenin in embryos before MBT is attributable to the broad synthesis of β-catenin followed by its being targeted for degradation by Xgsk-3 on the ventral side to a greater extent than on the dorsal side. This is likely a very direct effect since Xgsk-3 directly phosphorylates β-catenin in vitro and since deletion of the major in vitro phosphorylation site blocks most in vivo phosphorylation and leads to increased accumulation of β-catenin (Yost et al., 1996). A prediction of this model is that there are dorsal–ventral differences in Xgsk-3 activities, or differences in other components of the catabolic pathway for β-catenin. Further investigation is required to determine how the postfertilization cortical rotation may promote such differences.
Summary
In conclusion, we have demonstrated that β-catenin accumulates in the cytoplasm and nuclei of dorsal blastomeres of cleavage stage embryos beginning at the two-cell stage, in a manner dependent upon the postfertilization cortical rotation and regulated by Xgsk-3, a kinase in the Wnt signaling pathway. We have shown that ectopic expression of specific Xwnts , and inhibition of Xgsk-3, elevates levels of β-catenin before MBT, but the axis-inducing factors BVg1 and noggin do not. This leads us to conclude that neither endogenous Vg1 nor noggin is likely to play a role in establishing the observed asymmetries in endogenous β-catenin in cleavage stage embryos, while Xgsk-3 working directly or in response to Wnt signaling is further implicated in axis specification. These data also suggest that Xwnts stimulate ectopic axis formation through a pathway that is distinct from BVg1 and noggin.
Since elevation of steady-state levels and nuclear accumulation of dorsal β-catenin correlates with its ability to induce gene expression (Funayama et al., 1995, Kelly et al., 1995, Yost et al., 1996; Schneider et al., 1996), likely through interacting directly with HMG box architectural transcription factors (Behrens et al., 1996; Molenaar et al., 1996), we propose that greater levels of β-catenin in nuclei of dorsal blastomeres of cleavage stage embryos presage and influence dorso-ventral differences in gene expression at the blastula stage, when zygotic transcription commences. Thus, β-catenin is likely to play a key role in remodeling zygotic chromatin in dorsal blastomeres before the initiation of zygotic transcription. When zygotic transcription begins, these chromatin effects may work in a combinatorial manner with other growth factor pathways (for review see by Kimelman et al., 1992) to promote the expression of specific zygotic dorsal regulatory genes such as siamois (Lemaire et al., 1995), which is responsive to Wnt (Carnac et al., 1996; Yang-Snyder et al., 1996) and β-catenin (Brannon and Kimelman, 1996). Dorsal β-catenin may also function in other capacities, since β-catenin mimics the ability of _Xwnt_-8 to enhance gap junctional coupling in the absence of transcription (Guger and Gumbiner, 1995). Remaining challenges include determining whether the postfertilization cortical rotation of Xenopus eggs establishes the dorso-ventral differences in β-catenin levels through regulation of Xgsk-3 activity, and if so, whether this kinase is regulated in response to egg activation in a Wnt-independent or -dependent manner.
Acknowledgments
We thank M. Vo Sum (Howard Hughes Medical Institute, Seattle, WA) for assistance in preparing figures and J. Gerhart (University of California, Berkeley, CA) for discussions.
Abbreviations used in this paper
DAM
dorsal animal marginal
dnXgsk-3
dominant negative (kinase-dead) Xenopus glycogen synthase kinase-3
DVM
dorsal vegetal marginal
MBT
mid-blastula transition
MeOH
methanol
ptβ-catenin-myc
point mutant in β-catenin-myc
VAM
ventral animal marginal
VVM
ventral vegetal marginal
Xgsk-3
Xenopus glycogen synthase kinase-3
Xwnt
Xenopus Wnt
Footnotes
M. Torres, J. Miller, and R. Moon were supported by the Howard Hughes Medical Institute, and B. Rowning (HD27525 and HD 29360 to R.T. Moon), C. Yost (GM 07270), and D. Kimelman (HD27262) were supported by Public Health Service Awards from the National Institutes of Health. C. Larabell was supported by the Office of Health and Environmental Research, U.S. Department of Energy.
Note Added in Proof. β-Catenin was translated in vitro and labeled with [35S]methionine (as in Yost et al., 1996), then comparable amounts were injected into either dorsal or ventral blastomeres of four-cell embryos. Monitoring the injected β-catenin by SDS-PAGE and autoradiography demonstrates that it is more stable on the dorsal side of the embryo, as predicted by our experiments presented here and in Yost et al. (1996).
Address all correspondence to Randall T. Moon, Howard Hughes Medical Institute, Campus Box 357370, Room K536C Health Sciences Building, University of Washington School of Medicine, Seattle, WA 98195. Tel.: (206) 543-1722. Fax: (206) 543-0858.rtmoon@u.washington.edu
References
- Behrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature (Lond) 1996;382:638–642. doi: 10.1038/382638a0. [DOI] [PubMed] [Google Scholar]
- Bhanot P, Brink M, Samos CH, Hsieh J-C, Wang Y, Macke JP, Andrew D, Nathans J, Nusse R. A new member of the frizzled family from Drosophilafunctions as a Wingless receptor. Nature (Lond) 1996;382:225–230. doi: 10.1038/382225a0. [DOI] [PubMed] [Google Scholar]
- Brannon M, Kimelman D. Activation of Siamoisby the Wnt pathway. Dev Biol. 1996;180:344–347. doi: 10.1006/dbio.1996.0306. [DOI] [PubMed] [Google Scholar]
- Cardellini P. Reversal of dorsal ventral polarity in Xenopus laevisembryos by 180° rotation of the animal micromeres at the eight cell stage. Dev Biol. 1988;128:428–434. doi: 10.1016/0012-1606(88)90304-1. [DOI] [PubMed] [Google Scholar]
- Carnac G, Kodjabachian L, Gurdon JB, Lemaire P. The homeobox gene Siamoisis a target of the Wnt dorsalization pathway and triggers organizer activity in the absence of mesoderm. Development (Camb) 1996;122:2055–3065. doi: 10.1242/dev.122.10.3055. [DOI] [PubMed] [Google Scholar]
- Christian JL, Moon RT. Interactions between Xwnt-8 and Spemann organizer signaling pathways generate dorsoventral pattern in the embryonic mesoderm of Xenopus. . Genes Dev. 1993;7:13–28. doi: 10.1101/gad.7.1.13. [DOI] [PubMed] [Google Scholar]
- Christian JL, Olson DJ, Moon RT. Xwnt-8 modifies the character of mesoderm induced by bFGF in isolated Xenopusectoderm. EMBO (Eur Mol Biol Organ) J. 1992;11:33–41. doi: 10.1002/j.1460-2075.1992.tb05024.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui Y, Brown JD, Moon RT, Christian JL. Xwnt 8b: a maternally expressed Xenopus Wntgene with a potential role in establishing the dorsoventral axis. Development (Camb) 1995;121:2177–2186. doi: 10.1242/dev.121.7.2177. [DOI] [PubMed] [Google Scholar]
- Dale L, Matthews G, Colman A. Secretion and mesoderm-inducing activity of the TGFβ-related domain of XenopusVg1. EMBO (Eur Mol Biol Organ) J. 1993;12:4471–4480. doi: 10.1002/j.1460-2075.1993.tb06136.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeMarais A, Moon RT. The armadillo homologs β-catenin and plakoglobin are differentially expressed during early development of Xenopus laevis. . Dev Biol. 1992;153:337–346. doi: 10.1016/0012-1606(92)90118-z. [DOI] [PubMed] [Google Scholar]
- Domingues I, Itoh K, Sokol SY. Role of glycogen synthase kinase 3-β as a negative regulator of dorsoventral axis formation in Xenopusembryos. Proc Natl Acad Sci USA. 1995;92:8498–8502. doi: 10.1073/pnas.92.18.8498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du SJ, Purcell S, Christian JL, McGrew LL, Moon RT. Identification of distinct classes and functional domains of Wnts through expression of wild-type and chimeric proteins in Xenopusembryos. Mol Cell Biol. 1995;15:2625–2634. doi: 10.1128/mcb.15.5.2625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fagotto F, Gumbiner BM. β-catenin localization during Xenopusembryogenesis: accumulation at tissue and somite boundaries. Development (Camb) 1994;120:3667–3679. doi: 10.1242/dev.120.12.3667. [DOI] [PubMed] [Google Scholar]
- Fujisue M, Kobayakawa Y, Yamana K. Occurrence of dorsal axisinducing activity around the vegetal pole of an uncleaved Xenopusegg and displacement to the equitorial region by cortical rotation. Development (Camb) 1993;118:163–178. doi: 10.1242/dev.118.1.163. [DOI] [PubMed] [Google Scholar]
- Funayama N, Fagotto F, McCrea P, Grumbiner BM. Embryonic axis induction by the armadillorepeat domain of β-catenin: evidence for intracellular signaling. J Cell Biol. 1995;128:959–968. doi: 10.1083/jcb.128.5.959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giebelhaus DH, Zelus BD, Henchman SK, Moon RT. Changes in the expression of α-fodrin during embryonic development of Xenopus laevis. . J Cell Biol. 1987;105:843–853. doi: 10.1083/jcb.105.2.843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerhart, J.C., M. Danilchick, T. Doniach, S. Roberts, B. Rowning, and R. Stewart. 1989. Cortical rotation of the Xenopus egg: consequences for the anteroposterior pattern of embryonic dorsal development. Development (Camb.). 107(Suppl.):37–51. [DOI] [PubMed]
- Gimlich RL. Acquisition of developmental autonomy in the equatorial region of the Xenopusembryo. Dev Biol. 1986;115:340–352. doi: 10.1016/0012-1606(86)90254-x. [DOI] [PubMed] [Google Scholar]
- Grunz H. The four animal blastomeres of the eight-cell stage of Xenopus laevisare intrinsically capable of differentiating into dorsal mesodermal derivatives. Int J Dev Biol. 1994;38:69–76. [PubMed] [Google Scholar]
- Guger KA, Gumbiner BM. β-Catenin has Wnt-like activity and mimics the Nieuwkoop signaling center in Xenopusdorsal-ventral patterning. Dev Biol. 1995;172:115–125. doi: 10.1006/dbio.1995.0009. [DOI] [PubMed] [Google Scholar]
- Haegel H, Larue L, Ohsugi M, Fedorov L, Herrenknecht K, Kemler R. Lack of β-catenin affects mouse development at gastrulation. Development (Camb) 1995;121:3529–3537. doi: 10.1242/dev.121.11.3529. [DOI] [PubMed] [Google Scholar]
- He X, Saint JP, Jeannet, Woodgett JR, Varmus HE, Dawid IB. Glycogen synthase kinase-3 and dorsoventral patterning in Xenopusembryos. Nature (Lond) 1995;374:617–622. doi: 10.1038/374617a0. [DOI] [PubMed] [Google Scholar]
- Heasman J, Crawford A, Goldstone K, Garner P, Hamrick, Gumbiner B, McCrea P, Kintner C, Noro CY, Wylie C. Overexpression of cadherins and underexpression of β-catenin inhibit dorsal mesoderm induction in early Xenopusembryos. Cell. 1994;79:791–803. doi: 10.1016/0092-8674(94)90069-8. [DOI] [PubMed] [Google Scholar]
- Holowacz T, Elinson RP. Cortical cytoplasm, which induces dorsal axis formation in Xenopus, is inactivated by UV irradiation of the oocyte. Development (Camb) 1993;119:277–285. doi: 10.1242/dev.119.1.277. [DOI] [PubMed] [Google Scholar]
- Hoppler S, Brown JD, Moon RT. Expression of a dominant negative Wnt blocks induction of MyoD in Xenopusembryos. Genes Dev. 1996;10:2805–2817. doi: 10.1101/gad.10.21.2805. [DOI] [PubMed] [Google Scholar]
- Kageura H. Spatial distribution of the capacity to initiate a secondary embryo in the 32-cell embryo of Xenopus laevis. . Dev Biol. 1990;142:432–438. doi: 10.1016/0012-1606(90)90365-p. [DOI] [PubMed] [Google Scholar]
- Kelly GM, Erezyilmaz DF, Moon RT. Induction of a secondary embryonic axis in zebrafish occurs following the overexpression of β-catenin. Mech Dev. 1995;53:261–273. doi: 10.1016/0925-4773(95)00442-4. [DOI] [PubMed] [Google Scholar]
- Kimelman D, Christian JL, Moon RT. Synergistic principles of development: overlapping patterning systems in Xenopusmesoderm induction. Development (Camb) 1992;116:1–9. doi: 10.1242/dev.116.Supplement.1. [DOI] [PubMed] [Google Scholar]
- Klein PS, Melton DA. A molecular mechanism for the action of lithium during early development. Proc Natl Acad Sci USA. 1996;93:8455–8459. doi: 10.1073/pnas.93.16.8455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klymkowsky MW, Maynell LM, Polson AG. Polar asymmetry in the organization of the cortical cytokertain system of Xenopus laevisoocytes and embryos. Development (Camb) 1987;100:543–557. doi: 10.1242/dev.100.3.543. [DOI] [PubMed] [Google Scholar]
- Ku M, Melton DA. Xwnt-11: maternally expressed Xenopus Wntgene. Development (Camb) 1993;119:1161–1173. doi: 10.1242/dev.119.4.1161. [DOI] [PubMed] [Google Scholar]
- Larabell CA, Rowning BA, Wells J, Wu M, Gerhart JC. Confocal microscopy analysis of living Xenopuseggs and the mechanism of cortical rotation. Development (Camb) 1996;122:1281–1289. doi: 10.1242/dev.122.4.1281. [DOI] [PubMed] [Google Scholar]
- Lemaire P, Garrett N, Gurdon JB. Expression cloning of Siamois, a Xenopushomeobox gene expressed in dorsal-vegetal cells of blastulae and able to induce a complete secondary axis. Cell. 1995;81:85–94. doi: 10.1016/0092-8674(95)90373-9. [DOI] [PubMed] [Google Scholar]
- Li SH, Mao ZR, Yan SY, Grunz H. Isolated dorsal animal blastomeres of Xenopus laevisare capable to form mesodermal derivatives, while the ventral animal blastomeres differentiate into ciliated epidermis only. Zool Sci. 1996;13:125–131. [Google Scholar]
- McMahon AP, Moon RT. Ectopic expression of the proto-oncogene int-1 in Xenopusembryos leads to duplication of the embryonic axis. Cell. 1989;58:1075–1084. doi: 10.1016/0092-8674(89)90506-0. [DOI] [PubMed] [Google Scholar]
- Miller JR, Moon RT. Signal transduction through β-catenin and specification of cell fate during embryogenesis. Genes Dev. 1996;10:2527–2539. doi: 10.1101/gad.10.20.2527. [DOI] [PubMed] [Google Scholar]
- Molenaar M, van de Wetering M, Oosterwegel M, Peterson-Maduro J, Godsave S, Korinek V, Roose J, Destree O, Clevers H. XTcf-3 transcripton factor mediates β-catenin-induced axis formation in Xenopusembryos. Cell. 1996;86:391–399. doi: 10.1016/s0092-8674(00)80112-9. [DOI] [PubMed] [Google Scholar]
- Moon RT, Christian JL. Competence modifiers synergize with growth factors during mesoderm induction and patterning in Xenopus. . Cell. 1992;71:709–712. doi: 10.1016/0092-8674(92)90545-n. [DOI] [PubMed] [Google Scholar]
- Moon RT, Campbell RM, Christian JL, McGrew LL, Shih J, Fraser S. Xwnt-5A, a maternal Wnt that affects morphogenetic movements after overexpression in embryos of Xenopus laevis. . Development (Camb) 1993;119:97–111. doi: 10.1242/dev.119.1.97. [DOI] [PubMed] [Google Scholar]
- Pierce SB, Kimelman D. Regulation of Spemann organizer formation by the intracellular kinase Xgsk-3. . Development (Camb) 1995;121:755–765. doi: 10.1242/dev.121.3.755. [DOI] [PubMed] [Google Scholar]
- Sakai M. The vegetal determinants required for the Spemann organizer move equatorially during the first cell cycle. Development (Camb) 1996;122:2207–2214. doi: 10.1242/dev.122.7.2207. [DOI] [PubMed] [Google Scholar]
- Schneider S, Steinbeisser H, Warga RM, Hausen P. β-Catenin translocation into nuclei demarcates the dorsalizing centers in frog and fish embryos. Mech Dev. 1996;57:191–198. doi: 10.1016/0925-4773(96)00546-1. [DOI] [PubMed] [Google Scholar]
- Smith W, Harland RM. Expression cloning of noggin, a new dorsalizing factor localized in the Spemann organizer in Xenopusembryos. Cell. 1992;70:829–840. doi: 10.1016/0092-8674(92)90316-5. [DOI] [PubMed] [Google Scholar]
- Sokol S, Melton DA. Interaction of Wnt and activin in dorsal mesoderm induction in Xenopus. . Dev Biol. 1992;154:348–355. doi: 10.1016/0012-1606(92)90073-p. [DOI] [PubMed] [Google Scholar]
- Spencer M, Giebelhaus DH, Kelly GM, Bicknell J, Florio SK, Milam AH, Moon RT. Membrane skeleton protein 4.1 in developing Xenopus: expression in postmitotic cells of the retina. Dev Biol. 1990;139:279–291. doi: 10.1016/0012-1606(90)90297-v. [DOI] [PubMed] [Google Scholar]
- Thomsen GH, Melton DA. Processed Vg1 is an axial mesoderm inducer in Xenopus. . Cell. 1993;74:433–441. doi: 10.1016/0092-8674(93)80045-g. [DOI] [PubMed] [Google Scholar]
- Torres MA, Yang-Snyder JA, Purcell SM, DeMarais AA, McGrew LL, Moon RT. Activities of the Wnt-1 class of secreted signaling factors are antagonized by the Wnt-5A class and by a dominant negative cadherin in early Xenopusdevelopment. J Cell Biol. 1996;133:1123–1137. doi: 10.1083/jcb.133.5.1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang-Snyder J, Miller JR, Brown JD, Lai C-J, Moon RT. A frizzled homolog functions in a vertebrate Wntsignaling pathway. Curr Biol. 1996;6:1302–1306. doi: 10.1016/s0960-9822(02)70716-1. [DOI] [PubMed] [Google Scholar]
- Yost C, Torres M, Miller JR, Huang E, Kimelman D, Moon RT. The axis-inducing activity, stability, and subcellular distribution of β-catenin is regulated in Xenopusembryos by glycogen synthase kinase 3. Genes Dev. 1996;10:1443–1454. doi: 10.1101/gad.10.12.1443. [DOI] [PubMed] [Google Scholar]
- Yuge M, Kobayakawa Y, Fujisue M, Yamana K. A cytoplasmic determinant for dorsal axis formation in an early embryo of Xenopus laevis. . Development (Camb) 1990;110:1051–1056. doi: 10.1242/dev.110.4.1051. [DOI] [PubMed] [Google Scholar]