Dendritic Cell Maturation Overrules H-2d–Mediated Natural Killer T (Nkt) Cell Inhibition: Critical Role for B7 in Cd1d-Dependent Nkt Cell Interferon γ Production (original) (raw)
Abstract
Given the broad expression of H-2 class Ib molecules on hematopoietic cells, antigen presentation pathways among CD1d expressing cells might tightly regulate CD1d-restricted natural killer T (NKT) cells. Bone marrow–derived dendritic cells (BM-DCs) and not adherent splenocytes become capable of triggering NK1.1+/T cell receptor (TCR)int hepatic NKT cell activation when (a) immature BM-DCs lack H-2Db−/− molecules or (b) BM-DCs undergo a stress signal of activation. In such conditions, BM-DCs promote T helper type 1 predominant CD1d-restricted NKT cell stimulation. H-2 class Ia–mediated inhibition involves more the direct H-2Db presentation than the indirect Qa-1b pathway. Such inhibition can be overruled by B7/CD28 interactions and marginally by CD40/CD40L or interleukin 12. These data point to a unique regulatory role of DCs in NKT cell innate immune responses and suggest that H-2 class Ia and Ib pathways differentially control NKT cell recognition of DC antigens.
Keywords: inhibitory receptors, IFN-γ, costimulation, CD1d, NKT cells
Introduction
NKT cells are a recently described subpopulation of TCRα/β+CD4+ or CD4−CD8− T cells that have distinctive phenotypic and functional properties 1 2. These T cells were first identified in mice, where they can be distinguished from conventional T cells by their expression of the NK locus-encoded C type lectin molecule NK1.1. Another hallmark of murine NKT cells is their extremely restricted TCR repertoire, with the great majority expressing an invariant TCRα chain structure (Vα14-Jα281) paired preferentially with Vβ8, 7, or 2. Both mouse and human NKT cells rapidly secrete cytokines associated with both Th1 (IFN-γ) or Th2 (IL-4) responses upon TCR engagement 1 2 or stimulation with the synthetic CD1d ligand, the α-Galactosylceramide (α-GalCer) 3 4 5.
Recent results suggest a major role for murine NKT cells in the rejection of malignant tumors 6 7 8 9 10 and in regulating autoimmunity 11 and defense against certain pathogens. Indeed, NKT cells were shown to be relevant in innate antitumor immunosurveillance after IL-12 6 9 or α-GalCer administration 10 12 but also in spontaneous, endogenous IL-12–dependent tumor models in mice 8. Cytokines play an important part at the NKT cell effector phase 8 12. However, several conditions exist in which the apparent Th0 profile of cytokine production may be skewed toward a predominance of IFN-γ 11 12 or IL-4 13 14 and IL-13 15 leading to immunosuppression.
The origin and identity of the natural antigens recognized by CD1d-restricted T cells remain unknown. Given the canonical α chains and limited β chain diversity of the TCR of NKT cells, mCD1d may recognize a single or conserved set of antigens. Whether CD1d binding sphingolipid or phospholipid compounds represent self or foreign antigens 16, and whether the NKT cells responding to the synthetic α-GalCer are the same as those seeing phospholipids is unclear. However, mCD1d-restricted T cells varied in their recognition of different mCD1d transfected tumor cells suggesting that antigens presented by mCD1d molecules differ according to the cell type 17 18. Variations in CD1d-restricted antigen presentation could arise from differences in expression, trafficking, processing, or loading of antigens 19.
As dendritic cells (DCs) are cornerstones between innate 20 and cognate immune responses, we investigated the regulatory mechanisms underlying the DC/NKT cell cross-talk in vitro. Our data imply that potent H-2Db–mediated inhibition constitutively prevents DC-mediated IFN-γ production from resting NKT cells. The direct H-2D molecular pathway is partially involved in H-2 class I–mediated inhibition. However, after stress-induced maturation, DCs become electively capable of triggering CD1d-restricted NKT cells toward a Th1 predominant pattern in a B7/CD28-dependent manner, thereby overcoming H-2 class I–mediated inhibitory pathways. Our results provide further evidence for the pivotal role of DCs in NKT cell immune responses and identify DCs as a potential source of endogenous CD1d-restricted antigens.
Materials and Methods
Mice.
Female C57BL/6 (H-2b) mice were obtained from the Centre d' Elevage Janvier (Le Genest St Isle, France), the Centre d' Elevage Iffa Credo (L'Arbresle, France), and maintained in our animal facilities according to the Animal Experimental Ethics Committee Guidelines. Single H-2Kb−/− and H-2Db−/− and double H-2Kb−/−Db−/− knockout (KO) mice as well as triple β2microglobulin−/−H-2Kb−/−Db−/− KO mice were generated as described previously 21, bred in the animal facility of Institut Pasteur, and used at the sixth backcross generation onto C57BL/6 mice. CD1d−/− single and CD1d−/−H-2Kb−/−Db−/− triple KO mice were generated as described previously 22, bred at Princeton Animal Facility or Pasteur Institute, and used at the seventh backcross generation onto C57BL/6 mice. All female mice were used at 6–25 wk of age.
Purification of Hepatic NKT Cells and T Cells.
Hepatic NKT cells (defined as NK1.1+TCR-βint cells) and T cells (NK1.1− TCR-βhigh cells) were prepared as described previously 23 24. Briefly, hepatic mononuclear cells were stained with FITC-conjugated TCRβ (H57–597; BD PharMingen) and PE-conjugated NK1.1 (PK-136; BD PharMingen) and sorted by FACSVantage™ (Becton Dickinson). Purity of sorted cells exceeded 95% (data not shown). After overnight culture in complete medium, NKT cells could be restained using the anti-CD94 mAb coupled to PE (BD PharMingen).
Generation of DCs.
Bone marrow (BM)-derived DCs (BM-DCs) were propagated from BM progenitor cells in culture medium supplemented with 1,000 IU/ml of rmGM-CSF (R&D Systems) and 1,000 IU/ml of rmIL-4 (R&D Systems) as described previously 20. Culture medium was renewed at day 2 and 4. To induce stress and maturation of DCs, day 6 DCs were harvested, spun down, and transferred into new 6-well plates (referred to as “transferred” henceforth). For phenotypic analyses, cells were preincubated with culture supernatant of hybridoma (2.4G2) secreting anti-CD16/CD32 mAb, and were subsequently incubated with FITC-conjugated anti–I-Ab (AF6–120.1) and PE-conjugated anti-CD11c (HL-3), FITC-conjugated CD80 (16–10A1), CD86 (GL1), CD40 (3/23) or PE-conjugated CD1d (1B1) or the anti-H2Db (B22.249.R19; from H. Lemke, Institüt Für Genetik, Köln, Germany) or anti–H-2Kb (AF6–88.5.3 from American Type Culture Collection [HB-158]) mAb. All antibodies except 2.4G2 were purchased from BD PharMingen. Cells were gated according to size and granulosity with exclusion of PI-positive cells. Residual B lymphocytes (B220+ cells) and granulocytes (Gr1+ cells) were detected in the CD11c−/I-Ab− cells and constitute <20% of whole cell population. T and NK cells were not propagated in these DC culture conditions. BM-DCs derived from gene targeted mice were analyzed for MHC class Ia and Ib expression and exhibited levels of MHC class II, CD80, CD86, CD40 expression comparable to those of wild-type (wt) BM-DCs (see Fig. 1, and data not shown). Splenic adherent cells were generated by subjecting red blood cell deprived splenocytes to 3 h adherence at 37°C. Adherent cells were rinsed three times with PBS, incubated with 10 mM EDTA for 20 min, and analyzed in FACScan™ for CD11c, MHC class II, CD1d, CD40, CD80, or CD86 expression.
Figure 1.
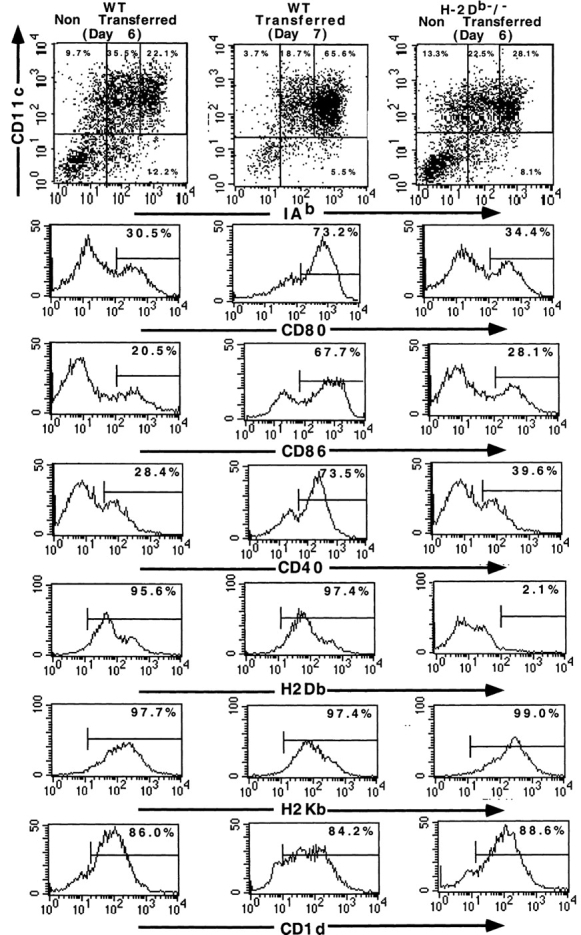
Phenotypic analyses of ex vivo propagated BM-DCs. BM-DCs (GM-CSF plus IL-4) at day 6 (wt, left panels, and H-2Db−/−, right panels) before/or at day 7 after harvesting and transfer into new wells (wt, middle panels) were subjected to two color staining (CD11c-PE and I-Ab-FITC) or one-color staining (CD1d-PE and CD40, CD80, CD86-FITC, indirect staining for H-2Db and H-2Kb expression) and immunostained cells were analyzed in flow cytometry.
In Vitro Cocultures.
The hepatic NK1.1+TCR-βint NKT cells or NK1.1−TCR-βhigh T cells (5 × 104) were cocultured with immature or mature BM-DCs (2 × 103–104–5 × 104) in 200 μl of complete medium in 96-well U-bottomed plates for 24–44 h at 37°C, 5% CO2. As positive controls, DCs were pulsed with 10 ng/ml of α-GalCer (provided by Pharmaceutical Research Laboratories, KIRIN Brewery Co., Gunma, Japan), washed in PBS, and cocultured with NKT cells. For blocking experiments, BM-DCs and NKT cells were cocultured in the presence of 50 μg/ml of neutralizing anti-CD40L mAb (MR; BD PharMingen), 50 μg/ml of CTLA-4 Ig fusion protein (gift from the Genetics Institute, Cambridge, MA), 25 μg/ml of anti–IL-12 mAb (C17.8; BD PharMingen), and of isotype-matched rat or hamster mAb (BD PharMingen). To mimic the stimulatory effects of B7 on mature DCs, we incubated immature day 6 BM-DCs with NKT cells along with 10 or 50 μg/ml of stimulating anti-CD28 mAb PV.1 (provided by C. June, University of Pennsylvania, Philadelphia, PA) plus or minus 100 IU rhIL-2/ml. To investigate the inhibitory pathways, immature DCs derived from wt mice were incubated with neutralizing anti-H2Db (B22.249.R19; from H. Lemke, Köln, Germany) or anti–H-2Kb (AF6–88.5.3 from American Type Culture Collection [HB-158]) mAb and immature DCs derived from Db−/− mice were incubated with increasing dosages (1–5-10 μM) of Qdm1 (Qa-1b binding peptide i.e. AMAPRTLLL) or R5K (mock Qa-1b binding peptide i.e. AMVPKTLLL provided by D. Raulet, University of Berkeley, Berkeley, CA) peptides for 24 h at 37°C. The Qa-1b mAb (clone 6A8.6F10.1A6) was purchased from BD PharMingen. All coculture conditions were performed in duplicates and experiments performed at least three times.
Cytokine Detection and Quantification (mIFNγ and mIL-4).
After 20–44 h DC/NKT or T coculture, supernatants were harvested, stored at −80°C, and assessed either directly or after 5–50 times dilution using commercial ELISA kits (OptEIATM ELISA kit; BD PharMingen). The sensitivity of the mIFNγ kit was >31.5 pg/ml and that of mIL-4 kit was >7.15 pg/ml.
Statistical Analyses of Cytokine Levels.
Fisher's exact method was used to compare means ± SE of IFN-γ production in-between various culture conditions and significant differences at 95% confidence are depicted with * on each graph.
Results and Discussion
Constitutive Inhibition of the DC/NKT Cell Cross-Talk.
We first investigated the capacity of ex vivo propagated BM-DCs at an immature stage of differentiation to trigger cytokine secretion from resting hepatic NK1.1+/TCRint NKT cells in vitro. After 6 d of in vitro culture in GM-CSF plus IL-4, 50–55% BM cells became CD11c+/MHC class II+, expressed CD1d, and a small fraction was MHC class IIbright and expressed CD80, CD86, and CD40 molecules (“non transferred DC (day6),” Fig. 1). Such immature DCs, when cocultured with liver NKT cells at various DC/NKT cell ratios, did not induce NKT cell triggering as already reported (3 12 23; Fig. 2, a–c). Similar data were achieved using immature DCs propagated in GM-CSF alone or DCs sorted from spleens of Flt3L-treated mice (data not shown). As reported previously 23, when immature CD1d+ DCs were pulsed with α-GalCer (1–10 ng/ml), high levels of IFN-γ (Fig. 3 a) and IL-4 (not shown) were measured in the supernatants of DC/NKT cell cocultures.
Figure 2.



Constitutive H-2D–mediated NKT cell inhibition by immature DCs. (a) Day 6 immature BM-DCs derived from B6.wt or H-2Kb−/−Db−/− double KO or H-2Kb−/−Db−/−β2m−/−triple KO mice were cocultured at a DC/NKT cell ratio of 1:1 or 1:25 with cell sorted hepatic NK1.1+/TCRint NKT cells for 40 h. (b) Similar experiments were performed comparing BM-DCs with adherent splenocytes derived from B6. wt versus H-2Kb−/−Db−/− double KO versus CD1d−/− Kb−/− Db−/− triple KO mice cocultured with NKT cells at a DC/NKT cell ratio of 1:1 for 40 h. (c) Similar experiments were performed comparing immature day 6 BM-DCs derived from single (H-2Kb−/− or H-2Db−/−) KO mice versus H-2 Kb−/−Db−/− double KO mice (left panels). 24 and 40 h supernatants were assayed for IFN-γ in ELISA. Right panel depicts IFN-γ levels in the supernatants of NKT cells with day 6 wt BM-DCs incubated with increasing amounts of neutralizing Ab anti–H-2Db or H-2Kb. Means ± SE are depicted in all graphs. Each graph represents either pooled data from three experiments performed in duplicate wells or a representative experiment out of three. Significant differences at 95% confidence using Fisher's exact method are outlined with *. All experiments were performed with KO female mice of 6–8 wk of age after 6–8 backcrosses on C57BL/6 mice. Phenotypic features were comparable in all gene targeted BM-DCs except for H-2 or CD1d molecule expression.
Figure 3.




Stress triggers DC maturation, promoting CD1d-restricted, B7-dependent NKT cell activation. (a) Day 6 immature BM-DCs pulsed or not pulsed with 10 ng/ml of α-GalCer were compared with mature day 7 BM-DCs in a coculture setting with hepatic NKT cells or T cells at a DC/NKT or T cell ratio of 1:5 for 40 h. (b) Cocultures of day 7 transferred DCs derived from B6.wt versus CD1d−/− mice with hepatic NK1.1+ TCR-βint NKT cells or hepatic NK1.1− TCR-βhigh T cells at various DC/NKT ratios were assayed for mIFN-γ (left panels) and at DC/NKT cell ratio of 1:5 for mIL-4 (right panel) using ELISA for 40 h. (c) Day 7 mature DCs were cocultured with NKT cells at a DC/NKT cell ratio of 1:5 for 24 h in the presence of 50 μg/ml of neutralizing mAb directed against mIL-12 (C17.8), mCD40L (MR1), or isotype-matched Ab or CTLA4Ig fusion proteins (left panel). Day 6 wt BM-DC were coincubated with NKT cells at a 1:5 ratio with increasing dosages of the stimulating anti-CD28 Ab PV1 plus or minus 100 IU/ml rhuIL-2 (right panel) for 48 h. Supernatants were assayed for IFN-γ in ELISA. Means ± SE are depicted in all graphs. Each graph depicts one representative experiment out of three performed in duplicate wells. Significant differences at 95% confidence using Fisher's exact method are outlined with *.
However, when derived from H-2Db−/−Kb−/− class I gene–targeted mice, immature DCs became capable of triggering IFN-γ secretion at nanogramm levels from hepatic NKT cells in a dose-dependent manner (Fig. 2 a). In contrast, low levels of IL-4 (100–300 pg/ml) were detected in the supernatants of NKT cells cocultured with H-2Db−/− Kb−/− gene-targeted BM-DCs (not shown). However, such nanogram levels of IFN-γ secretion were not found using BM-DCs propagated from H-2Db−/−Kb−/− β2microglobulin−/− triple KO mice instead of Db−/−Kb−/− double KO mice (Fig. 2 a) suggesting that the constitutive DC/NKT cell cross-talk involves β2microglobulin-associated H-2 class Ib molecules.
The DC/NKT Cell Cross-Talk Is Dependent on CD1d Molecules.
We formally demonstrated that CD1d play a dominant role in the constitutive DC/NKT cell cross-talk by comparing IFN-γ levels secreted by NKT cells in coculture with BM-DCs derived from H-2Db−/−Kb−/− CD1d−/− versus H-2Db−/−Kb−/− mice originating from similar backcrosses in B6 mice (Fig. 2 b). DCs were electively endowed with the constitutive capacity to present CD1d-restricted antigens since CD1d+/CD86+/I-Ab+ adherent splenocytes did not promote IFN-γ secretion from NKT cells even when derived from H-2Db−/−Kb−/−mice (Fig. 2 b).
Direct Role of H-2D Molecules in the Constitutive Inhibition of the DC/NKT Cell Cross-Talk.
It has been shown that NKT cell maturation is accompanied by extinction of Ly49 receptor expression. Liver NKT cells, as opposed to thymus NKT cells, are mature NKT cells and do not express Ly-49A, Ly-49C/I, or Ly-49-G 25. However, Skold and Cardell 26 pointed out the differential regulation of Ly49 expression on CD4+ and double negative NK1.1+ T cells with dramatic organ specific variations, stressing the complexity of NKT cell inhibitory pathways. Importantly, we were able to exclude a role for the H-2Kb/Ly49C pathway in the constitutive DC/NKT cell inhibition and to show a dominant inhibitory role of H-2Db molecules at early time points of the DC/NKT cell interaction (Fig. 2 c, left panel). Searching for the H-2 Db–specific inhibitory ligand, one should consider the possibility that H-2Db recognition could be direct and/or indirect. Indirect recognition is possible via the ubiquitously expressed Qa-1b molecules which preferentially associate with a H-2Db leader peptide (Qdm), this peptide representing ∼70% of those eluted from Qa-1 molecules 27 28. A CD94-NKG2A NKT cell inhibition would not rule out the coexistence of an additional inhibitory receptor interacting directly with H-2Db molecules. Therefore, we assessed the role of these inhibitory pathways by either (a) pulsing Qa-1b binding peptides (i.e. Qdm1 AMAPRTLLL or the mock peptides R5K (AMVPKTLLL) that does not bind Qa-1) at increasing dosages on H-2Db/− BM-DC, (b) or using the neutralizing mAb anti–H-2Db or anti–H-2Kb pulsed onto wt day 6 BM-DCs before NKT cell coculture. Only 20–25% of hepatic NKT cells (TCRβint/NK1.1+) express CD94 molecules (among which not all coexpress NKG2A molecules) i.e. up to 4% of bulk liver mononuclear cells (not shown). The amounts of IFN-γ secreted from NKT cells in coculture with day 6 immature wt versus H-2Db−/− BM-DCs incubated with increasing dosages of Qa-1b binding peptides were compared (1–30 μM of Qdm1 or R5K mock peptides). No significant inhibition of IFN-γ production was observed using Qdm1 peptides pulsed onto H-2Db−/− BM-DCs (not shown). In accordance with this result, the neutralizing anti–Qa-1b mAb did not allow IFN-γ production by NKT cells cocultured with wt day 6 BM-DCs (not shown). The right panel of Fig. 2 c investigates the direct inhibitory pathway and compares the secretion levels of IFN-γ by NKT cells stimulated by wt day 6 BM-DCs incubated with 10 to 50 μg/ml of neutralizing anti–H-2Db−/− or anti–H-2Kb−/− Ab. Significant IFN-γ secretion is found only using the anti–H-2Db−/− Ab, although nanogram levels were achieved using mature DCs (not shown on this graph). Therefore, in line with the well known heterogeneity of the NKT cell population, a complex inhibitory pathway likely participates in maintaining DC-mediated tolerance of liver resident NKT cells.
These data imply that H-2 class Ia and Ib molecules differentially and constitutively prevent DC-mediated IFN-γ production from resting liver NKT cells and that CD1d-restricted antigens are presented electively by DC to NKT cells in vitro.
Stressed DCs Become Capable of Stimulating NKT Cells.
However, in stress conditions shown to be mimicked by cell harvest and transfer into new plastic dishes 29 or by LPS, wt DCs became activated and acquired a more mature phenotype i.e. MHC class IIbright, CD80+, CD86+, and CD40+ (“transferred” day 7 DCs, Fig. 1) displaying potent allostimulatory capacity (not shown) while CD1d was not significantly overexpressed (Fig. 1). Moreover, the expression levels of the H-2K and H-2D molecules were comparable at day 7 (Fig. 1). After their transfer at day 6, day 7 wt DCs acquired the capacity to foster NKT cells toward a Th1 predominant secretion pattern while pulsing with KRN7000 in such conditions led to secretion of both IL-4 and IFN-γ at high levels (not shown). High levels of IFN-γ were measured in 22–40 h coculture supernatants (Fig. 3 a) while IL-4 was barely detectable (Fig. 3 b). Mature DCs stimulated NKT cells in a dose-dependent manner (Fig. 3 b), with optimal effects at a ratio of 1 DC:1 NKT cell (Fig. 3 b). Similar NKT cell activation was obtained using LPS-activated DCs (not shown). No significant thymidine incorporation by NKT cells was measured in proliferation assays and trypan blue exclusion did not show increased numbers of viable NKT cells after 40 h incubation with mature DCs (not shown). The mature DC/NKT cell cross-talk was significantly inhibited when DCs were generated from B6.CD1d−/− mice (Fig. 3 b). Inhibition was also clear for IL-4 secretion (Fig. 3 b). The phenotype of mature DCs in B6 wt and B6.CD1d−/− mice was comparable (not shown). Eberl et al. 30 pointed out a tissue-specific segregation of CD1d-dependent and -independent NKT cells, hepatic NKT cells being mostly CD1d dependent. In similar conditions of B6 liver cell sorting, CD3+/NK1.1− hepatic T cells also produce, albeit less efficiently, detectable amounts of IFN-γ (but no IL-4) when placed in close contact with mature DCs. It is noteworthy that, in contrast to NKT cell activation, T cell activation was not regulated by CD1d molecules nor α-GalCer (Fig. 3 b). It is unlikely that IFN-γ is secreted from DCs rather than NKT cells in these cocultures as only CD8+ DCs were shown to produce IFN-γ after stimulation with ng levels of both IL-12 and IL-18 31.
The DC maturation allowed to overcome H-2 class Ia–mediated constitutive NKT cell inhibition by DCs, as comparable levels of IFN-γ were secreted by NKT cells in contact with wt versus Kb−/−Db−/− mature DCs (data not shown). However, the synthetic α-GalCer–mediated CD1d stimulation was more potent, even at low dosage (1–10 ng/ml) than the self DC antigens and Th0 biased. The low levels of CD1d-dependent IL-4 secretion by NKT cells triggered by mature DCs could be accounted for by the relative weakness of the CD1d-mediated presentation of DC self-antigens (Fig. 3, a and b) and/or discrete threshold of reactivity of various hepatic NKT cell subsets.
The B7/CD28 Pathway Overrides the H-2Db-mediated NKT Cell Inhibition.
We next investigated the mechanisms accounting for NKT cell IFN-γ secretion after contact with stressed DCs. During α-GalCer–mediated NKT cell stimulation, IL-12 and CD40 were shown to play a critical role 3 12. However, despite the Th1 predominant pattern, IL-12 p70 was not detectable in ELISA in NKT cell cocultures with mature DCs where significant IFN-γ levels were measured (not shown). However, as neutralizing anti–IL-12 mAb decreased IFN-γ secretion, albeit not significantly (mostly at early time points 24 h of the coculture), it remains conceivable that IL-12 be involved after the initial CD1d/TCR interaction step (Fig. 3 c, left panel). Moreover, in transwell experiments whereby mature DCs and NKT cells are physically separated by a porous membrane, IFN-γ was not produced suggesting a dominant role for a membrane-associated molecule in the DC/NKT cell cross-talk (data not shown). Therefore, we next investigated whether costimulatory molecules upregulated upon maturation could account for circumvention of H-2 class I–mediated inhibition. Saturating concentrations of neutralizing mAb directed against CD40L or B7 molecules were used in the DC/NKT cell cocultures. While a partial but not significant inhibition was observed using neutralizing anti-CD40L mAb, CTLA4Ig fusion proteins were shown to be necessary and sufficient to completely abrogate the DC/NKT cell cross-talk in vitro. Both, NKT cell IFN-γ (Fig. 3 c, left panel) and IL-4 secretion (not shown) were suppressed. To confirm that CD28 engagement is sufficient to trigger hepatic NKT cell IFN-γ production by DCs, wt BM-DCs at day 6 (immature cells) were cocultured with hepatic NKT cells in the presence of stimulating anti-CD28 mAb (PV.1) with or without low doses of IL-2 (100 IU/ml). H-2 class I–mediated inhibition could be efficiently overcome using the anti-CD28 mAb plus IL-2 with a synergistic effects between the cytokine and the costimulatory factor (Fig. 3 c, right panel). The IFN-γ levels were fairly comparable with those achieved using mature BM-DCs (day7). Altogether, the B7/CD28 pathway is critical to account for the ability of activated DCs to trigger NKT cell IFN-γ production by circumventing class I inhibition.
A Schematic View of the DC/NKT Cell Cross-Talk In Vitro.
These data demonstrate that stress-induced DC activation allows to overcome constitutive inhibitory pathways through B7/CD28 interaction, thereby promoting CD1d-dependent IFN-γ production by NKT cells in vitro (Fig. 4). The role of CD1d and B7 expression levels, of DC endogenous antigens and/or tissue origin need to be investigated to ascribe these regulatory NKT activation pathways to BM-DCs. We showed that DCs and not adherent splenocytes (antigen-presenting cells expressing high levels of CD1d molecules, and low levels of I-Ab, B7.2 molecules), when devoid of inhibitory receptor ligand, trigger NKT cell activation. Such a difference may be accounted for by tissue origin. Indeed, while activated BM-DCs could trigger IFN-γ production, Flt3L splenic DCs after overnight activation were not able to activate NKT cells (not shown). It is also possible that B7.1 is critical to overcome H-2 class I–mediated inhibition because adherent splenocytes do not express B7.1. Intracellular trafficking of CD1d molecules may play a critical role in determining the antigens presented by DCs.
Figure 4.

Putative schematic representation of the DC/NKT cell cross-talk. Tissue resident DCs (immature stage) constitutively inhibit CD1d-dependent NKT cell IFN-γ production (first signal: CD1d/TCR) through a mechanism involving H-2 class I molecules (second signal: H-2D/Ly49? +/− CD94/NKG2A ?). The inhibitory pathway might be dominant because a third accessory pathway overruling the dominant negative signal is lacking on immature DCs. In contrast, after stress (LPS, transfer), DCs acquire a third accessory signal (B7/CD28) that is dominant over the second inhibitory pathway allowing CD1d-restricted DC antigen presentation. It is conceivable that in pathological conditions where high IFN-γ levels are required, DCs become capable of triggering NKT cell recognition of self and/or foreign antigens presented by MHC class Ib molecules to the canonical TCR.
It is noteworthy that DC-mediated NKT cell activation is leading predominantly to Th1 secretion pattern in vitro, suggesting that this regulatory pathway may be relevant in the setting of intracellular pathogens, viruses, or tumors. IFN-γ is a pleiotropic cytokine playing a central role in promoting innate and adaptive mechanisms of host defense. IFN-γ not only fosters T cell–mediated immunity but also activates macrophages, promotes antigen processing, and presentation by antigen-presenting cell and prevents viral replication, angiogenesis, and transformed cell growth 32. Therefore, it is conceivable that dysregulation of DC functions suggested in autoimmune disorders or tumors might foster NKT cell reactivity toward inappropriate Th pattern. Further investigations will be required to unravel the in vivo dynamics between DCs and NKT cells during infection, autoimmunity, or tumor development but such data outline that CD1d-restricted DC antigens might be relevant in such disorders.
Acknowledgments
We thank KIRIN Brewery Company Ltd. (Tokyo, Japan) for providing KRN7000 to the National Cancer Center. We are indebted to the animal facility staff of IGR and to Valérie Schiavon for technical assistance. We are grateful to Nadine Fernandez and D. Raulet for helpful discussion and advice.
Y. Ikarashi was supported by the research fellowship program of collaborative works for Japanese researchers in foreign institutes, i.e., the Japanese Foundation for Promotion of Cancer Research supported by the Ministry of Health and Welfare for the second Term Comprehensive 10-yr strategy for cancer control. R. Mikami was supported by the Ministry of Foreign Affairs. The work was also supported by the french fellowship GEFLUC, LIGUE française contre le cancer, and Association de la Recherche le Cancer (ARC).
Footnotes
Y. Ikarashi and R. Mikami contributed equally to this work.
References
- Bendelac A., Riversa M.N., Park S.H., Roark J.H. Mouse CD1-specific NK1+ T cellsdevelopment, specificity, and function. Annu. Rev. Immunol. 1997;15:535–562. doi: 10.1146/annurev.immunol.15.1.535. [DOI] [PubMed] [Google Scholar]
- MacDonald H.R. NK1.1+ T cell receptor α/β cellsnew clues to their origin, specificity and function. J. Exp. Med. 1995;182:633–638. doi: 10.1084/jem.182.3.633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawano T., Cui J., Koezuka Y., Toura I., Kaneko Y., Motoki K., Ueno H., Nakagawa R., Sato H., Kondo E. CD1d-restricted and TCR-mediated activation of Vα14 NKT cells by glycosylceramides. Science. 1997;278:1626–1629. doi: 10.1126/science.278.5343.1626. [DOI] [PubMed] [Google Scholar]
- Spada F.M., Koezuka Y., Porcelli S.A. CD1d-restricted recognition of synthetic glycolipid antigen by human natural killer T cell. J. Exp. Med. 1998;188:1529–1534. doi: 10.1084/jem.188.8.1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burdin N., Brossay L., Koezuka Y., Smily S.T., Gurusby M., Gui M., Taniguchi M., Hayakawa K., Kronenberg M. Selective ability of mouse CD1 to present glycolipids. α-galactosylceramide specifically stimulates Vα14 NKT lymphocytes. J. Immunol. 1998;161:3271–3281. [PubMed] [Google Scholar]
- Cui J., Shin T., Kawano T., Sato H., Kondo E., Toura I., Kaneko Y., Koseki H., Kanno M., Taniguchi M. Requirement for Vα14 NKT cells in IL-12-mediated rejection of tumor. Science. 1997;278:1623–1626. doi: 10.1126/science.278.5343.1623. [DOI] [PubMed] [Google Scholar]
- Kawano T., Cui J., Koezuka Y., Toura I., Kaneko Y., Sato H., Kondo E., Harada M., Koseki H., Nakayama T. Natural killer-like nonspecific tumor lysis mediated by specific ligand-activated Vα14 NKT cells. Proc. Natl. Acad. Sci. USA. 1998;95:5690–5693. doi: 10.1073/pnas.95.10.5690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smyth M.J., Thia K.Y.T., Street S.E.A., Cretney E., Trapani J.A., Taniguchi M., Kawano T., Pelikan S.B., Crowe N.Y., Godfrey D.I. Differential tumor surveillance by natural killer (NK) and NKT cells. J. Exp. Med. 2000;191:661–668. doi: 10.1084/jem.191.4.661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smyth M.J., Taniguchi M., Street S.E. The anti-tumor activity of IL-12mechanisms of innate immunity that are model and dose-dependent. J. Immunol. 2000;165:2265–2670. doi: 10.4049/jimmunol.165.5.2665. [DOI] [PubMed] [Google Scholar]
- Nakagawa R., Motoki K., Nakamura H., Ueno H., Iijima R., Yamaguchi A., Tsuyuki S., Inamoto T., Koezuka Y. Antitumor activity of α-galactosylceramide, KRN7000, in mice with EL-4 hepatic metastasis and its cytokine production. Oncol. Res. 1998;10:561–568. [PubMed] [Google Scholar]
- Wilson S.B., Kent S.C., Patton K.T., Orban T., Jackson R.A., Exely M., Porcelli S., Schatz D., Atkinson M.A., Balk S.P. Extreme Th1 bias of invariant Vα24JaQT cells in type 1 diabetes. Nature. 1998;391:177–181. doi: 10.1038/34419. [DOI] [PubMed] [Google Scholar]
- Kitamura H., Iwakabe K., Yahata T., Nishimura S., Ohta A., Ohmi Y., Sato M., Takeda K., Okumura K., Van Kaer L. The natural killer T (NKT) cell ligand α-galactosylceramide demonstrates its immunopotentiating effect by inducing interleukin (IL)-12 producing by dendritic cell and IL-12 receptor expression on NKT cells. J. Exp. Med. 1999;189:1121–1127. doi: 10.1084/jem.189.7.1121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burdin N., Brossay L., Kronenberg M. Immunization with α−galactosylceramide polarizes CD1d-reactive NK T cells towards Th2 cytokine synthesis. Eur. J. Immunol. 1999;29:2014–2025. doi: 10.1002/(SICI)1521-4141(199906)29:06<2014::AID-IMMU2014>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- Yoshimoto T., Bendelac A., Watson C., Hu-Li J., Paul W.E. Role of NK1.1+ T cells in a Th2 response and in IgE production. Science. 1995;270:1845–1847. doi: 10.1126/science.270.5243.1845. [DOI] [PubMed] [Google Scholar]
- Terabe, M., S. Matsui, N. Noben-Trauth, H. Chen, C. Watson, D.D. Donaldson, D.P. Carbone, W.E. Paul, and J.A. Berzofsky. NKT cell-immunosurveillance by IL-13 and the IL-4R-STAT6 pathway. Nat. Immunol. 1:515–520. [DOI] [PubMed]
- Gumperz J.E., Roy C., Makowaska A., Lum D., Sugita M., Podrebarac T., Koezuka Y., Porcelli S.A., Cardell S., Brenner M.B., Behar S.M. Murine CD1d-restricted T cell recognition of cellular lipids. Immunity. 2000;12:211–221. doi: 10.1016/s1074-7613(00)80174-0. [DOI] [PubMed] [Google Scholar]
- Brossay L., Tangri S., Bix M., Cardell S., Locksley R., Kornenberg M. Mouse CD1-autoreactive T cells have diverse patterns of reactivity to CD1+ targets. J. Immunol. 1997;160:3681–3688. [PubMed] [Google Scholar]
- Park S.H., Roark J.H., Bendelac A. Tissue-specific recognition of mouse CD1 molecules. J. Immunol. 1998;160:3128–3134. [PubMed] [Google Scholar]
- Chiu Y.H., Jayawardena J., Weiss A., Lee D., Park S.H., Dautry-Varsat V., Bendelac A. Distinct subset of CD1d-restricted T cells recognize self-antigen loaded in different cellular compartments. J. Exp. Med. 1999;189:103–110. doi: 10.1084/jem.189.1.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez N.C., Lozier A., Flament C., Ricciardi-Castagnoli P., Bellet D., Suter M., Perricaudet M., Tursz T., Maraskovsky E., Zitvogel L. Dendritic cells directly trigger NK cell functioncross-talk relevant in innate anti-tumor immune responses in vivo. Nat. Med. 1999;5:405–411. doi: 10.1038/7403. [DOI] [PubMed] [Google Scholar]
- Vugmeyster Y., Glas R., Pérarnau B., Lemonnier F.A., Eisen H., Ploegh H. Major histocompatibility complex (MHC) class I KbDb−/− deficient mice possess functional CD8+ T cells and natural killer cells. Proc. Natl. Acad. Sci. USA. 1998;95:1249–12497. doi: 10.1073/pnas.95.21.12492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park S.H., Guy-Grand D., Lemonnier F.A., Wang C.R., Bendelac A., Jabri B. Selection and expansion of CD8α/α+ T cell receptor α/β+ intestinal intraepithelial lymphocytes in the absence of both classical major histocompatibility complex class I and non classical CD1 molecules. J. Exp. Med. 1999;190:885–890. doi: 10.1084/jem.190.6.885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinohara K., Ikarashi Y., Maruoka H., Miyata M., Sugimura T., Terada M., Wakasugi H. Functional and phenotypical characteristics of hepatic NK-like T cells in NK1.1+ and − mouse strains. Eur. J. Immunol. 1999;29:1871–1878. doi: 10.1002/(SICI)1521-4141(199906)29:06<1871::AID-IMMU1871>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- Azuma M., Kato K., Ikarashi Y., Asada-Mikami R., Maruoka H., Takaue Y., Saito A., Wakasugi H. Cytokines production of U5A2-13-positive T cells by stimulation with glycolipid α-galactosylceramide. Eur. J. Immunol. 2000;30:2138–2146. doi: 10.1002/1521-4141(2000)30:8<2138::AID-IMMU2138>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- MacDonald H.R., Rosemary K., Lees K., Held W. Developmentally regulated extinction of Ly-49 receptor expression permits maturation and selection of NK1.1+ T cells. J. Exp. Med. 1998;187:2109–2114. doi: 10.1084/jem.187.12.2109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skold M., Cardell S. Differential regulation of Ly49 expression on CD4+ and CD4−CD8− (double negative) NK1.1+ T cells. Eur. J. Immunol. 2000;30:2488–2496. doi: 10.1002/1521-4141(200009)30:9<2488::AID-IMMU2488>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- Vance R.E., Kraft J.R., Altman J.D., Jensen P.E., Raulet D.H. Mouse CD94/NKG2A is a natural killer cell receptor for the nonclassical MHC class I molecule Qa-1b . J. Exp. Med. 1998;188:1841–1848. doi: 10.1084/jem.188.10.1841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kraft J.R., Vance R.E., Phol J., Martinet A.M., Raulet D.H., Jensen P.E. Analysis of Qa-1(b) peptide binding specificity and capacity of CD94/NKG2A to discriminate between Qa-1-peptide complexes. J. Exp. Med. 2000;192:613–624. doi: 10.1084/jem.192.5.613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallucci S., Lolkema M., Matzinger P. Natural adjuvantsendogenous activators of dendritic cells. Nat. Med. 1999;5:1249–1255. doi: 10.1038/15200. [DOI] [PubMed] [Google Scholar]
- Eberl G., Lees R., Smily S.T., Taniguchi M., Grusby M.J., MacDonald H.R. Tissue-specific segregation of CD1d-dependent and CD1d independent NKT cells. J. Immunol. 1999;162:6410–6419. [PubMed] [Google Scholar]
- Ohteki T., Fukao T., Suzue K., Maki C., Ito M., Nakamura M., Koyasu S. Interleukin 12-dependent interferon γ production by CD8α+ lymphoid dendritic cells. J. Exp. Med. 1999;189:1981–1986. doi: 10.1084/jem.189.12.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan D.H., Shankaran V., Dighe A.S., Stockert E., Aguet M., Old L.J., Schreiber R.D. Demonstration of an interferon γ−dependent tumor surveillance system in immunocompetent mice. Proc. Natl. Acad. Sci. USA. 1998;95:7556–7561. doi: 10.1073/pnas.95.13.7556. [DOI] [PMC free article] [PubMed] [Google Scholar]