Matrix Metalloproteinases (MMPs) Regulate Fibrin-invasive Activity via MT1-MMP–dependent and –independent Processes (original) (raw)
Abstract
Cross-linked fibrin is deposited in tissues surrounding wounds, inflammatory sites, or tumors and serves not only as a supporting substratum for trafficking cells, but also as a structural barrier to invasion. While the plasminogen activator-plasminogen axis provides cells with a powerful fibrinolytic system, plasminogen-deleted animals use alternate proteolytic processes that allow fibrin invasion to proceed normally. Using fibroblasts recovered from wild-type or gene-deleted mice, invasion of three-dimensional fibrin gels proceeded in a matrix metalloproteinase (MMP)-dependent fashion. Consistent with earlier studies supporting a singular role for the membrane-anchored MMP, MT1-MMP, in fibrin-invasive events, fibroblasts from MT1-MMP–null mice displayed an early defect in invasion. However, MT1-MMP–deleted fibroblasts circumvented this early deficiency and exhibited compensatory fibrin-invasive activity. The MT1-MMP–independent process was sensitive to MMP inhibitors that target membrane-anchored MMPs, and further studies identified MT2-MMP and MT3-MMP, but not MT4-MMP, as alternate pro-invasive factors. Given the widespread distribution of MT1-, 2-, and 3-MMP in normal and neoplastic cells, these data identify a subset of membrane-anchored MMPs that operate in an autonomous fashion to drive fibrin-invasive activity.
Keywords: matrix metalloproteinases, MT-MMP, fibrin, proteolysis, invasion
Introduction
In response to tissue damage, inflammation, or neoplastic growth, a provisional matrix largely composed of cross-linked fibrin and fibronectin is deposited at the affected site (1, 2). While the fibrin scaffolding functions initially as a structural support for infiltrating fibroblasts, endothelial cells, leukocytes, or tumor cells, the cross-linked network also acts as a physical barrier to cell traffic (2–14). Currently, invading cells are thought to use proteinases to access the fibrin matrix with proteolysis purposefully restricted to the pericellular milieu of the ingressing cells (3, 14, 15). In this manner, the overall structural integrity of the matrix would be preserved as a mechanical support for propulsive movement (3, 5, 6, 11–15).
Based on the ability of the plasminogen activator-plasminogen axis to rapidly dissolve intravascular fibrin deposits, it has been generally assumed that cell surface-associated plasmin plays a key, if not necessary, role in driving the fibrin-invasive phenotype in the extravascular compartment (5, 6, 11, 12). However, in plasminogen-null animals, fibrin-rich tissues are infiltrated efficiently by normal as well as neoplastic cell types despite significant defects in intravascular fibrinolysis (3, 9, 16–18). Consequently, increased attention has focused on the identification of alternate proteolytic systems that would allow cells to infiltrate fibrin matrices via a plasminogen-independent process. Recently, we identified an unexpected role for matrix metalloproteinases (MMPs)* in fibrin-invasive activity (3). The MMP gene family consists of 25 proteinases, of which 18 are secreted and 7 are membrane-associated either as transmembrane- or GPI-anchored enzymes (19–21). In our earlier studies, a membrane-anchored MMP, termed MT1-MMP, which is expressed in fibroblasts and endothelial cells, was able to confer a fibrin-invasive phenotype to invasion-incompetent, recipient cells after transfection (3). However, in the absence of specific MT1-MMP inhibitors or gene-deleted animals, the functional importance of this metalloproteinase in a physiologically relevant cell population has remained the subject of conjecture. This issue has been further complicated by the fact that more recent studies have demonstrated that an expanding number of secreted MMPs likewise express fibrin(ogen)olytic activity (3, 22–27). Given the recent generation of MT1-MMP–deleted mice (28, 29), we undertook studies to determine the ability of this proteinase as well as other fibrin(ogen)olytic MMPs to regulate fibrin invasion. We now demonstrate that whereas wild-type fibroblasts utilize MMPs to penetrate cross-linked fibrin gels under physiologic conditions, MT1-MMP–null fibroblasts unexpectedly display only a transient defect in fibrin-invasive activity. Further studies reveal that an additional subset of membrane-anchored MMPs, including MT2-MMP and MT3-MMP, but not MT4-MMP, allow cells to traverse fibrin matrices independently of MT1-MMP. These findings identify a triad of MT-MMPs whose expression, alone or in combination, is sufficient to confer fibrin-invasive activity. We posit that normal or neoplastic cells expressing any of these three MT-MMPs will display potent fibrin-invasive activity that functions independently of the plasminogen activator-plasminogen axis.
Materials and Methods
Cell Culture.
MDCK and CHO-K1 cell lines (both from American Type Culture Collection) were maintained in DMEM and RPMI 1640 (GIBCO BRL), respectively, supplemented with 10% fetal bovine serum (Hyclone), 2 mM L-glutamine, and antibiotics as described (30). Stably transfected MDCK cells were maintained in medium containing 700 μg/ml G418 (Life Technologies). Mouse fibroblasts were isolated from dermal explants of MT1-MMP null or wild-type littermates as described (31) and used between the third and sixth passages. All cultures were maintained at 37°C in 5% CO2/95% air.
Expression Vector Construction and Transfection.
Expression vectors including FLAG-tagged MT1-MMP, MT2-MMP, and MT3-MMP as well as stably transfected MDCK cells overexpressing MMPs 2, 3, 7, 9, 11, 13, MT1-, 2-, and 3-MMPs have been described in detail previously (30). To generate FLAG-tagged full-length MT4-MMP, primers encoding the FLAG epitope (5′-GACTACAACGACGACGATGACAAG-3′) were inserted after the COOH-terminal leucine of full-length MT4-MMP (provided by M. Seiki, University of Tokyo, Tokyo, Japan) and the epitope-tagged construct cloned into the pSG5 vector. Soluble His-tagged MT1-, MT2-, and MT3-MMPs were generated by inserting primers encoding the His epitope (5′-ATGATGATGATGATGATG-3′) after the COOH-terminal glycine of soluble MT1-MMP (Met1-Gly535), the COOH-terminal asparagine of soluble MT2-MMP (Met1-Asn625), and the COOH-terminal alanine of soluble MT3-MMP (Met1-Ala564) using a PCR-based method and cloned into pCR3.1-Uni. An expression vector for α1 antitrypsin Portland (α1PDX) was constructed from cDNA provided by G. Thomas (Oregon Health Sciences Center, Portland, OR [32]). CHO-K1 cells were transiently transfected with purified plasmid DNA using Fugene (Roche) according to the manufacturer's instructions. The activity of the soluble MT-MMPs was monitored with the quenched fluorescent peptide (7-methoxycoumarin-4-yl)acetyl-prolyl-leucyl-glycyl-leucyl-[3-(2,4-dinitrophenyl)-L-2,3-diaminopropionyl]-alanyl-arginamide (Bachem) as described (33).
Reverse Transcription PCR Analysis.
RNA was isolated from MT1-MMP wild-type or null fibroblasts using TRIzol reagent (Life Technologies). Reverse transcription was performed as described previously (3). cDNAs of MMPs were amplified by PCR using specific oligonucleotide primers for MT1-MMP (sense 5′-TGCGACGGGAACTTTGACACCG-3′; antisense 5′-TCCTCCTCAGTCCCCTCATC-3′; 607 bp product from 1201T to 1808A), MT2-MMP (sense 5′-ACATGTCCACCATGCGCTCT-3′; antisense 5′-TACCATGATGTCAGCCTCC-3′; 391 bp product from 222A to 613A), MT3-MMP (sense 5′-CGGTGTACCAGACCAGACAA-3′; antisense 5′-ATTGGGATTTCCCAGTGTCC-3′; 402 bp product from 357C to 756T). The identity of the isolated cDNAs was confirmed by sequence analysis (3).
Invasion, Motility, and Fibrinolysis Assays.
Fibrin gels were cast in the upper wells of 24-mm Transwell dishes (3 μm pore size; Corning, Inc.) by combining 0.5 ml of plasminogen-free human fibrinogen (6 mg/ml; Calbiochem) with 0.5 ml of human thrombin (4 U/ml; Sigma-Aldrich) in Hanks Balanced Salt Solution (pH 7.4) alone or supplemented with 200 μg/ml aprotinin (Sigma-Aldrich [3, 10]). Type I collagen gels were prepared as described previously (30). Because the fibrinogen preparation contains transglutaminase and fibronectin, the fibrin gel contains γ-γ crosslinks, α-polymers, and covalently-incorporated fibronectin (3, 4). Where indicated, fibrinogen preparations were depleted of fibronectin by gelatin-affinity chromatography (34). The gels were incubated at 37°C for 2 h to allow complete gelling. To vary fibrin fiber size, polymerization rates were varied by incubating thrombin-fibrinogen mixtures at pH 6.9, 7.4, or 8.0 (35). After the fibrin gels were cast at the indicated pH, 1–5 × 105 MDCK cells, CHO-K1 cells, wild-type, or MT1-MMP null fibroblasts in complete medium containing 10% fetal bovine serum were cultured atop the fibrin matrix. After 24 h, recombinant scatter factor/hepatocyte growth factor (SF/HGF) was added to the lower chamber at 50 ng/ml for MDCK cells (30). CHO-K1 cells were transiently transfected with the indicated plasmids and SF/HGF (50 ng/ml) added to the bottom well 24 h later. Wild-type or MT1-MMP null fibroblasts were similarly treated, but invasion stimulated with platelet-derived growth factor (PDGF)-BB (herein referred to as PDGF, 15 ng/ml; Calbiochem). Fibroblast motility was monitored by culturing cells atop fibrin gels whose surface was partially covered with glass cloning chips (3). After confluent layers of fibroblasts were formed, the chips were removed leaving denuded areas wherein the distance migrated by the advancing front of cells (three cells or more) was measured during a 24 h incubation. Where indicated, recombinant tissue inhibitor of metalloproteases-1 (TIMP-1; 50 μg/ml), tissue inhibitor of metalloproteases-2 (TIMP-2; 20 μg/ml), or the synthetic MMP inhibitor, BB-94 (5 μM final concentration in 0.1% DMSO; gift of British Biotechnology, Oxford, UK) was included in the fibrin gel and added to the medium (3, 30). Endotoxin-free TIMP-1 and TIMP-2 (Fuji Chemicals) were active-site titrated with gelatinase A before use (specific activity of TIMP-2 was 2.5 times greater than that of TIMP-1). All media, including SF/HGF or PDGF and inhibitors was exchanged every 2 d during the assay period (MDCK invasion was routinely terminated at 5 d, CHO-K1 invasion at 5 d, and fibroblast invasion at 6 d unless indicated otherwise). The number of invasive foci and depth of invasion (mean depth of the three front-leading cells/field) were determined from digitized images of 10 randomly selected 40× fields of hematoxylin and eosin (H and E)-stained cross sections (3, 30). All results were confirmed with stable transfectants in three or more cloned cell lines.
Fibrinolytic activity was determined by culturing cells atop fibrin gels containing 125I-labeled fibrinogen (Amersham Pharmacia Biotech) in the presence of aprotinin (200 μg/ml) with or without BB-94 as described above. Fibrin degradation products in the cell-free media and in 5 M urea extracts of the fibrin gel were pooled and quantitated by γ-scintillation counting (3). Spontaneous release of 125I-fibrin from gels incubated in the absence of cells was subtracted from all values and the results presented as mean cpm ± 1 SD.
Western Blots.
The expression of MMP proteins in transiently transfected CHO cells was assessed by Western blotting. FLAG-tagged MT-MMP proteins were recovered from Triton-X114 extracted cells as described previously (32). To detect soluble, His-tagged MT1, MT2, and MT3-MMPs, CHO-K1 cells were transferred to serum-free medium 24 h after transfection and incubated for an additional 24 to 48 h. Cell-free supernatants and detergent extracts (in the case of the full-length MT-MMPs) were resolved on 10% polyacrylamide gels under reducing conditions and the proteins transferred to nitrocellulose membranes (32). FLAG-tagged proteins were detected with mouse monoclonal antibody M2 against a FLAG epitope (1:1,000; Sigma-Aldrich), while soluble MT-MMPs were detected with mouse monoclonal Penta-His antibody (1:2,000; QIAGEN). Blots were probed with a horseradish peroxidase-conjugated anti–mouse IgG secondary antibody and bands visualized using the ECL system according to the manufacturer's instructions (Pierce Chemical Co.).
Histology and Electron Microscopy.
For histological sections, fibrin or type I collagen gels were fixed in 4% paraformaldehyde in phosphate buffered saline (pH 7.4), dehydrated through a graded ethanol series, and embedded in paraffin. Sections were cut at 7 μm and stained with hematoxylin and eosin (3, 30). For electron microscopy, fibrin gels were fixed in 2% glutaraldehyde/1.5% paraformaldehyde in 0.1 M sodium cacodylate buffer, postfixed in 1% osmium tetroxide, and dehydrated through graded ethanol. Samples were then embedded in epoxy resin, 500 nm sections cut, and stained with uranyl acetate and lead citrate. For scanning electron microscopy (SEM) analyses, after dehydration in ethanol, samples were incubated in 50:50 ethanol:hexamethyldisilazane followed by a 20-min incubation in 100% hexamethyldisilazane. Air dried specimens were mounted, sputter coated with gold, and viewed on an Amray 1000-B scanning electron microscope.
Results
Fibrinolytic and Fibrin-Invasive Activity of MMPs in Stably Transfected Cells.
Matrix remodeling events at sites of fibrin deposition in vivo are associated with the expression of a complex mix of soluble MMPs, including MMP-1 (collagenase-1), MMP-13 (collagenase-3), MMP-2 (gelatinase A), MMP-9 (gelatinase B), MMP-3 (stromelysin-1), MMP-11 (stromelysin-3), and MMP-7 (matrilysin) (1, 16, 18, 36). Each of these MMPs has been reported to degrade fibrin(ogen) under serum-free and cell-free conditions (3, 22–24, 26, 27), but the ability of these proteinases to mediate fibrin-degradative or invasive events by intact cells under physiologically relevant conditions is unknown. Consequently, invasion-incompetent cells engineered to stably overexpress each of these fibrinolytic MMPs were cultured atop 125I-labeled fibrin gels in the presence of 10% serum and motility stimulated by exogenous HGF/SF. Fibrinolysis as well as invasion were then monitored during a 5-d incubation period. As shown in Fig. 1 A, each of the MMP transfectants as well as the control-transfected cells expressed fibrinolytic activity that could be suppressed by the peptidomimetic MMP inhibitor, BB-94 (5 μM). However, neither the control-transfected cells nor any of the clones that overexpressed the secreted MMPs penetrated the underlying fibrin gels (Fig. 1, A and B). By contrast, stable clones overexpressing the membrane-anchored proteinase, MT1-MMP, not only expressed fibrinolytic activity, but as described previously (3), displayed fibrin-invasive activity as well (Fig. 1, A and B). As expected, MT1-MMP–dependent fibrinolytic and invasive activity were both inhibited in the presence of BB-94 (77 ± 6% and 100 ± 0% for fibrinolysis and invasion, respectively; mean ± 1 SD, n = 4).
Figure 1.


Fibrin-degradative and invasive activities of MDCK stable transfectants overexpressing secreted MMPs or MT1-MMP. (A) Control vector-transfected or MMP-expressing transfectants were cultured atop 125I-labeled fibrin gels in complete medium with 10% FBS and SF/HGF (50 ng/ml) for 5 d in the absence (light gray bars) or presence of 5 μM BB-94 (dark gray bars). Solubilized 125I-fibrin was monitored as described in the Materials and Methods and expressed as the mean cpm ± 1 SD (n = 4). (B) H and E–stained transverse sections of control vector-transfected and MMP-overexpressing clones cultured atop fibrin gels as described above in the presence of SF/HGF for 5 d. Only MT1-MMP transfectants expressed invasive behavior (arrows) which was completely blocked in the presence of BB-94. The double-headed arrow in the first panel demarcates the underlying fibrin gel which is largely transparent in H and E–stained sections. Bar = 100 μm.
Regulation of Fibroblast-Fibrin Interactions by MT1-MMP.
Despite the fact that MT1-MMP (a) uniquely conferred fibrin-invasive activity to transfected cells relative to all of the secreted MMPs tested and (b) is known to be expressed in fibroblasts at sites of fibrin deposition in vivo (16, 18, 36), the role of this, or other, MMPs in regulating fibroblast-fibrin interactions has not been defined previously. To determine whether fibroblast-derived MMPs regulate fibrin-invasive activity, wild-type mouse cells were cultured atop fibrin gels and motility stimulated with PDGF in the absence or presence of class-specific antiproteinases. After 2 d in culture, PDGF-stimulated fibroblasts had penetrated the underlying gel with peak invasion occurring by day 6 (Fig. 2, A and C) . As assessed by transmission electron microscopy (TEM), 4 to 6-d-old cultures revealed zones of cleared fibrin juxtaposed to the invading cells (Fig. 2 B) which correlated with the detection of degraded fibrin in the fluids expressed from the gels (at day 4, wild-type fibroblasts released 6,540 cpm 125I-fibrin and 17,380 cpm at day 6; n = 2). While fibroblast invasion was unaffected by inhibitors directed against serine proteinases (aprotinin, soybean trypsin inhibitor, or plasminogen-depletion), cysteine proteinases (E-64), or aspartyl proteinases (pepstatin A), PDGF-stimulated cells were unable to penetrate the fibrin gels in the presence of BB-94 during the 6 d culture period (Fig. 2 A; 100 ± 0% inhibition; mean ± 1 SD, n = 3).
Figure 2.


Invasive and migratory behavior of MT1-MMP+/+ and MT1-MMP−/− fibroblasts. (A) H and E–stained transverse sections of wild-type littermate and MT1-MMP-null fibroblasts cultured atop fibrin gels in the presence of 10% FBS with invasion stimulated by the addition of PDGF to the basilar compartment. Invasion of MT1-MMP+/+ fibroblasts commenced at day 2 (top left panel) and peaked at day 6 (middle panel). Invasion of MT1-MMP-null fibroblasts was not observed in most sections at day 2 (bottom left panel), but accelerated to near-normal values by day 6. In the presence of BB-94, invasion of wild-type and null fibroblasts was inhibited completely. Bar = 100 μm. (B) TEM analysis of fibrin-invasive MT1-MMP+/+ and MT1-MMP−/− fibroblasts. Invasive wild-type and null fibroblasts were both pocketed by zones of cleared fibrin (arrows). Bars = 5 μm for MT1-MMP+/+ and 2 μm for MT1-MMP−/− samples. (C) MT1-MMP+/+ and MT1-MMP−/− fibroblasts were cultured atop fibrin gels in the absence or presence of PDGF and the number of invasive foci monitored at the indicated times. Results are expressed as the mean number of invading cells/field ± 1 SD (n = 3). (D) Two-dimensional migration of PDGF-stimulated MT1-MMP+/+ and MT1-MMP−/− fibroblasts that were cultured atop fibrin gels and denuded by wounding in the absence or presence of BB-94 for 24 h. Arrow marks the edge of the confluent fibroblast layer after wounding as viewed by phase contrast microscopy. Bar = 100 μm.
Wound-associated fibroblasts have been shown to express MMP-2, MMP-11, and MT1-MMP in vivo (16, 18, 36), but only the latter proteinase scored positive in our screen of pro-invasive MMPs. Consequently, MT1-MMP-null fibroblasts were isolated from the gene-deleted animals and assessed for their ability to invade the fibrin gels under conditions identical to those used for the wild-type cells. After the first 2 d in culture, PDGF-stimulated MT1-MMP null cells displayed a striking deficiency in invasive potential (Fig. 2, A and C) with the number of invasive foci reaching levels only 27 ± 0.3% of control value (mean ± 1 SD; n = 3). Surprisingly, however, the defect in invasive activity was only transient as the null cells began to invade the fibrin matrix after a 3-d lag where invasion reached 60–70% of control values by day 6 (Fig. 2 C). Similar to the behavior of the wild-type cells, zones of fibrin clearing were found in association with the invading null fibroblasts which also proteolyzed fibrin comparably to control cells (i.e., 16,260 cpm 125I-fibrin released by null fibroblasts versus 17,380 cpm for wild-type cells by day 6; n = 2; Fig. 2 B).
To determine whether the delayed fibrin-invasive activity of MT1-MMP–deficient fibroblasts could be linked to the expression of other MMP family members, the null fibroblasts were stimulated with PDGF atop fibrin gel in the presence of BB-94. As shown in Fig. 2 A, BB-94 completely prevented fibrin invasion by the null cells (100 ± 0% inhibition; mean ± 1 SD, n = 3). Significantly, while MMP activity has been linked to the migratory response of other cell types across matrix-coated surfaces (e.g., references 37–41), the two-dimensional movement of PDGF-stimulated wild-type or null cells cultured atop fibrin gels was not inhibited by BB-94 (Fig. 2 D) and leading front determinations demonstrated that wild-type cells incubated in the absence or presence of BB-94 migrated 0.69 ± 0.04 mm and 0.67 ± 0.03 mm, respectively, while null cells migrated 0.65 ± 0.07 mm and 0.66 ± 0.04 mm in the absence or presence of BB-94 (mean ± 1 SD, n = 3).
Characterization of MT1-MMP-Independent Fibrin-Invasive Activity.
As BB-94 inhibits secreted as well as membrane-anchored MMPs with near equivalent efficiency (3, 30), the MT1-MMP-independent process used by the null fibroblasts could be mediated by either group of MMPs. Consequently, we took advantage of the ability of the endogenous MMP inhibitor, TIMP-1, to preferentially inhibit secreted MMPs in contrast to the ability of the related inhibitor, TIMP-2, to target secreted as well as membrane-anchored MMPs (3, 19, 30). Hence, wild-type and MT1-MMP null fibroblasts were cultured atop fibrin gels and stimulated with PDGF in the absence or presence of equimolar concentrations of active TIMP-1 or TIMP-2. As shown in Fig. 3 , while the invasive activity of wild-type or null fibroblasts was unaffected by TIMP-1, invasion was completely inhibited by TIMP-2 (Fig. 3, A and B). The inhibitory effects of TIMP-2 were confined to three-dimensional invasion as neither TIMP-2 nor TIMP-1 affected the two-dimensional migratory response of fibroblasts stimulated atop fibrin gels (data not shown).
Figure 3.


Effect of TIMP-1 and TIMP-2 on the fibrin-invasive activity of MT1-MMP+/+ and MT1-MMP−/− fibroblasts. (A) H and E–stained transverse sections of PDGF-stimulated MT1-MMP+/+ and null fibroblasts cultured atop fibrin gels in either 10% FBS alone or supplemented with TIMP-1 (50 μg/ml) or TIMP-2 (20 μg/ml) for 6 d. Bar = 100 μm. (B) The number of invasive foci and depth of invasion was quantified for MT1-MMP+/+ and MT1-MMP−/− fibroblasts cultured in the absence or presence of TIMP-1 and TIMP-2. Results are expressed as the mean ± 1 SD of three experiments. (C) RT-PCR analysis of MT1-MMP and MT3-MMP expression in wild-type and null cells cultured atop fibrin gels and stimulated with PDGF for 72 h. The 402 bp MT-3 transcript was confirmed by sequence analysis. MT2-MMP-specific transcripts were not detected under these conditions.
Among the other membrane-anchored MMPs that are preferentially sensitive to TIMP-2, MT2-MMP, and MT3-MMP appear to be among the most widely expressed in vitro or in vivo (30, 42–47). To determine whether fibroblasts expressed either of these MT-MMPs, wild-type or MT1-MMP null cells were cultured atop fibrin gels, stimulated with PDGF for 3 d, and isolated RNA amplified by reverse transcription (RT)-PCR using the corresponding MT-MMP-specific primers (Fig. 3 C). Interestingly, while MT2-MMP could not be detected in either fibroblast population, both the wild-type and MT1-MMP-null fibroblasts expressed MT3-MMP. Consistent with these data, MT1-MMP-null fibroblasts retained a diminished ability to activate proMMP-2 via a process inhibitable by TIMP-2 or BB-94, but not TIMP-1 (data not shown).
To determine directly whether MT3-MMP can confer a fibrin-invasive activity comparable to that observed for MT1-MMP, an invasion-incompetent cell population (i.e., CHO-K1 cells) was transiently transfected with either of the epitope-tagged MT-MMPs and fibrin invasion monitored in the presence of 10% serum as described. While control transfected CHO-K1 cells showed little, if any, BB-94-sensitive invasive activity, MT1-MMP transfected cells readily penetrated the gels (Fig. 4, A and D) . Significantly, MT3-MMP-transient as well as stable transfectants displayed similar activity despite the fact that the levels of MT3-MMP protein expressed (as assessed by Western blot analysis with anti-M2 FLAG monoclonal antibody) were consistently lower than those detected for MT1-MMP (Fig. 4, A–C). Finally, in a fashion similar to that observed in the wild-type or MT1-MMP null fibroblasts, MT3-MMP–dependent invasion was completely resistant to TIMP-1, but equally sensitive to BB-94 or TIMP-2 (Fig. 4 D). Cells cotransfected with MT1-MMP and MT3-MMP did not display synergistic invasive behavior (data not shown). Hence, the fibrin-invasive properties of MT1-MMP can be extended to include the membrane-anchored metalloproteinase, MT3-MMP.
Figure 4.
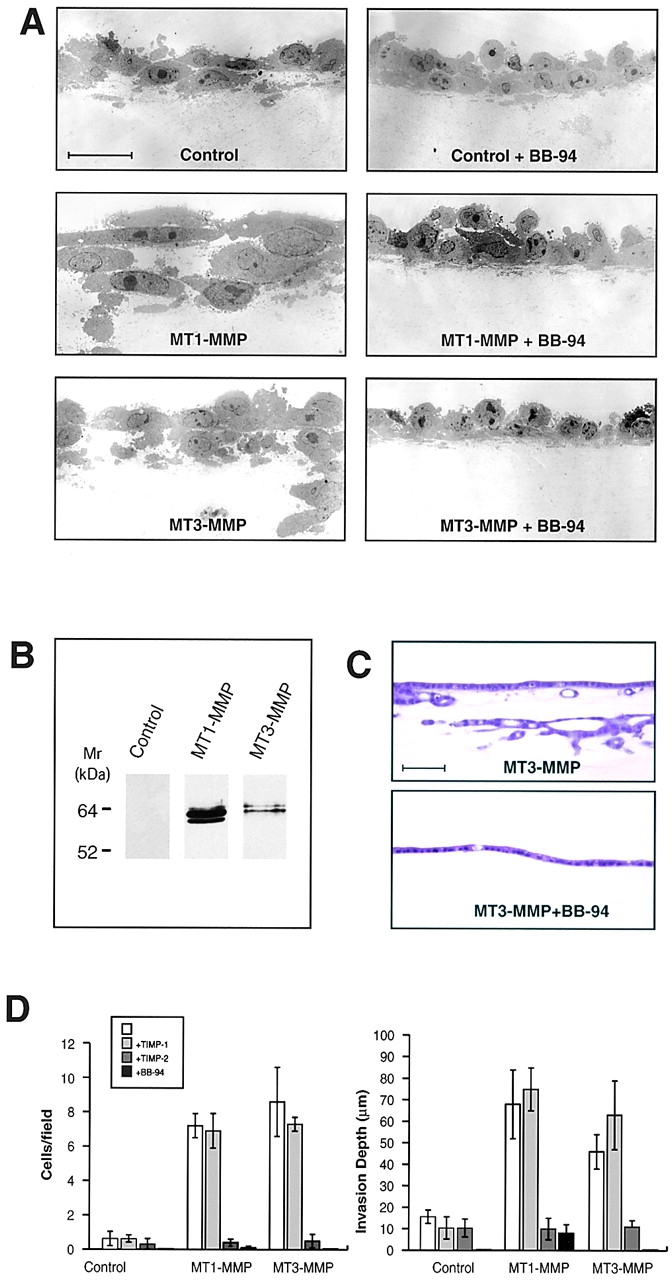
MT3-MMP-dependent fibrin-invasive activity. (A) TEM sections of control vector-, MT1-MMP-, and MT3-MMP–transfected CHO-K1 cells cultured atop fibrin gels in 10% FBS and stimulated with SF/HGF in the absence or presence of BB-94 for 5 d. Bar = 10 μm. (B) Western blot analysis of CHO-K1 cells transfected with a control vector, FLAG-tagged MT1-MMP, or FLAG-tagged MT3-MMP. (C) H and E–stained transverse sections of MDCK stable transfectants overexpressing MT3-MMP cultured atop fibrin gels and stimulated with SF/HGF for 5 d in the absence or presence of BB-94. Bar = 100 μm. (D) CHO-K1 cells transiently transfected with control-, MT1-MMP-, or MT3-MMP- expression vectors were cultured atop fibrin gels for 5 d and the number of invasive foci/field and invasion depth were quantified in the absence or presence of TIMP-1, TIMP-2, or BB-94. Results are expressed as the mean ± 1 SD of three experiments.
Fibrin-Invasive Activity Can Be Extended to MT2-MMP, but not MT4-MMP.
While fibroblasts can preferentially rely on either MT1-MMP or MT3-MMP to negotiate fibrin barriers, a variety of normal as well as neoplastic cell populations display different repertoires of membrane-anchored metalloproteinases which include MT2-MMP and/or MT4-MMP (25, 42–47). To determine whether fibrin-invasive activity is restricted to MT1- and MT3-MMP, cells were transfected with either MT2- or MT4-MMP and invasive activity monitored. Significantly, despite the fact that the isolated catalytic domain of MT2-MMP has been reported to express little, if any, fibrinolytic activity (23), transiently transfected CHO-K1 cells (as well as stably transfected cells) expressed an invasive phenotype comparable to that observed for MT1-MMP and MT3-MMP (Fig. 5, A–C) . Likewise, the invasion by MT2-MMP transfectants could be blocked completely by TIMP-2 or BB-94, but not TIMP-1 (Fig. 5, B, C, and E).
Figure 5.
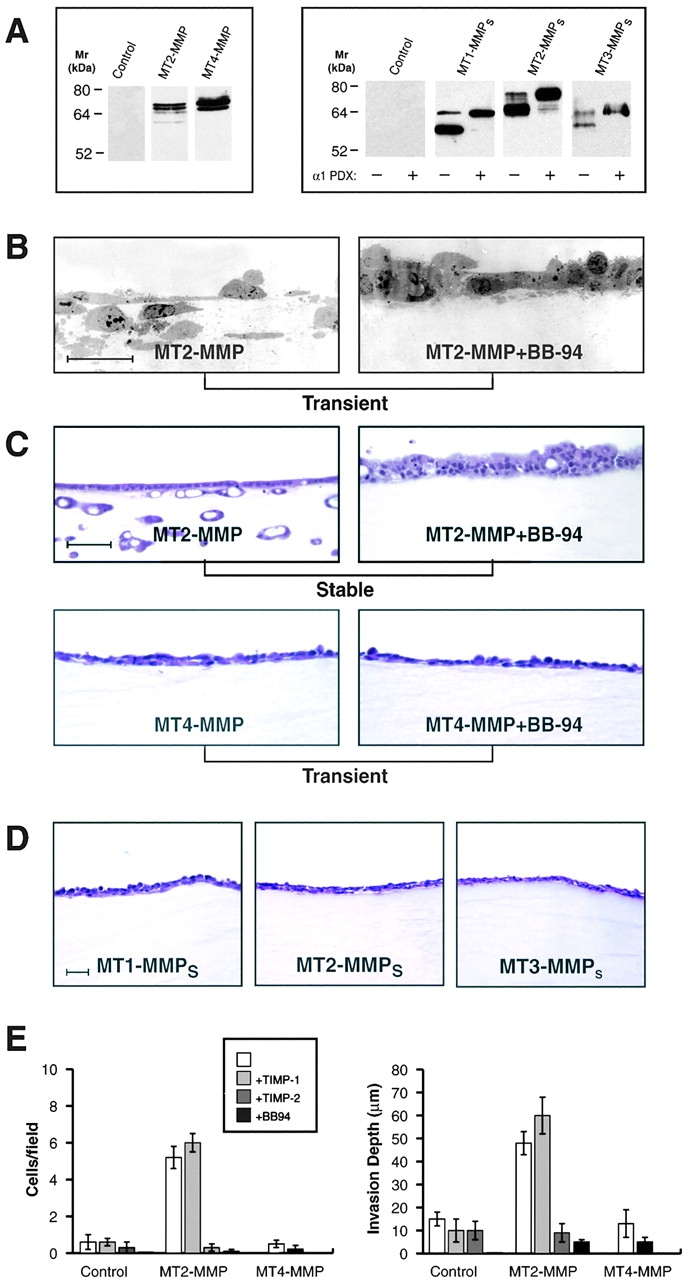
Fibrin-invasive potential of MT2-MMP, MT4-MMP, and transmembrane-deleted mutants. (A) Western blot analysis of CHO-K1 cells transiently transfected with a control vector, FLAG-tagged full-length MT2-MMP, or FLAG-tagged full-length MT4-MMP (left panel). Alternatively, CHO-K1 cells were transiently transfected with a control vector, transmembrane-deleted MT1-MMP (MT1-MMPs), MT2-MMPs, or MT3-MMPs containing a His-tag epitope at their carboxy termini in the absence or presence of an α1PDX expression vector. In the presence of α1PDX, the proform of each of the soluble MT-MMPs was the predominant species detected. (B) TEM sections of CHO-K1 cells transiently transfected with MT2-MMP cultured atop a fibrin gel in 10% FBS with SF/HGF in the absence or presence of BB-94. Bar = 10 μm. (C) H and E–stained sections of MDCK cells stably transfected with MT2-MMP or CHO-K1 cells transiently transfected with MT4-MMP and cultured atop fibrin gels as described above, in the absence or presence of BB-94. Bar = 100 μm. (D) CHO-K1 cells transiently transfected with MT1-MMPs, MT2-MMPs, or MT3-MMPs did not invade the underlying fibrin gel after a 5 d incubation period. Bar = 50 μm. (E) Invasive foci/field and invasion depth for SF/HGF-stimulated CHO-K1 cells transiently transfected with a control vector, MT2-MMP, or MT4-MMP and cultured atop fibrin gels in the absence or presence of TIMP-1, TIMP-2, or BB-94 was determined as described above. Results are expressed as the mean ± 1 SD of three experiments.
The ability of MT1-, 2-, and 3-MMP to promote fibrin invasion contrasts with the inability of each of the secreted MMPs to confer similar activities (see Fig. 1). Among the MMPs tested, only MMP-11 (stromelysin-3) and MT-MMP prodomains include a proprotein recognition motif that allows these proteinases to undergo efficient processing to their enzymically active forms (19, 32, 48). To determine whether MT-MMPs confer invasive activity when secreted as fully active, but no longer membrane tethered, enzymes, each of the proteinases were alternatively expressed as epitope-tagged, transmembrane-deleted mutants. As shown in Fig. 5 A (right panel), the soluble MT-MMPs were all expressed as a higher Mr proform and a lower Mr active enzyme whose activity was confirmed by fluorometric assay (see Materials and Methods). The identity of the slower migrating bands as the MT-MMP proforms was confirmed by cotransfecting cells with the proprotein convertase inhibitor, α1PDX32, which blocked soluble MT-MMP processing (Fig. 5 A). However, despite the fact that transfected cells secreted active MT-MMPs efficiently, no fibrin-invasive activity was detected (Fig. 5 D).
Given the apparent requirement that fibrinolytic MMPs must be membrane-anchored to drive invasion, the activity of MT4-MMP, a GPI-anchored metalloproteinase, was assessed. However, despite the fact that soluble MT4-MMP can degrade fibrin(ogen) (25), the GPI-anchored proteinase did not promote fibrin-invasive activity under any of the conditions tested (Fig. 5, A, C, and E). Although MT4-MMP, unlike MT1-, 2-, or 3-MMP cannot process gelatinase A to its active form (25), the pro-invasive phenotypes displayed by MT1-, MT2-, or MT3-MMP–transfected cells was not affected under either gelatinase A–depleted conditions or when cells were cotransfected with gelatinase A (data not shown).
MT-MMP–dependent Invasion.
In vivo_,_ networks of polymerized fibrin can vary dramatically in terms of fiber size, density, rigidity, and sensitivity to proteolytic degradation (35, 49–51). To determine the ability of MT-MMP–transfected cells to degrade and invade fibrin gel networks exhibiting different physical properties, fibrinogen was polymerized at pH 6.9–8.0 (Fig. 6) . At acidic pH, the polymerization process is slowed, favoring the formation of thicker, more rigid fibrillar networks while at alkaline pH, polymerization is accelerated generating thinner, more malleable structures (35, 49–51). Though thinner fibrils are more resistant to proteolytic attack by the plasminogen activator-plasminogen system (35, 51), stably transfected cells overexpressing MT1-MMP, MT2-MMP, or MT3-MMP rapidly spread on each of the matrices and degraded the underlying gel at comparable rates (Fig. 6, A and B). Likewise, no differences were detected in the ability of the transfectants to penetrate or invade the gels (Fig. 6, C and D). Fibrin fiber size and gel characteristics are also affected by the covalent incorporation of fibronectin into the fibrillar network by transglutaminase (35, 50). As MT1, 2, and 3-MMPs are able to degrade fibronectin (33, 52–54), we considered the possibility that the degradation of gel-incorporated fibronectin may have contributed to the invasive phenotype. However, stable transfectants cultured atop transglutaminase cross-linked fibrin gels formed from fibronectin-depleted fibrinogen exhibited similar degradative and invasive behavior (data not shown). Finally, while the ability of MT1-, 2-, and 3-MMP transfectants to degrade and/or invade fibrin barriers in comparable fashion suggests that each of these metalloproteinases share overlapping substrate profiles, their interchangeable phenotypic behavior was limited to the fibrin substratum (Fig. 7) . In paired experiments, when MT-MMP transfectants were cultured atop type I collagen gels, only MT1-MMP accelerated invasion while the MT2-MMP and MT3-MMP transfectants displayed a reduced invasive activity relative to the control transfected cells (Fig. 7). Hence, whereas a different repertoire of MT-MMP activities must be used for traversing the type I collagen-rich interstitium (30), each of the three MT-MMPs arm cells with the ability to negotiate fibrin barriers with equal efficiency.
Figure 6.

Effect of fibrin diameter on fibrinolytic and invasive activity of MT1-MMP-, MT2-MMP-, or MT3-MMP–expressing stable transfectants. (A) Fibrin gels prepared at pH 6.9, 7.4, or 8.0 form fibrils with mean diameters of 211 ± 60 nm, 149 ± 51 nm, and 81 ± 30 nm (mean ± 1 SD) as assessed by SEM (top three panels; Bar = 1 μm), and supported comparable cell adhesion (bottom three panels; Bar = 10 μm). B, C, and D depict the fibrinolytic activity, number of invasive foci and depth of invasion for control-, MT1-MMP-, MT2-MMP-, and MT3-MMP-stable MDCK transfectants stimulated with SF/HGF and cultured atop fibrin gels prepared at pH 6.9 (dark gray bars), 7.4 (white bars), or 8.9 (light gray bars) for 10 d. Results are expressed as the mean ± 1 SD of three experiments.
Figure 7.
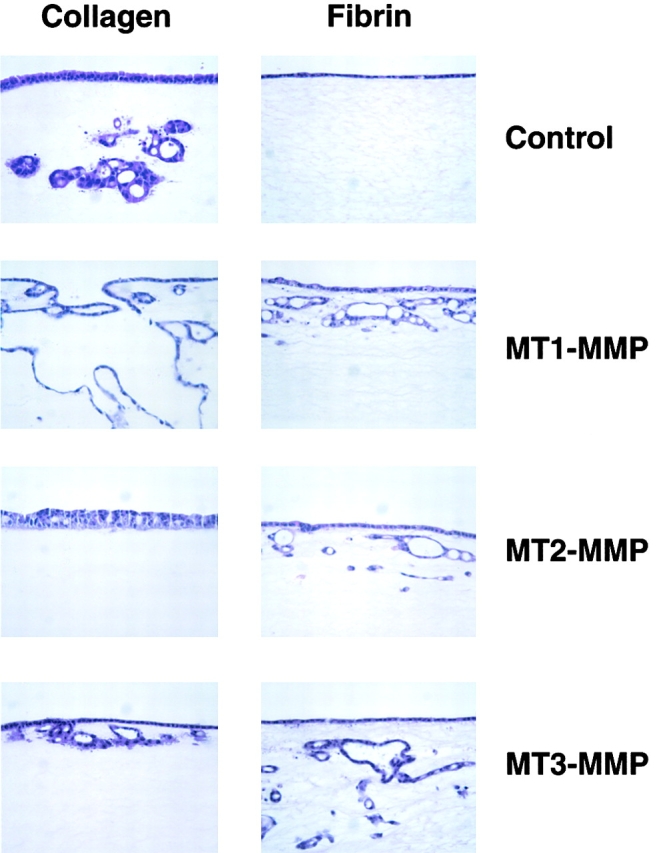
Differential regulation of type I collagen- and fibrin-invasive activity by MT-MMPs. H and E–stained cross-sections of the invasion pattern displayed by MDCK cell clones stably transfected with control-, MT1-MMP-, MT2-MMP-, or MT3-MMP-expression vectors and cultured atop type I collagen (left series of panels) or fibrin (right series of panels) in 10% FBS and stimulated with SF/HGF for 10 d. While control MDCK invaded type I collagen, no invasive activity occurred atop fibrin gels. Clones overexpressing MT1-MMP and cultured atop collagen gels underwent a disrupted tubulogenesis program and expressed heightened invasive activity. In the case of MT2-MMP– or MT3-MMP–overexpressing clones, collagen invasive activity was inhibited. These patterns of invasion contrasted with the comparable fibrin-invasive activity of the MT1-, MT2-, and MT3-MMP–overexpressing clones.
Discussion
The proteolytic processes used by normal or neoplastic cells to penetrate the fibrin deposits that surround wounds, inflammatory sites, or tumor foci have remained largely undefined (2–13). Given the efficiency with which plasmin degrades fibrin relative to other proteinases, it has been generally assumed that the plasminogen activator-plasminogen axis would serve as the sole determinant of invasive activity (5, 6, 11, 12). While plasmin can participate in fibrin-invasive events in vitro or in vivo (5, 6, 11, 12, 55), studies in plasminogen-deficient mice have demonstrated that alternate proteolytic systems exist that allow cells to readily infiltrate fibrin-rich tissues in pathophysiologic settings (e.g., references 9, 16–18, and 56). Coincident with these findings, increased awareness has focused on the fact that the substrate repertoire of MMPs can be extended from structural components of the “mature” extracellular matrix (i.e., collagens, elastins, glycoproteins, and proteoglycans) to fibrin(ogen), the major component of the provisional matrix (3, 22, 23, 25, 26). Nonetheless, fibrin(ogen)olytic activity need not correlate with the ability of a given proteinase to confer an invasive phenotype as pericellular degradation must proceed in an orchestrated fashion that allows the cell to tunnel through the substratum while retaining sufficient matrix to act as a structural support for movement (3, 10, 14, 15, 57).
Recently, we reported that invasion-incompetent cells can be induced to penetrate fibrin matrices after transfection with MT1-MMP cDNA (3). However, the role that the proteinase plays in regulating the fibrin-invasive activity of wild-type cells has remained unclear. Further, the relative importance of MT1-MMP in promoting fibrin invasion has been questioned as a large series of soluble MMPs have been subsequently reported to degrade fibrin(ogen) with similar, if not greater, efficiency (3, 22–27). To order the fibrin(ogen)olytic MMPs in terms of their pro-invasive potential, invasion-incompetent cells were transiently or stably transfected with fibrin(ogen)olytic metalloproteinases. Significantly, whereas MT1-MMP overexpressing cells readily invaded cross-linked fibrin gels, neither collagenases (MMP-1 or MMP-13), gelatinases (MMP-2 or MMP-9), stromelysins (MMP-3 or MMP-11), nor matrilysin conferred invasive activity.
Given that fibroblasts migrating into fibrin-rich tissues in vivo express MT1-MMP (16–18, 36), the ability of fibroblasts isolated from wild-type or MT1-MMP–deleted mice to penetrate fibrin gels was examined. While the mechanisms used by wild-type fibroblasts has remained controversial (e.g., see references 4, 5, and 12), PDGF-stimulated fibroblasts invaded fibrin gels via an MMP-dependent process that proved sensitive to BB-94 or TIMP-2, but not TIMP-1. As BB-94 and TIMP-2 effectively inhibit all MMP family members while TIMP-1–dependent inhibition is predominately restricted to secreted MMPs (3, 19, 25, 30), this profile supported a potential role for MT1-MMP in fibrin invasion. Interestingly, secreted as well as membrane-anchored MMPs have recently been implicated in migratory responses of cells cultured atop various two-dimensional substrata including type I collagen and laminin-5 (37–41). The promigratory response has been alternatively linked with degradation of the underlying substratum, integrin processing, or proteolysis of surface-associated transglutaminase (37–41, 58). However, in our system, the inhibitory effects of BB-94 and TIMP-2 were confined solely to effects on invasive behavior. Caution should, however, be exercised in necessarily ascribing all effects of these inhibitors to their ability to target MMPs. For example, BB-94 and other peptidomimetic inhibitors can affect cell responses by inhibiting metalloproteinases in the adamalysin family responsible for the proteolytic shedding of membrane-anchored ligands that can trigger growth factor receptors (59–61). In contrast, TIMP-2 has been reported to suppress mitogenesis independently of its antiproteolytic activity (62). Nonetheless, despite these potential caveats, neither fibroblast adhesion, proliferation, nor migration were affected by BB-94 or TIMP-2 under our experimental conditions. Though we had previously reported that MMP inhibitors did not completely block fibroblast/myofibroblast migration from fibrin-embedded tissue explants (3), subsequent studies have demonstrated that cell egress in the presence of BB-94 or TIMP-2 is significantly impaired and confined to a narrow zone surrounding the tissue (unpublished observation).
Given that fibroblasts use an MMP-dependent process to invade fibrin gels and express MT1-MMP in vivo (18, 36), we predicted that MT1-MMP–null fibroblasts would fail to penetrate fibrin matrices. Indeed, during the first 2 d of the culture period, the MT1-MMP–deleted cells expressed limited invasive behavior despite normal adhesive and migratory responses atop fibrin gels. Surprisingly, however, the activity of null cells rebounded and reached levels comparable to those observed in the wild-type cells. While the compensatory activity of the knockout cells to express an invasive phenotype might have reflected the use of an alternate class of proteinases, the residual activity remained sensitive to BB-94 and TIMP-2, and hence, was consistent with the participation of an alternate MT-MMP. As predicted, MT1-MMP+/+ as well as MT1-MMP-null fibroblasts expressed MT3-MMP in a fashion similar to that reported for wild-type smooth muscle cells stimulated with PDGF (47). Though neither MT3-MMP specific inhibitors nor gene-deleted animals have been described, we confirmed the ability of MT3-MMP to directly confer fibrin-invasive activity. We are currently unable to compare MT1-MMP and MT3-MMP expression at the protein level in wild-type or null fibroblasts, as mouse-specific antibodies are unavailable. However, using epitope-tagged constructs, both MT-MMPs displayed comparable activity when expressed at similar levels in transiently transfected cells. Taken together, these data support our contention that either MT1-MMP or MT3-MMP can drive the fibrin-invasive phenotype in PDGF-stimulated fibroblasts. Consistent with this interpretation, MT1-MMP null cells were unable to traverse a type I collagen matrix as MT3-MMP expresses minimal activity toward this substrate (30; unpublished observation). While other TIMP-2–sensitive MMPs might further modify fibrin-invasive activity in fibroblasts, neither MT5-MMP, MT6-MMP, nor MMP-19 (a secreted MMP recently shown to be preferentially sensitive to inhibition by TIMP-2 [63]) were able to promote fibrin-invasive activity under our assay conditions (unpublished observation).
Fibroblasts can alternatively use MT1-MMP or MT3-MMP to invade fibrin-rich deposits, but other cell populations (e.g., synoviocytes, leukocytes, trophoblasts, and tumor cells) express distinct repertoires of membrane-anchored metalloproteinases which include MT2-MMP and MT4-MMP (19, 25, 42–47). Whereas the isolated catalytic domain of MT2-MMP has been reported to be devoid of fibrinolytic activity (23), a similar construct of MT4-MMP was recently shown to degrade fibrinogen as well as fibrin (25). In contrast to these reports, full-length MT2-MMP, but not MT4-MMP, efficiently conferred fibrin-invasive potential. These findings highlight the importance of using the intact enzymes rather than isolated domains for functional characterization and are consistent with reports documenting marked differences in activity between catalytic domains, transmembrane-deleted constructs and the intact MMPs (33, 53, 64). Likewise, our studies with MT4-MMP-transfected cells indicate that the fibrin(ogen)olytic activity of an isolated enzyme may not predict its ability to mediate invasive activity. Unlike MT1, 2, or 3-MMP, MT4-MMP is GPI-anchored, its catalytic domain does not include a characteristic insert (i.e., the so-called IS-2 domain), and the proteinase cannot activate progelatinase A (25, 65). However, these properties do not likely underlie its inability to confer invasive activity as MT1-MMP retains fibrin-invasive activity when GPI-anchored or its IS-2 domain is deleted (unpublished observation). Further, gelatinase A does not express significant fibrinolytic activity nor does its exclusion affect fibrin-invasive activity (3, 23). Instead, the inability of MT4-MMP to drive invasive activity may reside in the fact that the proteinase cannot, in contrast to MT1-MMP, efficiently degrade the high molecular weight α-polymers of fibrin (3, 25). We speculate that the specific cleavage sites chosen in the fibrin scaffolding may be important in allowing the cell to penetrate the matrix while maintaining the structural stability of the supporting network.
As MT1-MMP, MT2-MMP, or MT3-MMP are widely distributed among normal and neoplastic cells (e.g., references 19, 25, and 42–47), these metalloproteinases likely constitute the plasminogen-independent arm of the fibrin-invasive phenotype observed in vivo. However, unlike the plasminogen activator-plasminogen axis wherein fibrinolytic activity is affected by fibrin structure or composition (49–51), MT-MMP–dependent proteolysis and invasion proceeded in an autonomous fashion. The lack of a plasminogen-dependent component in this invasion program is also consistent with the proMT-MMP activation process. While plasminogen mediates the activation of a number of secreted MMPs (19), each of the MT-MMPs contains a basic motif (i.e., RXK/RR) at the COOH terminus of their respective prodomains that serves as a recognition sequence for one or more members of the proprotein convertase family of serine proteinases (19, 32, 48). The convertases activate the MT-MMP zymogens either intracellularly in the trans-Golgi network or at the cell surface, thus allowing the cell to directly control its proteolytic potential in a trans-dependent fashion (19, 32, 48). Interestingly, though the transmembrane-deleted MT-MMP mutants are also processed efficiently by the proprotein convertases, only the membrane-anchored forms of the proteinases expressed functional activity in our invasion assay system. Given that soluble MT1-MMP expresses fibrinolytic activity (3, 23, 26), these results lend credence to the notion that proteinases must be tethered to the cell surface to efficiently match degradative activity with motility (14, 15, 30). MT-MMPs may further require localization to specific regions of the membrane actively involved in the invasive process (66, 67), but cytosolic domain-deleted forms of MT1-MMP remain functional (3, 30). Either surface localization signals are no longer required when the proteinase is overexpressed or targeting information is encoded within the extracellular domain itself.
Our findings have focused on the fibrin-invasive activity of MT-MMPs, but increasing evidence suggests that multiple metalloproteinases may participate in the bulk resorption of fibrin deposits and affect coagulation by targeting fibrinogen as well as Factor XII (22–27). Significantly, fibrin deposits were increased at wound sites in normal or plasminogen-deleted mice that were treated with a broad-spectrum MMP inhibitor (18). Coincident with these effects, the delayed wound healing response of plasminogen-null mice was further affected by the metalloproteinase inhibitor, but this latter result should be interpreted cautiously as effects on adamalysin-dependent shedding of surface-associated growth factors (e.g., HB-EGF) must be considered (68). This caveat aside, fibrin-degradation products consistent with MMP-dependent proteolysis have been detected in rheumatoid synovial fluid and MMP-9 has recently been implicated in fibrinolytic events in an in vivo nephritis model (22, 23, 27). In the latter studies, renal damage was exacerbated in MMP-9 null mice and linked to increased fibrin deposition (27). Further, while MMP-9 has been reported to degrade fibrin rather inefficiently relative to other secreted MMPs (e.g., MMP-3 or MMP-7; references 22 and 23), Lelongt et al. demonstrated that the proteinase could efficiently degrade fibrin under their in vitro conditions (27). In vivo, however, the situation is more complex as ECM components such as type I collagen are deposited into fibrin networks by fibroblasts (1). As these ECM-fibrin composites display a heightened resistance to plasmin-dependent fibrinolysis (69), secreted MMPs may play an important role in fibrin clearance by either attacking fibrin(ogen) directly or by collaborating with plasmin to dissolve the stabilized fibrin deposits. Given evidence that MT-MMPs maybe shed from the cell surface or expressed directly as soluble enzymes (44, 70), this class of enzymes may either promote invasion or function as bulk fibrinolysins dependent on their presentation at the cell surface.
In summary, we have identified a subset of MT-MMPs as the lead candidates responsible for allowing mammalian cells to traverse fibrin-rich tissues in a plasminogen-independent fashion. We note, however, that the comparable activities of MT1-MMP, MT2-MMP, and MT3-MMP with regard to fibrin-invasive activity do not similarly extend to other matrix barriers such as type I collagen. Nonetheless, despite only ∼50% homology between MT1-MMP and MT2-MMP or MT3-MMP at the amino acid level (19), each of the membrane-anchored metalloproteinases allows expressing cells to display comparable fibrin-invasive and fibrinolytic activity. The ability of MT2-MMP and MT3-MMP, but not other fibrinolytic MMPs, to confer fibrin-invasive activity in the absence or presence of MT1-MMP, identifies a new pro-invasive system for traversing the provisional matrix under pathophysiologic conditions.
Acknowledgments
This work was supported by National Institutes of Health grants CA71699 and CA88308.
K.B. Hotary, I. Yana, and F. Sabeh contributed equally to this work.
Footnotes
*
Abbreviations used in this paper: H and E, hematoxylin and eosin; MMP, matrix metalloproteinase; PDGF, platelet-derived growth factor; RT, reverse transcription; SF/HFG, scatter factor/hepatocyte growth factor; TIMP, tissue inhibitor of metalloproteases.
References
- 1.Singer, A.J., and R.A.F. Clark. 1999. Cutaneous wound healing. N. Engl. J. Med. 341:738–746. [DOI] [PubMed] [Google Scholar]
- 2.Dvorak, H.F., L.F. Brown, M. Detmar, and A.M. Dvorak. 1995. Vascular permeability factor, vascular endothelial growth factor, microvascular hyperpermeability, and angiogenesis. Am. J. Pathol. 146:1029–1039. [PMC free article] [PubMed] [Google Scholar]
- 3.Hiraoka, N., E. Allen, I.J. Apel, M.R. Gyetko, and S.J. Weiss. 1998. Matrix metalloproteinases regulate neovascularization by acting as pericellular fibrinolysins. Cell. 95:365–377. [DOI] [PubMed] [Google Scholar]
- 4.Brown, L.F., N. Lanir, J. McDonagh, K. Tognazzi, A.M. Dvorak, and H.F. Dvorak. 1993. Fibroblast migration in fibrin gel matrices. Am. J. Pathol. 142:273–283. [PMC free article] [PubMed] [Google Scholar]
- 5.Greiling, D., and R.A.F. Clark. 1997. Fibronectin provides a conduit for fibroblast transmigration from collagenous stroma into fibrin clot provisional matrix. J. Cell Sci. 110:861–870. [DOI] [PubMed] [Google Scholar]
- 6.Ronfard, V., and Y. Barrandon. 2001. Migration of keratinocytes through tunnels of digested fibrin. Proc. Natl. Acad. Sci. USA. 98:4504–4509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mach, F., U. Schonbeck, R.P. Fabunmi, C. Murphy. E. Atkinson, J.-Y. Bonnefoy, P. Graber, and P. Libby. 1999. T lymphocytes induce endothelial cell matrix metalloproteinase expression by a CD40L-dependent mechanism. Am. J. Pathol. 154:229–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lanir, N., P.S. Ciano, L. Van De Water, J. McDonagh, A.M. Dvorak, and H.F. Dvorak. 1988. Macrophage migration in fibrin gel matrices. J. Immunol. 140:2340–2349. [PubMed] [Google Scholar]
- 9.Bugge, T.H., K.W. Kombrinck, Q. Xiao, K. Holmback, C.C. Daugherty, D.P. Witte, and J.L. Degen. 1997. Growth and dissemination of Lewis lung carcinoma in plasminogen-deficient mice. Blood. 90:4522–4531. [PubMed] [Google Scholar]
- 10.Montesano, R., M.S. Pepper, J.-D. Vassalli, and L. Orci. 1987. Phorbol ester induces cultured endothelial cells to invade a fibrin matrix in the presence of fibrinolytic inhibitors. J. Cell. Physiol. 132:509–516. [DOI] [PubMed] [Google Scholar]
- 11.Kookwijk. P., M.G.M. van Erck, W.J.A. de Vree, M.A. Vermeer, H.A. Weich, R. Hanemaaijer, and V.W.M. van Hinsbergh. 1996. Cooperative effect of TNFα, bFGF, and VEGF on the formation of tubular structures of human microvascular endothelial cells in a fibrin matrix. Role of urokinase activity. J. Cell Biol. 132:1177–1188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Knox, P., S. Crooks, M.C. Scaife, and S. Patel. 1987. Role of plasminogen, plasmin, and plasminogen activators in the migration of fibroblasts into plasma clots. J. Cell. Physiol. 132:501–508. [DOI] [PubMed] [Google Scholar]
- 13.Dvorak, H.F., V.S. Harvey, P. Estrella, L.F. Brown, J. McDonagh, and A.M. Dvorak. 1987. Fibrin containing gels induce angiogenesis. Implications for tumor stroma generation and wound healing. Lab. Invest. 57:673–686. [PubMed] [Google Scholar]
- 14.Murphy, G., and J. Gavrilovic. 1999. Proteolysis and cell migration: creating a path? Curr. Opin. Cell Biol. 11:614–621. [DOI] [PubMed] [Google Scholar]
- 15.Quaranta, V. 2000. Cell migration through extracellular matrix: membrane-type metalloproteinases make the way. J. Cell Biol. 149:1167–1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Romer, J., T.H. Bugge, C. Pyke, L.R. Lund, M.J. Flick, J.L. Degen, and K. Dano. 1996. Impaired wound healing in mice with a disrupted plasminogen gene. Nat. Med. 2:287–292. [DOI] [PubMed] [Google Scholar]
- 17.Bugge, T.H., M.J. Flick, M.J.S. Danton, C.C. Daugherty, J. Romer, K. Dano, P. Carmeliet, D. Collen, and J.L. Degen. 1996. Urokinase-type plasminogen activator is effective in fibrin clearance in the absence of its receptor or tissue-type plasminogen activator. Proc. Natl. Acad. Sci. USA. 93:5899–5904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lund, L.R., J. Romer, T.H. Bugge, R.S. Nielsen, T.L. Frandsen, J.L. Degen, R.W. Stephens, and K. Dano. 1999. Functional overlap between two classes of matrix-degrading proteases in wound healing. EMBO J. 18:4645–4656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nagase, H., and J.F. Woessner, Jr. 1999. Matrix metalloproteinases. J. Biol. Chem. 274:21491–21494. [DOI] [PubMed] [Google Scholar]
- 20.Marchenko, G.N., and A.Y. Strongin. 2001. MMP-28 a new human matrix metalloproteinase with an unusual cysteine-switch sequence is widely expressed in tumors. Gene. 265:87–93. [DOI] [PubMed] [Google Scholar]
- 21.Lohi, J., C.L. Wilson, J.D. Roby, and W.C. Parks. 2001. Epilysin, a novel human matrix metalloproteinase (MMP-28) expressed in testis and keratinocytes and in response to injury. J. Biol. Chem. 276:10134–10144. [DOI] [PubMed] [Google Scholar]
- 22.Bini, A., Y. Itoh, B.J. Kudryk, and H. Nagase. 1996. Degradation of cross-linked fibrin by matrix metalloproteinase 3 (stromelysin 1): Hydrolysis of the γ Gly 404-Ala 405 peptide bond. Biochemistry. 35:13056–13063. [DOI] [PubMed] [Google Scholar]
- 23.Bini, A., D. Wu, J. Schnuer, and B.J. Kudryk. 1999. Characterization of stromelysin 1 (MMP-3), matrilysin (MMP-7), and membrane type 1 matrix metalloproteinase (MT1-MMP) derived fibrin(ogen) fragments D-dimer and D-like monomer: NH2-terminal sequences of late-stage digest fragments. Biochemistry. 38:13928–13936. [DOI] [PubMed] [Google Scholar]
- 24.Lijnen, H.R., F. Ugwu, M.-C. Rio, and D. Collen. 1998. Plasminogen/plasmin and matrix metalloproteinase system function in mice with targeted inactivation of stromelysin-3 (MMP-11). Fibrinolysis Proteol. 12:155–164. [Google Scholar]
- 25.English, W.R., X.S. Puente, J.M.P. Freije, V. Knauper, A. Amour, A. Merryweather, C Lopez-Otin, and G. Murphy. 2000. Membrane type 4 matrix metalloproteinase (MMP17) has tumor necrosis factor-α convertase activity but does not activate pro-MMP2. J. Biol. Chem. 275:14046–14055. [DOI] [PubMed] [Google Scholar]
- 26.Hiller, O., A. Lichte, A. Oberpichler, A. Kocourek, and H. Tschesche. 2000. Matrix metalloproteinases collagenase-2, macrophage elastase, collagenase-3 and membrane type 1-matrix metalloproteinase impair clotting by degradation of fibrinogen and factor XII. J. Biol. Chem. 275:33008–33013. [DOI] [PubMed] [Google Scholar]
- 27.Lelongt, B., S. Bengatta, M. Delauche, L.R. Lund, Z. Werb, and P.M. Ronco. 2001. Matrix metalloproteinase 9 protects mice from anti-glomerular basement membrane nephritis through its fibrinolytic activity. J. Exp. Med. 193:793–802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Holmbeck, K., P. Bianco, J. Caterina, S. Yamada, M. Kromer, S.A. Kuznetsov, M. Mankani, P.G. Robey, A.R. Poole, I. Pidoux, et al. 1999. MT1-MMP-deficient mice develop dwarfism, osteopenia, arthritis, and connective tissue disease due to inadequate collagen turnover. Cell. 99:81–92. [DOI] [PubMed] [Google Scholar]
- 29.Zhou, Z., S.S. Apte, R. Soininen, R. Cao, G.Y. Baaklini, R.W. Rauser, J. Wang, Y Cao, and K. Tryggvason. 2000. Impaired endochondral ossification and angiogenesis in mice deficient in membrane-type matrix metalloproteinase 1. Proc. Natl. Acad. Sci. USA. 97:4052–4057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hotary, K., E. Allen, A. Punturieri, I. Yana, and S.J. Weiss. 2000. Regulation of cell invasion and morphogenesis in a 3-dimensional type I collagen matrix by membrane-type matrix metalloproteinases 1, 2 and 3. J. Cell Biol. 149:1309–1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Havemose-Poulsen, A., P. Holmstrup, K. Stoltze, and H. Birkedal-Hansen. 1998. Dissolution of type I collagen fibrils by gingival fibroblasts isolated from patients of various periodontitis categories. J. Periodontal Res. 33:280–291. [DOI] [PubMed] [Google Scholar]
- 32.Yana, I., and S.J. Weiss. 2000. Regulation of membrane type-1 matrix metalloproteinase activation by proprotein convertases. Mol. Biol. Cell. 11:2387–2401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.D'Ortho, M.-P., H. Will, S. Atkinson, G. Butler, A. Messent, J. Gavrilovic, B. Smith, R. Timpl, L. Zardi, and G. Murphy. 1997. Membrane-type matrix metalloproteinases 1 and 2 exhibit broad-spectrum proteolytic capacities comparable to many matrix metalloproteinases. Eur. J. Biochem. 250:751–757. [DOI] [PubMed] [Google Scholar]
- 34.Corbett, S.A., C.L. Wilson, and J.E. Schwarzbauer. 1996. Changes in cell spreading and cytoskeletal organization are induced by adhesion to a fibronectin-fibrin matrix. Blood. 88:158–166. [PubMed] [Google Scholar]
- 35.Nehls, V., and R. Herrmann. 1996. The configuration of fibrin clots determines capillary morphogenesis and endothelial cell migration. Microvasc. Res. 51:347–364. [DOI] [PubMed] [Google Scholar]
- 36.Okada, A., C. Tomasetto, Y. Lutz, J.-P. Belocq, M.-C. Rio, and P. Basset. 1997. Expression of matrix metalloproteinases during rat skin wound healing: Evidence that membrane type-1 matrix metalloproteinase is a stromal activator of pro-gelatinase A. J. Cell Biol. 137:67–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pilcher, B.K., J.A. Dumin, B.D. Sudbeck, S.M. Krane, H.G. Welgus, and W.C. Parks. 1997. The activity of collagenase-1 is required for keratinocyte migration on a type I collagen matrix. J. Cell Biol. 137:1445–1457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Koshikawa, N., G. Giannelli, V. Cirulli, K. Miyazaki, and V. Quaranta. 2000. Role of cell surface metalloprotease MT1-MMP in epithelial cell migration over laminin-5. J. Cell Biol. 148:615–624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Belkin, A.M., S.S. Akimov, L.S. Zaritskaya, B.I. Ratnikov, E.I. Deryugina, and A.Y. Strongin. 2001. Matrix-dependent proteolysis of surface transglutaminase by membrane-type metalloproteinase regulates cancer cell adhesion and locomotion. J. Biol. Chem. 276:18415–18422. [DOI] [PubMed] [Google Scholar]
- 40.Behrendtsen, O., and Z Werb. 1997. Metalloproteinases regulate parietal endoderm differentiating and migrating in cultured mouse embryos. Dev. Dyn. 208:255-265. [DOI] [PubMed] [Google Scholar]
- 41.Makela, M., H. Larjava, E. Pirila, P. Maisi, T. Salo, T. Sorsa, and V.-J. Uitto. 1999. Matrix metalloproteinase 2 (gelatinase A) is related to migration of keratinocytes. Exp. Cell Res. 251:67–78. [DOI] [PubMed] [Google Scholar]
- 42.Kim, M.H., R.P. Kitson, P. Albertsson, U. Nannmark, P.H. Basse, P.J.K. Kuppen, M.E. Hodland, and R.H. Goldfarb. 2000. Secreted and membrane-associated matrix metalloproteinases of IL-2-activated NK cells and their inhibitors. J. Immunol. 164:5883–5889. [DOI] [PubMed] [Google Scholar]
- 43.Yamanaka, H., K. Makino, M. Takizawa, H. Nakamura, N. Fujimoto, H. Moriya, R. Nemori, H. Sato, M. Seiki, and Y. Okada. 2000. Expression and tissue localization of membrane-types 1, 2, and 3 matrix metalloproteinases in rheumatoid synovium. Lab. Invest. 80:677–687. [DOI] [PubMed] [Google Scholar]
- 44.Shofuda, K.-I., H. Yasumitsu, A. Nishihashi, K. Miki, and K. Miyazaki. 1997. Expression of three membrane-type matrix metalloproteinases (MT-MMPs) in rat vascular smooth muscle cells and characterization of MT3-MMPs with and without transmembrane domain. J. Biol. Chem. 272:9749–9754. [DOI] [PubMed] [Google Scholar]
- 45.Fjorn, S.F., N. Hastrup, J.F. Larsen, L.R. Lund, and C. Pyke. 2000. Messenger RNA for membrane-type 2 matrix metalloproteinase, MT2-MMP, is expressed in human placenta of first trimester. Placenta. 21:170–176. [DOI] [PubMed] [Google Scholar]
- 46.Nakada, M., H. Nakamura, E. Ikeda, N. Fujimoto, J. Yamashita, H. Sato, M. Seiki, and Y. Okada. 1999. Expression and tissue localization of membrane-type 1, 2, and 3 matrix metalloproteinases in human astrocytic tumors. Am. J. Pathol. 154:417–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Shofuda, K.-I., Y. Nagashima, K. Kawahara, H. Yasumitsu, K. Miki, and K. Miyazaki. 1998. Elevated expression of membrane-type 1 and 3 matrix metalloproteinases in rat vascular smooth muscle cells activated by arterial injury. Lab. Invest. 78:915–923. [PubMed] [Google Scholar]
- 48.Pei, D., and S.J. Weiss. 1995. Furin-dependent intracellular activation of the human stromelysin-3 zymogen. Nature. 375:244–247. [DOI] [PubMed] [Google Scholar]
- 49.Carr, M.E., Jr., and B.M. Alving. 1995. Effect of fibrin structure on plasmin-mediated dissolution of plasma clots. Blood Coagul. Fibrinolysis. 6:567–573. [DOI] [PubMed] [Google Scholar]
- 50.Carr, M.E., Jr., D.A. Gabriel, and J. McDonagh. 1987. Influence of factor XIII and fibronectin on fiber size and density in thrombin-induced fibrin gels. J. Lab. Clin. Med. 110:747–752. [PubMed] [Google Scholar]
- 51.Collet, J.P., D. Park, C. Lesty, J. Soria, C. Soria, G. Montalescot, and J.W. Weisel. 2000. Influence of fibrin network conformation and fibrin fiber diameter on fibrinolysis speed. Arterioscler. Thromb. Vasc. Biol. 20:1354–1361. [DOI] [PubMed] [Google Scholar]
- 52.Pei, D., and S.J. Weiss. 1996. Transmembrane-deletion mutants of the membrane-type matrix metalloproteinase-1 activate progelatinase A and express intrinsic matrix-degrading activity. J. Biol. Chem. 271:9135–9140. [DOI] [PubMed] [Google Scholar]
- 53.Ohuchi, E., K. Imai, Y. Fujii, H. Sato, M. Seiki, and Y. Okada. 1997. Membrane type 1 matrix metalloproteinase digests interstitial collagens and other extracellular matrix macromolecules. J. Biol. Chem. 272:2446–2451. [DOI] [PubMed] [Google Scholar]
- 54.Shimada, T., H. Nakamura, E. Ohuchi, Y. Fujii, Y. Murakami, H. Sato, M. Seiki, and Y. Okada. 1999. Characterization of a truncated recombinant form of human membrane type 3 matrix metalloproteinase. Eur. J. Biochem. 262:907–914. [DOI] [PubMed] [Google Scholar]
- 55.Creemers, E., J. Cleutjens, J. Smits, S. Heymans, L. Moons, D. Collen, M. Daemen, and P. Carmeliet. 2000. Disruption of the plasminogen gene in mice abolishes wound healing after myocardial infarction. Am. J. Pathol. 156:1865–1873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Shi, C., A. Patel, D. Zhang, H. Wang, P. Carmeliet, G.L. Reed, M.-E. Lee, E. Haber, and N.E.S. Sibinga. 1999. Plasminogen is not required for neointima formation in a mouse model of vein graft stenosis. Circ. Res. 84:883–890. [DOI] [PubMed] [Google Scholar]
- 57.Bajou, K., V. Masson, R.D. Gerald, P.M. Schmitt, V. Albert, M. Praus, L.R. Lund, T.L. Frandsen, N. Brunner, K. Dano, et al. 2001. The plasminogen activator inhibitor PAI-1 controls in vivo tumor vascularization by interaction with proteases, not vitronectin: Implications for antiangiogenic strategies. J. Cell Biol. 152:77–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Deryugina, E.I., M.A. Bourdon, K. Jungwirth, J.W. Smith, and A.Y. Strongin. 2000. Functional activation of integrin αVβ3 in tumor cells expressing membrane-type 1 matrix metalloproteinase. Int. J. Cancer. 86:15–23. [DOI] [PubMed] [Google Scholar]
- 59.Dong, J., L.K. Opresko, P.J. Dempsey, D.A. Lauffenburger, R.J. Coffey, and H.S. Wiley. 1999. Metalloprotease-mediated ligand release regulates autocrine signaling through the epidermal growth factor receptor. Proc. Natl. Acad. Sci. USA. 96:6235–6240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Eguchi, S., P.J. Dempsey, G.D. Frank, E.D. Motley, and T. Inagami. 2001. Activation of MAPKs by angiotensin II in vascular smooth muscle cells. Metalloprotease-dependent EGF receptor activation is required for activation of ERK and p38 MAPK but not for JNK. J. Biol. Chem. 276:7957–7962. [DOI] [PubMed] [Google Scholar]
- 61.Nath, D., N.J. Williamson, R. Jarvis, and G. Murphy. 2001. Shedding of c-Met is regulated by crosstalk between a G-protein coupled receptor and the EGF receptor and is mediated by a TIMP-3 sensitive metalloproteinase. J. Cell. Sci. 114:1313–1220. [DOI] [PubMed] [Google Scholar]
- 62.Hoegy, S.E., H.-R. Oh, M.L. Corcoran, and W.G. Stetler-Stevenson. 2001. Tissue inhibitor of metalloproteinases-2 (TIMP-2) suppresses TKR-growth factor signaling independent of metalloproteinase inhibition. J. Biol. Chem. 276:3203–3214. [DOI] [PubMed] [Google Scholar]
- 63.Stracke, J.O., M. Hutton, M. Stewart, A.M. Pendas, B. Smith, C. Lopez-Otin, G. Murphy, and V. Knauper. 2000. Biochemical characterization of the catalytic domain of human matrix metalloproteinase 19. J. Biol. Chem. 275:14809–14816. [DOI] [PubMed] [Google Scholar]
- 64.Miyamori, H., T. Takino, M. Seiki, and H. Sato. 2000. Human membrane type-2 matrix metalloproteinase is defective in cell-associated activation of progelatinase A. Biochem. Biophys. Res. Commun. 267:796–800. [DOI] [PubMed] [Google Scholar]
- 65.Itoh, Y., M. Kajita, H. Kinoh, H. Mori, A. Okada, and M. Seiki. 1999. Membrane type 4 matrix metalloproteinase (MT4-MMP, MMP-17) is a glycosylphosphatidylinositol-anchored proteinase. J. Biol. Chem. 274:34260–34266. [DOI] [PubMed] [Google Scholar]
- 66.Nakahara, H., L. Howard, E.W. Thompson, H. Sato, M. Seiki, Y. Yeh, and W.-T. Chen. 1997. Transmembrane/cytoplasmic domain-mediated membrane type 1-matrix metalloprotease docking to invadopodia is required for cell invasion. Proc. Natl. Acad. Sci. USA. 94:7959–7964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lehti, K., H. Valtanen, S. Wickstrom, J. Lohi, and J. Keski-Oja. 2000. Regulation of membrane-type 1 matrix metalloproteinase activity by its cytoplasmic domain. J. Biol. Chem. 275:15006–15013. [DOI] [PubMed] [Google Scholar]
- 68.Tokumaru, S., S. Higashiyama, T. Endo, T. Nakagawa, J.-I., Miyagawa, K. Yamamori, Y. Hanakawa, H. Ohmoto, K. Yoshino, Y. Shirakata, et al. 2000. Ectodomain shedding of epidermal growth factor receptor ligands is required for keratinocyte migration in cutaneous wound healing. J. Cell Biol. 151:209–219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Siebenlist, K.R., and M.W. Mosessons. 1994. Progressive cross-linking of fibrin γ chains increases resistance to fibrinolysis. J. Biol. Chem. 269:28414–28419. [PubMed] [Google Scholar]
- 70.Imai, K., E. Ohuchi, T. Aoki, H. Nomura, Y. Fujii, H. Sato, M. Seiki, and Y. Okada. 1996. Membrane-type matrix metalloproteinase 1 is a gelatinolytic enzyme and is secreted in a complex with tissue inhibitor of metalloproteinases 2. Cancer Res. 56:2707–2710. [PubMed] [Google Scholar]