Self Antigens Expressed by Solid Tumors Do Not Efficiently Stimulate Naive or Activated T Cells: Implications for Immunotherapy (original) (raw)
Abstract
Induction and maintenance of cytotoxic T lymphocyte (CTL) activity specific for a primary endogenous tumor was investigated in vivo. The simian virus 40 T antigen (Tag) expressed under the control of the rat insulin promoter (RIP) induced pancreatic β-cell tumors producing insulin, causing progressive hypoglycemia. As an endogenous tumor antigen, the lymphocytic choriomeningitis virus (LCMV) glycoprotein (GP) was introduced also under the control of the RIP. No significant spontaneous CTL activation against GP was observed. However, LCMV infection induced an antitumor CTL response which efficiently reduced the tumor mass, resulting in temporarily normalized blood glucose levels and prolonged survival of double transgenic RIP(GP × Tag2) mice (137 ± 18 d) as opposed to control RIP-Tag2 mice (88 ± 8 d). Surprisingly, the tumor-specific CTL response was not sustained despite the facts that the tumor cells continued to express MHC class I and LCMV-GP–specific CTLs were present and not tolerized. Subsequent adoptive transfer of virus activated spleen cells into RIP(GP × Tag2) mice further prolonged survival (168 ± 11 d), demonstrating continued expression of the LCMV-GP tumor antigen and MHC class I. The data show that the tumor did not spontaneously induce or maintain an activated CTL response, revealing a profound lack of immunogenicity in vivo. Therefore, repetitive immunizations are necessary for prolonged antitumor immunotherapy. In addition, the data suggest that the risk for induction of chronic autoimmune diseases is limited, which may encourage immunotherapy against antigens selectively but not exclusively expressed by the tumor.
Tumor-specific cytotoxic T lymphocytes (CTLs) can prevent or eradicate tumors in a number of experimental systems (1–5). However, in other models CTLs failed to control tumor growth, which resembles the situation for most cancer patients. Possible reasons for the lack of antitumor CTL activity are selection of tumor antigen or MHC-negative tumor cells (6–9), defective lymphocyte homing to the tumor (10), lack of immunogenicity and costimulation for CTL activation (11–15), or functional or physical absence of tumor antigen–specific CTLs (16). Alternatively, efficient antitumor CTL activity may initially be induced but subsequently declines. It is of interest to characterize why tumor immunosurveillance fails so that antitumor immunotherapy can be improved.
Tumor antigens recognized by CTLs may be encoded by viral genes or by mutated cellular genes (1–3). These antigens are foreign to the tumor-bearing host and are thus tumor specific. However, detailed analyses have shown that most defined human tumor antigens are tumor selective but not entirely tumor specific, since physiological expression is also detected in testis, placenta, or melanocytes (17– 19). Since such antigens are expressed only in selected tissues and/or at low levels they may be less accessible for lymphocytes, but it is still possible that autoimmune disease may occur if CTLs are activated for therapeutic purposes. Nonetheless, if tissue-specific self antigens were used as targets for immunotherapy, the development and application of tumor vaccines might be simplified by targeting the same protein or major epitopes in different patients. Therefore, it is important to evaluate whether CTL activation against such antigens results in damage of healthy tissues and whether this would trigger chronic autoimmune disease.
To assess these questions in an in vivo model, we took advantage of the simian virus 40 tumor T antigen (Tag)1 which is a potent oncogene, largely due to its ability to inactivate two tumor suppressors, Rb and p53 (20, 21). We used transgenic mice expressing Tag under the control of the rat insulin promoter (RIP). The RIP-Tag2 line of transgenic mice develops pancreatic β-cell tumors leading to progressive autonomous insulin secretion and hypoglycemia, which is lethal when the mice reach the age of 3–4 mo (22). Histological sections show both hyperplasia and tumor formation by proliferating β cells. The transgenic mouse line RIP-Tag2 used in this study expresses Tag early during embryogenesis leading to immune tolerance such that Tag-specific lymphocytes are not detectable in the T cell repertoire (16).
As a tumor-associated antigen, we introduced the lymphocytic choriomeningitis virus (LCMV) glycoprotein (GP), also expressed under the control of the RIP (24). Like the RIP-Tag2 mice (23), these RIP-GP mice do not mount a spontaneous immune response against the transgenic product. However, in contrast to the RIP-Tag2 mice, there is no specific T cell tolerance to the transgenic neoantigen. As a consequence, infection with LCMV activates GP-specific CTLs which destroy the pancreatic β cells and induce type I autoimmune diabetes (24). LCMV is a natural mouse pathogen which replicates rapidly in vivo. It induces a strong immune response dominated by LCMV-specific cytotoxic T cells which proliferate to high numbers (∼25% of splenic mononuclear cells). These CTLs are responsible for rapid virus elimination occurring within ∼10 d and give rise to a long-term protective CTL memory (25–27). This strong CTL response was well suited for an in vivo model of tumor immunotherapy.
The two transgenic lines were crossed to obtain mice with β-islet cell tumors expressing LCMV-GP. The effectiveness of antitumor responses was evaluated by addressing the following questions in vivo: (a) Are LCMV-GP specific T cells spontaneously activated in tumor-bearing mice? (b) Does immunological unresponsiveness to LCMV-GP develop? (c) Do these mice mount a cytotoxic T cell response after immunization? (d) Will chronic autoimmune disease develop after induction of the tumor-specific LCMV-GP response? Our data show that the tumor antigen–specific CTLs were not spontaneously activated but efficiently reduced the tumor mass upon activation by virus infection. The tumor cells were profoundly incapable of stimulating an LCMV-GP–specific response in vivo since the antitumor CTL activity was not maintained but declined in parallel with the virus specific response.
Materials and Methods
Mice and Typing.
C57BL/6 (H-2b) mice were purchased from The Jackson Laboratory (Bar Harbor, ME). The Tag-transgenic mouse line RIP-Tag2, originally described as line RIP1-Tag2 (22), was crossed with GP-transgenic mice RIP-GP (24) to obtain RIP(GP × Tag2) double transgenic mice. Mice transgenic for a Vα2/Vβ8 TcR specific for H-2Db and the major LCMV epitope, i.e., the GP-derived peptide aa 33-41 (p33), were generated as described (27) and crossed with the RIP-Tag2 and RIP(GP × Tag2) transgenic mouse lines. All mice were H-2b; they were typed for the presence of the transgenes by PCR. The primer sequences and PCR conditions for the RIP-Tag2 transgene were GGACAAACCACAACTAGAATGCAGTG/CAGAGCAGAATTGTGGAGTGG and 60 s/94°C, 60 s/60°C, and 180 s/72°C; and for the RIP-GP transgene they were CAAGCAAGATGTAGAGTCTGCC/GGCTTTGGACATGAACCGCCC and 15 s/94°C, 30 s/53°C, and 30 s/72°C.
Tumor Monitoring, In Vivo Antitumor Treatment, and Adoptive Transfer Experiments.
Blood glucose levels were measured from blood obtained from the tail vein. The Haemogluco Test Kit was used according to the instructions of the supplier (Boehringer Mannheim Biochemicals, Indianapolis, IN). Tumor growth and pancreatic islet cell structure were examined by histology. For the induction of tumor-specific CTLs, the transgenic mice were immunized i.v. with LCMV Armstrong strain (2 × 105 PFU for TCR transgenic mice, 2,000 PFU for all other mice). For adoptive transfer experiments, spleen cells from C57BL/6 mice immunized with LCMV Armstrong (2,000 PFU) were collected 8 d after infection. 5 × 107 spleen cells were injected into the tail vein of control mice and of RIP(GP × Tag2) with recurrent hypoglycemia.
Proliferation Assays.
Responder spleen cells (105) were cultured with 20 Gy-irradiated stimulator macrophages (2 × 104) previously incubated with 10 nM LCMV-GP–derived peptide aa 33-41 (p33) recognized by LCMV-GP–specific CTLs. The cultures were maintained in 96-well flat-bottomed microtiter plates at 37°C in a 5% CO2 humidified atmosphere. IMDM was supplemented with 10% FCS, 5 × 10−5 M 2-mercaptoethanol, penicillin, and streptomycin (complete IMDM). After 3 d, the cultures were pulsed for 16 h with 1 μCi of [3H]thymidine, harvested, and counted on a Matrix 96 direct beta counter (Canberra Packard). The data are expressed as mean cpm of triplicate cultures.
Cytotoxicity Tests.
To detect secondary effector activity in mice analyzed 30 d after LCMV infection, spleen cells (3 × 106/ well) were cocultured with irradiated C57BL/6J stimulator macrophages (3 × 105/well) previously incubated with peptide p33 (10 nM) in complete IMDM at 37°C for 5 d. Effector functions from untreated mice or 8 d after LCMV infection were tested directly ex vivo. Effector cells were tested in duplicate cultures and serial dilutions in 96-well round-bottomed plates during 5 h in 51Cr (NEN/DuPont, Boston, MA) release assays using MC57G fibroblast target cells (H-2b) previously incubated with 10 nM peptide p33 for 1 h at 37°C. The supernatant of the cytotoxicity cultures was counted in a Cobra II Gamma Counter (Canberra Packard, Meriden, CT). Percentage of specific lysis was calculated as (experimental release − spontaneous release)/(total release − spontaneous release) × 100.
Immunohistology.
Freshly removed organs were immersed in HBSS and snap-frozen in liquid nitrogen. Tissue sections of 5-μm thickness were cut in a cryostat and fixed in acetone for 10 min. Sections were then incubated with the primary rat mAbs M1/42 specific for mouse MHC class I (28), YTS 191 specific for CD4, or YTS 169 specific for CD8 (29) for 30 min at room temperature, followed by a two-step indirect immunoenzymatic staining procedure. First, alkaline phosphatase–labeled rat-specific goat immunoglobulins (Tago, Inc., Burlingame, CA) were applied for 30 min at room temperature, followed by alkaline phosphatase– labeled goat-specific immunoglobulins (Jackson Immunoresearch Labs, Inc., West Grove, PA) for another 30 min. Dilutions were performed in 0.1 M Tris-HCl (pH 7.4) containing 5% normal mouse serum. Alkaline phosphatase was then detected by a red color reaction using naphthol AS-BI phosphate and New Fuchsin as a substrate. Endogenous alkaline phosphatase was blocked by Levamisol. Sections were counterstained with Mayer's hemalum for 2 min.
Results
Hypoglycemia Due to Insulin Overproduction by β-islet Tumor Cells.
To generate double transgenic RIP(GP × Tag2) mice, tumor-susceptible RIP-Tag2 mice were crossed with transgenic RIP-GP mice expressing the LCMV-GP on pancreatic β cells. To monitor tumor progression, blood glucose was measured regularly. Pathological hypoglycemia (i.e., multiple values <5 mM glucose) corresponded with a large tumor burden since the β-islet tumor cells autonomously released high quantities of insulin (22). This occurred in tumor-bearing RIP-Tag2 and RIP(GP × Tag2) mice, which both developed hypoglycemia at the age of 70–85 d (Fig. 1 A). As expected, control C57BL/6 and RIP-GP mice showed normal blood glucose levels. The RIP(GP × Tag2) mice developed tumors and hypoglycemia with identical kinetics to RIP-Tag2 mice suggesting that the LCMV-GP specific T cells did not spontaneously develop tumor protective activity.
Figure 1.

Blood glucose levels reflect the tumor burden and the effectiveness of tumor immunotherapy in vivo. (A) RIP(GP × Tag2) and RIP-Tag2 mice developed hypoglycemia indicative of β-cell hyperplasia and insulin overproduction, while control RIP-GP and C57BL/6 mice showed normal glucose levels. (B) Tag2 transgenic animals with two consecutive blood glucose readings <5 mM (at an age of 70–85 d) and age-matched control mice were infected with LCMV to induce tumor-specific CTLs. After infection, the GP-expressing mice (but not those without GP) increased the blood glucose levels. This effect was reversed in the RIP(GP × Tag2) mice and hypoglycemia relapsed within 4 wk. In 1 out of 10 RIP(GP × Tag2) mice the LCMV-induced hyperglycemia was very high and lethal. The figures show 1 mouse/group representative for 7–20 mice/group (4 mice for C57BL/6). ▪, RIP-GP; •, RIP(GP × Tag2); ○, RIP-Tag2; □, C57BL/6.
No Detectable Spontaneous T Cell Responses Against the LCMV-GP Tumor Antigen.
It was important to directly examine the GP-specific T cell responses by the tumor-bearing mice. Spleen cells were stimulated with irradiated macrophages pulsed with peptide p33, the major epitope of LCMV-GP in the H-2b haplotype (30). Fig. 2 A (closed bars) shows that the T cells from untreated RIP-GP, RIP-Tag2, and double transgenic mice did not proliferate significantly. Proliferation could be observed in spleen cells derived from mice transgenic for a GP-specific TCR. Because this assay can easily detect GP-specific memory CTLs from LCMV-immunized C57BL/6 mice (see below), the results indicated that no efficient spontaneous lymphocyte activation against the LCMV-GP tumor antigen had occurred in vivo (23, 24).
Figure 2.


GP-specific T cells were neither spontaneously activated nor rendered unresponsive by the tumor. (A) LCMV-GP–specific proliferative responses. Spleen cells were taken from untreated mice (closed bars), 8 d after LCMV infection (open bars), or 30 d after LCMV infection (hatched bars) and incubated in vitro with stimulator macrophages treated with the LCMV glycoprotein peptide p33. Data are mean ± SD of triplicate cultures. Control cultures using macrophages without peptide had background levels <3,000 cpm (not shown). (B–E) Normal LCMV-specific cytotoxic responses were detected 8 d after LCMV infection (B and C), or 30 d after LCMV infection upon restimulation in vitro for 5 d (D and E). In the RIP(GP × Tag2) mice studied 30 d after LCMV infection the tumors had grown back and blood glucose was low (3–5 mM). No data were obtained for day 30 RIP-GP or RIP-Tag2 mice (n.d., not determined) because these animals were killed due to diabetes or tumor burden. Target cells were MC57G fibroblasts untreated (C and E) or treated with peptide p33 (B and D). The results shown are representative for three independent experiments and analysis of at least four mice per group. ▪, RIP-GP; •, RIP(GP × Tag2); ○, RIP-Tag2; □, C57BL/6.
Tumor Regression upon LCMV Infection.
To activate tumor-specific T cells, mice were immunized i.v. with LCMV. This was done at the age of 80 d or, in tumor-bearing mice, after two consecutive blood glucose readings <5 mM. Fig. 1 B shows that early after infection, the single transgenic RIP-GP mice developed hyperglycemia as seen in autoimmune diabetes due to β-cell destruction by the GP specific CTLs (24). Importantly, the blood glucose levels of the tumor-bearing RIP(GP × Tag2) mice increased also. This suggested that the CTLs destroyed insulin producing tumor cells and normal β-islet cells. However, hypoglycemia recurred, probably due to the waning of the LCMV response and the continuous expansion of insulin-producing tumor cells. The control mice not expressing GP in their islets remained normoglycemic (C57BL/6) or showed progressive hypoglycemia due to insulin production by the β-cell tumor (RIP-Tag2) which was similar to that in uninfected mice (not shown).
The functional LCMV-GP–specific response was also examined in these animals. Spleen cells taken 8 d after LCMV infection showed strong in vitro proliferative responses in all the mice (Fig. 2 A, open bars). GP-specific cytolytic T cell activity was measured in 51Cr release assays, revealing strong cytotoxic responses specific for p33 in all the mice tested (Fig. 2, B and C). Together, these data demonstrated that LCMV infection led to efficient functional CTL activation which was associated with specific regression of those tumors which expressed LCMV-GP.
To further examine tumor burden and the consequences of the induction of an antiviral/antitumor response, the pancreata were examined histologically. This confirmed that the β cell tumors in single transgenic RIP-Tag2 mice were not affected by the LCMV infection. In contrast, in RIP(GP × Tag2) mice pancreatic β-islets were infiltrated with inflammatory cells including CTLs at maximal levels 8 d after infection, leading to tumor mass reduction and increased blood glucose levels. Fig. 3 shows that the tumors in RIP(GP × Tag2) transgenic mice were infiltrated by numerous CD8+ and many CD4+ lymphocytes, whereas the tumors in RIP-Tag2 mice had only limited lymphocyte infiltration. These findings correlated with the in vitro results (Fig. 2) of minimal spontaneous lymphocyte responses in uninfected tumor-bearing mice but very strong CTL activity upon viral immunization. Thus, the tumor-specific CTLs were efficiently activated by LCMV infection but not by the tumor.
Figure 3.
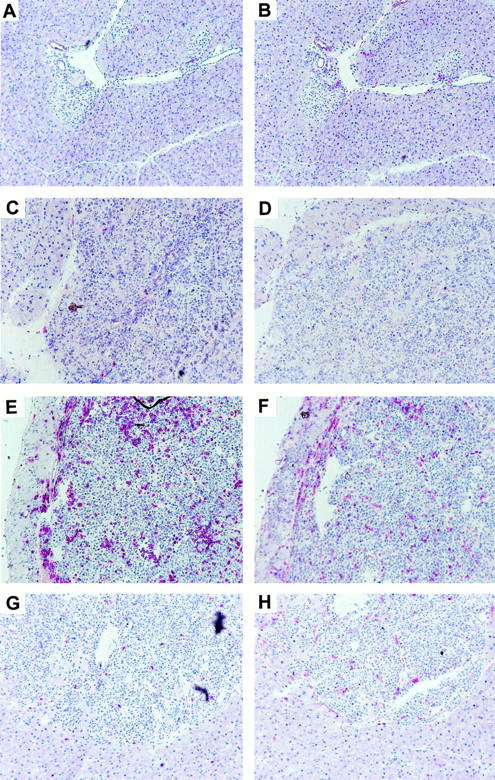
Intense lymphocytic infiltration after LCMV infection in double transgenic RIP(GP × Tag2) but not in RIP-Tag2 single transgenic mice. Mice were immunized with LCMV i.v. and killed on day 8 (A–F). Tag transgenic mice showed islet hyperplasia and tumor formation by proliferating β cells (C–H). Immunohistochemical analysis with CD8-specific (left) or CD4-specific (right) antibodies showed intense lymphocytic infiltration in the tumors of RIP-GP × Tag2 (E and F) but not RIP-Tag2 (C and D) mice or nontransgenic mice (A and B). In uninfected mice (G and H), the tumors showed minimal lymphocyte infiltration (only RIP-GP × Tag2 shown).
Systematic comparisons of histological sections confirmed that significant CD8+ cell infiltration was found only in LCMV-immunized mice where the β-islet cells expressed the GP (Table 1). Interestingly, the lymphocytic infiltrations in small β-cell hyperplasias and large tumors were of similar intensity (data not shown), indicating that even advanced tumors remained susceptible to CTLs. Morphometric analysis of pancreas sections further revealed that the insulinomas in LCMV-treated RIP(GP × Tag2) mice had many regions with relatively low density of viable tumor cells. Finally, the insulinomas in LCMV-infected RIP(GP × Tag2) mice were significantly smaller than in RIP-Tag2 mice or in uninfected controls. These data confirmed that LCMV-activated CTLs directly destroyed GP expressing tumor cells and reduced tumor mass.
Table 1.
Effect of Virus Infection on Infiltration of CD8+ Cells, Tumor Cell Density, and Tumor Size
| RIP (GP × Tag2) | P | RIP-Tag2 | |
|---|---|---|---|
| 8 d after LCMV infection | |||
| CD8+ cell infiltration (%) | 39.13 ± 8.5 | <0.0001 | 2.59 ± 2.40 |
| Tumor cell density (%) | 71.25 ± 10.7 | <0.001 | 92.65 ± 4.39 |
| Tumor size (mm2) | 0.25 ± 0.07 | 0.002 | 0.46 ± 0.46 |
| Uninfected | |||
| CD8+ cell infiltration (%) | 3.88 ± 1.90 | NS | 2.11 ± 1.66 |
| Tumor cell density (%) | 94.13 ± 4.01 | NS | 95.11 ± 2.42 |
| Tumor size (mm2) | 0.51 ± 0.29 | NS | 0.64 ± 0.20 |
Prolonged Survival Due to Immunotherapy.
To evaluate whether the CTL-mediated tumor destruction was of extended therapeutic value, we monitored disease progression and survival. All mice were infected with LCMV after two consecutive blood glucose readings <5 mM. Tumor-bearing RIP-Tag2 mice had a mean life expectancy of 88 ± 8 d (Table 2), similar to uninfected RIP-Tag2 and RIP(GP × Tag2) mice (not shown). However, LCMV infection of RIP(GP × Tag2) mice lead to a significant increase in survival with a mean of 137 ± 18 d (P <0.001). Thus, a single acute LCMV infection with its known short duration of ∼10 d (25) prolonged survival considerably, demonstrating the intense immune response elicited by LCMV and the potential for tumor immunotherapy.
Table 2.
Life Span of Tumor Mice after Virus Infection
| Lineage | No. of mice | Life span* | P |
|---|---|---|---|
| RIP-Tag2 | 7 | 88 ± 8 | |
| <0.001 | |||
| RIP(GP × Tag2) | 9 | 137 ± 18 | |
| <0.005 | |||
| RIP(GP × Tag2) + AT | 4 | 168 ± 11 | |
| TCR RIP-Tag2 | 4 | 90 ± 17 | |
| <0.0001 | |||
| TCR RIP(GP × Tag2) | 4 | 183 ± 5 |
We crossed the RIP(GP × Tag2) mice with mice transgenic for an LCMV-GP–specific TCR to investigate whether a high frequency of tumor-specific CTLs would increase the efficiency of virus-mediated antitumor therapy. Indeed, the survival of LCMV-infected triple transgenic TCR RIP(GP × Tag2) mice was much longer as compared to the survival of TCR RIP-Tag2 mice (Table 2). Thus, the TCR transgenic mice showed an even greater prolongation of survival due to more efficient CTL-mediated tumor destruction. These data show that virus-induced CTL-mediated immunotherapy significantly controlled pancreatic β-cell tumor progression. Ultimately, the tumor-bearing mice nevertheless succumbed to lethal hypoglycemia, demonstrating that a single virus challenge did not trigger a chronic antitumor response and permanent CTL activity.
Persistence of Functionally Competent CTLs.
We investigated whether the GP-specific CTLs had been functionally or physically eliminated during or after infection because of the presence of the GP-expressing tumors. Spleen cells from immunized RIP(GP × Tag2), control C57BL/6, or control TCR transgenic mice obtained 30 d after in vivo infection proliferated normally upon GP-specific in vitro stimulation with peptide p33–treated macrophages (Fig. 2 A, hatched bars). After 5 d of such in vitro stimulation, a normal peptide-specific cytotoxic response was also detected (Fig. 2, D and E). These data show that tumor antigen–specific CTLs functioned normally and were not tolerized in vivo.
Continued Tumor Susceptibility to CTL-mediated Immunotherapy.
We examined other parameters which may have limited the antitumor therapy in the RIP(GP × Tag2) mice. Immunohistological sections from pancreas revealed positive staining for MHC class I expression in tumors from RIP(GP × Tag2) mice 30 d after LCMV infection (data not shown), suggesting that the tumor cells did not downregulate MHC class I. But did the tumors also continue to present the cognate peptide? We performed functional experiments to determine whether those tumor cells which had survived a previous immunotherapy would remain susceptible to tumor antigen–specific CTLs. Spleen cells from LCMV immune C57BL/6 mice were adoptively transferred into RIP(GP × Tag2) mice which had been immunized previously with LCMV and had redeveloped hypoglycemia. As shown in Table 2, this additional treatment significantly (P <0.005) prolonged the survival with a mean of 168 ± 11 d as compared to mice that had only been immunized once with LCMV (137 ± 18 d). Therefore, the tumor cells had not developed secondary immune escape mechanisms but remained susceptible to CTL-mediated immunotherapy.
Discussion
Most models which examine tumor immunity are based on transplanted syngeneic tumors. However, primary tumors which naturally develop and grow in vivo are likely to represent more realistic models for human cancers. Using the latter approach, this study was designed to examine the potential for efficient tumor immunotherapy in vivo. No spontaneous tumor regression was detectable despite the presence of LCMV-GP–specific CTLs and LCMV-GP expressed by tumor cells. However, virus infection induced tumor-specific CTLs infiltrating small and even large β-cell tumors and mediating efficient immunotherapy with significantly prolonged survival of tumor-bearing mice. Nevertheless, after viral clearance, the CTLs were no longer active and no chronic autoimmune diabetes was induced despite continuous expression of antigen/MHC by the tumor and persistence of functional tumor-specific CTLs. This experimental model thus demonstrates an efficient tumor therapy (by virus infection) capable of treating established primary tumors. However, the tumors themselves were profoundly nonimmunogenic, since they could not activate T cells or maintain an activated T cell response. The data show that the tumors behaved similarly to normal nonlymphoid peripheral tissues which fail to activate immune responses despite the existence of self-specific CTLs (24, 31).
These results relate to several important issues for tumor immunotherapy: (a) Why are peripheral epithelial, neurectodermal or mesenchymal tumors antigenic but fail to induce an immune response? Are they ignored by T cells or do they anergize or tolerize T cells? (b) How can an antitumor T cell response be maintained to effectively control tumors? (c) Will the T cell response cause chronic autoimmune disease?
(a) Immunogenicity Versus Antigenicity of Peripheral Tumors.
That tumors are often antigenic but not immunogenic has been recognized for many years (32–35). More recently, the detailed analysis of the so called tum− variants obtained by in vitro treatment with mutagens revealed that mice which had rejected tum− variants were significantly protected against a challenge with the original tumor (11, 12). This indicated that the latter was antigen positive. Since then it has been shown that many tumors express antigens which may be recognized by CTLs, and thus display the necessary targets for potential immunotherapy.
Several mechanisms may contribute to the failure of tumor immunosurveillance. Models have demonstrated that downregulation of peptide/MHC expression (6–9), lack of costimulation (11–15), the induction of lymphocyte unresponsiveness (16), local fibrin formation or alterations in the glycocalyx, or changes in homing receptor expression (10) may all contribute to tumor evasion from the immune system. In our model, virus infection and/or adoptive transfer of virus-activated lymphocytes remained an efficient means to counteract tumor expansion in vivo. Thus, there was no obvious tolerization of tumor-specific CTLs, and the tumors have not actively escaped immunosurveillance. Although we cannot entirely exclude the possibility that some of the above mentioned escape mechanisms have occurred to some degree in our model, it is evident that such mechanisms were not dominant. Rather, the lack of immunogenicity of tumor cells was the major feature with respect to the failure of immunosurveillance.
In vivo, adequate spontaneous activation of tumor specific lymphocytes either does not occur or often results in inefficient tumor protection. This is in part because the tumor cell itself is not a professional antigen-presenting cell and therefore is relatively inefficient at activating tumor-specific effector cells. Previous studies have shown that if a tumor cell or nonprofessional antigen-presenting cell reaches secondary lymphoid organs, measurable cytotoxic T cell responses are nevertheless induced against epithelial and mesenchymal malignancies. Thus, whether or not a T cell immune response is induced is related to the physiological location of the tumor itself (32, 36). In many cases, the tumor is not in an environment which enhances an antitumor response, or tumor metastasis to organized lymphatic tissues occurs too late to induce an effective antitumor response in vivo.
Recognition of antigen by naive T cells in the absence of costimulation has been postulated to induce T cell unresponsiveness. Since most carcinomas do not express costimulatory molecules, it has been suggested that tumor therapy may only be feasible in patients with recent onset cancers, whereas in advanced disease all the tumor-specific T cells will have become unresponsive (4). However, our data do not support this notion since they show that tumors may progress without significantly altering tumor-specific T cell responsiveness. This is in parallel to antigens of healthy tissues that neither induce immunological tolerance nor immune responses but remain invisible for T cells (24, 31). Thus, carcinomas and sarcomas, similar to solid tissue antigens which do not reach secondary lymphoid organs, may be ignored by T cells. Appropriate immunization could therefore induce an efficient T cell immunity against such tumor antigens even after extended periods of tumor burden.
(b) Maintenance of an Antitumor T Cell Response.
Our data indicate that the lack of tumor immunogenicity may be more profound than previously suspected, since the tumor was not even sufficiently immunogenic to maintain an activated and ongoing strong immune response. Despite persistence of functionally competent specific CTLs and of antigen-expressing tumor cells, the CTL effector function declined. This result indicates that antigen must be presented in an immunogenic form not only for induction but also for maintenance of T cell responses, including protective T cell memory. The fact that the CTL effector function was not sustained by the tumor is a novel finding, which has important implications for immunotherapy. It indicates that repetitive vaccination or adoptive transfer of activated tumor-specific CTL is necessary to sustain antitumor activity in cancer patients.
(c) Antitumor Immunity and Autoimmune Disease.
Damage to healthy organs may be acceptable to some extent, as widely illustrated by many radiation and chemotherapeutic techniques which are partially but not highly specific for tumor tissues. However, in the case of immunotherapy specific for tumor antigens which are also expressed by some healthy tissues, there is the possibility of inducing uncontrolled and progressive chronic autoimmune disease. It has been demonstrated that the lymphocytes from patients with vitiligo may be activated against tyrosinase and other melanocyte-specific antigens (37–39) and that spontaneous appearance of vitiligo in persons with metastatic melanoma was associated with improved prognosis (40, 41). Discrete and limited damage of normal skin has recently also been demonstrated experimentally where passive immunization with antibodies directed against the melanocyte differentiation antigen gp75 (tyrosinase-related protein 1) was successfully used to treat melanomas in mice (42). Both clinical and experimental evidence suggests that the damage of healthy tissues remains limited despite effective antitumor immune activity (37–43). In our model, not only tumor cells but also normal β cells were targets for CTLs (24), and it is possible that tumor cells compensated for reduced insulin production by normal β cells. As can be expected from the sharing of a specific antigen by tumor and normal cells, the CTL cytotoxicity was not tumor specific. However, the important finding was that the CTL activity was limited in time. Thus, although reactivity to healthy tissue may be difficult to avoid, there was no development of progressive chronic autoimmune disease. These data indicate that CTL activity can be controlled by the intervals and intensity of the applied immunization procedures.
We conclude that solid tumors may not be sufficiently immunogenic either to initiate or to maintain a beneficial or therapeutic immune response. Therefore, efficient tumor surveillance must include both improved and accelerated induction by using vaccines localizing tumor antigens to secondary lymphoid organs and booster immunizations to sustain the antitumor effect. Antigens selectively but not exclusively expressed by the tumor may be considered targets for antitumor CTL therapy, since CTL activity is limited in time and thus induction of chronic autoimmune disease is unlikely.
Acknowledgments
We would like to thank Ming Tsao for his help with histology and Drs. Maurice Zauderer and Ruth Ganss for comments to the manuscript.
This work was supported by the Medical Research Council (MRC) of Canada and the Canadian Diabetes Association. P.S. Ohashi is supported by an MRC scholarship.
Footnotes
1
Abbreviations used in this paper: GP, glycoprotein; LCMV, lymphocytic choriomeningitis virus; RIP, rat insulin promoter; Tag, T antigen.
References
- 1.Boon T, Cerottini J-C, Van den Eynde B, van der Bruggen P, Van Pel A. Tumor antigens recognized by T lymphocytes. Annu Rev Immunol. 1994;12:337–365. doi: 10.1146/annurev.iy.12.040194.002005. [DOI] [PubMed] [Google Scholar]
- 2.Slingluff CL, Jr, Hunt DF, Engelhard VH. Direct analysis of tumor-associated peptide antigens. Curr Opin Immunol. 1994;6:733–740. doi: 10.1016/0952-7915(94)90077-9. [DOI] [PubMed] [Google Scholar]
- 3.Melief CJM, Kast WM. T-cell immunotherapy of tumors by adoptive transfer of cytotoxic T lymphocytes and by vaccination with minimal essential epitopes. Immunol Rev. 1994;145:167–177. doi: 10.1111/j.1600-065x.1995.tb00081.x. [DOI] [PubMed] [Google Scholar]
- 4.Hellström KE, Hellström I, Chen L. Can co-stimulated tumor immunity be therapeutically efficacious? . Immunol Rev. 1994;145:123–145. doi: 10.1111/j.1600-065x.1995.tb00079.x. [DOI] [PubMed] [Google Scholar]
- 5.Ostrand-Rosenberg S. Tumor immunotherapy: the tumor cell as an antigen-presenting cell. Curr Opin Immunol. 1994;6:722–727. doi: 10.1016/0952-7915(94)90075-2. [DOI] [PubMed] [Google Scholar]
- 6.Ye X, McCarrick J, Jewett L, Knowles BB. Timely immunization subverts the development of peripheral nonresponsiveness and suppresses tumor development in simian virus 40 tumor antigen-transgenic mice. Proc Natl Acad Sci USA. 1994;91:3916–3920. doi: 10.1073/pnas.91.9.3916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cromme FV, Airey J, Heemels M-T, Ploegh HL, Keating PJ, Stern PL, Meijer CJLM, Walboomers JMM. Loss of transporter protein, encoded by the TAP-1 gene, is highly correlated with loss of HLA expression in cervical carcinomas. J Exp Med. 1994;179:335–340. doi: 10.1084/jem.179.1.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cohen EP, Kim TS. Neoplastic cells that express low levels of MHC class I determinants escape host immunity. Semin Cancer Biol. 1994;5:419–428. [PubMed] [Google Scholar]
- 9.Garrido F, Ruiz-Cabello F, Cabrera T, Perez-Villar JJ, Lopez-Botet M, Duggan-Keen M, Stern PL. Implications for immunosurveillance of altered HLA class I phenotypes in human tumours. Immunol Today. 1997;18:89–95. doi: 10.1016/s0167-5699(96)10075-x. [DOI] [PubMed] [Google Scholar]
- 10.Onrust SV, Hartl PM, Rosen SD, Hanahan D. Modulation of L-selectin ligand expression during an immune response accompanying tumorigenesis in transgenic mice. J Clin Invest. 1996;97:54–64. doi: 10.1172/JCI118406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Boon T, Kellermann O. Rejection by syngeneic mice of cell variants obtained by mutagenesis of a malignant teratocarcinoma cell line. Proc Natl Acad Sci USA. 1977;74:272–275. doi: 10.1073/pnas.74.1.272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Boon T, Van Pel A. Teratocarcinoma cell variants rejected by syngeneic mice: protection of mice immunized with these variants against other variants and against the original malignant cell line. Proc Natl Acad Sci USA. 1978;75:1519–1523. doi: 10.1073/pnas.75.3.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen L, Ashe S, Brady WA, Hellström I, Hellström KE, Ledbetter JA, McGowan P, Linsley PS. Costimulation of antitumor immunity by the B7 counterreceptor for the T lymphocyte molecules CD28 and CTLA-4. Cell. 1992;71:1093–1102. doi: 10.1016/s0092-8674(05)80059-5. [DOI] [PubMed] [Google Scholar]
- 14.Townsend SE, Allison JP. Tumor rejection after direct costimulation of CD8+T cells by B7-transfected melanoma cells. Science (Wash DC) 1993;259:368–370. doi: 10.1126/science.7678351. [DOI] [PubMed] [Google Scholar]
- 15.Huang AYC, Bruce AT, Pardoll DM, Levitsky HI. Does B7-1 expression confer antigen-presenting cell capacity to tumors in vivo? . J Exp Med. 1996;183:769–776. doi: 10.1084/jem.183.3.769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Förster I, Hirose R, Arbeit JM, Clausen BE, Hanahan D. Limited capacity for tolerization of CD4+ T cells specific for a pancreatic β cell neo-antigen. Immunity. 1995;2:573–585. doi: 10.1016/1074-7613(95)90002-0. [DOI] [PubMed] [Google Scholar]
- 17.Houghton AN. Cancer antigens: immune recognition of self and altered self. J Exp Med. 1994;180:1–4. doi: 10.1084/jem.180.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nanda NK, Sercarz EE. Induction of anti–self-immunity to cure cancer. Cell. 1995;82:13–17. doi: 10.1016/0092-8674(95)90047-0. [DOI] [PubMed] [Google Scholar]
- 19.Van Pel A, van der Bruggen P, Coulie PG, Brichard VG, Lethe B, Van den Eynde B, Uyttenhove C, Renauld J-C, Boon T. Genes coding for tumor antigens recognized by cytolytic T lymphocytes. Immunol Rev. 1995;145:229–250. doi: 10.1111/j.1600-065x.1995.tb00084.x. [DOI] [PubMed] [Google Scholar]
- 20.Livingston, D.M. 1992. Functional analysis of the retinoblastoma gene product and of RB-SV40 T antigen complexes. In Tumor Suppressor Genes, the Cell Cycle and Cancer. A.J. Levine and L.M. Franks, editors. Imperial Cancer Research Fund, Cold Spring Harbor, NY. 153–160. [PubMed]
- 21.Ludlow JW. Interactions between SV40 large-tumor antigen and the growth suppressor proteins pRB and p53. FASEB (Fed Am Soc Exp Biol) J. 1993;7:866–871. doi: 10.1096/fasebj.7.10.8344486. [DOI] [PubMed] [Google Scholar]
- 22.Hanahan D. Heritable formation of pancreatic β-cell tumours in transgenic mice expressing recombinant insulin/ simian virus 40 oncogenes. Nature (Lond) 1985;315:115–122. doi: 10.1038/315115a0. [DOI] [PubMed] [Google Scholar]
- 23.Adams TE, Alpert S, Hanahan D. Non-tolerance and autoantibodies to a transgenic self antigen expressed in pancreatic β cells. Nature (Lond) 1987;325:223–228. doi: 10.1038/325223a0. [DOI] [PubMed] [Google Scholar]
- 24.Ohashi PS, Oehen S, Bürki K, Pircher H, Ohashi CT, Odermatt B, Malissen B, Zinkernagel R, Hengartner H. Ablation of “tolerance” and induction of diabetes by virus infection in viral antigen transgenic mice. Cell. 1991;65:305–317. doi: 10.1016/0092-8674(91)90164-t. [DOI] [PubMed] [Google Scholar]
- 25.Lehmann-Grube F. Lymphocytic choriomeningitis virus. Virol Monogr. 1971;10:1–173. [Google Scholar]
- 26.Moskophidis D, Assmann-Wischer U, Simon MM, Lehmann-Grube F. The immune response of the mouse to lymphocytic choriomeningitis virus. V. High numbers of cytolytic T lymphocytes are generated in the spleen during acute infection. Eur J Immunol. 1987;17:937–942. doi: 10.1002/eji.1830170707. [DOI] [PubMed] [Google Scholar]
- 27.Pircher H, Bürki K, Lang R, Hengartner H, Zinkernagel R. Tolerance induction in double specific T-cell receptor transgenic mice varies with antigen. Nature (Lond) 1989;342:559–561. doi: 10.1038/342559a0. [DOI] [PubMed] [Google Scholar]
- 28.Springer, T.A. 1980. Cell-surface differentiation in the mouse. In Monoclonal Antibodies. R.H. Kennett, T.J. McKearn, and K.B. Bechtol, editors. Plenum Press, New York. 185–217.
- 29.Cobbold SP, Jayasuriya A, Nash A, Prospero TD, Waldmann H. Therapy with monoclonal antibodies by elimination of T cell subsets in vivo. Nature (Lond) 1984;312:548–551. doi: 10.1038/312548a0. [DOI] [PubMed] [Google Scholar]
- 30.Klavinskis LS, Whitton JL, Joly E, Oldstone MBA. Vaccination and protection from a lethal viral infection: identification, incorporation, and use of a cytotoxic T lymphocyte glycoprotein epitope. Virology. 1990;178:393–400. doi: 10.1016/0042-6822(90)90336-p. [DOI] [PubMed] [Google Scholar]
- 31.von Herrath MG, Dockter J, Oldstone MBA. How virus induces a rapid or slow onset insulin-dependent diabetes mellitus in a transgenic model. Immunity. 1994;1:231–242. doi: 10.1016/1074-7613(94)90101-5. [DOI] [PubMed] [Google Scholar]
- 32.Prehn RT, Main JM. Immunity to methylcholanthrene-induced sarcomas. J Natl Cancer Inst. 1957;18:769–782. [PubMed] [Google Scholar]
- 33.Klein E, Klein G. Antigenic properties of lymphomas induced by the Moloney agent. J Natl Cancer Inst. 1964;32:547–565. [PubMed] [Google Scholar]
- 34.Kripke ML. Antigenicity of murine skin tumors induced by ultraviolet light. J Natl Cancer Inst. 1974;53:1333–1336. doi: 10.1093/jnci/53.5.1333. [DOI] [PubMed] [Google Scholar]
- 35.Zinkernagel RM, Pircher H, Ohashi PS, Oehen S, Odermatt B, Mak TW, Arnheiter H, Bürki K, Hengartner H. T and B cell tolerance and responses to viral antigens in transgenic mice: implications for the pathogenesis of autoimmune versus immunopathological disease. Immunol Rev. 1991;122:133–171. doi: 10.1111/j.1600-065x.1991.tb00601.x. [DOI] [PubMed] [Google Scholar]
- 36.Kündig TM, Bachmann MF, DiPaolo C, Simard JJL, Battegay M, Lother H, Gessner A, Kühlcke K, Ohashi PS, Hengartner H, Zinkernagel RM. Fibroblasts as efficient antigen-presenting cells in lymphoid tissues. Science (Wash DC) 1995;268:1343–1347. doi: 10.1126/science.7761853. [DOI] [PubMed] [Google Scholar]
- 37.Naughton GK, Eisinger M, Bystryn JC. Autoantibodies to normal human melanocytes in vitiligo. J Exp Med. 1983;158:246–251. doi: 10.1084/jem.158.1.246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Song Y-H, Conner E, Li Y, Zorovich B, Balducci P, Maclaren N. The role of tyrosinase in autoimmune vitiligo. Lancet. 1994;344:1049–1052. doi: 10.1016/s0140-6736(94)91709-4. [DOI] [PubMed] [Google Scholar]
- 39.Visseren MJW, van Elsas A, van der Voort EIH, Ressing ME, Kast WM, Schrier PI, Melief CJM. CTL specific for the tyrosinase autoantigen can be induced from healthy donor blood to lyse melanoma cells. J Immunol. 1995;154:3991–3998. [PubMed] [Google Scholar]
- 40.Nordlund JJ, Kirkwood JM, Forget BM, Milton G, Albert DM, Lerner AB. Vitiligo in patients with metastatic melanoma: a good prognostic sign. J Am Acad Derm. 1983;9:689–695. doi: 10.1016/s0190-9622(83)70182-9. [DOI] [PubMed] [Google Scholar]
- 41.Kawakami Y, Eliyahu S, Delgado CH, Robbins PF, Sakaguchi K, Appella E, Yanelli JR, Adema GJ, Miki T, Rosenberg SA. Identification of a human melanoma antigen recognized by tumor-infiltrating lymphocytes associated with in vivo tumor rejection. Proc Natl Acad Sci USA. 1994;91:6458–6462. doi: 10.1073/pnas.91.14.6458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hara I, Takechi Y, Houghton AN. Implicating a role for immune recognition of self in tumor rejection: immunization against the brown locus protein. J Exp Med. 1995;182:1609–1614. doi: 10.1084/jem.182.5.1609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hu J, Kindsvogel W, Busby S, Bailey MC, Shi YY, Greenberg PD. An evaluation of the potential to use tumor-associated antigens as targets for antitumor T cell therapy using transgenic mice expressing a retroviral tumor antigen in normal lymphoid tissues. J Exp Med. 1993;177:1681–1690. doi: 10.1084/jem.177.6.1681. [DOI] [PMC free article] [PubMed] [Google Scholar]