Casein kinase Iɛ in the Wnt pathway: Regulation of β-catenin function (original) (raw)
Abstract
Wnt and its intracellular effector β-catenin regulate developmental and oncogenic processes. Using expression cloning to identify novel components of the Wnt pathway, we isolated casein kinase Iɛ (CKIɛ). CKIɛ mimicked Wnt in inducing a secondary axis in Xenopus, stabilizing β-catenin, and stimulating gene transcription in cells. Inhibition of endogenous CKIɛ by kinase-defective CKIɛ or CKIɛ antisense-oligonucleotides attenuated Wnt signaling. CKIɛ was in a complex with axin and other downstream components of the Wnt pathway, including Dishevelled. CKIɛ appears to be a positive regulator of the pathway and a link between upstream signals and the complexes that regulate β-catenin.
Wnt regulates developmental and oncogenic processes through its downstream effector, β-catenin (1–3). Intracellular protein complexes, including Dishevelled (Dvl/Dsh), glycogen synthase kinase-3β (GSK-3β), axin and adenomatous polyposis coli (APC) protein, regulate cytosolic β-catenin protein levels. However, little is known about how Wnt or other upstream stimuli regulate these complexes. Most components of the Wnt pathway have been found by genetic approaches in Drosophila. Mutations in several molecules in the wingless (Wg, the Drosophila homologue of Wnt) pathway, such as Dvl/Dsh, β-catenin, and lymphoid enhancer factor-1 (Lef-1)/T cell factor (Tcf), caused segment polarity phenotypes in Drosophila similar to the Wg phenotype, suggesting that these molecules are positive regulators of the pathway (4, 5). Genetic studies in Drosophila also revealed that GSK-3β is a negative regulator of this pathway. GSK-3β is in a complex containing other negative regulators, axin and APC protein, and a positive regulator, β-catenin (6–10). β-catenin is an extensively studied effector in the pathway and has a pivotal role in both developmental processes and oncogenesis (2, 3). Upon Wnt stimulation, β-catenin protein is stabilized and moves to the nucleus where it forms a complex with and activates Lef-1/Tcf transcription factors (11, 12). Mutated forms of β-catenin appear to be involved in cancer and induce Lef-1/Tcf-dependent transcription even in the absence of Wnt stimulation (13, 14). The molecular mechanism by which Wnt regulates β-catenin is not yet fully understood. Here we show that casein kinase I (CKI)ɛ is an important regulator of β-catenin in the Wnt pathway and is a component of these complexes. CKIɛ mimicked Wnt in inducing a secondary axis in Xenopus, stabilizing β-catenin, and stimulating β-catenin-dependent gene transcription. Inhibition of endogenous CKIɛ by the kinase-defective form of CKIɛ (KN-CKIɛ) or antisense-oligonucleotide attenuated gene transcription stimulated by Wnt. CKIɛ was found in a complex with axin and other downstream components of the Wnt pathway, including Dvl. We propose that CKIɛ is a positive regulator of the Wnt pathway and is a possible functional link between upstream signals and the intracellular axin signaling complex that regulates β-catenin.
Materials and Methods
Plasmids.
Human CKIδ cDNA was a gift from J. Kusuda (National Institute of Infectious Diseases, Tokyo). Human CKIα cDNA was isolated by PCR. Lysine-38 in mouse CKIɛ was mutated to phenylalanine to make KN-CKIɛ as described (15). CKIɛ, KN-CKIɛ, and ΔC-CKIɛ were constructed in pCS2+ vector (16). CKIδ and CKIα were constructed in pcDNA3.1 vector (Invitrogen). Myc-tagged Axin construct has been described (10). Myc-taggd Dv13 was from D. Yan (Chiron).
Library Screening.
E14 mouse embryonic cDNA library [oligo(dT)-primed] was constructed in pCS2+ vector (16). Pools of RNA derived from the library were injected into the four-cell stage of the Xenopus embryos at the ventral side. A total of 6 × 105 independent clones were screened. Each pool for injection contains 25–50 clones.
Xenopus Experiments.
mRNAs were synthesized by using a mMESSAGE mMACHINE kit (Ambion, Austin, TX). The RNA samples (1–5 ng) were injected into the ventral side of the four-cell stage blastomeres. Embryos with secondary axis structure were counted 48–72 h after injection. β-catenin or XWnt-8 RNA was injected as a positive control (17, 18).
Reverse Transcription–PCR (RT-PCR).
mRNA for RT-PCR was prepared from ventral halves of the Xenopus embryos at stage 10–10 1/2. RT-PCRs and primers were as described (19).
Cell Culture, Immunoprecipitation, and Western Blotting.
S2 stable cell lines were generated by transfecting CKIɛ and sgg under the control of methallothionein promoter with pMK33 vector that contains hygromycin-resistant gene for selection marker. S2 cells were lysed 24 h after induction by CuSO4. Transfection of 293 cells and immunoprecipitation was performed as described (10). Cytosolic fraction of 293 cells were prepared from supernatant by ultracentrifugation (100,000 g × 30 min) after lysis in hypotonic buffer.
Antisense Oligonucleotide Transfection.
Antisense oligonucleotides against human CKIɛ (CK-ASa; 5′-gcggcagaagttgaggtatgttgag-3′, CK-ASb; 5′-cgtaggtaagagtagtcgggcttgt-3′) or control oligonucleotide (5′-cgccgtcttcaactccatacaactc-3′) (final concentration 100 nM) were transfected into 293 cells by using cationic peptoid reagents (20) followed by transfection with Lef-1, Lef-1 reporter, and Wnt-1 plasmids using Lipofectamine (Life Technologies, Grand Island, NY).
Results and Discussion
To find additional regulators in the Wnt pathway, we used a screen for molecules that could mimic the developmental effects of Wnt. In Xenopus embryos, ectopic expression of Wnt elicits formation of a secondary axis (21). We injected pools of RNA derived from a mouse embryonic cDNA library into the ventral side of Xenopus embryos and searched for a gene that caused secondary axis formation. In this screen, we isolated several clones including β-catenin and Wnt-1 that have been shown previously to induce a secondary axis, validating the efficacy of this approach in discovering genes in the Wnt pathway. We also isolated a full-length cDNA for mouse CKI, which is 98.8% identical to the human CKIɛ isoform. The CKI gene family consists of seven different genes in mammals, CKIα, β, γ1, γ2, γ3, δ, and ɛ (15, 22). CKIɛ and δ isoforms, the most closely related, share 98% identity in the kinase domain and are 53% identical in a C-terminal domain that is not present in other CKI isoforms. This C-terminal domain appears to negatively regulate kinase activity (15, 23). CKIα is 74% identical to CKIɛ in the kinase domain and has no C-terminal extension. We showed that the ventral injection of CKIɛ RNA induced a secondary axis in Xenopus embryos (Fig. 1 a and b). CKIδ, like CKIɛ but not CKIα, induced a secondary axis when injected at the ventral side of the embryos (Fig. 1b). A point mutant of CKIɛ that was defective in kinase activity (KN-CKIɛ) did not induce a secondary axis (Fig. 1b). These data suggested that axis-inducing activity is specific for the CKIɛ/δ isoform and depends on its kinase activity. Because the C-terminal domain is unique for CKIɛ/δ isoforms, we tested whether deletion of this domain altered activity. When the truncated CKIɛ (ΔC-CKIɛ) RNA was injected into Xenopus embryos, we did not see any effects on axis formation (Fig. 1b). Both CKIα and ΔC-CKIɛ had kinase activity in vitro comparable to or greater than that of wild-type CKIɛ when they were expressed in mammalian cells (on a per-cell basis assessed by kinase assays of CKIɛ immunoprecipitates) or _in vitro_-translated (data not shown) even though they were not effective in inducing a secondary axis. Therefore, CKI kinase activity was not sufficient for mimicking Wnt. The requirement for the C-terminal domain of CKIɛ suggested that this part of the molecule is involved in linking CKIɛ to the Wnt pathway. Indeed further studies (see below) confirmed that this domain is important for the interaction of CKIɛ with a signaling complex.
Figure 1.

CKIɛ/δ induces a secondary axis in Xenopus embryos. (a) Examples of the embryos injected with CKIɛ or β-galactosidase (β-gal) RNA. (b) Percentage of embryos with duplicated axis injected with β-gal, β-catenin, CKIɛ, CKIδ, CKIα, KN-CKIɛ, or ΔC-CKIɛ RNA as indicated. The number of the embryos with duplicated axis relative to the total number of injected embryos is indicated above each bar. (c) Axin inhibits secondary axis formation induced by CKIɛ. CKIɛ RNA was coinjected with β-gal or axin RNA (1–2 ng). (d) Induction of Siamois by CKIɛ. Siamois expression was analyzed by RT-PCR from dorsal halves of the embryos injected with XWnt-8, CKIɛ, or β-gal. EF-1 expression was a loading control.
To demonstrate that CKIɛ activates Wnt signaling, we coinjected CKIɛ with axin, which is a known inhibitor of the Wnt pathway that acts by linking β-catenin to GSK-3β (6–10, 17). Axin inhibited the induction of a secondary axis by CKIɛ (Fig. 1c). This finding suggests that the CKIɛ effect is mediated through β-catenin in a manner analogous to the effects of Wnt. One of the downstream target genes of β-catenin in Xenopus is Siamois (18, 19). Siamois is a homeobox gene induced by Wnt and responsible for its dorsalizing activity. CKIɛ overexpression at the ventral side of Xenopus embryo also induced Siamois expression detected by RT-PCR (Fig. 1d and ref. 19). Our observations in the Xenopus experiments that CKIɛ mimicked Wnt, both in its gene regulation and developmental effects, suggested that CKIɛ might be a component of the Wnt pathway.
To understand the mechanism by which CKIɛ activates the Wnt pathway, we examined the effect of CKIɛ on β-catenin protein level. Wnt-1 stabilizes cytosolic β-catenin protein by suppressing GSK-3β (24, 25). We made Drosophila Schneider S2 cell lines that stably expressed CKIɛ controlled by a metallothionein promoter. Overexpression of CKIɛ caused accumulation of endogenous armadillo (arm) protein, the Drosophila homologue of β-catenin (Fig. 2a). We also showed that β-catenin protein level was increased by transiently overexpressing CKIɛ in 293 cells (Fig. 2b). These results suggest that CKIɛ activates the Wnt pathway by stabilizing β-catenin.
Figure 2.
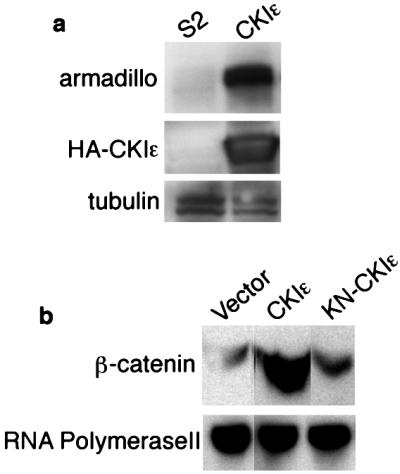
β-catenin stabilization induced by CKIɛ. (a) Drosophila S2 Schneider cell lysates blotted with armadillo antibody and hemagglutinin (HA) antibody recognized transfected CKIɛ. Tubulin was a loading control. (b) Cytosolic fraction from 293 cells transfected with vector, Wnt-1, CKIɛ, and KN-CKIɛ blotted with β-catenin antibody. RNA polymerase II was a loading control.
To further study the role of CKIɛ in the Wnt pathway, we measured Lef-1 reporter gene activity in mammalian cells. The transcription factor Lef-1/Tcf forms a complex with β-catenin in response to Wnt stimulation (11, 12). When expressed in 293 cells, Wnt-1 stimulated the expression of a Lef-1 reporter gene transcription 4- to 6-fold over vector transfected cells (10). CKIɛ and CKIδ activated the Lef-1 reporter gene about 10-fold (Fig. 3a). However CKIα and ΔC-CKIɛ did not activate the Lef-1 reporter gene (Fig. 3a), consistent with the Xenopus injection experiments (Fig. 1). Coexpressing axin inhibited the Lef-1 reporter activation induced by CKIɛ (Fig. 3b). These data confirm the results from Xenopus experiments, suggesting that CKIɛ activates the Wnt pathway through an effect on the β-catenin-Lef-1/Tcf complex.
Figure 3.

Lef-1 reporter gene activity induced by CKIɛ. Lef-1 reporter gene assay was performed as described (10). Representative data from several independent experiments are shown. (a) The effects of CKI isoforms on Lef-1 activity. (b) Axin inhibits Lef-1 reporter gene activity induced by CKIɛ. (c) KN-CKIɛ blocks Lef-1 activity induced by Wnt-1. (d) CKIɛ antisense-oligonucleotides inhibit Lef-1 reporter gene activity induced by Wnt-1. (Left) Wnt-1 induced Lef-1 reporter activity. CKI-ASa and CKI-ASb are two different antisense-oligonucleotides from the human CKIɛ coding sequence. The control-oligonucleotide is the reverse sequence of ASa. (Right) Endogenous level of CKIɛ normalized to levels of 14-3-3 protein (loading control) after transfection of oligonucleotides.
KN-CKIɛ inhibited the activation of Lef-1 reporter by Wnt-1 (Fig. 3c), suggesting the involvement of endogenous CKIɛ during the Wnt signal. These data suggest that KN-CKIɛ acts as a dominant negative kinase and blocks the upstream signal coming from Wnt-1. To further assess the physiological importance of CKIɛ in the Wnt pathway, we used antisense-oligonucleotides to reduce the endogenous CKIɛ protein level, which resulted in the inhibition of Lef-1 reporter activity induced by Wnt-1 (Fig. 3d). Taken together the Xenopus, Drosophila, and mammalian cell experiments showed that CKIɛ activates the Wnt pathway and appears to be a significant positive regulatory component that is required for a full Wnt effect.
Some of the downstream molecules in the Wnt pathway have been shown to form complexes containing negative regulators (GSK-3β, axin, and adenomatous polyposis coli tumor suppressor protein) and a positive regulator (β-catenin) in vivo (6–10). We examined the possibility that CKIɛ is also in a complex with these molecules. We found that axin, a negative regulator of the Wnt pathway, bound to CKIɛ. Endogenous CKIɛ was coimmunoprecipitated with overexpressed axin (Fig. 4a). Binding of axin to ΔC-CKIɛ and ΔC-KN-CKIɛ was much reduced compared to wild-type CKIɛ and KN-CKIɛ (Fig. 4b). These results suggest that the C-terminal domain of CKIɛ is important for its interaction with axin, which may be the reason that ΔC-CKIɛ and CKIα did not activate the Wnt pathway in Xenopus or mammalian cells (Figs. 1 and 3). To further study the complex of CKIɛ with axin and GSK-3β, we showed that GSK-3β coimmunoprecipitated with CKIɛ, but much less with ΔC-CKIɛ, and only in the presence of axin (Fig. 4c). This finding suggests that CKIɛ is a positive regulatory molecule in the Wnt pathway and its interaction through its C terminus with the axin-GSK-3β complex is likely to be important for its activity. Furthermore, we also detected endogenous CKIɛ in the complex with overexpressed Dvl3 (Fig. 4d), an upstream molecule of the axin-GSK-3β complex.
Figure 4.

CKIɛ forms a complex with the other molecules in the Wnt pathway. (a) Endogenous CKIɛ coimmunoprecipitated with transfected myc-tagged axin. (b) The C-terminus domain of CKIɛ is required for binding to axin. Myc-axin and hemagglutinin (HA)-CKIɛ constructs (indicated by arrows) were cotransfected in 293 cells. Myc-axin immune complexes were analyzed by immunoblotting with myc (detect axin) and HA (detect CKIɛ) antibodies. Dye front was marked as ★. (c) GSK-3β is in a complex with CKIɛ and axin. Myc-axin and HA-CKIɛ constructs (indicated) were cotransfected in 293 cells. HA-CKIɛ immune complexes were analyzed by immunoblotting with myc (detect axin) and GSK-3β antibodies. (d) Endogenous CKIɛ is coimmunoprecipitated with transfected myc-tagged Dvl3.
In this study, we cloned a CKIɛ gene by using the Xenopus system as an indicator of Wnt-inducing effects. Recently a CKIɛ gene of Drosophila was identified as the clock gene (26). Homozygous mutation in the Drosophila CKIɛ gene produces embryonic lethality, which also suggests the involvement of CKIɛ in an early process that is probably unrelated to its “clock” function. It will be interesting to examine in detail the embryonic phenotype of CKIɛ-defective flies related to pathway genes.
In summary we report a signaling molecule in the Wnt pathway, CKIɛ, that induces secondary axis formation in Xenopus, Lef-1-dependent transcription, and stabilization of β-catenin in both fly and mammalian cells. A critical role of CKIɛ in Wnt signaling is supported by several observations: inhibition of endogenous CKIɛ by KN-CKIɛ or by antisense-oligonucleotides blocked Wnt-1 effects; endogenous CKIɛ is present in a complex with axin, GSK-3β, and Dvl, known components of the Wnt pathway; and the C-terminal domain of CKIɛ is required for its interaction with the axin complex and for the ability of CKIɛ to mimic Wnt. It will be important to determine the substrates of CKIɛ in this pathway and the upstream signals between Wnt receptors and CKIɛ. It is also possible that stimuli other than Wnt regulate the ability of CKIɛ to impinge on this pathway.
Acknowledgments
We thank J. Martin for his role in performing the Xenopus expression cloning and constructing plasmids, M. Wu for Xenopus injection assay, C. Reinhardt and M. Del Rosario for the advice in antisense-oligonucleotides technologies, R. Zuckermann for cationic peptoid reagents, and W. Fantl for reading the manuscript. This work was partially supported by the Howard Hughes Medical Institute and Bristol-Myers Squibb Company (C.S.).
Abbreviations
CKI
casein kinase I
GSK-3β
glycogen synthase kinase-3β
Lef-1
lymphoid enhancer factor-1
Tcf
T cell factor
RT-PCR
reverse transcription–PCR
KN-CKIɛ
kinase-defective form of CKIɛ
References
- 1.Cadigan K M, Nusse R. Genes Dev. 1997;11:3286–3305. doi: 10.1101/gad.11.24.3286. [DOI] [PubMed] [Google Scholar]
- 2.Gumbiner B M. Curr Biol. 1997;7:R443–R446. doi: 10.1016/s0960-9822(06)00214-4. [DOI] [PubMed] [Google Scholar]
- 3.Willert K, Nusse R. Curr Opin Genet Dev. 1998;8:95–102. doi: 10.1016/s0959-437x(98)80068-3. [DOI] [PubMed] [Google Scholar]
- 4.Klingensmith J, Nusse R. Dev Biol. 1994;166:396–414. doi: 10.1006/dbio.1994.1325. [DOI] [PubMed] [Google Scholar]
- 5.Perrimon N. Cell. 1994;76:781–784. doi: 10.1016/0092-8674(94)90351-4. [DOI] [PubMed] [Google Scholar]
- 6.Behrens J, Jerchow B A, Wurtele M, Grimm J, Asbrand C, Wirtz R, Kuhl M, Wedlich D, Birchmeier W. Science. 1998;280:596–599. doi: 10.1126/science.280.5363.596. [DOI] [PubMed] [Google Scholar]
- 7.Ikeda S, Kishida S, Yamamoto H, Murai H, Koyama S, Kikuchi A. EMBO J. 1998;17:1371–1384. doi: 10.1093/emboj/17.5.1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Itoh K, Krupnik V E, Sokol S Y. Curr Biol. 1998;8:591–594. doi: 10.1016/s0960-9822(98)70229-5. [DOI] [PubMed] [Google Scholar]
- 9.Hart M J, de los Santos R, Albert I N, Rubinfeld B, Polakis P. Curr Biol. 1998;8:573–581. doi: 10.1016/s0960-9822(98)70226-x. [DOI] [PubMed] [Google Scholar]
- 10.Sakanaka C, Weiss J B, Williams L T. Proc Natl Acad Sci USA. 1998;95:3020–3023. doi: 10.1073/pnas.95.6.3020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Behrens J, von Kries J P, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Nature (London) 1996;382:638–642. doi: 10.1038/382638a0. [DOI] [PubMed] [Google Scholar]
- 12.Molenaar M, van de Wetering M, Oosterwegel M, Peterson-Maduro J, Godsave S, Korinek V, Roose J, Destree O, Clevers H. Cell. 1996;86:391–399. doi: 10.1016/s0092-8674(00)80112-9. [DOI] [PubMed] [Google Scholar]
- 13.Morin P J, Sparks A B, Korinek V, Barker N, Clevers H, Vogelstein B, Kinzler K W. Science. 1997;275:1787–1790. doi: 10.1126/science.275.5307.1787. [DOI] [PubMed] [Google Scholar]
- 14.Rubinfeld B, Robbins P, El-Gamil M, Albert I, Porfiri E, Polakis P. Science. 1997;275:1790–1792. doi: 10.1126/science.275.5307.1790. [DOI] [PubMed] [Google Scholar]
- 15.Fish K J, Cegielska A, Getman M E, Landes G M, Virshup D M. J Biol Chem. 1995;270:14875–14883. doi: 10.1074/jbc.270.25.14875. [DOI] [PubMed] [Google Scholar]
- 16.Rupp R A, Snider L, Weintraub H. Genes Dev. 1994;8:1311–1323. doi: 10.1101/gad.8.11.1311. [DOI] [PubMed] [Google Scholar]
- 17.Zeng L, Fagotto F, Zhang T, Hsu W, Vasicek T J, Perry W L, 3rd, Lee J J, Tilghman S M, Gumbiner B M, Costantini F. Cell. 1997;90:181–192. doi: 10.1016/s0092-8674(00)80324-4. [DOI] [PubMed] [Google Scholar]
- 18.Fan M J, Gruning W, Walz G, Sokol S Y. Proc Natl Acad Sci USA. 1998;95:5626–5631. doi: 10.1073/pnas.95.10.5626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fagotto F, Guger K, Gumbiner B M. Development (Cambridge, UK) 1997;124:453–460. doi: 10.1242/dev.124.2.453. [DOI] [PubMed] [Google Scholar]
- 20.Murphy J E, Uno T, Hamer J D, Cohen F E, Dwarki V, Zuckermann R N. Proc Natl Acad Sci USA. 1998;95:1517–1522. doi: 10.1073/pnas.95.4.1517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Moon R T, Brown J D, Torres M. Trends Genet. 1997;13:157–162. doi: 10.1016/s0168-9525(97)01093-7. [DOI] [PubMed] [Google Scholar]
- 22.Gross S D, Anderson R A. Cell Signal. 1998;10:699–711. doi: 10.1016/s0898-6568(98)00042-4. [DOI] [PubMed] [Google Scholar]
- 23.Cegielska A, Gietzen K F, Rivers A, Virshup D M. J Biol Chem. 1998;273:1357–1364. doi: 10.1074/jbc.273.3.1357. [DOI] [PubMed] [Google Scholar]
- 24.Hinck L, Nelson W J, Papkoff J. J Cell Biol. 1994;124:729–741. doi: 10.1083/jcb.124.5.729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.van Leeuwen F, Samos C H, Nusse R. Nature (London) 1994;368:342–344. doi: 10.1038/368342a0. [DOI] [PubMed] [Google Scholar]
- 26.Kloss B, Price J L, Saez L, Blau J, Rothenfluh A, Wesley C S, Young M W. Cell. 1998;94:97–107. doi: 10.1016/s0092-8674(00)81225-8. [DOI] [PubMed] [Google Scholar]