Plasmacytoid Dendritic Cells Mediate Oral Tolerance (original) (raw)
. Author manuscript; available in PMC: 2013 Jan 15.
SUMMARY
Oral tolerance prevents oral sensitization to dietary antigens (Ags), including proteins and haptens, and development of delayed-type hypersensitivity (DTH) responses. We showed here that plasmacytoid dendritic cells (pDCs) prevented oral T cell priming and were responsible for systemic tolerance to CD4+ and CD8+ T cell-mediated DTH responses induced by Ag feeding. Systemic depletion of pDCs prevented induction of tolerance by antigen feeding. Transfer of oral Ag-loaded liver pDCs to naive recipient mice induced Ag-specific suppression of CD4+ and CD8+ T cell responses to protein and hapten, respectively. Liver is a site of oral Ag presentation, and pDCs appeared to induce anergy or deletion of Ag-specific T cells in the liver relatively rapidly via a CD4+ T cell-independent mechanism. These data demonstrate that oral tolerance relies on Ag presentation by pDC to T cells and suggest that pDC could represent a key therapeutic target for intestinal and systemic inflammatory diseases.
INTRODUCTION
Oral tolerance is the physiologic mechanism by which the mucosal immune system prevents deleterious T cell-mediated delayed-type hypersensitivity (DTH) reactions to self and exogenous dietary and environmental antigens (Ag) including food proteins as well as small chemical molecules also known as haptens (i.e., drugs, dyes, conservatives, and flavor enhancers) (Dubois et al., 2005; Mowat, 2003). Studies in animal models have documented that Ag feeding, prior to systemic immunization with the same Ag, prevents the development of specific T cell-mediated immunopathologies (Faria and Weiner, 2005) and CD4+ and CD8+ T cell-mediated DTH responses to proteins (Mowat, 2003) and haptens (Dubois et al., 2003), respectively. Clinical studies have confirmed that systemic T cell tolerance can be induced by Ag feeding in humans and primarily affects T cell-mediated DTH responses but not B cell responses (Husby et al., 1994). Along these lines, breakdown of oral tolerance to dietary Ag or to the gut flora can cause chronic intestinal inflammatory diseases, as illustrated by celiac disease and inflammatory bowel disease (IBD), respectively. Although CD4+ T cells have always been thought to contribute to intestinal inflammation, emerging data indicate that in both celiac disease and IBD, CD8+ T cells mediating DTH might initiate inflammation by exerting cytotoxicity against epithelial cells (Hue et al., 2004; Nancey et al., 2006).
Several mechanisms contribute to oral tolerance including anergy or deletion of antigen-specific T cells and activation of regulatory CD4+ T cells that encompass natural CD4+CD25+Foxp3+ and antigen-specific regulatory T (Treg) cells producing the cytokines TGF-β and/or IL-10 (Dubois et al., 2005). Although dendritic cells (DC) likely play a role in oral tolerance induction (Viney et al., 1998), there is as yet no direct evidence of the implication of myeloid or other types of DC, and the sites of oral Ag presentation by DC remain elusive. Although orally administered Ag can be presented to T cells by DC in both Peyer’s patches (PP) and mesenteric lymph nodes (MLN) (Kunkel et al., 2003), oral tolerance requires MLN (Spahn et al., 2001; Worbs et al., 2006) but not PP (Spahn et al., 2001). Oral Ag can be captured by DCs in the intestinal lamina propria (LP) and transported to MLN via the mesenteric lymph (Mowat, 2003). The recent observation that myeloid DC of both MLN and LP can convert CD4+ T cells into Foxp3+ Treg in vitro (Coombes et al., 2007; Sun et al., 2007), or induce IL-10-producing CD4+ Treg cells (Bilsborough et al., 2003) might explain their suspected role in oral tolerance (Viney et al., 1998). However, oral tolerance also involves systemic diffusion of free Ag via blood. Indeed, a variable fraction of food protein entering blood vessels in the lamina propria reaches the liver via the portal vein and disseminates systemically, intact or in the form of polypeptides, in peripheral lymphoid organs (Swarbrick et al., 1979). Direct evidence for a role of the liver in tolerance is provided by the observation that portocaval anastomosis, which prevents draining into the liver of Ags from the gut, abrogates oral tolerance to proteins or haptens (Cantor and Dumont, 1967; Yang et al., 1994). Along these lines, it has been shown that T cell tolerance in the liver and in the spleen can occur within hours after Ag feeding (Gutgemann et al., 1998; Ju and Pohl, 2001; Li et al., 2004), suggesting that Ag presentation by antigen-presenting cells (APCs) outside the gut-associated lymphoid tissues (GALT) can be involved in oral tolerance.
Plasmacytoid DC (pDC) represent an unconventional type of APC that is typified by their ability to contribute to antiviral innate immunity by producing type I interferon (IFN) upon activation (Asselin-Paturel et al., 2001). They also participate in adaptive immune responses by directing either T helper 1 (Th1) cell orTh2 cell development (Boonstra et al., 2003). However, pDC at the immature state have a poor capacity to support T cell proliferation (Asselin-Paturel et al., 2001) and can even suppress T cell responses (Bilsborough et al., 2003; Martin et al., 2002). Indeed, pDC contribute to peripheral T cell tolerance including transplantation tolerance (Ochando et al., 2006), tumor escape (Munn et al., 2004), and mucosal tolerance in an animal model of asthma (De Heer et al., 2004). However, whether and how pDC contribute to oral tolerance has not been previously studied.
The goal of this study was to identify the type and location of DC that could play an essential role in oral tolerance and could achieve efficient suppression of CD4+ and CD8+ T cell-mediated DTH responses. To monitor oral tolerance of CD8+ T cell-mediated DTH responses to a soluble Ag, we used a model of hapten-specific skin DTH, induced by CD8+ CTL independently of CD4+ T cell help (Desvignes et al., 2000). In this model, covalent binding of the hapten to proteins generates hapten-modified self peptides, which are presented by MHC class I molecules and induce priming of IFNγ-producing CD8+ CTL effectors mediating contact dermatitis (Dubois et al., 2003; Kehren et al., 1999) or colitis (Nancey et al., 2006). We also analyzed the induction of oral tolerance in a classical model of DTH, mediated by specific CD4+ T cells, to a more conventional dietary protein, ovalbumine (OVA). We showed here that oral tolerance relies on pDCs, which suppress both CD4+ and CD8+ T cell-mediated DTH responses.
RESULTS
Oral Ag Is Rapidly Captured in the Liver and in Peripheral Lymphoid Organs
The localization and fate of oral Ag in normal mice was examined using 2.4-dinitrofluorobenzene (DNFB) as a model Ag, as its binding to self proteins allows in situ detection by staining with a DNP-specific mAb. Kinetic immunohistochemical analysis revealed that the hapten is detectable as early as 30 min after gavage in the gut epithelium, appears by 6 hr in the intestinal lamina propria, and disappears at 24 hr (Figure 1A). Because after local uptake the Ag was not identifiable by immunohistochemical analysis in lymphoid and nonlymphoid organs, we set up a functional ex vivo antigen presentation assay to determine the presence of Ag-presenting cells in different organs after intragastric administration of DNFB. To this aim, leukocytes were isolated from MLN, spleen, liver or different lymph nodes (LN) and used to stimulate the proliferation of CD8+ T cells from day 5 DNFB-sensitized mice. CD8+ T cell proliferation was induced by leukocytes isolated from all organs of DNFB-fed mice except the spleen (Figure 1B). Leukocytes from MLN and inguinal or axillary LN induced specific CD8+ T cell proliferation at 2 and 24 hr after DNFB gavage. However, liver and hepatic LN cells were able to induce proliferation only when isolated at 2 hr but not at 24 hr, suggesting that the hapten rapidly disappeared from these two sites or that Ag-loaded APC from liver and hepatic LN become unable to stimulate CD8+ T cell proliferation. These data illustrate that orally administered hapten rapidly diffuses to the liver and hepatic LN and also reaches systemic LN including MLN.
Figure 1. Organ Distribution of Orally Administered Hapten.

(A) Immunohistochemical analysis of DNP capture in the small intestine, at various time points after DNFB gavage (initial magnification 320). Cryostat section (5 μm) of small intestine were stained with a biotinylated DNP mAb and streptavidin peroxidase, and the reaction was developed using amino-ethyl carbazole.
(B) Peyer’s patches, spleen, MLN, liver, hepatic LN, or axillaries and inguinal LN were harvested at 2 hr after vehicle feeding (white bars) or 2 hr (hatched bars) and 24 hr (black bars) after DNFB gavage. Isolated leukocytes (105 cells/well) were cocultured for 3 days with CD8+ T cells (105 cells/well) from day 5 DNFB-sensitized mice. Proliferation was determined by 3H-thymidine uptake during the last 8 hr of culture. Results are expressed as mean cpm ± SD of triplicate wells containing pooled cells from four mice per group. Data are representative of one out of three independent experiments.
Ag Delivery by Oral or Intraportal Route Prevents CD8+ T Cell-Mediated DTH
We previously documented that oral administration of DNFB, prior to skin sensitization with the hapten, completely prevented the development of hapten-specific CD8+ T cells mediating contact hypersensitivity (CHS) (Desvignes et al., 2000; Dubois et al., 2003). To determine whether systemic diffusion of dietary antigen via the hepatic portal venous system could contribute to CD8+ T cell tolerance, we compared the CHS response in mice previously exposed to the Ag by gavage or by injection in the portal vein. Control mice injected with PBS in the portal vein developed an ear swelling response that peaked at 48 hr after DNFB challenge (Figure 2A) and correlated with the priming of hapten-specific interferon-γ (IFN-γ)-producing CD8+ T cells (Figure 2B). Oral gavage with DNFB or intraportal injection of DNBS (a soluble form of DNFB) were similarly efficient at inhibiting the DNFB-specific CHS response (Figure 2A) and prevented the differentiation of hapten-specific IFN-γ-producing CD8+ T cells (Figure 2B). Thus, oral gavage and immunization through the hepatic portal vein are equally efficient at inducing CD8+ T cell tolerance.
Figure 2. Hapten Delivery by Either Oral or Intraportal Routes Induces Tolerance of CD8+ T Cells Mediating CHS.
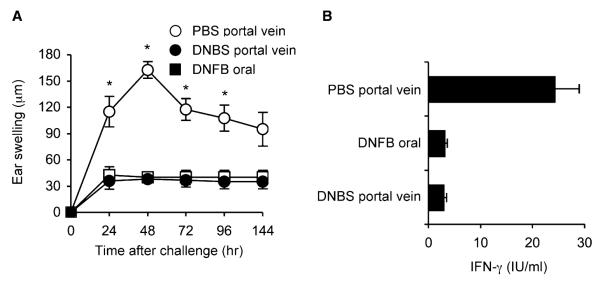
(A) Hapten-specific CHS. On day −7, mice were fed with 0.1% DNFB diluted in acetone and olive oil (1:10 v/v) (closed squares) or injected via the portal vein with 125 mg/kg DNBS diluted in PBS (closed circles) or with PBS alone (open circles). All mice were DNFB sensitized on day 0 and ear challenged on day 5, and CHS response was determined by ear swelling (μm) at various times after hapten challenge. Results are expressed as mean ear swelling (μm ± SD) for five to seven mice per group and are representative of one out of two independent experiments. *p < 0.01 PBS-injected mice compared to DNBS-injected mice.
(B) Hapten-specific CD8+ T cell response. Mice were treated and sensitized as in Figure 2A. Purified CD8+ T cells (2 × 105) harvested from inguinal and axillary lymph nodes on day 5 after skin sensitization were restimulated in vitro for 3 days with irradiated spleen cells (5 × 105) either untreated or pulsed with DNBS, and IFN-γ was titrated by ELISA in day 3 supernatants. Results are expressed as mean IFN-γ production (UI/ml) ± SD of triplicate wells containing pooled CD8+ T cells from four mice per group. Data are representative of one out of two independent experiments.
Plasmacytoid Dendritic Cells Are the Tolerogenic CD11c+ DC of the Liver
To analyze whether hepatic DC after oral administration of Ag had tolerogenic functions, CD11c+ cells were purified from the liver and tested for their capacity to induce or suppress hapten-specific T cell and CHS responses. CD11c+ cells account for as much as 15%–20% of CD45+ liver leukocytes (Figure 3A) and exhibit an immature phenotype with low expression of MHC class II, CD80, and CD86, and a low allostimulatory potential (data not shown), confirming previous data (Lu et al., 2001; Watanabe et al., 2003). Unlike in vitro-generated bone marrow-derived DCs (BMDCs), liver CD11c+ cells pulsed in vitro with DNBS were unable to induce a CHS response when injected subcutaneously (s.c.) into naive mice (Figure 3B). Moreover, unpulsed liver CD11c+ DCs delivered at the site and time of DNFB skin sensitization completely prevented induction of the CHS response (Figure 3C), demonstrating that liver DCs exert an inhibitory effect.
Figure 3. Liver Plasmacytoid DC Are Tolerogenic In Vivo.

(A) FACS analysis and sorting of hepatic DC subsets. Liver leukocytes were stained with anti-CD11c, anti-CD11b and anti-NK1.1. Gated CD11c+ cells (left dot plot) were analyzed for CD11b and NK1.1 expression (right dot plot) and were FAC-sorted into three subsets: CD11c+CD11b+or−NK1.1+ (R1), CD11c+CD11b+NK1.1− (R2), and CD11c+CD11b−NK1.1− (R3).
(B) Liver CD11c+ cells are unable to prime a CHS response to DNFB. Mice were immunized on day 0 by s.c. transfer of 105 BM-DC (open squares) or MAC-sorted CD11c+ liver cells (closed squares) pulsed in vitro with DNBS. CHS was determined by ear swelling (μm ± SD of five to seven mice per group) after DNFB challenge on day 5. *p < 0.01.
(C) Liver CD11c+ cells inhibit CHS to DNFB. Unpulsed (105) liver CD11c+ cells (closed circles) or PBS (open circles) were injected subcutaneously in naive mice at the time and site of skin painting with DNFB. CHS was determined by ear swelling (μm ± SD of five mice per group) after DNFB challenge 5 days later. *p < 0.05.
(D–F) In vivo adoptive transfer experiments. Each subset of R1 (closed squares), R2 (open circles), and R3 (closed circles) sorted CD11c+ liver cells shown in (C)was transferred s.c. (105 cells/mouse) into naive mice at the site and time of skin sensitization with DNFB (day 0). Untransferred but DNFB skin-sensitized mice (open squares) were used as controls. In (D) and (F), all mice were ear challenged on day 5, and CHS to DNFB was determined by ear swelling (μm ± SD of five mice per group) at (D) various times or (F) 24 hr after DNFB challenge. *p < 0.05 between the untransferred group and the R3-transferred group. In (E), at day 5, the frequency of hapten-specific IFN-γ-spot-forming cells (SFC) in draining lymph nodes was determined by ELISPOT. Results are expressed as the mean number of SFC per 106 CD8+ T cells ± SD of triplicate wells containing pooled CD8+ T cells from four mice per group. In (E), right panel, and (F), in an independent experiment, CD11c+CD11b−NK1.1− liver leukocytes were further sorted into 120G8+ pDC and 120G8− cells and tested for their capacity to suppress priming of (E) hapten-specific IFN-γ SFC or a (F) CHS response. Each data set is representative of two (D–F) to four (B and C) independent experiments.
CD11c+ liver cells contained around 50% of NK1.1+ cells that likely correspond to NK cells expressing CD11c as well as 40% of CD11b− NK1.1− cells and 10% of CD11b+ NK1.1− cells (Figure 3A), both of which mostly contained DCs (Figure S1A). These three populations of CD11c+ cells sorted on the basis of CD11b and NK1.1 expression (Figure 3A) were compared for their relative ability to inhibit the CHS response upon s.c. transfer at the time of skin sensitization. The CHS response was not affected by transfer of either CD11c+CD11b+NK1.1−- or CD11c+NK1.1+ cell subsets. In contrast, transfer of CD11c+CD11b−NK1.1− cells conferred almost complete inhibition of the CHS response (Figure 3D) and prevented priming of hapten-specific IFN-γ-producing CD8+ T cells in skin draining LN (Figure 3E).
Interestingly, 60%–80% of CD11c+CD11b−NK1.1− liver cells expressed the plasmacytoid DC marker BST-2, as revealed by staining with the 120G8 mAb (Figure S1B) (Asselin-Paturel et al., 2003). Indeed, as described for pDC of lymphoid organs (Asselin-Paturel et al., 2001, 2003; Blasius et al., 2006), liver CD11c+120G8+ cells expressed B220, Ly6C, BST-2, Siglec-H, and varying levels of CD8α (Figures S1B, S1C, and S3C) and produced type I IFN upon in vitro culture with influenza virus (data not shown). pDC represented up to 30% of total CD11c+ liver cells and were contained within the CD11c+CD11b−NK1.1− subpopulation of liver DC (Figure S1C). We thus examined whether pDC accounted for the capacity of liver CD11c+ DC to induce hapten-specific T cell tolerance. CD11c+NK1.1−CD11b− leukocytes were sorted into 120G8+ (pDC) and 120G8− cell subsets and injected s.c. into naive C57Bl/6 mice at the time of skin sensitization with DNFB. Adoptive transfer of FAC-sorted 120G8− cells did not affect the frequency of hapten-specific IFN-γ-producing CD8+ T cells (Figure 3E), which are effector cells of CHS (Kehren et al., 1999). In contrast, sorted 120G8+ pDC transfer decreased by 60% the frequency of hapten-specific IFN-γ-producing CD8+ T cells (Figure 3E) and completely prevented the CHS response (Figure 3F). Thus, pDC were immunoregulatory and accounted for the tolerogenic property of liver DC.
pDC Are Required for Oral Tolerance of CD8+ and CD4+ T Cell-Mediated DTH Responses
To directly address the role of pDC in oral tolerance, pDC were depleted in vivo using the RB6-8C5 anti-Gr1 mAb. Anti-Gr1 efficiently depleted ~80% of liver pDC as assessed both by evaluating the remaining frequency of liver pDC by flow cytometry and by measuring the loss of ability of MAC-sorted CD11c+ liver DC to produce IFN-α in response to virus stimulation, a functional property of pDC (Figures S2A and S2B). Mice injected with the control rat mAb GL113 were tolerized by oral administration of the hapten, whereas oral tolerance was impaired in Gr1 mAb injected mice, as demonstrated by restoration of CHS (Figure 4A) and increased IFN-g production by CD8+ T cells (Figure 4B). Because the Gr1 mAb in addition to pDC also depleted neutrophils, eosinophils, and other Gr1+ cells, we tested whether tolerance could be reversed by injection of another mAb targeting pDC, i.e., the 120G8 mAb (Asselin-Paturel et al., 2003). Treatment with 120G8 efficiently depleted pDC from the liver (Figure S2C), MLN, and spleen (data not shown), as assessed by flow cytometry with anti-BST-2 or anti-SiglecH mAbs. Tolerance to DNFB could not be induced in 120G8-treated mice, as demonstrated by the similar intensity of the CHS (Figure 4C) and CD8+ T cell responses (Figure 4D) in both vehicle-fed and hapten-fed mice. Furthermore, systemic sensitization could be induced by a single DNFB gavage (without skin painting) in 120G8 mAb-treated mice, as evidenced by the development of a CHS response upon ear challenge 5 days later (Figure 4E). Thus, pDC are required for induction of CD8+ T cell tolerance by oral hapten delivery and their absence permits oral sensitization and development of a CHS response.
Figure 4. Antibody Depletion of pDC Prevents Induction of Oral Tolerance and Allows for Oral Priming.

(A–D) Mice were injected with either anti-Gr1 RB8.6C5 mAb or anti-BST2 120G8 mAb to deplete pDC, or with the control rat GL113 mAb, and tested for induction of oral tolerance as monitored by the CHS (A and C) and CD8+ T cell responses to DNFB (B and D), as described in the Experimental Procedures and in the legend of Figure 2. In (A), *p < 0.01 between control mAb- and anti-Gr1-treated DNFB-fed mice. In (B), *** p < 0.0001; ** p = 0.0071. Data in (A)–(D) are representative of three reproducible experiments using five to seven mice per group.
(E) To test for oral priming, mice were fed with DNFB or vehicle and ear challenged 5 days later, and the ear swelling response was determined after 48 hr (one representative experiment out of two).
To confirm whether pDC were also required for oral tolerance to a more conventional dietary protein (ovalbumine, OVA), we used the classical model of DTH to OVA, mediated by specific CD4+ T cells. We showed that intragastric delivery of 20 mg of OVA 3 times on days −13, −10, and −7 (protocol depicted on Figure 5A) nearly completely prevented the DTH response to OVA (80% of suppression of footpad swelling; Figure 5B). In vivo depletion of pDC by 120G8 mAb treatment of OVA-fed mice, prevented suppression of footpad swelling and of IFN-γ production by OVA-specific T cells, by roughly 50% (Figures 5B and 5C). Similar results were obtained in mice fed from day − 14 to −7 with 20 mg/ml OVA in drinking water (data not shown). These data demonstrate that pDC also inhibit DTH responses induced by gavage with a protein Ag.
Figure 5. Antibody-Mediated Depletion of pDC Affects Oral Tolerance to Protein Antigens.

(A) Mice were injected or not with 120G8 mAb and were fed three times either with PBS or 20 mg of OVA as depicted in the flow charts, where # indicates the days of 120G8 mAb injections. Seven days after the last gavage, all mice were immunized by subcutaneous injection of OVA in PBS and CFA emulsion and challenged 1 week later with aggregated OVA injected in the footpad.
(B and C) Tolerance induction was analyzed by measurement of the DTH reaction 24 hr after challenge (mean ear swelling ± SD of five mice per group) and (C) OVA-specific T cell IFN-γ production by spleen cells collected 48 hr after challenge and restimulated in vitro with 1 mg/ml OVA (mean UI/ml ± of triplicate wells). *** p = 0.001; ** p = 0.0018.
Comparison of the Tolerogenic Potential of pDC from Liver, MLN, and Spleen of Ag-Fed Mice
We next examined the efficacy of tolerance induction by pDC from Ag-fed mice and asked whether it depended on their tissue origin. Graded numbers of MAC-sorted liver pDC, isolated from DNFB- or vehicle-fed mice (Figure S3A), were transferred into B6 recipient mice at the time of immunization with hapten-pulsed BM-DC and tested for their ability to inhibit CD8+ T cell-mediated CHS responses. pDC enriched from liver leukocytes of vehiclefed mice had a modest inhibitory effect only at the highest dose (105). In contrast, a dose-dependent suppression of the ear swelling response was achieved by transfer of pDC-enriched leukocytes from DNFB-fed mice, with significant inhibition with as few as 104 cells (Figure 6A). Importantly, transfer of 105 FAC-sorted pDC (which contained >97% pure pDC; Figures S3B and S3C) from liver or MLN cells of hapten-fed mice were similarly efficient to prevent in vivo priming of hapten-specific IFN-γ-producing CHS effectors, whereas T cell priming was only moderately affected by transfer of splenic pDC either purified from vehicle- or DNFB-fed mice (Figure 6B). In addition, pDC purified from the liver, but not spleen, efficiently suppressed the proliferation of hapten-specific CD8+ T cells in vitro (Figure S4A). Thus, pDC from the liver and MLN, but not spleen, of hapten-fed mice can induce T cell tolerance and suppression of DTH responses in vivo.
Figure 6. pDC from Liver or MLN of Ag-Fed mice Efficiently Suppress DTH Responses in an Antigen-Specific Fashion.

(A) DNBS-pulsed BMDCs (105) were transferred s.c. into naive mice in the presence or absence of graded numbers of pDCs (102–105) purified by MACS from the liver of vehicle- or DNFB-fed mice. All mice were ear challenged with DNFB at day 5, and CHS to DNFB was determined by ear swelling 24 hr later. Data are expressed as mean ear swelling (μm ± SD) and are representative of one out of two experiments with four mice per group. ***p < 0.0005 between recipients of 105 or 104 liver pDC from DNFB-fed donors and control mice that did not receive pDC.
(B) DNBS-pulsed BM-DC (105) were injected s.c. into naive mice, either alone or together with 105 MAC-sorted pDC from liver or spleen of mice that received DNFB or vehicle by gavage 18 hr before. In an independent experiment, pDC were purified from the liver or MLN of DNFB-fed mice by FACS and similarly transferred into naive mice. At day 5 after DC transfer, the frequency of IFN-γ-secreting T cells was determined by an ELISPOT in skin-draining lymph nodes. Results are expressed as the mean number of IFN-γ SFC per 106 T cells ± SD of triplicate wells containing pooled lymph node cells from three mice per group.
(C and D) OVA-pulsed BM-DC (105) (C) or DNBS-pulsed BM-DC (D) were injected s.c. alone or together with 105 liver pDC FACS purified from mice that were fed 18 hr earlier with either DNFB, OXA, or OVA. Five days after immunization, recipients of DNBS-pulsed BM-DC were ear challenged with DNFB, and the ear swelling response was determined after 48 hr (C) *** p = 0.0002. All mice that received OVA-pulsed BM-DC were challenged with aggregated OVA in the footpad 7 days after immunization, and the DTH response, as measured by footpad swelling, was determined after 48 hr (D) * p = 0.03. These data are representative of two independent experiments.
Suppression of DTH Responses by Liver pDC from Ag-Fed Mice Is Antigen Specific
To further analyze whether liver pDCs carrying oral Ag can control CD4+ or CD8+ T cell-mediated DTH reactions in an Ag-specific way, adoptive transfer experiments were performed. Transfer of liver pDCs purified by FACS from OVA-fed mice suppressed the DTH response induced by immunization with OVA-pulsed BMDCs (Figure 6C). In contrast, liver pDCs from DNFB-fed mice or from oxazolone (OXA)-fed animals did not modify at all the DTH response. Likewise, analysis in the model of CD8+ T cell mediated CHS response to DNFB, induced by immunization with DNBS-pulsed BMDCs, revealed that only pDCs from DNFB-fed mice could suppress skin inflammation, whereas pDCs from mice fed with the hapten OXA or the protein Ag OVA were inefficient (Figure 6D). Similarly, only pDC from DNFB-fed animals efficiently suppressed the proliferation of CD8+ T cells in vitro in an Agspecific way (Figure S4B). Together with the results described above (Figure 5), these observations demonstrate that soon after feeding with a protein Ag or a hapten, liver pDC exhibit the capacity to present the oral Ag to T cells and to perform Ag-specific suppression of DTH responses in vivo.
Plasmacytoid DC Induce Anergy or Deletion of Ag-Specific T Cells in the Liver Shortly after Ag Feeding
The above results suggest that the liver constitutes a key organ where presentation of oral Ag by pDC might induce tolerance. To analyze the mechanism by which pDC in the liver induce tolerance, we analyzed the fate of Ag-specific CD8+ T cells after DNFB feeding. To circumvent the lack of tools to directly identify hapten-specific T cells, we developed a CHS model by transferring CD8+ T cells into T cell-deficient CD3ε-deficient mice. CD8+ T cells were purified from donor mice 1 or 6 days after vehicle or DNFB gavage, and adoptively transferred into Cd3ε−/− recipient mice. Three weeks later, recipient mice were skin sensitized with DNFB and tested for their capacity to initiate a CHS response upon DNFB challenge. Recipients of CD8+ T cells isolated either from the liver or MLN of vehicle-fed control B6 donors were able to mount a CHS response after DNFB sensitization and challenge (Figure 7A). In contrast, adoptive transfer of CD8+ T cells from day 6 DNFB-fed donors hardly induced any ear swelling response in Cd3ε−/− recipients and had a reduced capacity to differentiate into IFN-γ-secreting effector T cells upon skin sensitization (Figure 7B). These results demonstrate that Ag feeding of B6 mice induces, in MLN and liver, a massive decrease of the number of hapten-specific T cells able to differentiate into DTH effector cells. Importantly, this loss of Ag-reactive T cells, revealed by an impaired capacity to differentiate into CHSeffectors, was observed in MLN only at 6 days but not at 1 (Figure 7B) or 3 days (data not shown) after DNFB feeding. In contrast, the impaired capacity to differentiate into CHS effectors occurred in the liver as early as day 1 and persisted up to day 6 after gavage. Thus, hapten feeding resulted in anergy or deletion of hapten-reactive CD8+ T cells that is rapid and long lasting in the liver but delayed in the MLN. Further, depletion of pDC prior to hapten feeding of donor mice completely prevented this anergy or deletion of hapten-reactive MLN CD8+ T cells and restored their ability to differentiate into CHS effectors when transferred into Cd3ε−/− mice (Figure 7C). Finally, hapten feeding of wild-type B6 or MHC class II Aβ-deficient mice similarly impaired the capacity of MLN CD8+ T cells to generate a CHS response after adoptive transfer into _Cd3ε−/−_-recipient mice (Figure 7D), indicating that the orally induced anergy or deletion of Ag-specific T cells does not involve CD4+ T cells.
Figure 7. pDC Are Required for Orally Induced Functional Deletion of Hapten-Specific CD8+ T Cells.

(A–D) On day 0, 8–10 C57BL/6 or (D) MHC class II-deficient donor mice were fed with either DNFB or vehicle as control, and sacrificed 1 (B) or 6 (A–D) days later for isolation of CD8+ T cells. CD8+ T cells (3 3 105, A and B, or 2.5 3 106, C and D) purified from the (A and B) liver or (A–D) MLN were transferred intravenously into CD3ε-deficient recipient mice, that were sensitized 3 weeks later by skin painting with DNFB. Five days after sensitization, all mice were ear challenged with DNFB and the (A) CHS response of individual, and (B–D) the frequency of hapten-specific IFN-γ SFC in pooled skin draining lymph nodes (mean values of SFC ± SD of triplicate wells) were determined.
(C) pDC were depleted in donor mice by injection of the 120G8 mAb at days −1, 0 (day of feeding), +2, and +5. Results are representative of one out two independent experiments using five to eight Cd3 ε−/− recipients per group. Data of Figure 7B represent mean values of SFC ± SD of triplicate wells, containing CD8+ T cells pooled from five recipients. Statistics are indicated as p values in the figures.
Altogether, these data show that anergy or deletion of T cells specific for the oral Ag occurs at early times in the liver and later in MLN, and is mediated by pDC independently of CD4+ T cells.
DISCUSSION
This study demonstrates that pDC presenting dietary Ags are responsible for induction of oral tolerance and immune suppression affecting both CD4+ and CD8+ T cell responses. Using models of oral tolerance to a classical dietary protein Ag (OVA) and to the allergenic hapten DNFB, we showed that suppression of systemic CD4+ or CD8+ T cell-mediated DTH responses requires pDC. Indeed, in vivo depletion of pDC using either the Gr1 (RB6.8C5) mAb (Asselin-Paturel et al., 2001) or the more selective BST-2 (120G8) mAb (Asselin-Paturel et al., 2003; Blasius et al., 2006) abrogated oral tolerance and suppression of DTH responses in both models. A single intragastric hapten gavage in pDC-depleted naive mice resulted in oral T cell priming and development of a CHS response after skin hapten challenge. In addition, similarly to Ag feeding (Desvignes et al., 2000), adoptive transfer of pDC from Ag-fed mice prevented T cell priming and induction of DTH responses in an Ag-specific way. The important role of pDCs in oral tolerance is in line with the previous observation that Flt3-L treatment, which by increasing the number of DC including pDC (Gilliet et al., 2002), results in more efficient oral tolerance (Viney et al., 1998).
Anergy or deletion of Ag-specific T cells and active suppression by regulatory T cells are the two major mechanisms involved in orally induced T cell tolerance (Dubois et al., 2005; Mowat, 2003). We provide in vivo evidence that the main mechanism by which pDCs contribute to oral tolerance is by inducing anergy and/or deletion of nearly 70%–80% of Ag-specific T cells. Indeed, Ag feeding impaired the capacity of Ag-reactive T cells from liver and MLN of B6 donor mice to differentiate into DTH effectors upon adoptive transfer into T cell-deficient Cd3ε−/− mice, and this effect was completely prevented by pDC depletion of donor mice. Interestingly, pDC-mediated functional inactivation of Ag-specific T cells did not require CD4+ T cells indicating that the mechanism by which pDCs reduces the size of the circulating pool of Ag-specific T cells is independent of their capacity to trigger CD4+ Treg cell differentiation. It can be postulated that pDCs, which harbor low amounts of MHC peptide complexes and costimulatory molecules (Asselin-Paturel et al., 2001), make short or weak interactions with naive T cells. Similarly to conventional immature DC (Gett et al., 2003), pDCs may thus induce abortive proliferation and/or desensitization of Ag-specific T cells that fail to differentiate into CD4+ or CD8+ effector or memory T cells and die by neglect. The newly recognized cytotoxic potential of pDCs and the production of molecules favoring T cell apoptosis, like IDO (Sharma et al., 2007) and TNF-α (Kingham et al., 2007), could explain their suppressive function.
Due to its unique vascular supply, the liver receives via the portal vein a majority of soluble Ag from the gut lamina propria and has long been recognized as an immune-privileged organ that contributes to oral tolerance. Previous studies reported that creating a portocaval shunt to bypass the liver and prevent Ag transport from the gut to the liver via the portal vein, abrogates oral tolerance in response to either protein or hapten (Cantor and Dumont, 1967; Yang et al., 1994), Ag known to induce CD4+ and CD8+ T cell-mediated DTH. More recent studies using orthotopic liver transplants from OVA-fed donors documented that the liver within hours after gavage is sufficient to transfer oral tolerance and prevent systemic DTH to OVA and that this effect could be reproduced by transfer of liver nonparenchymal cells (Li et al., 2004). The possible importance of the liver as an inductive site of oral tolerance is further supported by our present data showing that the liver is highly enriched with pDC endowed with a tolerogenic potential, and is a site where pDCs induce anergy or deletion of Ag-specific T cells early after Ag feeding. pDCs, which account for up to 30% of liver CD11c+ cells, represented the only DC subset of the liver able to suppress a CHS response by preventing in vivo priming of hapten-specific IFN-g-producing CD8+ effectors, when transferred at the time and site of hapten sensitization. Although oral hapten rapidly distributes from the gut lamina propria to the liver as well as to the GALT and systemic lymphoid organs, anergy or deletion of Ag-reactive T cells is detected by day 1 in the liver and only several days later in MLN, suggesting that the very early CD4+ T cell-independent phase of oral tolerance to haptens is initiated by the Ag diffusion through the hematic route rather than by Ag transport into the draining MLN. Furthermore, we document that T cell tolerance and ability to prevent CHS to DNFB is achieved equally well by oral and intraportal antigen delivery.
pDCs might also exert their suppressive function in lymphoid organs during induction of oral tolerance. In this respect, pDCs harvested from MLN and liver of hapten-fed mice have the same efficacy to prevent hapten-specific CD8+ T cell priming, if transferred in similar numbers into recipient mice, indicating that, on a per cell basis, they share similar tolerogenic properties, although their in vivo tolerogenic potential could be conditioned in vivo by the local microenvironment (Bilsborough et al., 2003; Contractor et al., 2007). The fact that pDC from spleen of Ag-fed mice were weakly suppressive upon in vivo transfer may relate to the limited amount of oral Ag reaching the spleen, inasmuch as they could suppress hapten-specific CD8+ T cells when pulsed in vitro with the Ag.
Although our study highlights an unanticipated and essential role of pDCs in the rapid and efficient anergy or deletion of T cells specific to orally administrated Ag, we demonstrated earlier that oral tolerance to DNFB also critically depends on CD4+ regulatory T cells (Desvignes et al., 2000; Dubois et al., 2003). This strongly supports the idea that oral tolerance involves two sequential and complementary steps with distinct underlying mechanisms. The initial “mucosal” phase of oral tolerance is induced shortly after Ag penetration through the gut epithelium and consists of pDC-mediated deletion or inactivation of a high proportion of Ag-specific T cells, an event that is independent of regulatory CD4+ T cells (this study). In parallel, oral Ag promotes activation and/or differentiation of CD4+ Treg cells (Thorstenson and Khoruts, 2001; Zhang et al., 2001) with increased suppressive functions (Dubois et al., 2003). The subsequent “systemic” phase of oral tolerance operates in lymphoid organs upon systemic immunization with the same Ag and consists in the complete suppression of specific T cell priming mediated by CD4+ Treg cells (Desvignes et al., 2000). These include CD4+CD25+foxp3+ T cells, which control Ag-specific CD4+ and CD8+ T cell responses (Sakaguchi, 2004). That oral tolerance is abrogated by CD4+ T cell depletion at the time of systemic immunization with the same Ag (Desvignes et al., 2000; Dubois et al., 2003) supports that Treg cells induced or activated by Ag feeding (Thorstenson and Khoruts, 2001; Zhang et al., 2001) prevent systemic priming of Ag-reactive T cells that have escaped anergy or deletion or may have recently emigrated from the thymus.
Recent studies using elegant approaches of surgical ablation of MLN as well as leukocyte sequestration in secondary lymphoid organs, provided convincing evidence that MLN were essential for oral tolerance in a model of CD4+ T cell-mediated DTH response (Worbs et al., 2006). Thus, it is tempting to speculate that MLN and liver may play complementary roles with a division of labor with different temporal requirements to achieve efficient oral tolerance. On the one hand, the liver due to its anatomical characteristics, and the constant blood flow is the first site exposed to large quantities of oral Ag in soluble form and pDC enriched in this organ cause rapid and efficient deletion of oral Ag-specific T cells (this study) constantly trafficking from the blood. This is consistent with the observation that the liver constitutes a privileged site for apoptosis of both CD8+ T cells (Crispe, 2003) and CD4+ T cells (Watanabe et al., 2003). On the other hand, MLN which collects via lymph intestinal Ags loaded onto myeloid DC emigrating from lamina propria might constitute the preferential site of activation/differentiation of regulatory T cells specific for dietary Ag (Thorstenson and Khoruts, 2001; Zhang et al., 2001). In this respect, myeloid CD103+ DC from the MLN have the potential to drive the differentiation of functional Foxp3+ Treg in vitro in the presence of retinoic acid and TGF-β (Coombes et al., 2007), although the role of CD103+ DC in oral tolerance remains to be formally demonstrated.
Our data clearly demonstrate that pDC perform efficient CD4+ and CD8+ T cell tolerance to orally administered haptens and proteins. The notion that pDCs may control the emergence of food allergy, drug-induced allergic DTH reactions, as well as intestinal inflammatory diseases, and also favor transplantation tolerance opens up new avenues for the development of therapeutic approaches targeting pDC.
EXPERIMENTAL PROCEDURES
Mice
Female C57BL/6 mice (2–4 months old) were purchased from Charles River Laboratories (L’Arbresle, France). MHC-class II (H2-Ab−/−)-deficient mice were kindly provided by D.Mathis and C. Benoist, and CD3ε−/− mice were obtained from M. Malissen. These mice were bred as homozygotes at the institute’s animal facility (Plateau de Biologie Expérimentale de la Souris, PBES, Ecole Normale Supérieure de Lyon) under specific pathogen-free conditions. All experiments have been approved by the “Comité regional d’éthique pour l’expérimentation animale” (CREEA) and were performed according to institutional guidelines.
Contact Hypersensitivity (CHS) to DNFB
Mice were sensitized epicutaneously on abdominal skin with 0.5% DNFB (Sigma, St. Quentin Fallavier, France) diluted in acetone-olive oil (4:1, v/v) and ear-challenged 5 days later with 0.15% DNFB. CHS was determined by the mouse ear swelling test as previously described (Dubois et al., 2003). Ear swelling in unsensitized but ear challenged mice was usually <20 μm. In some experiments (tolerance induced by the portal vein), mice were sensitized on the dorsal skin with 0.2% of DNFB. CHS was also induced by s.c. injection of BM-DC (105 cells) pulsed with 1.6 mM of DNBS.
Delayed Type Hypersensitivity (DTH) to OVA
Mice were immunized subcutaneously with 50 μg OVA (Seikagaku corporation, Tokyo, Japan) in 50 μl PBS/CFA emulsion, or 105 OVA-pulsed BM-DC, and were challenged 7 days later by s.c. injection of 250 μg aggregated OVA in 30 μl of PBS in the left hind footpad. The right footpad received PBS as control. OVA-specific DTH was determined by footpad swelling at various times after challenge, as described (Krasteva et al., 1998).
Tolerance Induction by Intragastric or Intraportal Antigen Delivery
Oral tolerance to DNFB was induced by intragastric administration of 0.1% DNFB (300 μl) in acetone/olive oil (1/10, v/v) 7 days before skin sensitization with DNFB, as described (Desvignes et al., 2000). For induction of tolerance to OVA, mice were fed three times 20 μg of OVA (grade II, Sigma-Aldrich) in 200μl PBS by gavage on days −13, −10, and −7 beforeOVA/CFA immunization.
Tolerance via the portal route was induced by injection of 125 mg/kg of DNBS (Sigma) in 100 μl of PBS into the portal vein of mice under isoflurane anesthesia. For depletion of pDC, mice received multiple injections (200 μl) of either of anti-Gr1 antibody (clone RB6-8C5) as 1/5 dilution of ascitic fluid in PBS, or 0.5 mg of purified rat IgG anti-BST-2 mAb (clone 120G8) (Asselin-Paturel et al., 2003).
Isolation of Liver, Spleen, and MLN Leucocytes
Leucocytes were isolated from liver, MLN, and spleen by enzymatic digestion at 37°C for 30 min using collagenase type IA (1 mg/ml) and DNase I (0.1 mg/ml) (each from Sigma, St Quentin Fallavier, France). Liver (Lian et al., 2003) leukocytes were separated from contaminating hepatocytes by centrifugation over a 40%–80% Percoll gradient (Percoll, density 1,130, Amersham Biosciences) (Lian et al., 2003).
Flow Cytometry Analysis
Phenotypic studies were conducted by direct immunofluorescence staining using the following Ab panel from Becton Dickinson (Le Pont de Claix, France): biotinylated or phycoerythrin (PE)-conjugated hamster anti-mouse CD11c (clone HL3), FITC, or PE-Cy5.5-conjugated rat anti-mouse CD11b (clone M1/70), PE-conjugated rat anti-mouse CD8α (clone 53-6.7), PE-conjugated mouse anti mouse NK1.1 (clone PK136). We also used Alexa488-conjugated rat anti mouse pDCs (clone 120G8) (Asselin-Paturel et al., 2003). Fluorescence was measured with a FACS can equipped with CellQuest software (Becton Dickinson, Mountain View, CA).
Purification of CD11c+ DC Subsets
DC were enriched from liver leukocytes by magnetic sorting using anti-CD11c-coated microbeads (Miltenyi Biotec, France). This routinely yielded to approximately 90% pure CD11c+ cells. DC subsets were isolated by FACS (FACStar cell sorter, BD Biosciences) using biotin-conjugated anti-CD11c mAb and APC-conjugated streptavidin, PE-Cy5.5-conjugated anti-CD11b, PE-conjugated anti-NK1.1, and Alexa488-conjugated 120G8. Several subsets, including CD11c+ CD11b+ NK1.1−, CD11c+ CD11b− NK1.1−, CD11c+ CD11b+or− NK1.1+ cells, CD11c+ CD11b− NK1.1− 120G8− (pDC depleted), and CD11c+ CD11b− NK1.1− 120G8+ (sorted pDC) (Figure 3E), were obtained with a purity >97%. Alternatively, pDC were enriched from the liver, MLN, or spleen by MACS using mPDCA1-coated microbeads and MS columns (Miltenyi Biotec). This yielded 70%–80% pure pDC expressing CD11c and BST-2 (Figure S3A). For certain experiments, >97% pure CD11cintBST-2+ pDC were obtained by FACS (Figure S3B).
Ag-Specific T Cell Responses
The presence of hapten-specific T cells was determined by proliferation assay, IFN- ELISA, and IFN-γ ELISPOT after in vitro restimulation of either MAC-sorted CD8+ T cells or total LN cells with hapten-pulsed spleen cells or BM-DC as described (Dubois et al., 2003; Le Borgne et al., 2006). For ex vivo antigen presentation assay, 105 leukocytes isolated from either DNFB or vehicle-fed mice were used as APC and cocultured with hapten-specific CD8+ T cells. Hapten-specific T cell proliferation was determined on day 3 of culture by 3H-thymidine incorporation (1 μCi/well) during the last 8 hr of culture. OVA-specific T cell responses were determined by measuring IFN-γ production by spleen cells stimulated for 48 hr in vitro with 1 mg/ml OVA.
Statistical Analysis
Statistics were performed using the Student’s t test. P values <0.05 were considered to be statistically significant.
Supplementary Material
Figures
ACKNOWLEDGMENTS
We are grateful to Dr. Amine Bouheddi and Pr. Thierry Roger from Ecole Nationale Vétérinaire de Lyon, Unité d’Anatomie, Marcy l’Etoile, for their expertise and skilful assistance in performing portocaval injections. We thank Pierre-Yves Durand and Jean-Benoit Le Luduec for their expert technical assistance. We would also like to thank the staff of the Plateau de Biologie Expérimentale de la Souris (PBES) of Ecole Normale Supérieure de Lyon for breeding and maintaining mouse strains and Chantal Bella from the flow cytometry facility of IFR-128 Biosciences Gerland-Lyon Sud for cell sorting. Anne Goubier was supported by grants from INSERM (poste d’accueil), Fondation pour la Recherche Médicale, and Académie de Médecine.
Footnotes
REFERENCES
- Asselin-Paturel C, Boonstra A, Dalod M, Durand I, Yessaad N, Dezutter-Dambuyant C, Vicari A, O’Garra A, Biron C, Briere F, Trinchieri G. Mouse type I IFN-producing cells are immature APCs with plasmacytoid morphology. Nat. Immunol. 2001;2:1144–1150. doi: 10.1038/ni736. [DOI] [PubMed] [Google Scholar]
- Asselin-Paturel C, Brizard G, Pin JJ, Briere F, Trinchieri G. Mouse strain differences in plasmacytoid dendritic cell frequency and function revealed by a novel monoclonal antibody. J. Immunol. 2003;171:6466–6477. doi: 10.4049/jimmunol.171.12.6466. [DOI] [PubMed] [Google Scholar]
- Bilsborough J, George TC, Norment A, Viney JL. Mucosal CD8alpha+ DC, with a plasmacytoid phenotype, induce differentiation and support function of T cells with regulatory properties. Immunology. 2003;108:481–492. doi: 10.1046/j.1365-2567.2003.01606.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blasius AL, Giurisato E, Cella M, Schreiber RD, Shaw AS, Colonna M. Bone marrow stromal cell antigen 2 is a specific marker of type I IFN-producing cells in the naive mouse, but a promiscuous cell surface antigen following IFN stimulation. J. Immunol. 2006;177:3260–3265. doi: 10.4049/jimmunol.177.5.3260. [DOI] [PubMed] [Google Scholar]
- Boonstra A, Asselin-Paturel C, Gilliet M, Crain C, Trinchieri G, Liu YJ, O’Garra A. Flexibility of mouse classical and plasmacytoid-derived dendritic cells in directing T helper type 1 and 2 cell development: dependency on antigen dose and differential toll-like receptor ligation. J. Exp. Med. 2003;197:101–109. doi: 10.1084/jem.20021908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantor HM, Dumont AE. Hepatic suppression of sensitization to antigen absorbed into the portal system. Nature. 1967;215:744–745. doi: 10.1038/215744a0. [DOI] [PubMed] [Google Scholar]
- Contractor N, Louten J, Kim L, Biron CA, Kelsall BL. Cutting edge: Peyer’s patch plasmacytoid dendritic cells (pDCs) produce low levels of type I interferons: possible role for IL-10, TGFbeta, and prostaglandin E2 in conditioning a unique mucosal pDC phenotype. J. Immunol. 2007;179:2690–2694. doi: 10.4049/jimmunol.179.5.2690. [DOI] [PubMed] [Google Scholar]
- Coombes JL, Siddiqui KR, Arancibia-Carcamo CV, Hall J, Sun CM, Belkaid Y, Powrie F. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-{beta}- and retinoic acid-dependent mechanism. J. Exp. Med. 2007;204:1757–1764. doi: 10.1084/jem.20070590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crispe IN. Hepatic T cells and liver tolerance. Nat. Rev. Immunol. 2003;3:51–62. doi: 10.1038/nri981. [DOI] [PubMed] [Google Scholar]
- De Heer HJ, Hammad H, Soullie T, Hijdra D, Vos N, Willart MA, Hoog-steden HC, Lambrecht BN. Essential role of lung plasmacytoid dendritic cells in preventing asthmatic reactions to harmless inhaled antigen. J. Exp. Med. 2004;200:89–98. doi: 10.1084/jem.20040035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desvignes C, Etchart N, Kehren J, Akiba I, Nicolas JF, Kaiserlian D. Oral administration of hapten inhibits in vivo induction of specific cytotoxic CD8+ T cells mediating tissue inflammation: a role for regulatory CD4+ T cells. J. Immunol. 2000;164:2515–2522. doi: 10.4049/jimmunol.164.5.2515. [DOI] [PubMed] [Google Scholar]
- Dubois B, Chapat L, Goubier A, Papiernik M, Nicolas JF, Kaiserlian D. Innate CD4+CD25+ regulatory T cells are required for oral tolerance and inhibition of CD8+ T cells mediating skin inflammation. Blood. 2003;102:3295–3301. doi: 10.1182/blood-2003-03-0727. [DOI] [PubMed] [Google Scholar]
- Dubois B, Goubier A, Joubert G, Kaiserlian D. Oral tolerance and regulation of mucosal immunity. Cell. Mol. Life Sci. 2005;62:1322–1332. doi: 10.1007/s00018-005-5036-0. [DOI] [PubMed] [Google Scholar]
- Faria AM, Weiner HL. Oral tolerance. Immunol. Rev. 2005;206:232–259. doi: 10.1111/j.0105-2896.2005.00280.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gett AV, Sallusto F, Lanzavecchia A, Geginat J. T cell fitness determined by signal strength. Nat. Immunol. 2003;4:355–360. doi: 10.1038/ni908. [DOI] [PubMed] [Google Scholar]
- Gilliet M, Boonstra A, Paturel C, Antonenko S, Xu XL, Trinchieri G, O’Garra A, Liu YJ. The development of murine plasmacytoid dendritic cell precursors is differentially regulated by FLT3-ligand and granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 2002;195:953–958. doi: 10.1084/jem.20020045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutgemann I, Fahrer AM, Altman JD, Davis MM, Chien YH. Induction of rapid T cell activation and tolerance by systemic presentation of an orally administered antigen. Immunity. 1998;8:667–673. doi: 10.1016/s1074-7613(00)80571-3. [DOI] [PubMed] [Google Scholar]
- Hue S, Mention JJ, Monteiro RC, Zhang S, Cellier C, Schmitz J, Verkarre V, Fodil N, Bahram S, Cerf-Bensussan N, Caillat-Zucman S. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity. 2004;21:367–377. doi: 10.1016/j.immuni.2004.06.018. [DOI] [PubMed] [Google Scholar]
- Husby S, Mestecky J, Moldoveanu Z, Holland S, Elson CO. Oral tolerance in humans. T cell but not B cell tolerance after antigen feeding. J. Immunol. 1994;152:4663–4670. [PubMed] [Google Scholar]
- Ju C, Pohl LR. Immunohistochemical detection of protein adducts of 2,4-dinitrochlorobenzene in antigen presenting cells and lymphocytes after oral administration to mice: lack of a role of Kupffer cells in oral tolerance. Chem. Res. Toxicol. 2001;14:1209–1217. doi: 10.1021/tx0100587. [DOI] [PubMed] [Google Scholar]
- Kehren J, Desvignes C, Krasteva M, Ducluzeau MT, Assossou O, Horand F, Hahne M, Kagi D, Kaiserlian D, Nicolas JF. Cytotoxicity is mandatory for CD8(+) T cell-mediated contact hypersensitivity. J. Exp. Med. 1999;189:779–786. doi: 10.1084/jem.189.5.779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kingham TP, Chaudhry UI, Plitas G, Katz SC, Raab J, DeMatteo RP. Murine liver plasmacytoid dendritic cells become potent immunostimulatory cells after Flt-3 ligand expansion. Hepatology. 2007;45:445–454. doi: 10.1002/hep.21457. [DOI] [PubMed] [Google Scholar]
- Krasteva M, Kehren J, Horand F, Akiba H, Choquet G, Ducluzeau MT, Tedone R, Garrigue JL, Kaiserlian D, Nicolas JF. Dual role of dendritic cells in the induction and down-regulation of antigen-specific cutaneous inflammation. J. Immunol. 1998;160:1181–1190. [PubMed] [Google Scholar]
- Kunkel D, Kirchhoff D, Nishikawa S, Radbruch A, Scheffold A. Visualization of peptide presentation following oral application of antigen in normal and Peyer’s patches-deficient mice. Eur. J. Immunol. 2003;33:1292–1301. doi: 10.1002/eji.200323383. [DOI] [PubMed] [Google Scholar]
- Le Borgne M, Etchart N, Goubier A, Lira SA, Sirard JC, van Rooijen N, Caux C, Ait-Yahia S, Vicari A, Kaiserlian D, Dubois B. Dendritic Cells Rapidly Recruited into Epithelial Tissues via CCR6/CCL20 Are Responsible for CD8(+) T Cell Crosspriming In Vivo. Immunity. 2006;24:191–201. doi: 10.1016/j.immuni.2006.01.005. [DOI] [PubMed] [Google Scholar]
- Li W, Chou ST, Wang C, Kuhr CS, Perkins JD. Role of the liver in peripheral tolerance: induction through oral antigen feeding. Am. J. Transplant. 2004;4:1574–1582. doi: 10.1111/j.1600-6143.2004.00551.x. [DOI] [PubMed] [Google Scholar]
- Lian ZX, Okada T, He XS, Kita H, Liu YJ, Ansari AA, Kikuchi K, Ikehara S, Gershwin ME. Heterogeneity of dendritic cells in the mouse liver: identification and characterization of four distinct populations. J. Immunol. 2003;170:2323–2330. doi: 10.4049/jimmunol.170.5.2323. [DOI] [PubMed] [Google Scholar]
- Lu L, Bonham CA, Liang X, Chen Z, Li W, Wang L, Watkins SC, Nalesnik MA, Schlissel MS, Demestris AJ, et al. Liver-derived DEC205+B220+CD19- dendritic cells regulate T cell responses. J. Immunol. 2001;166:7042–7052. doi: 10.4049/jimmunol.166.12.7042. [DOI] [PubMed] [Google Scholar]
- Martin P, Del Hoyo GM, Anjuere F, Arias CF, Vargas HH, Fernandez LA, Parrillas V, Ardavin C. Characterization of a new subpopulation of mouse CD8alpha+ B220+ dendritic cells endowed with type 1 interferon production capacity and tolerogenic potential. Blood. 2002;100:383–390. doi: 10.1182/blood.v100.2.383. [DOI] [PubMed] [Google Scholar]
- Mowat AM. Anatomical basis of tolerance and immunity to intestinal antigens. Nat. Rev. Immunol. 2003;3:331–341. doi: 10.1038/nri1057. [DOI] [PubMed] [Google Scholar]
- Munn DH, Sharma MD, Hou D, Baban B, Lee JR, Antonia SJ, Mes-sina JL, Chandler P, Koni PA, Mellor AL. Expression of indo-leamine 2,3-dioxygenase by plasmacytoid dendritic cells in tumor-draining lymph nodes. J. Clin. Invest. 2004;114:280–290. doi: 10.1172/JCI21583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nancey S, Holvoet S, Graber I, Joubert G, Philippe D, Martin S, Nicolas JF, Desreumaux P, Flourie B, Kaiserlian D. CD8+ cytotoxic T cells induce relapsing colitis in normal mice. Gastroenterology. 2006;131:485–496. doi: 10.1053/j.gastro.2006.05.018. [DOI] [PubMed] [Google Scholar]
- Ochando JC, Homma C, Yang Y, Hidalgo A, Garin A, Tacke F, Angeli V, Li Y, Boros P, Ding Y, et al. Alloantigen-presenting plasmacytoid dendritic cells mediate tolerance to vascularized grafts. Nat. Immunol. 2006;7:652–662. doi: 10.1038/ni1333. [DOI] [PubMed] [Google Scholar]
- Sakaguchi S. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu. Rev. Immunol. 2004;22:531–562. doi: 10.1146/annurev.immunol.21.120601.141122. [DOI] [PubMed] [Google Scholar]
- Sharma MD, Baban B, Chandler P, Hou DY, Singh N, Yagita H, Azuma M, Blazar BR, Mellor AL, Munn DH. Plasmacytoid dendritic cells from mouse tumor-draining lymph nodes directly activate mature Tregs via indoleamine 2,3-dioxygenase. J. Clin. Invest. 2007;117:2570–2582. doi: 10.1172/JCI31911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spahn TW, Fontana A, Faria AM, Slavin AJ, Eugster HP, Zhang X, Koni PA, Ruddle NH, Flavell RA, Rennert PD, Weiner HL. Induction of oral tolerance to cellular immune responses in the absence of Peyer’s patches. Eur. J. Immunol. 2001;31:1278–1287. doi: 10.1002/1521-4141(200104)31:4<1278::aid-immu1278>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, Belkaid Y. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. J. Exp. Med. 2007;204:1775–1785. doi: 10.1084/jem.20070602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swarbrick ET, Stokes CR, Soothill JF. Absorption of antigens after oral immunisation and the simultaneous induction of specific systemic tolerance. Gut. 1979;20:121–125. doi: 10.1136/gut.20.2.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorstenson KM, Khoruts A. Generation of anergic and potentially immunoregulatory CD25+CD4 T cells in vivo after induction of peripheral tolerance with intravenous or oral antigen. J. Immunol. 2001;167:188–195. doi: 10.4049/jimmunol.167.1.188. [DOI] [PubMed] [Google Scholar]
- Viney JL, Mowat AM, O’Malley JM, Williamson E, Fanger NA. Expanding dendritic cells in vivo enhances the induction of oral tolerance. J. Immunol. 1998;160:5815–5825. [PubMed] [Google Scholar]
- Watanabe T, Katsukura H, Shirai Y, Yamori M, Nishi T, Chiba T, Kita T, Wakatsuki Y. A liver tolerates a portal antigen by generating CD11c+ cells, which select Fas ligand+ Th2 cells via apoptosis. Hepatology. 2003;38:403–412. doi: 10.1053/jhep.2003.50343. [DOI] [PubMed] [Google Scholar]
- Worbs T, Bode U, Yan S, Hoffmann MW, Hintzen G, Bernhardt G, Forster R, Pabst O. Oral tolerance originates in the intestinal immune system and relies on antigen carriage by dendritic cells. J. Exp. Med. 2006;203:519–527. doi: 10.1084/jem.20052016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang R, Liu Q, Grosfeld JL, Pescovitz MD. Intestinal venous drainage through the liver is a prerequisite for oral tolerance induction. J. Pediatr. Surg. 1994;29:1145–1148. doi: 10.1016/0022-3468(94)90297-6. [DOI] [PubMed] [Google Scholar]
- Zhang X, Izikson L, Liu L, Weiner HL. Activation of CD25(+)CD4(+) regulatory T cells by oral antigen administration. J. Immunol. 2001;167:4245–4253. doi: 10.4049/jimmunol.167.8.4245. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figures