Antitumor activity of tumor-targeted RNA replicase-based plasmid that expresses interleukin-2 in a murine melanoma model (original) (raw)
. Author manuscript; available in PMC: 2014 Jun 3.
Published in final edited form as: Mol Pharm. 2013 May 17;10(6):2404–2415. doi: 10.1021/mp400033m
Abstract
Double stranded RNA (dsRNA) has multiple antitumor mechanisms that may be used to control tumor growth. Previously we have shown that treatment of solid tumors with a plasmid that encodes Sindbis viral RNA replicase complex, pSIN-β, significantly inhibited the growth of tumors in mice. In the present study, we evaluated the feasibility of further improving the antitumor activity of the pSIN-β plasmid by incorporating interleukin-2 (IL2) gene into the plasmid. The resultant pSIN-IL2 plasmid was delivered to mouse melanoma cells that overexpress sigma receptor. Here we report that the pSIN-IL2 plasmid was more effective at controlling the growth of B16 melanoma in mice when complexed with sigma receptor-targeted liposomes than with the untargeted liposomes. Importantly, the pSIN-IL2 plasmid was more effective than pSIN-β plasmid at controlling the growth of B16 melanoma in mice, and B16 tumor-bearing mice that were treated with pSIN-IL2 had an elevated number of activated CD4+, CD8+, and natural killer cells, as compared to those treated with pSIN-β. The RNA replicase-based, IL2-expressing plasmid may have application in melanoma gene therapy.
Keywords: dsRNA therapy, sigma receptor, anisamide, IL2 therapy, melanoma
Introduction
Advanced melanoma remains a significant problem although cancer therapy has improved dramatically over the past decades. Melanoma is a malignancy with the worst prognosis, and death rates have been rising faster than those of other cancers.1 The median survival time for patients with metastatic melanoma is between 6 and 10 months.2 Metastatic melanoma is one of the most resistant cancers to single agent chemotherapy, combination chemotherapy, and immunotherapy.
IL2 therapy is clinically efficacious in patients with advanced melanoma and renal cell carcinoma.3 IL2 is the only FDA-approved immunotherapeutic agents for patients with metastatic melanoma with an overall response rate of 13–17%.4 IL2 has no direct impact on cancer cells; its antitumor effects are due to its ability to modulate immune responses.5 IL2 affects cytotoxic CD8+ T-cells, CD4+ T-cells, natural killer (NK) cells, B-cells, and macrophages.6, 7 Both CD8+ and CD4+ lymphocytes are required for tumor regression in multiple tumor models.6 The short half-life of IL2 in vivo is a major limitation of IL2 treatment, and toxic side effects are typically observed at high doses. Alternatives to overcome these limitations include continuous infusion,3, 8 prolonged low dose daily injection, and local regional injections of IL2 into the tumor area.3, 9 An additional method for IL2 delivery is by intratumoral injection of plasmid DNA that expresses IL2. There are a number of phase I-II clinical trials that evaluate whether locally administrated IL2-expressing plasmid improves patient outcome and induces intratumoral immune responses (www.clinicaltrials.gov). In fact, ultrasound-guided intralesional injection of IL2-expressing plasmid was shown to be safe and, in some cases, led to a durable objective responses in several clinical trials in patients with prostate cancer,10 metastatic renal cell carcinoma,11 and metastatic melanoma.12
Introducing IL2 gene into tumor cells may also help overcome the limitations associated with in vivo administration of IL2, including avoiding the side effects caused by high doses of IL2 necessary for its antitumor activity. 3 Expression of IL2 in tumor cells has been shown to reduce tumor cell tumorigenicity in vivo and induce specific antitumor immunity.12–14 Treatment with IL2 expressing plasmid has also been shown to improve the efficacy of IL2 immunotherapy, while avoiding the toxicity associated with high doses of IL2.12 Repeated intratumoral injection of IL2 plasmid complexed with liposomes was shown to be safe and well tolerated.12 The predicted outcome is that tumor cells transfected with IL2 gene will produce IL2 at a level that is low enough to prevent significant side effects to the patients, but sufficient to create an antitumor response through the immune system. 3
It is known that double-stranded (dsRNA) molecules have multiple direct and indirect pro-apoptotic, anti-proliferative, anti-angiogenic, and immunostimulatory activities.15–17 Double-stranded RNA is also a potent inducer of type I interferons (IFN-α/β), which have pro-apoptotic and anti-angiogenic effects. 17, 18 Data from recent studies showed that intracellular dsRNA is more effective than extracellular dsRNA in promoting cell apoptosis and in orchestrating innate and adaptive immune responses.19–21 Previously, we proposed and demonstrated the feasibility of using an RNA replicase-based plasmid (pSIN-β) to generate dsRNA intracellularly in the transfected tumor cells to control tumor growth in culture and in mice. 22 We have shown that treatment of tumor-bearing mice with the pSIN-β plasmid complexed with cationic liposomes by peritumoral injections significantly inhibited the growth of tumors pre-established in a mouse model and, in many cases, caused complete tumor regression, suggesting the use of RNA replicase-based plasmid as a novel tumor molecular therapy. In addition we have shown that actively targeting the pSIN-β plasmid to tumors using liposomes surface-conjugated with a ligand to a receptor overexpressed by tumor cells more effectively controlled the tumor growth, as compared to utilizing conventional liposomes.23
Sigma receptors are ubiquitously expressed membrane bound proteins that are highly conserved in mammalian systems. The presence of sigma 1 and sigma 2 receptor subtypes was reported in human amelanotic melanoma.24 Anisamide, a benzamide derivative, acts as a ligand to sigma receptors and is not subtype specific.25 Anisamide was previously used as a functional ligand of the sigma 1 receptor to successfully target liposomes to sigma receptor-overexpressing melanoma cells.26–28
The present study is designed to further improve the antitumor activity of the pSIN-β plasmid by incorporating the IL2 gene into the plasmid. The resultant pSIN-IL2 plasmid was targeted to melanoma cells that overexpress sigma receptors using cationic liposomes surface-conjugated with anisamide. We reason that the IL2 produced by tumor cells transfected with the pSIN-IL2 plasmid will help improve the antitumor activity of the dsRNA produced by the pSIN plasmid.
Materials and methods
Plasmids and cells
Plasmid pSIN-β was kindly provided by Dr. Richard Weiss (University of Salzburg, Salzburg, Austria). Plasmid pCMV-β was from the American Type Culture Collection (ATCC, Manassas, VA). The pORF-mIL2 plasmid was from InvivoGen (San Diego, CA). Plasmids were amplified in E. coli DH5α under selective growth conditions and purified using a QIAGEN midiprep kit (Valencia, CA) according to the manufacturer’s instruction. Large-scale plasmid purification was performed by GenScript (Piscataway, NJ). The B16-OVA cells were kindly provided by Dr. Edith M. Lord and Dr. John Frelinger (University of Rochester Medical Center, Rochester, NY) and cultured in RPMI 1640 (Invitrogen, Carlsbad, CA) medium supplemented with 5% fetal bovine serum (FBS) and 400 µg/ml of G418 (Sigma-Aldrich, St. Louis, MO).
Construction of pSIN-IL2 plasmid
To construct pSIN-IL2 plasmid, the murine interleukin-2 (IL2) gene from the pORF-mIL2 plasmid was PCR-amplified with primers IL2 F (5’-ACA AGT TCT AGA CAC CGG CGA AGG AGG GCC-3’) and IL2 R (5’-CCT AGA GCA TGC ATT GAG GGC TTG TTG AGA-3’). The PCR product was digested with XbaI and SphI, and the IL2 gene was used to replace the p-galactosidase gene downstream of the subgenomic promoter in the pSIN-β.29 Positive clones were screened for insert using restriction digestion and further confirmed by DNA sequencing (ABI 3730XL DNA analyzer, Applied Biosystems, Foster City, CA). A map of the pSIN-IL2 plasmid is included in the Supporting Information (Fig. S1). In vitro transfection of the pSIN-IL2 into B16-OVA murine melanoma cells using Lipofectamine® 2000 reagent (Invitrogen) confirmed the expression of IL2 (Fig. S2).
Synthesis of DSPE-PEG-anisamide (DSPE-PEG-AA)
DSPE-PEG-AA was synthesized according to Bangerjee et. al. with slight modifications.26 Briefly, _ρ_-anisoyl chloride (3.412 g, 0.02 mmol) in benzene (9 ml) at 50°C was added to an aqueous solution of 2-bromo ethylamine hydrobromide (3.73 g, 18.2 mmol) in water (27 ml). Sodium hydroxide 5% (33.5 ml) was gradually added to the emulsion while shaking and cooling in running water. After 2 h of stirring, solid amide was filtered and washed with 0.1 M sodium carbonate. After lyophilization, 1H NMR spectra were obtained to confirm the production of _N_-(2-bromoethyl)-4-methoxybenzamide. _N_-(2-bromoethyl)-4-methoxybenzamide (50 mg, 0.4 mmol) was then reacted with DSPE-PEG-2000-NH2 (50 mg, 23.2 mmol) in acetonitrile (2.5 ml) in the presence of _N,N_-diisopropylethylamine (DIPEA) (15 µl, 0.1 mmol) at 65–70°C for 8 h. Methanol (2.5 ml) was added to the reaction mixture followed by excess ether (25 ml), and the mixture was kept at −80°C for 24 h. The precipitate was collected after centrifugation, and recrystallization was performed with the addition of methanol (2.5 ml) followed by ether (17.5 ml) at 4°C for 12 h. The overall yield on average was 46%. The product DSPE-PEG-AA was confirmed by 1H NMR and ESI mass spectrometry.
Preparation of anisamide-conjugated PEGylated liposomes (AA-PEG-LP)
Cationic liposomes were prepared using cholesterol (Sigma-Aldrich), egg phosphatidylcholine (Avanti Polar Lipids, Inc, Alabaster, AL), 1,2,-dioleoyl-3-trimethylamonium-propane (DOTAP, Avanti) and DSPE-PEG-(2000)-methoxy (Avanti), or DSPE-PEG-(2000)-AA at a molar ratio of 4.6:10.8:19.6:1.5. To fluorescently label the liposomes, 1,2,-dioleoyl glycero-3-phosphoethanolamine-N-(lissamine rhodamine B sulfonyl) (Rho-DOPE) (Avanti 1% w/w) was included in the lipids. Lipids were mixed in a 20-ml glass scintillation vial followed by solvent evaporation and the formation of a thin film by placing the vial under a constant stream of nitrogen gas. The film was hydrated with phosphate buffered saline (PBS) (10 mM, pH 7.4) and sonicated for 1 h, followed by a 24 h hydration at room temperature with intermittent vortexing. The liposome suspension was forced through polycarbonate filters (1.0, 0.4, and 0.1 µm, sequentially) using a mini-extruder (Avanti).28 The final concentration of DOTAP in the liposomes was 10 mg/ml.
The particle size and zeta potential of the cationic liposomes, PEG-liposomes, and anisamide-conjugated PEGylated liposomes (AA-PEG-LP) were measured using a Malvern Zetasizer® Nano ZS (Westborough, MA). Liposomes were mixed with plasmid DNA at various ratios to prepare lipoplexes. The stability of the lipoplexes was determined in simulated biological medium.30 Briefly, the lipoplexes were diluted in normal saline with 10% FBS, and their sizes were measured immediately (0 min) and after 30 min of incubation at 37°C.
Plasmid DNA uptake assay
B16-OVA cells (1 × 105) were seeded in 24-well plates (n = 6) and incubated at 37°C, 5% CO2 for 24 h or until 60% confluency. The pSIN-IL2 plasmid was labeled using a Label IT® fluorescein nucleic acid labeling kit (Mirus, Madison, WI) according to the manufacturer’s instruction. Freshly labeled pSIN-IL2 (0.75 µg) was complexed with the AA-PEG-liposomes or the PEG-liposomes (DOTAP, 3.75 µg) and incubated for at least 15 min at room temperature. The resultant lipoplexes were added to each well and incubated for 1 h at 37°C, 5% CO2. Cells were washed with PBS and lysed using Triton X-100 (in 0.5% in 20 mM Tris, 100 mM NaCl, and 1 mM EDTA) following by incubation at −80°C for 1 h. The fluorescence intensity was measured at 492/518 nm in a black bottom plate using a BioTek Synergy® Multi-Mode Microplate Reader (Winooski, CT).
In vitro transfection
B16-OVA cells (2.5 × 105 cells/well) were seeded into 24-well plates (n = 3). After overnight incubation, cells were incubated with pSIN-IL2 complexed with AA-PEG-LP or PEG- LP (DNA : DOTAP, 1:10, w/w) for 24 h. Briefly 1 µg of plasmid was diluted in 50 µl of serum-free medium, and 1 µl of corresponding liposomes were diluted in 50 µl of serum-free medium. The diluted plasmid DNA samples were mixed with the diluted liposome suspension and incubated at room temperature for 15 min. The complexes were added to B16-OVA cell-containing wells. The time when the complexes were applied to the cell culture medium was defined as 0 h. After incubation, the supernatant was collected and analyzed for IL-2 using a mouse IL-2 ELISA kit (BD Biosciences).
Caspase-3 activity assay
Caspase-3 activity was determined using a caspase-3 assay kit from Sigma-Aldrich. B16-OVA cells (100,000 cells/well in 300 µl) were seeded in 24-well plates. After overnight incubation, the cells were transfected with pSIN-IL2 (1 µg) complexed with AA-PEG-LP or PEG-LP (DNA : DOTAP, 1:10, w/w). Forty-eight hours later, cells were detached and washed with PBS, centrifuged, re-suspended in 50 µl of cell lysis buffer, and incubated at 4°C for 15 min. The cell lysate was centrifuged at 12,000 × g for 15 min at 4°C, and the supernatant was mixed with the assay substrate, acetyl-Asp-Glu-Val-Asp-7-amido-4-methylcourmarin (Ac-DEVD-AMC). The mixture was incubated for 48 h for the hydrolysis of the Ac-DEVD-AMC by caspase-3 to release the fluorescent AMC. AMC was quantified by measuring the fluorescence intensity at 360/460 nm using a BioTek Synergy™ HT Multi-Mode Microplate Reader.
Plasmid DNA uptake detected by fluorescence microscopy
B16-OVA cells (2 × 106) were seeded on poly-D-lysine-coated glass coverslips and incubated in 6-well plates at 37°C, 5% CO2 for 24 h. Cells were further incubated in the presence of pSIN-IL2/AA-PEG-liposome lipoplexes or pSIN-IL2/PEG-liposome lipoplexes (DNA : DOTAP, 3.75 µg : 18.75 µg) in reduced growth medium for 1 h at 37°C. After the incubation, cells were washed twice with PBS and fixed in 3% paraformaldehyde for 20 min at room temperature. Cells were washed with PBS three times, and coverslips were mounted on slides using a mounting medium containing 4’,6-diamidino-2-phenylindole (DAPI) as a nuclear counter stain (vectashield H-1200) from Vector laboratories (Burlingame, CA). Cells were viewed using an Olympus BX53 Microscope with a DP72 digital camera (Olympus America, Inc., Center Valley, PA). Images were acquired using the CellSens dimension imaging software (Olympus America, Inc., Center Valley, PA).
Intracellular trafficking of rhodamine-labeled lipoplexes determined using confocal microscopy
B16-OVA cells (1 × 106 cells/well) were seeded in a 35 mm glass bottom dish (Mattek Corporation, Ashland, MA) and incubated overnight. Rhodamine-labeled liposomes (Rho-AA-PEG-liposomes and Rho-PEG-liposomes) were prepared by including rhodamine-labeled DOPE (Rho-DOPE, 1% w/w) in the liposomes. To study the intracellular localization of pSIN-IL2/Rho-AA-PEG-liposome lipoplexes or pSIN-IL2/Rho-PEG-liposome lipoplexes (DNA : DOTAP, 3.75 µg : 18.75 µg), the cells were incubated at 37°C with the lipoplexes, followed by the addition of 0.2 mM Hoechst 33342 (Ex/Em 345/478 nm) (AnaSpec Inc. Fremont, CA). The cells were viewed live at time points indicated, and single focus images were acquired using a Leica TCS-SP5 × Supercontinum confocal microscope with an oil immersion objective (63 × 1.4 NA) (Leica Microsystems GmbH, Mannheim, Germany). Images were processed using the NIH Image J software.
Animal studies
All animal studies were carried out following National Institutes of Health guidelines for animal care and use. The animal protocol was approved by the Institutional Animal Care and Use Committee at the University of Texas at Austin. Female C57BL/6 mice (18–20 g) were from Charles River laboratories, Inc. (Wilmington, MA). Mice were subcutaneously (s.c.) injected in the right flank with B16-OVA cells (5 × 105). We intended to use an animal model that recapitulates human cutaneous melanoma progression. The spontaneous C57BL/6-derived B16 melanoma cell line is well established and is a widely used tumor model. For an early stage efficacy test, the subcutaneously grafted B16 model is likely sufficient. When tumors reached an average diameter of 2–4 mm, the lipoplexes (DNA : DOTAP, 25 : 125 µg/day) were injected subcutaneously peritumorally (s.c., p.t.) into mice (n = 5–8) for 5 consecutive days.22 In one experiment, mice were also given four additional injections starting on day 15. Tumor size was measured using a digital caliper, and tumor volume was calculated using the following equation: tumor volume = (length × width2)/2.
Immune cell profiles
B16-OVA tumor-bearing mice (n = 5/group) were treated (s.c., p.t.) for 5 consecutive days with plasmid complexed with AA-PEG-liposomes, starting when tumors were 2–3 mm. Mice were sacrificed 24 h after the last treatment to collect peripheral blood and spleens. The peripheral blood was collected and immediately mixed with 125 mM EDTA. Peripheral blood lymphocytes (PBL) were isolated by density gradient centrifugation following manufacturer’s instruction (Lympholyte cell separation media, Cedar Lane, Hornby, Canada). Lymphocytes (1 × 106) were blocked with anti-mouse CD16/CD32 for 10 min, washed and further stained with a cocktail of antibodies containing PE-labeled anti-mouse CD4 (Clone RM4-4, cat# 12–0043, eBioscience, San Diego, CA), Pe-Cy5-labeled anti-mouse CD8a (Clone 53–6.7, cat# 15–0081, eBioscience), FITC-labeled anti-mouse CD69 (Clone H1.2F3, cat# 11–0691, eBioscience), and APC-labeled anti-mouse CD49b (Clone DX5, cat# 17–5971, eBioscience). Stained cells were analyzed using a flow cytometer (Guava Easycyte 8HT Flow cytometry System, Millipore, Hayward, CA). Data was analyzed using the FlowJo flow cytometry analysis software (Ashland, OR). Quadrants were positioned based on corresponding unstained samples.
Splenocytes were isolated from B16-OVA tumor-bearing mice following treatment with lipoplexes as previously described.31 Spleens were removed from each mouse and placed in 10 mM PBS containing 2% FBS. Spleens were homogenized in fresh PBS by passing through a cell strainer using a syringe plunger. Red blood cells were lysed by incubating the cell suspension with a red blood cell lysis buffer (Sigma) for 5 min at 4°C. Splenocytes were stained with the cocktail of antibodies and analyzed as mentioned above.
In vivo expression of IL2 in B16-OVA tumors
B16-OVA tumors were harvested from mice that had been treated for 5 days with plasmids complexed with AA-PEG-liposomes. Twenty-four hours after the last treatment, tumors were removed and flash frozen in liquid nitrogen and stored at −80°C until further analysis. Total RNA was isolated from tumors by homogenization using TRIzol reagent (Invitrogen) and RNeasy kit (Qiagen) according to manufacturer’s instructions. High capacity cDNA Reverse Transcription kit (Applied Biosystems, Foster City, CA) was utilized for the RT-PCR. Real-time PCR of IL-2 gene was carried out using the Power SYBR Green PCR Master Mix kit (Applied Biosystems, Foster City, CA) with the following primers: 5’-CCT GAG CAG GAT GGA GAA TTA CA-3’ (forward) and 5’-TCC AGA ACA TGC CGC AGA G-3’ (reverse). All samples were performed in triplicates, and normalized to β-actin (5’-TGT GAT GGT GGG AAT GGG TCA GAA-3’ (forward) and 5’-TGC CAC AGG ATT CCA TAC CCA AGA-3’ (reverse)). Data were analyzed using the Applied Biosystems ViiA™ 7 Software (Applied Biosystems).
Hematoxylin and eosin staining
B16 tumors in mice that were treated for 5 consecutive days with plasmids were collected, fixed in formalin, embedded in paraffin, and sectioned. Sections of 4 µm were stained with hematoxylin and eosin (H&E). Slides were scanned and images were acquired using the ScanScope XT (Aperio Technologies, Vista, CA).
Statistical analysis
Statistical analyses were performed using analysis of variance followed by Fisher’s protected least significant difference procedure. Mouse survival curves were compared using the Kaplan–Meier survival analysis (GraphPad Prism®, La Jolla, CA). The p-value was computed using the log-rank test (Mantel-Cox test) or Gehan-Breslow-Wilcoxon test. A p-value of < 0.05 (two-tail) was considered statistically significant.32
Results
Preparation and characterization of pSIN-IL2/anisamide-conjugated liposome lipoplexes
Anisamide-conjugated, PEGylated cationic liposomes (AA-PEG-liposomes) were prepared by mixing cationic DOTAP liposomes with anisamide-conjugated DSPE-PEG (2000). The final concentration of DOTAP in the liposomes was 10 mg/ml. The anisamide-free, PEGylated cationic liposomes (PEG-liposomes) were prepared similarly except that an equivalent amount of DSPE-PEG (2000) was used. The diameter and zeta potential of the AA-PEG-liposomes were 90.3 ± 0.1 nm and 31.1 ± 1.0 mV, respectively. The AA-free, PEG-liposomes were 97.8 ± 0.2 nm, with a zeta potential of 24.1 ± 0.4 mV (Fig. 1A).
Fig. 1. Physicochemical characteristics of lipoplexes prepared by complexing pSIN-IL2 with anisamide-conjugated PEG-liposomes.

(A). Mean diameters (black bars) and zeta potentials (white squares) of liposomes (LP), PEG-liposomes (PEG-LP), and AA-PEG-liposomes (AA-PEG-LP).
(B). Mean diameters (bars) and zeta potentials (squares) of pSIN-IL2-anisamide-PEG-liposome lipoplexes prepared at various ratios of DOTAP vs. DNA. Equal volumes of DNA (25 µg) solution and AA-PEG-liposome suspension were mixed and allowed to incubate at room temperature for at least 15 min (L = AA-PEG-LP alone).
(C). Uptake of pSIN-IL2 in lipoplexes by B16-OVA cells in culture. Cells (1 × 105/well) were incubated with fluorescein-labeled pSIN-IL2 complexed with AA-PEG-liposomes at various ratios for 1 h at 37°C (pSIN = pSIN-IL2 alone, LP = AA-PEG-LP alone).
(D). Overlay of the dynamic light scattering spectra of lipoplexes prepared by complexing AA-PEG-LP with pSIN-IL2 at 2:1 or 10:1 ratios (w/w), immediately after preparation (green) and 30 min (red) after incubation at 37°C in a simulated biological medium.
In A–C, data shown are mean ± S.E.M. (n = 4).
The following experiments were completed to identify the optimal ratio of pSIN-IL2 to liposomes in the lipoplexes. Various amounts of AA-PEG-liposomes were complexed with a fixed amount of pSIN-IL2 to form different pSIN-IL2/liposome lipoplexes. The particle diameters and the zeta potentials of the resultant lipoplexes are reported in Fig. 1B. At the ratio of 1:1 and 2:1 (DOTAP vs. pSIN-IL2, w/w), the lipoplexes appeared unstable and aggregated, whereas lipoplexes prepared at other ratios had smaller sizes around 150 nm (Fig. 1B).
In order to identify the liposomes (or DOTAP lipid) to DNA ratio that is optimal in transfecting the pSIN-IL2 plasmid into tumor cells, the uptake of pSIN-IL2 in various lipoplexes by the B16-OVA cells was evaluated. The weight ratio of 5:1 to 20:1 (DOTAP : DNA) were found optimum based on the high levels of cellular uptake (Fig. 1C). The stability of the lipoplexes at various ratios was evaluated in a simulated physiological medium. The lipoplexes prepared at 5:1 to 30:1 ratio were physically stable within the tested period (Fig. 1D). Therefore, the lipoplexes prepared with a DOTAP (in liposomes) to pSIN-IL2 ratio of 5:1 or 10:1 (w/w) were used for further studies.
In vitro uptake of pSIN-IL2 in targeted lipoplexes by B16-OVA melanoma cells
The uptake of fluorescein-labeled pSIN-IL2 by B16-OVA cells expressing high levels of sigma receptor was evaluated after the cells were incubated for 1 h with lipoplexes prepared with the AA-PEG-LP or PEG-LP. The uptake of the pSIN-IL2 in the lipoplexes prepared with AA-PEG-LP was significantly higher compared to that prepared with PEG-LP (Fig. 2A), a 50 % increase. To further investigate whether the uptake of the pSIN-IL2 in the lipoplexes produced IL2, we incubated the lipoplexes with B16-OVA cells and measured IL2 expression. The expression of IL2 was significantly higher in B16-OVA cells transfected with pSIN-IL2 complexed with the AA-PEG-LP than with the PEG-LP (Fig. 2B). Cells transfected with pSIN-IL2 complexed with the PEG-liposomes produced an elevated amount of IL2 as compared to cells transfected with pSIN-IL2 alone.
Fig. 2. In vitro uptake of pSIN-IL2 in AA-PEG-LP or PEG-LP lipoplexes by B16-OVA tumor cells and in vivo antitumor activities of pSIN-IL2 complexed with AA-PEG-LP or PEG-LP.

(A). B16-OVA cells (1 × 106/well) were incubated with fluorescein-labeled pSIN-IL2 (0.75 µg) complexed with AA-PEG-LP or PEG-LP (DOTAP, 3.75 µg) for 1 h at 37°C. Fluorescence intensity was measured at 492/518 nm (n = 6).
(B). B16-OVA cells (2.5 × 105/well) were incubated with pSIN-IL2 (1 µg) complexed with AA-PEG-LP or PEG-LP (DOTAP, 10 µg) for 24 h at 37°C. The supernatant was collected to determine IL2 concentration using a mouse IL2 ELISA kit (n = 3). In A–B, pDNA means pSIN-IL2, and the a, b, c indicate significant differences (p < 0.05).
(C). Caspase-3 activity in B16-OVA cells transfected cells with pSIN-IL2 complexed with PEG-LP or AA-PEG-LP. Cell lysates were incubated with assay substrate for 48 h to determine caspase-3 activity (n = 3). (*) The values of the pSIN-IL2/AA-PEG-LP and pSIN-IL2/PEG-LP are different (p < 0.05). In A–C, data reported are mean ± SEM.
(D). B16-OVA (5 × 105) cells were established in female C57BL/6 mice on day 0. Starting on day 3, mice were peritumorally injected with PBS, pSIN-IL2/AA-PEG-LP or pSIN-IL2/PEG-LP for 5 consecutive days. The dose of pSIN-IL2 was 25 µg/mouse/injetion, 125 µg of the DOTAP. * Starting from day 13, the values of the pSIN-IL2/AA-PEG-LP and pSIN-IL2/PEG-LP are different from each other (p < 0.05).
(E). Survival curves of mice in D (p = 0.04, pSIN-IL2/AA-PEG-LP vs. pSIN-IL2/PEG-LP, Gehan-Breslow-Wilcoxon test). The number of mice per group in D–E was 5 for PBS, 7 for pSIN-IL2/AA-PEG-LP and pSIN-IL2/PEG-LP.
Additionally, we also evaluated the pSIN-IL2 plasmid’s ability to induce tumor cell apoptosis when complexed with AA-PEG-LP or PEG-LP by determining caspase 3 activation. The B16-OVA cells were transfected with the pSIN-IL2 plasmid complexed to either PEG-LP or AA-PEG-LP liposomes for 48 h. The caspase-3 activity in cells transfected with pSIN-IL2 complexed with the AA-PEG-LP was significantly higher than in cells transfected with pSIN-IL2 plasmid complexed with the PEG-LP (Fig. 2C), indicating that the in vitro cytotoxicity of the pSIN-IL2 was also enhanced by complexing it with the sigma receptor-targeting anisamide-conjugated liposomes. The effects of the AA-PEG-LP and PEG-LP alone on the viability of the B16-OVA cells were not different after 24 h of incubation (data not shown).
The pSIN-IL2 plasmid was more effective at controlling the growth of B16-OVA tumor cells in mice when complexed with the AA-PEG-LP than with the PEG-LP
The antitumor activity of the pSIN-IL2 complexed with the AA-PEG-LP or PEG-LP was evaluated against B16-OVA tumors in vivo. The pSIN-IL2/AA-PEG-liposome lipoplexes were significantly more effective than the pSIN-IL2/PEG-liposome lipoplexes at controlling the growth of the tumors (Fig. 2D). Starting on day 13, tumors in mice that were treated with the pSIN- IL2/AA-PEG-liposome lipoplexes became significantly smaller than those treated with the pSIN-IL2/PEG-liposome lipoplexes (Fig. 2D). The median survival time for mice treated with sterile PBS was 18 d, 22 d for mice treated with the pSIN-IL2/PEG-liposomes lipoplexes, and 27 d for mice treated with the pSIN-IL2/AA-PEG-liposome lipoplexes (Fig. 2E). Gross toxicity from treatment with the lipoplexes was not observed in terms of body weight loss (data not shown). H&E staining of the skin sites that were injected with the lipoplexes (i.e., pSIN-IL2/AA-PEG-LP) or the liposomes alone (i.e., AA-PEG-LP) did not reveal any significant histopathological difference (Fig. S3).
Intracellular trafficking of lipoplexes in B16 melanoma cells
The cellular distribution of the lipoplexes was examined with fluorescein-labeled pSIN-IL2 complexed with either rhodamine-labeled AA-PEG-liposomes or rhodamine-labeled PEG-liposomes. We examined the distribution of pSIN-IL2, liposomes, and the lipoplexes by fluorescence microscopy (Fig. 3). The fluorescence emission from the fluorescein-labeled pSIN-IL2 is shown in green, rhodamine-labeled liposomes in red, and the emission from the lipoplexes in yellow. Overall lipoplexes prepared with AA-free liposomes had less internalization compared to lipoplexes prepared with AA-PEG-LP (Fig. 3). Disassociation of pSIN-IL2 from the Rho-PEG-LP occurred at 180 min whereas disassociation of pSIN-IL2 from the Rho-AA-PEG-LP occurred at 60 min.
Fig. 3. Fluorescence images of cells incubated with fluorescein-labeled pSIN-IL2-rhodamine-labeled liposome lipoplexes.

B16-OVA cells (2 × 106) were incubated with fluorescein-labeled pSIN-IL2 (FITC-pSIN-IL2) complexed with rhodamine-labeled AA-PEG-LP (Rho-AA-PEG-LP) or rhodamine-labeled PEG-LP (Rho-PEG-LP) (DNA : DOTAP, 3.75 µg : 18.75 µg) for up to 180 min at 37°C. Cell nuclei were stained with DAPI (blue) (bar = 50 µm).
Subcellular distribution of lipoplexes
We further evaluated the intracellular localization of the lipoplexes using confocal microscopy. The lipoplexes were prepared using rhodamine-labeled liposomes. The lipoplexes prepared with the PEG-LP were primarily located on the plasma membrane after 30 min of incubation (Fig. 4). By 180 min, a large portion of the lipoplexes remained near the plasma membrane with some penetration into the cytoplasm. The lipoplexes prepared with AA-PEG-LP had a significantly higher level of internalization at 30 min, compared with PEG-LP, with some particles remaining on the plasma membrane. By 180 min, the lipoplexes were located inside the cells, adjacent to the nucleus.
Fig. 4. Subcellular distribution of rhodamine-labeled lipoplexes.

B16-OVA cells (1 × 106) were incubated with pSIN-IL2 complexed with rhodamine-labeled AA-PEG-LP (Rho-AA-PEG-LP) or rhodamine-labeled PEG-LP (Rho-PEG-LP) (DNA : DOTAP, 3.75 µg : 18.75 µg) for up to 180 min at 37°C. The image in the white box is shown at a higher magnification in adjacent image on the right. Cell nuclei were stained with Hoechst (blue).
The pSIN-IL2 plasmid was more effective at controlling the growth of B16-OVA tumor cells in mice than the pSIN-β plasmid
To evaluate the extent at which incorporating IL2 gene into the pSIN-β plasmid will improve the antitumor activity of the pSIN-β plasmid, B16-OVA tumor cells were implanted in mice. When tumors reached an average of 3 mm, mice were treated with pSIN-IL2 or pSIN-β plasmid complexed with the AA-PEG-LP daily for five consecutive days, and four additional days starting on day 15 (post implantation) if the mice were still alive. Tumors grew significantly slower in mice treated with the pSIN-IL2 than in mice treated with the pSIN-β plasmid or the pIL2 plasmid (Fig. 5A). Mice treated with the pSIN-IL2 plasmid also survived significantly longer than those treated with pSIN-β or pIL2 (Fig. 5B). The antitumor activities of the pSIN-β and pIL2 were not different from each other, but were significantly different from the sterile PBS as a vehicle control (Fig. 5A).
Fig. 5. The pSIN-IL2 plasmid is more effective than the pSIN-β plasmid at controlling the growth of B16-OVA tumor cells in mice.
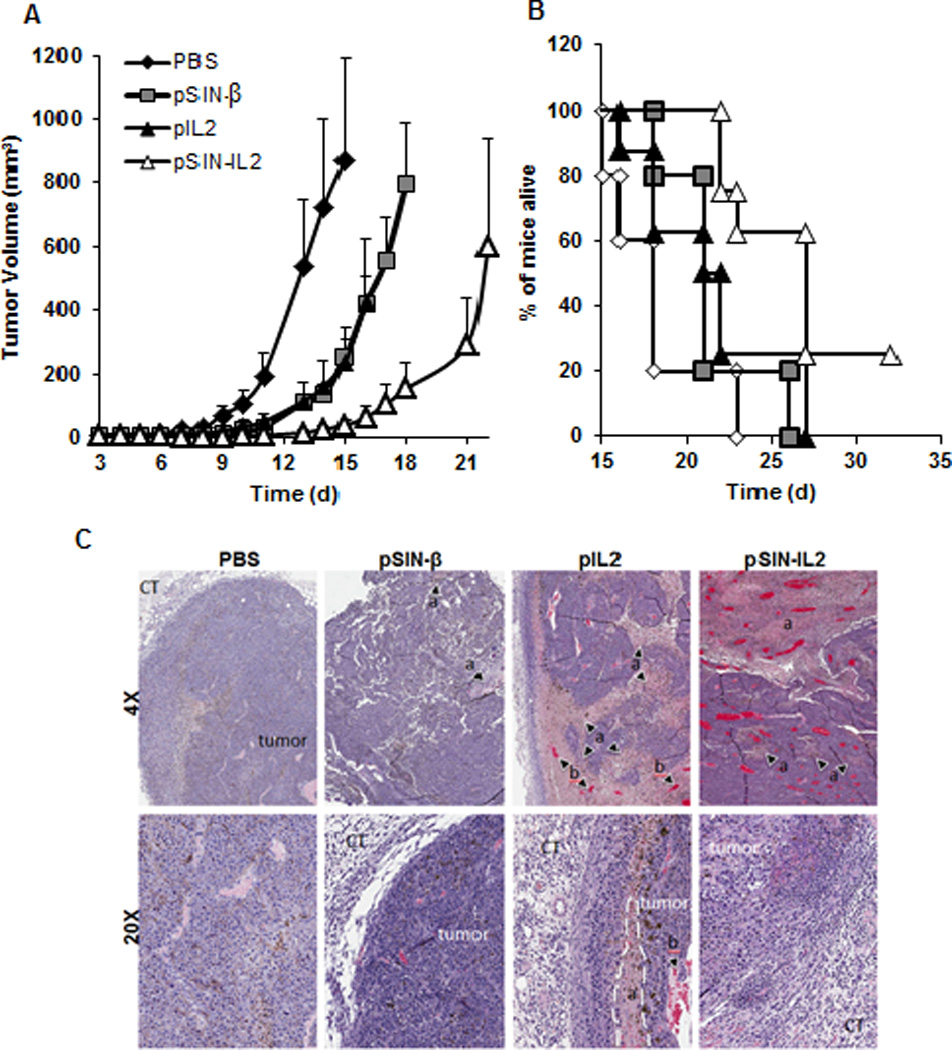
(A). B16-OVA (5 × 105) cells were established in female C57BL/6 mice (n = 5–8) on day 0. Starting on day 3, mice were peritumorally injected with pSIN-IL2, pSIN-β, or pIL2, all complexed with AA-PEG-LP for 5 consecutive days. Starting on day 15, all live mice were treated for an additional 4 consecutive days. Starting from day 7, the values of the pSIN-IL2/AA-PEG-LP and pSIN-β/AA-PEG-LP or pIL2/AA-PEG-LP are different from each other (p < 0.05). The values of pSIN-β/AA-PEG-LP and pIL2/AA-PEG-LP are not different from each other on any day. The number of mice used per group was 5 for PBS and pSIN-β/AA-PEG-LP, 8 for pIL2/AA-PEG-LP and pSIN-IL2/AA-PEG-LP. Data reported are mean ± SEM.
(B). Survival curves of mice in A (p = 0.005, pSIN-IL2 vs. pSIN-β; p = 0.03, pSIN-IL2 vs. pIL2; Log-rank Mantel-Cox test).
(C). H & E micrographs. In pSIN-β, (a) indicates necrotic areas. In pIL2, (a) indicates necrotic areas; (b) indicates hemorrhagic regions; and area within the dotted line indicates severe necrosis. In pSIN-IL2, (a) in dotted line indicates extensive central necrotic regions. Tumors were highly hemorrhagic (all red lacunas; note: despite the appearance in the low magnification picture, red blood cells were not contained by vessels), and there is a poorly defined border between connective tissue (CT) and the tumor per se.
Histology
B16-OVA tumors in mice that were treated with sterile PBS had a homogenous epithelial-like cellular appearance (Fig. 5C). Well-defined blood vessels were present, necrosis was rare, and hemorrhage was not observed. Further infiltration by inflammatory cells was minimal. Tumors from mice that were treated with pSIN-β have necrotic areas mainly on the periphery of the tumors, and inflammatory cells were found surrounding the mass. Tumors from mice that were treated with pIL2 plasmid showed extensive hemorrhagic regions and severe necrosis, and inflammatory cells including neutrophils and T cells were found on the periphery of the tumors with minor infiltration into the mass. Tumors in mice that were treated with pSIN-IL2 showed extensive central necrotic regions and were highly hemorrhagic. Severe inflammatory cell infiltration was found in both the center and the periphery of the tumors (Fig. 5C).
Treatment with pSIN-IL2 plasmid increases activated CD4+ T cells, CD8+ T cells, and NK cells in the peripheral blood and spleen of B16-OVA tumor-bearing mice
Peripheral blood and spleens were collected from B16-OVA tumor-bearing mice after 5 treatments with different plasmids. Isolated lymphocytes and splenocytes were stained with a cocktail of antibodies containing anti-mouse CD4, anti-mouse CD8a, anti-mouse CD69, and anti-mouse CD49b. CD49 is a marker for cell activation.33 The percentage of CD49+CD69+ cells in the PBL of mice treated with the pSIN-IL2 was 8.6 times higher than in mice treated with PBS, and 2.1 times higher than in the mice treated with pSIN-β (Fig. 6A). The percentage of CD4+CD69+ and CD8+CD69+ cells in PBL of mice treated with pSIN-IL2 was 1.6-fold and 3-fold higher than in mice treated with the pSIN-β , respectively (Fig. 6A). The percentages of CD4+CD69+, CD8+CD69+, and CD49+CD69+ cells in PBL in mice treated with pSIN-IL2 were not significantly different from those in mice treated with the pIL2 plasmid (Fig. 6A). A similar trend was observed for the CD4+CD69+, CD8+CD69+, and CD49+CD69+ cells in the splenocytes of the treated mice (Fig. 6C, D). Finally, the total CD4+, CD8+, and CD49+ cells were not different among all treatment groups in the PBL and splenocyte samples (data not shown).
Fig. 6. Activation of CD8+ T cells, CD4+ T cells, and NK cells in the peripheral blood and spleens of tumor-bearing mice after treatment with pSIN-IL2 complexed with AA-PEG-LP.


B16-OVA tumor-bearing mice (n = 5) were treated for 5 consecutive days with plasmid complexed with AA-PEG-liposomes, starting when tumors reached 2–3 mm in diameter. Mice were sacrificed 24 h after the last treatment to collect peripheral blood and spleens.
(A). Percentage of CD4+CD69+, CD8+CD69+, and CD49+CD69+ lymphocytes in the PBL of mice treated with PBS, pSIN-β, pIL2, or pSIN-IL2. All plasmids were complexed with AA-PEG-LP. (*) the values of the pSIN-IL2 and pSIN-β are different from each other (p < 0.05).
(B). Representative dot plots indicating the percentages of activated lymphocytes in PBL of mice treated with PBS, pSIN-β, pIL2, or pSIN-IL2.
(C). Percentage of CD4+CD69+, CD8+CD69+, and CD49+CD69+ splenocytes of mice treated with PBS, pSIN-β, pIL2, or pSIN-IL2. (*) the values of the pSIN-IL2 and pSIN-β are different from each other (p < 0.05).
(D). Representative dot plots indicating percentage of activated splenocytes of mice treated with PBS, pSIN-β, pIL2, or pSIN-IL2. In A and C, data shown are mean ± SEM (n = 4–5).
(E)In vivo expression of IL2 in B16-OVA tumors. Twenty hours after the last treatment with PBS, or pSIN-β, pIL2, or pSIN-IL2, all complexed with AA-PEG-LP, tumors were collected. Total RNA was extracted from tumor tissues to quantify IL2 expression using qRT-PCR. Data shown are mean ± SEM (n = 3–4). Different letters indicate significant differences (p < 0.05).
In vivo expression of IL2 in B16-OVA tumors
As shown in Fig. 6E, a significant amount of IL2 mRNA was detected in B16-OVA tumors in mice that were treated with pSIN-IL2, but not in tumors in mice that were treated with pSIN-β. A similar trend for IL2 protein level was observed in blood, although not statistically different, likely due to the smaller sample size of the blood samples (data not shown).
Discussion
We have previously reported that the RNA replicase-based plasmid, pSIN-β, has a strong antitumor activity by enabling tumor cells transfected with it to produce dsRNA, which is pro-apoptotic and activates innate immunity.16, 22 The pSIN-IL2 multigene plasmid construct used in the present study represents an improvement over the pSIN-β plasmid for melanoma treatment. In order to improve the antitumor activity of the pSIN-β, we cloned mouse IL2 into it. A high level of IL2 was detected in cells transfected with pSIN-IL2 (Fig. S2A), and the level of IL2 produced by the pSIN-IL2 transfected cells was dependent on the dose of the pSIN-IL2 used (Fig. S2B). The inclusion of the IL2 gene in the pSIN-β plasmid enabled the plasmid to effectively activate T cells and NK cells in mice (Fig. 6), which may explain the significantly stronger antitumor activity of the pSIN-IL2 plasmid in B16-OVA tumor-bearing mice, compared to the pSIN-β (Fig. 5).
The antitumor activity of dsRNA is well documented.16, 18, 34–36 Polyriboinosinic-polyribocytidylic acid (poly I:C), a synthetic dsRNA, has been used as the standard to evaluate the effect of dsRNA in many animal models as well as in clinical trials.15, 16, 35, 37 Poly (I:C) was found to slightly delay tumor growth, and it is not practical to increase its antitumor activity by increasing its dose because of the dose-limiting adverse effects.37–39 We have previously reported that the dsRNA-producing pSIN-β plasmid significantly inhibits tumor cell growth both in vitro and in vivo,22 likely due to the production of the pro-apoptotic and immunostimulatory dsRNA by tumor cells transfected with the pSIN-β plasmid.22
IL2 is approved for use in the treatment of advanced melanoma due to its ability to modulate immune response and stimulate the activation and proliferation of T-cells and NK cells.4 In the present study, we tested whether including IL2 gene into the pSIN-β plasmid will improve the resultant antitumor activity. We found that treatment of tumor-bearing mice with the pSIN-IL2 plasmid significantly increased the percentages of activated CD4+ T cells, CD8+ T cells, and CD49+ NK cells in peripheral blood and splenocytes of the mice, as compared to treatment with the pSIN-β plasmid (Fig. 6A, C), indicating that the IL2 gene in the pSIN-IL2 plasmid had helped to incite the activation of circulating T cells and NK cells (Fig. 6). H&E micrographs also showed that the pSIN-IL2 plasmid induced an elevated number of infiltrating immune cells both in the center and in the periphery of the tumors, as compared to pIL2 or pSIN-β (Fig. 5C). The activation of the T cells and NK cells by the IL2 may explain the enhanced antitumor activity from the pSIN-IL2 (Fig. 5). It is known that IL2 regulates the Treg lymphocyte balance, which suppresses antitumor immune responses.40, 41 Yao and colleagues showed that treatment of mice with an IL2-expressing plasmid led to a reduction of Foxp3+ Tregs populations in peripheral blood.42 We found high levels of IL6, a proinflammatory cytokine, in the serum samples of mice that were treated with pSIN-IL2 (data not shown). It was shown that IL6 decrease Foxp3 expression and down-regulates Tregs in vitro and in vivo,43, 44 which may also explain the pSIN-IL2 plasmid’s ability to activate T cells and NK cells. In our studies, the median survival time for mice treated with sterile PBS was 18 days, 22 days for mice treated with the pSIN-IL2/PEG-liposomes lipoplexes, and 27 days for mice treated with the pSIN-IL2/AA-PEG-liposome lipoplexes (Fig. 2E). The B16 melanoma cells are very aggressive. Some of the tumor-bearing mice that were left untreated had to be euthanized as early as two weeks after injection of tumor cells (5 × 105 per mouse). Therefore, the 9-day extension of overall survival time by treatment with the pSIN-IL2/AA-PEG-liposome lipoplexes is significant. Patients with advanced metastatic melanoma have a life expectancy of 6–10 months.2, 45 Any improvement in life expectancy and/or quality of life is beneficial to patients. It is difficult to predict the clinical performance of our pSIN-IL2 plasmid without conducting a clinical trial as human conditions are typically far more complex. However, we expect that the pSIN-IL2 construct can be given to patients as an adjuvant therapy to improve the life expectancy and/or quality of life of the patients.
The present study is also designed to actively target the RNA replicase-based IL2- expressing plasmid into tumor cells more specifically, therefore allowing the tumor cells to preferentially take up the plasmid, generate intracellular dsRNA and IL2, and then undergo apoptosis by committing “suicide”. We chose to target the sigma receptor because it is overexpressed in many tumor cells.24, 46 Radiolabeled sigma-receptor ligands have been extensively used as imaging agents in melanoma,47 breast cancer,48 and prostate cancer,49, 50 In the present study, anisamide was chemically conjugated onto the cationic liposomal carrier, which was then complexed with the pSIN-IL2 plasmid to target it into sigma receptor-overexpressing murine tumor cells in vitro and in vivo. Data from previous in vitro and in vivo studies showed that anisamide has the similar binding affinity to sigma receptors as I123-N-(2-diethyl aminoethyl) 4-iodobenzamide (I123-IDAB), a radiolabeled benzamide, used to visualize sigma receptors in vivo.51, 52 B16-OVA cells were derived from the highly aggressive B16-F10 mouse melanoma cells.53 They express high level of sigma receptors.54 Our in vitro data showed that the anisamide-conjugated cationic liposomes more effectively delivered the pSIN-IL2 plasmid into the B16-OVA tumor cells than the anisamide-free liposomes (Fig. 2A). Importantly, pSIN-IL2 complexed with the anisamide-conjugated liposomes also more effectively controlled B16-OVA tumor growth in mice than pSIN-IL2 complexed with the anisamide-free liposomes (Fig. 2D–E), confirming the feasibility of using anisamide or similar sigma receptor ligands for future targeted gene therapy of melanoma.
Understanding the uptake mechanism and intracellular distribution of plasmid liposome complexes is important in order to design optimal gene delivery systems. The uptake of pSIN-IL2/Rho-AA-PEG-LP complexes were much higher than pSIN-IL2/Rho-PEG-LP after incubation at 30 min (Fig. 4). The pSIN-IL2/Rho-PEG-LP complexes appear on the cell surface; with minor internalization at 180 min. The anisamide conjugated on the surface of the liposomes increased the interaction of the lipoplexes with cell membrane as well as the total amount of the lipoplexes entering the cytosol. There is strong correlation between the cellular uptake of complexes and the transfection efficiency of the pSIN-IL2/AA-PEG-LP complexes (Fig. 2). Interestingly, we observed that the dissociation of pSIN-IL2 from the pSIN-IL2/Rho-AA-PEG-LP lipoplexes occurred as early as 60 min after the addition of the lipoplexes into the cell culture, whereas the dissociation of pSIN-IL2 from the pSIN-IL2/Rho-PEG-LP lipoplexes did not occur until around 180 min later (Fig. 3). Dissociation of the plasmid from the lipoplexes is critical for nuclear translocation of the plasmid. The anisamide on the lipoplexes facilitated the uptake of the lipoplexes by the B16-OVA cells and thereby increased the expression of the IL2 gene (Fig. 2B).
Similar to other cytotoxic anticancer drugs, the pSIN-IL2 plasmid is also expected to be cytotoxic to non-tumor cells if they are transfected, because the dsRNA produced inside the transfected cells will be pro-apoptotic.23, 55 In the present study, we took advantage of the overexpression of sigma-receptor by the B16 melanoma cells to increase the uptake of the pSIN-IL2 plasmid by the tumor cells to more effectively kill the tumor cells. Due to the difficulty in directly injecting a sufficient volume of lipoplexes into tumors in mice, especially when tumors were small, we injected the anisamide-conjugated pSIN-IL2 lipoplexes peritumorally in the subcutaneous space. It is therefore likely that non-cancer cells close to the tumors may have also been transfected. However, we expect that the tumor cells had taken up the lipoplexes more efficiently because they overexpress sigma-receptor. We observed an elevated IL2 mRNA expression in tumor tissues 24 h after peritumoral injection of the pSIN-IL2 plasmid (Fig. 6E), clearly indicating that the lipoplexes were taken up by the B16 tumor cells, and the B16 tumor cells expressed the IL2 gene. In future clinical trials, the pSIN-IL2 plasmid can certainly be injected directly into tumor tissues in patients to further minimize its uptake by non-tumor cells.
In conclusion, we reported a novel strategy to further improve the antitumor activity the RNA replicase-based pSIN-β plasmid by including an IL2 gene in the plasmid to boost the antitumor immunity by activating T cells and NK cells. We also confirmed the sigma receptor as a potential target for future targeted gene therapy of melanoma.
Supplementary Material
1_si_001
Acknowledgement
This work was supported in part by National Cancer Institute grants CA135274 (to ZC) and CA135274-S1 (to BLR and ZC). We would like to thank Dr. Yue Li at the UT-Austin DPRI Microscopy Core Facility for assistance in acquiring the confocal microscopic images. We would also like to thank Dr. Irena Fernandez at the DPRI Flow Cytometry Core Facility for her invaluable assistance with the flow cytometry data analysis.
Footnotes
Supporting Information Available:
A map of the pSIN-IL2 plasmid (Fig. S1), the expression of IL2 by B16-OVA cells transfected with pSIN-IL2 using Lipofectamine® (Fig. S2), and the H&E histological images of the skin sites injected with pSIN-IL2/AA-PEG-LP (Fig. S3) are included in the Supporting Information. This information is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Blank CU, Hooijkaas AI, Haanen JB, Schumacher TN. Combination of targeted therapy and immunotherapy in melanoma. Cancer Immunol Immunother. 2011;60(10):1359–1371. doi: 10.1007/s00262-011-1079-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Puzanov I, Flaherty KT. Targeted molecular therapy in melanoma. Semin Cutan Med Surg. 2010;29(3):196–201. doi: 10.1016/j.sder.2010.06.005. [DOI] [PubMed] [Google Scholar]
- 3.Foa R, Guarini A, Gansbacher B. IL2 treatment for cancer: from biology to gene therapy. Br J Cancer. 1992;66(6):992–998. doi: 10.1038/bjc.1992.400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Riker AI, Radfar S, Liu S, Wang Y, Khong HT. Immunotherapy of melanoma: a critical review of current concepts and future strategies. Expert Opin Biol Ther. 2007;7(3):345–358. doi: 10.1517/14712598.7.3.345. [DOI] [PubMed] [Google Scholar]
- 5.Rosenberg SA, Yang JC, White DE, Steinberg SM. Durability of complete responses in patients with metastatic cancer treated with high-dose interleukin-2: identification of the antigens mediating response. Ann Surg. 1998;228(3):307–319. doi: 10.1097/00000658-199809000-00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ali G, Boldrini L, Lucchi M, Picchi A, Dell'Omodarme M, Prati MC, Mussi A, Corsi V, Fontanini G. Treatment with interleukin-2 in malignant pleural mesothelioma: immunological and angiogenetic assessment and prognostic impact. Br J Cancer. 2009;101(11):1869–1875. doi: 10.1038/sj.bjc.6605438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jackaman C, Bundell CS, Kinnear BF, Smith AM, Filion P, van Hagen D, Robinson BW, Nelson DJ. IL-2 intratumoral immunotherapy enhances CD8+ T cells that mediate destruction of tumor cells and tumor-associated vasculature: a novel mechanism for IL-2. J Immunol. 2003;171(10):5051–5063. doi: 10.4049/jimmunol.171.10.5051. [DOI] [PubMed] [Google Scholar]
- 8.West WH, Tauer KW, Yannelli JR, Marshall GD, Orr DW, Thurman GB, Oldham RK. Constant-infusion recombinant interleukin-2 in adoptive immunotherapy of advanced cancer. N Engl J Med. 1987;316(15):898–905. doi: 10.1056/NEJM198704093161502. [DOI] [PubMed] [Google Scholar]
- 9.Cortesina G, De Stefani A, Giovarelli M, Barioglio MG, Cavallo GP, Jemma C, Forni G. Treatment of recurrent squamous cell carcinoma of the head and neck with low doses of interleukin-2 injected perilymphatically. Cancer. 1988;62(12):2482–2485. doi: 10.1002/1097-0142(19881215)62:12<2482::aid-cncr2820621205>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 10.Belldegrun A, Tso CL, Zisman A, Naitoh J, Said J, Pantuck AJ, Hinkel A, deKernion J, Figlin R. Interleukin 2 gene therapy for prostate cancer: phase I clinical trial and basic biology. Hum Gene Ther. 2001;12(8):883–892. doi: 10.1089/104303401750195854. [DOI] [PubMed] [Google Scholar]
- 11.Galanis E, Burch PA, Richardson RL, Lewis B, Pitot HC, Frytak S, Spier C, Akporiaye ET, Peethambaram PP, Kaur JS, Okuno SH, Unni KK, Rubin J. Intratumoral administration of a 1,2-dimyristyloxypropyl-3-dimethylhydroxyethyl ammonium bromide/dioleoylphosphatidylethanolamine formulation of the human interleukin-2 gene in the treatment of metastatic renal cell carcinoma. Cancer. 2004;101(11):2557–2566. doi: 10.1002/cncr.20653. [DOI] [PubMed] [Google Scholar]
- 12.Galanis E, Hersh EM, Stopeck AT, Gonzalez R, Burch P, Spier C, Akporiaye ET, Rinehart JJ, Edmonson J, Sobol RE, Forscher C, Sondak VK, Lewis BD, Unger EC, O'Driscoll M, Selk L, Rubin J. Immunotherapy of advanced malignancy by direct gene transfer of an interleukin-2 DNA/DMRIE/DOPE lipid complex: phase I/II experience. J Clin Oncol. 1999;17(10):3313–3323. doi: 10.1200/JCO.1999.17.10.3313. [DOI] [PubMed] [Google Scholar]
- 13.Fearon ER, Pardoll DM, Itaya T, Golumbek P, Levitsky HI, Simons JW, Karasuyama H, Vogelstein B, Frost P. Interleukin-2 production by tumor cells bypasses T helper function in the generation of an antitumor response. Cell. 1990;60(3):397–403. doi: 10.1016/0092-8674(90)90591-2. [DOI] [PubMed] [Google Scholar]
- 14.Zier KS, Gansbacher B. IL-2 gene therapy of solid tumors: an approach for the prevention of signal transduction defects in T cells. J Mol Med (Berl) 1996;74(3):127–134. doi: 10.1007/BF01575444. [DOI] [PubMed] [Google Scholar]
- 15.Fujimura T, Nakagawa S, Ohtani T, Ito Y, Aiba S. Inhibitory effect of the polyinosinic-polycytidylic acid/cationic liposome on the progression of murine B16F10 melanoma. Eur J Immunol. 2006;36(12):3371–3380. doi: 10.1002/eji.200636053. [DOI] [PubMed] [Google Scholar]
- 16.Absher M, Stinebring WR. Toxic properties of a synthetic double-stranded RNA. Endotoxin-like properties of poly Ipoly C, an interferon stimulator. Nature. 1969;223(5207):715–717. doi: 10.1038/223715a0. [DOI] [PubMed] [Google Scholar]
- 17.Chawla-Sarkar M, Lindner DJ, Liu YF, Williams BR, Sen GC, Silverman RH, Borden EC. Apoptosis and interferons: role of interferon-stimulated genes as mediators of apoptosis. Apoptosis. 2003;8(3):237–249. doi: 10.1023/a:1023668705040. [DOI] [PubMed] [Google Scholar]
- 18.Friedrich I, Shir A, Klein S, Levitzki A. RNA molecules as anti-cancer agents. Semin Cancer Biol. 2004;14(4):223–230. doi: 10.1016/j.semcancer.2004.04.001. [DOI] [PubMed] [Google Scholar]
- 19.Schulz O, Diebold SS, Chen M, Naslund TI, Nolte MA, Alexopoulou L, Azuma YT, Flavell RA, Liljestrom P, Reis e Sousa C. Toll-like receptor 3 promotes cross-priming to virus-infected cells. Nature. 2005;433(7028):887–892. doi: 10.1038/nature03326. [DOI] [PubMed] [Google Scholar]
- 20.Cui Z, Le UM, Qiu F, Shaker DS. Learning from viruses: the necrotic bodies of tumor cells with intracellular synthetic dsRNA induced strong anti-tumor immune responses. Pharm Res. 2007;24(9):1645–1652. doi: 10.1007/s11095-007-9293-5. [DOI] [PubMed] [Google Scholar]
- 21.McBride S, Hoebe K, Georgel P, Janssen E. Cell-associated double-stranded RNA enhances antitumor activity through the production of type I IFN. J Immunol. 2006;177(9):6122–6128. doi: 10.4049/jimmunol.177.9.6122. [DOI] [PubMed] [Google Scholar]
- 22.Rodriguez BL, Yu Z, Chung WG, Weiss R, Cui Z. Replicase-based plasmid DNA shows anti-tumor activity. BMC Cancer. 2011;11:110. doi: 10.1186/1471-2407-11-110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rodriguez BL, Li X, Kiguchi K, DiGiovanni J, Unger EC, Cui Z. Control of solid tumor growth in mice using EGF receptor-targeted RNA replicase-based plasmid DNA. Nanomedicine (Lond) 2012;7(4):475–491. doi: 10.2217/nnm.11.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vilner BJ, John CS, Bowen WD. Sigma-1 and sigma-2 receptors are expressed in a wide variety of human and rodent tumor cell lines. Cancer Res. 1995;55(2):408–413. [PubMed] [Google Scholar]
- 25.Hou C, Tu Z, Mach R, Kung HF, Kung MP. Characterization of a novel iodinated sigma-2 receptor ligand as a cell proliferation marker. Nucl Med Biol. 2006;33(2):203–209. doi: 10.1016/j.nucmedbio.2005.10.001. [DOI] [PubMed] [Google Scholar]
- 26.Banerjee R, Tyagi P, Li S, Huang L. Anisamide-targeted stealth liposomes: a potent carrier for targeting doxorubicin to human prostate cancer cells. Int J Cancer. 2004;112(4):693–700. doi: 10.1002/ijc.20452. [DOI] [PubMed] [Google Scholar]
- 27.Chen Y, Bathula SR, Yang Q, Huang L. Targeted nanoparticles deliver siRNA to melanoma. J Invest Dermatol. 2010;130(12):2790–2798. doi: 10.1038/jid.2010.222. [DOI] [PubMed] [Google Scholar]
- 28.Le UM, Cui Z. Long-circulating gadolinium-encapsulated liposomes for potential application in tumor neutron capture therapy. Int J Pharm. 2006;312(1–2):105–112. doi: 10.1016/j.ijpharm.2006.01.002. [DOI] [PubMed] [Google Scholar]
- 29.Scheiblhofer S, Weiss R, Gabler M, Leitner WW, Thalhamer J. Replicase-based DNA vaccines for allergy treatment. Methods Mol Med. 2006;127:221–235. doi: 10.1385/1-59745-168-1:221. [DOI] [PubMed] [Google Scholar]
- 30.Sloat BR, Sandoval MA, Li D, Chung WG, Lansakara PD, Proteau PJ, Kiguchi K, DiGiovanni J, Cui Z. In vitro and in vivo anti-tumor activities of a gemcitabine derivative carried by nanoparticles. Int J Pharm. 2011;409(1–2):278–288. doi: 10.1016/j.ijpharm.2011.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cui Z, Qiu F. Synthetic double-stranded RNA poly(I:C) as a potent peptide vaccine adjuvant: therapeutic activity against human cervical cancer in a rodent model. Cancer Immunol Immunother. 2006;55(10):1267–1279. doi: 10.1007/s00262-005-0114-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Festing MF, Altman DG. Guidelines for the design and statistical analysis of experiments using laboratory animals. ILAR J. 2002;43(4):244–258. doi: 10.1093/ilar.43.4.244. [DOI] [PubMed] [Google Scholar]
- 33.Ziegler SF, Ramsdell F, Alderson MR. The activation antigen CD69. Stem Cells. 1994;12(5):456–465. doi: 10.1002/stem.5530120502. [DOI] [PubMed] [Google Scholar]
- 34.Hirabayashi K, Yano J, Inoue T, Yamaguchi T, Tanigawara K, Smyth GE, Ishiyama K, Ohgi T, Kimura K, Irimura T. Inhibition of cancer cell growth by polyinosinic-polycytidylic acid/cationic liposome complex: a new biological activity. Cancer Res. 1999;59(17):4325–4333. [PubMed] [Google Scholar]
- 35.Le UM, Kaurin DG, Sloat BR, Yanasarn N, Cui Z. Localized irradiation of tumors prior to synthetic dsRNA therapy enhanced the resultant anti-tumor activity. Radiother Oncol. 2009;90(2):273–279. doi: 10.1016/j.radonc.2008.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Le UM, Yanasarn N, Lohr CV, Fischer KA, Cui Z. Tumor chemo-immunotherapy using gemcitabine and a synthetic dsRNA. Cancer Biol Ther. 2008;7(3):440–447. doi: 10.4161/cbt.7.3.5423. [DOI] [PubMed] [Google Scholar]
- 37.Okada C, Akbar SM, Horiike N, Onji M. Early development of primary biliary cirrhosis in female C57BL/6 mice because of poly I:C administration. Liver Int. 2005;25(3):595–603. doi: 10.1111/j.1478-3231.2005.01043.x. [DOI] [PubMed] [Google Scholar]
- 38.Pimm MV, Baldwin RW. Treatment of transplanted rat tumours with double-stranded RNA(BRL 5907). IITreatment of pleural and peritoneal growths. Br J Cancer. 1976;33(2):166–171. doi: 10.1038/bjc.1976.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Meier H, Myers DD, Huebner RJ. Ineffectiveness of poly rI:rC on transplanted tumors induced by methylcholanthrene. Naturwissenschaften. 1970;57(5):248–249. doi: 10.1007/BF01010279. [DOI] [PubMed] [Google Scholar]
- 40.Shimizu J, Yamazaki S, Sakaguchi S. Induction of tumor immunity by removing CD25+CD4+ T cells: a common basis between tumor immunity and autoimmunity. J Immunol. 1999;163(10):5211–5218. [PubMed] [Google Scholar]
- 41.Tanaka H, Tanaka J, Kjaergaard J, Shu S. Depletion of CD4+ CD25+ regulatory cells augments the generation of specific immune T cells in tumor-draining lymph nodes. J Immunother. 2002;25(3):207–217. doi: 10.1097/00002371-200205000-00003. [DOI] [PubMed] [Google Scholar]
- 42.Yao H, Ng SS, Huo LF, Chow BK, Shen Z, Yang M, Sze J, Ko O, Li M, Yue A, Lu LW, Bian XW, Kung HF, Lin MC. Effective melanoma immunotherapy with interleukin-2 delivered by a novel polymeric nanoparticle. Mol Cancer Ther. 2011;10(6):1082–1092. doi: 10.1158/1535-7163.MCT-10-0717. [DOI] [PubMed] [Google Scholar]
- 43.Lal G, Zhang N, van der Touw W, Ding Y, Ju W, Bottinger EP, Reid SP, Levy DE, Bromberg JS. Epigenetic regulation of Foxp3 expression in regulatory T cells by DNA methylation. J Immunol. 2009;182(1):259–273. doi: 10.4049/jimmunol.182.1.259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Korn T, Mitsdoerffer M, Croxford AL, Awasthi A, Dardalhon VA, Galileos G, Vollmar P, Stritesky GL, Kaplan MH, Waisman A, Kuchroo VK, Oukka M. IL-6 controls Th17 immunity in vivo by inhibiting the conversion of conventional T cells into Foxp3+ regulatory T cells. Proc Natl Acad Sci U S A. 2008;105(47):18460–18465. doi: 10.1073/pnas.0809850105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Di Giacomo AM, Ascierto PA, Pilla L, Santinami M, Ferrucci PF, Giannarelli D, Marasco A, Rivoltini L, Simeone E, Nicoletti SV, Fonsatti E, Annesi D, Queirolo P, Testori A, Ridolfi R, Parmiani G, Maio M. Ipilimumab and fotemustine in patients with advanced melanoma (NIBIT-M1): an open-label, single-arm phase 2 trial. Lancet Oncol. 2012;13(9):879–886. doi: 10.1016/S1470-2045(12)70324-8. [DOI] [PubMed] [Google Scholar]
- 46.Bem WT, Thomas GE, Mamone JY, Homan SM, Levy BK, Johnson FE, Coscia CJ. Overexpression of sigma receptors in nonneural human tumors. Cancer Res. 1991;51(24):6558–6562. [PubMed] [Google Scholar]
- 47.John CS, Bowen WD, Saga T, Kinuya S, Vilner BJ, Baumgold J, Paik CH, Reba RC, Neumann RD, Varma VM, et al. A malignant melanoma imaging agent: synthesis, characterization, in vitro binding and biodistribution of iodine-125-(2-piperidinylaminoethyl)4-iodobenzamide. J Nucl Med. 1993;34(12):2169–2175. [PubMed] [Google Scholar]
- 48.Caveliers V, Everaert H, John CS, Lahoutte T, Bossuyt A. Sigma receptor scintigraphy with N-[2-(1'-piperidinyl)ethyl]-3-(123)I-iodo-4-methoxybenzamide of patients with suspected primary breast cancer: first clinical results. J Nucl Med. 2002;43(12):1647–1649. [PubMed] [Google Scholar]
- 49.John CS, Vilner BJ, Geyer BC, Moody T, Bowen WD. Targeting sigma receptor-binding benzamides as in vivo diagnostic and therapeutic agents for human prostate tumors. Cancer Res. 1999;59(18):4578–4583. [PubMed] [Google Scholar]
- 50.Kashiwagi H, McDunn JE, Simon PO, Jr., Goedegebuure PS, Xu J, Jones L, Chang K, Johnston F, Trinkaus K, Hotchkiss RS, Mach RH, Hawkins WG. Selective sigma-2 ligands preferentially bind to pancreatic adenocarcinomas: applications in diagnostic imaging and therapy. Mol Cancer. 2007;6:48. doi: 10.1186/1476-4598-6-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Megalizzi V, Le Mercier M, Decaestecker C. Sigma receptors and their ligands in cancer biology: overview and new perspectives for cancer therapy. Med Res Rev. 2012;32(2):410–427. doi: 10.1002/med.20218. [DOI] [PubMed] [Google Scholar]
- 52.Everaert H, Flamen P, Franken PR, Verhaeghe W, Bossuyt A. Sigma-receptor imaging by means of I123-IDAB scintigraphy: clinical application in melanoma and non-small cell lung cancer. Anticancer Res. 1997;17(3B):1577–1582. [PubMed] [Google Scholar]
- 53.Brown DM, Fisher TL, Wei C, Frelinger JG, Lord EM. Tumours can act as adjuvants for humoral immunity. Immunology. 2001;102(4):486–497. doi: 10.1046/j.1365-2567.2001.01213.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Li SD, Chono S, Huang L. Efficient oncogene silencing and metastasis inhibition via systemic delivery of siRNA. Mol Ther. 2008;16(5):942–946. doi: 10.1038/mt.2008.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Leitner WW, Hwang LN, Bergmann-Leitner ES, Finkelstein SE, Frank S, Restifo NP. Apoptosis is essential for the increased efficacy of alphaviral replicase-based DNA vaccines. Vaccine. 2004;22(11–12):1537–1544. doi: 10.1016/j.vaccine.2003.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
1_si_001