Magnetic orientation of garden warblers (Sylvia borin) under 1.4 MHz radiofrequency magnetic field (original) (raw)
Abstract
We report on the experiments on orientation of a migratory songbird, the garden warbler (Sylvia borin), during the autumn migration period on the Courish Spit, Eastern Baltics. Birds in experimental cages, deprived of visual information, showed the seasonally appropriate direction of intended flight with respect to the magnetic meridian. Weak radiofrequency (RF) magnetic field (190 nT at 1.4 MHz) disrupted this orientation ability. These results may be considered as an independent replication of earlier experiments, performed by the group of R. and W. Wiltschko with European robins (Erithacus rubecula). Confirmed outstanding sensitivity of the birds' magnetic compass to RF fields in the lower megahertz range demands for a revision of one of the mainstream theories of magnetoreception, the radical-pair model of birds' magnetic compass.
Keywords: orientation, magnetoreception, radiofrequency fields, radical-pair model
1. Introduction
The question as to how migrating animals find their way over great expanses of land and sea is one of the hottest topics of modern science [1]. The development of tracking technologies, including satellite-based, uncovers details of these seasonal movements that were beyond imagination just a decade ago—such as the 8 day non-stop flight of bar-tailed godwits (Limosa lapponica baueri) across the Pacific Ocean [2]. Such long-distance travels require a precise and reliable navigation system. Biophysical, physiological and neural bases of animal navigation are so far rather poorly understood, though, in certain directions, remarkable progress has been achieved. One of those is related to compass orientation in migratory birds.
Owing to conspicuous changes in physiology and behaviour, demonstrated by many bird species during the migration period, and to their obvious motivation for direction finding, birds can be used as an object for studying navigation abilities under controlled experimental conditions in the laboratory. This way, birds were shown to have at least three mechanisms of direction finding, namely the sun compass [3,4], stellar compass [5–8] and the magnetic compass [9]. The latter is especially interesting, because the well-documented existence of the magnetic compass in birds remains one of few unambiguous demonstrations of the elusive magnetic sense of animals, the physical and physiological origins of which are still unclear [10]. Based, again, mainly on laboratory experiments with birds, two theoretical models of magnetoreception in terrestrial animals were singled out as the most viable: one involving magnetic nanocrystals of iron oxides [11] and the other based on magneto-sensitive photochemical reactions in the eye [12,13] (the radical-pair model, RPM).
While within the iron-oxide model magnetic fields are supposed to be detected through the torque they exert on the nanocrystal having a huge magnetic moment (largely like the magnetic needle in a technical compass), the physics behind the RPM is more subtle [14]. Here, the magnetic sensitivity is provided by extremely short-lived (within the microsecond range) directional correlation of magnetic moments of just two electrons. These two electrons reside in radicals, which can be formed by splitting of a specific molecule (or by an electron transfer within the molecule) upon absorption of a photon. If the magnetic field affects these two magnetic moments in different ways (this may happen if hyperfine [15] or spin–orbit [16] interactions in the two radicals are different), then their mutual orientation changes as a function of the field, and this change becomes more pronounced with time elapsed after separation of the radicals. Because, as known from quantum chemistry, covalent chemical bonds are formed by electrons with antiparallel magnetic moments (singlet spin state), the conversion of the pair of magnetic moments from antiparallel (singlet) to parallel (triplet) configuration and vice versa strongly affects the probability of recombination of the two radicals, and ultimately the yield of the entire photochemical reaction. The sensitivity of radical-pair reactions to magnetic fields is well known in chemistry [17]; recently, such a sensitivity was demonstrated in vitro for cryptochrome, the photosensitive protein that is considered as the candidate magnetoreceptor molecule of the bird compass [18]. However, the field intensities, required to considerably affect the photochemical process in the cryptochrome, were two orders of magnitude higher than that of the geomagnetic field. Much higher sensitivity was reached with an artificial molecule comprising porphyrine, carotenoid and fullerene, the use of which realized a model ‘chemical compass’ [19].
Although the results of many behavioural experiments suggested that the bird's compass actually worked on radical-pair reactions (for example, its sensitivity to the spectrum of the ambient light [20]), the crucial evidence was lacking. The situation has apparently changed with the demonstration of the effect of weak oscillating magnetic fields (OMFs) on the orientation ability of European robins (Erithacus rubecula) by the group of R. and W. Wiltschko in Frankfurt [21–24]. These experiments were specially designed to test the radical-pair theory of magnetoreception, which predicted sensitivity not only to static, but also to RF magnetic fields in the few-MHz frequency range (close to the electron spin resonance in typical geomagnetic fields). Their overwhelming success was, and often continues to be, considered as an indication that the orientational magnetoreception of birds is based on the radical-pair mechanism. However, it soon turned out that the experiments had been, in a sense, too successful; in fact, birds appeared to be RF field-sensitive to the extent that the radical-pair theory was unable to explain [25]; see also discussion in [26].
To understand the origin of this problem, one needs to recall the abovementioned time-dependence of the magnetic-field effect on the mutual orientation of electron magnetic moments. In the extreme and most favourable for the magnetic sensitivity case when the direction of one of the electron moments is fixed by strong and anisotropic hyperfine interaction, whereas the other one is free (the ‘reference–probe’ model [23]), the probabilities of finding the pair of electrons in the spin-singlet state oscillates at the Larmor frequency _f_L = γ_e_B, where B is the magnetic field intensity and γ_e ≈ 28 Hz nT−1 is the electron gyromagnetic ratio. The amplitude of the probability oscillation depends on the direction of the magnetic field as A(1 − cos2_θ), where θ is the angle between the magnetic field and the anisotropy axis of the ‘reference’ radical, and A < 1/2 is a constant depending on the details of the radical-pair formation process. A considerable difference in the singlet yield for radical pairs with different directions of the anisotropy axis with respect to the magnetic field can develop if both the lifetime of the radical pair and disorientation time of electron moments (spin relaxation time) are comparable with or longer than (2_πf_L)−1. For B = 50 µT, this time amounts to 0.1 µs. With the typical radical-pair lifetimes of the order of a few microseconds, this would give a very good directional sensitivity (provided controllable orientation of the molecules in the magnetoreceptor; see discussion of this issue in [26]). Comparison with the results of in vitro experiments performed on plant cryptochrome [18] shows, however, that RP systems in cryptochromes may be very far from optimal, most likely because spin relaxation times are short and hyperfine couplings in both radicals are of comparable strength. Because similar experiments with bird cryptochromes have not been done, it remains an open question whether and to what extent their sensitivity to magnetic fields could be improved by evolution (see [27] for a discussion of possible ways for such improvements).
Now, if we turn to the effect of RF fields, the situation looks much less optimistic even in the ideal ‘reference–probe’ model. If a weak RF field with the amplitude _B_RF and the frequency _f_B is applied at the angle φ to the static field B, then, even at θ = 0, oscillations between the singlet and triplet configurations will occur at the Rabi frequency _f_R = 1/2_γ_e_B_RF sin φ. If all the characteristic times in the system were long enough, this would equalize the singlet yield for all orientations of the molecule and, therefore, disrupt the work of the radical-pair compass. However, because now both the lifetime and spin relaxation time must be longer than (2_πf_R)−1, and 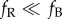 (because
(because 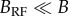 ), the criterion becomes much more severe: for the lowest RF amplitude that was reported to disrupt orientation of European robins, i.e. 15 nT applied at the angle of 24° to the static field [22], this would mean 2 ms that exceeds the most daring estimates of electron spin relaxation time in organic radicals. Further, one must take into account that the static magnetic field varied from 46 to 47.4 µT for different experimental cages in [22], but the RF field with the amplitude _B_RF ≥ 15 nT and frequency 1.315 MHz always caused disorientation, while for most cages it was, in fact, off the resonance.1 The theory of magnetic resonance says that the expected effect of the RF field, in this case, is at least 100 times weaker that one of the strictly resonant RF field; therefore, the required spin relaxation time shifts to utterly unbelievable 0.2 s. A similar estimate (about 1 s) was obtained in reference [25] from the observed effect of a broadband (0.1–10 MHz) RF field with the amplitude of 85 nT [24]. No need to say that any remotely realistic model of the radical pair would give worse sensitivity to the RF field than the ‘reference–probe’ model considered above. Several theoretical works published in recent years have attempted to reproduce some of the experimental results [28–30], invoking various quantum and statistical effects to suppress decoherence and enhance sensitivity. With rather liberal choice of relaxation times, the effect of a strictly resonant RF field with the amplitude of 150 nT could be reproduced [28]. No one has ever tried to explain the effect of 15 nT monochromatic or of 85 nT broadband fields. The scattering of static field intensities (and therefore of resonance frequencies) in the experiment of reference [22] has never been taken into account either.
), the criterion becomes much more severe: for the lowest RF amplitude that was reported to disrupt orientation of European robins, i.e. 15 nT applied at the angle of 24° to the static field [22], this would mean 2 ms that exceeds the most daring estimates of electron spin relaxation time in organic radicals. Further, one must take into account that the static magnetic field varied from 46 to 47.4 µT for different experimental cages in [22], but the RF field with the amplitude _B_RF ≥ 15 nT and frequency 1.315 MHz always caused disorientation, while for most cages it was, in fact, off the resonance.1 The theory of magnetic resonance says that the expected effect of the RF field, in this case, is at least 100 times weaker that one of the strictly resonant RF field; therefore, the required spin relaxation time shifts to utterly unbelievable 0.2 s. A similar estimate (about 1 s) was obtained in reference [25] from the observed effect of a broadband (0.1–10 MHz) RF field with the amplitude of 85 nT [24]. No need to say that any remotely realistic model of the radical pair would give worse sensitivity to the RF field than the ‘reference–probe’ model considered above. Several theoretical works published in recent years have attempted to reproduce some of the experimental results [28–30], invoking various quantum and statistical effects to suppress decoherence and enhance sensitivity. With rather liberal choice of relaxation times, the effect of a strictly resonant RF field with the amplitude of 150 nT could be reproduced [28]. No one has ever tried to explain the effect of 15 nT monochromatic or of 85 nT broadband fields. The scattering of static field intensities (and therefore of resonance frequencies) in the experiment of reference [22] has never been taken into account either.
An alternative approach to this puzzle is to say that the change of the reaction yield under the RF field might be very small, should the bird's sensory system be so sensitive to this signal that at the final stages of neural transduction it could interfere with the signal from the static field. This explanation is, however, no less problematic (see discussion in reference [26])—for instance, it would require an unrealistically large number of cryptochrome molecules to provide the required sensitivity. Ultimately, it is unclear why it could be advantageous in evolutionary terms to develop an outstanding sensitivity to tiny signals from RF fields which birds apparently never met in nature.
To summarize this long introduction, experimental works [21,22,24] and consequent theoretical studies aimed at their explanation [25,28] have led to a dilemma: either the experiments of the Frankfurt group on the RF field effect upon orientation of European robins were flawed and their results are incorrect, or the RPM in its present form does not correctly reproduce the physics of the bird's magnetic compass. As the RPM is one of the mainstream theories of magnetoreception, and there exists a considerable body of experimental evidence supporting the idea that compass magnetoreception is based on the vision system of birds, independent experimental studies aimed at confirmation or negation of the results of previous studies [21,22,24] are of key importance for the further development of the science of magnetoreception [10].
Here, we report the results of our experiments which were aimed to independently replicate the claims that small-amplitude RF field disrupts magnetic orientation in migratory songbirds.
2. Material and methods
We performed orientation tests in autumn 2013 with garden warblers (S. borin) captured during their first autumn migration on the Courish Spit. Experimental birds were kept in an outdoor aviary, they experienced natural photoperiod, natural geomagnetic field and had access to astronomic orientation cues (sun, sunset polarization patterns and stars) during their time in captivity. They were provided food (mealworms) and water ad libitum.
In the first series of tests, migratory orientation of garden warblers was studied in the natural magnetic field (NMF) in Emlen funnels [31] under simulated total overcast. On top of Emlen funnels, we put lids made of milky glass which completely obscured star and any other patterns but let enough light in. Thus, the only orientation cue available to experimental birds during the test was geomagnetic field. Each test lasted for 40 min and started at the beginning of astronomical twilight. We used modified Emlen funnels made of aluminium (top diameter 300 mm, bottom diameter 100 mm, slope 45° with the top opening covered by netting). The directionality of the birds' activity was recorded as scratches left by their claws as they hopped in the funnels on a print film covered with a dried mixture of whiting and glue. Two researchers (N.C. and A.P.) independently determined each bird's mean direction from the distribution of the scratches. In most cases, the mean direction could be very precisely identified using the simple visual estimation method [32]. If a pattern of scratches was not clear, both persons independently counted scratches in each of 36 × 10° sectors (some birds leave so many scratches that an exact number of scratches cannot be counted, but must be estimated) and used circular statistics software to assess the directionality based on the numbers of scratches. The mean of the two observer's determined directions was recorded as the orientation result. If both observers considered the scratches to be randomly distributed or if the two mean directions deviated by more than 30°, then the bird was considered to be not oriented in the given test. The same counting procedure was used also in all the tests with applied magnetic fields (see below). In view of the limited number of birds we could test in parallel during our outdoor testing sessions, we opted for not using the fully double-blind procedure, which would not have allowed us to complete the experiments within one field season; this choice was justified, in our view, by absence of obvious preferences in our group as to the expected outcome of the tests: both confirmation and non-confirmation of the results of Frankfurt experiments was considered equally interesting.
After testing the birds for at least three and up to five times and receiving at least two and up to five results when the birds were sufficiently active (i.e. left at least 40 and nearly always more than 100 scratches on underlying paper) and showed orientation sufficiently concentrated (i.e. it would have passed the Rayleigh test [33] at the 5% significance level had the movements in the funnel been independent events), the birds were transferred to the second phase of the experiment. Inactive (fewer than 40 scratches) tests and not sufficiently concentrated trials were excluded from analysis. From the active and oriented tests, the mean directions of each individual were calculated using vector addition.
In the second series of tests, the same birds were tested in RF OMF. The field was created by coils 0.75 m in diameter, fed from a commercial stabilized high-frequency generator and a custom-made broadband power amplifier, which served to match the generator output with the 50 Ohm coaxial cable 40 m in length. Each coil comprised three wraps of isolated copper wire 1 mm in cross section, fixed to a wooden frame. The cable transmitted the RF current to coils and allowed us to place them at a distance from the RF source assembly (generator, amplifier, power supply). The source assembly was put into a wooden case, placed on a clearing in reeds approximately in 25 m along the straight line from the centre of another clearing, where two wooden tables for experimental cages were build. The sound of working devices was not discernible already 2 m apart from the case, and near experimental tables it was by far below natural environmental noises. On the tables, up to four coils could be installed, two on each. Coils were connected in parallel by small lengths of coaxial cable to the main RF cable and mounted, so that one side of the coil was raised above the table top to increase the angle between OMF and NMF, which as a result was approximately 30°. Emlen funnels were placed on tables, one in the centre of each coil. In all the experiments, monochromatic RF fields with the amplitude 190 nT and frequency 1.403 MHz (matching the Larmor frequency of a free-standing electrons spin in the local GMF of 50 100 nT) were used. The parameters of the OMF were controlled before and after each test. To this end, we used a single wire loop 20 cm in diameter, connected to a digital oscilloscope. The loop was placed consequently in the centre of each coil. The periodic in time induction EMF was visualized, its frequency and amplitude measured, and the latter recalculated into the amplitude of RF magnetic field. Using this device and applying real-time fast Fourier transform to the measured signal, we also analysed the spectrum of background noise in the frequency range 0.1–100 MHz. Its rms amplitude was found to be always below 0.5 nT.
The tests were again performed under milky glass, so that experimental birds had no access to astronomic orientation cues.
After the birds were tested in OMF at least three times (and up to five times) and produced at least two and up to three usable directions in individual tests (see above for criteria for usability), they were tested again in stationary magnetic field, which was however rotated 120° anticlockwise in the horizontal plane (rotated magnetic field, RMF). The total intensity and vertical component of this rotated field were identical to the natural values. To produce the artificial rotated field, electric current flowed through the magnetic coils (Helmholtz coil system with two coils per axis) custom built by Pushkov Institute of Terrestrial Magnetism, Ionosphere and Radiowave Propagation (Russian Academy of Sciences, St Petersburg, Russia). The coils were managed by custom-written software. These coils were the same as the ones used in reference [34]. This last experiment was performed to test (i) whether RF fields had any lasting effect on the birds’ orientation performance and (ii) whether they indeed used the magnetic information and not anything else for their orientation (even though the use of celestial cues was ruled out by putting milky glass lids on top of the funnels).
We tested a total of 19 garden warblers in NMF and in OMF. One bird never showed directed and sufficiently concentrated orientation in OMF, in spite of being tested five times. Unfortunately, we managed to test only eight garden warblers in RMF before the end of September when migratory activity and orientation sharply decreased. The reason for this could be that our experimental birds remained in the Baltic area, whereas in the wild, they should have been in the Iberian peninsula or already crossing the western Sahara [35]. They may have lacked some crucial cues and sharply decreased their activity because of that (cf. [34,36,37]). Thus, here we report the data on eight individuals that were successfully tested in all three conditions, NMF, OMF and RFM. To avoid pseudo-replication, the mean values of successful tests for each bird (two to five tests) were used in analyses, so that sample size was equal to the number of experimental birds (eight), not to the number of successful tests (24 in NMF, 18 in OMF, 22 in RMF; table 1).
Table 1.
Orientation behaviour of garden warblers during autumn migration on the Courish Spit in three testing conditions: natural magnetic field, radiofrequency oscillating magnetic field (OMF), and rotated magnetic field (RMF). RMF was rotated 120° anticlockwise in the horizontal plane as compared to the NMF, i.e. to make the results in RMF comparable with the results of the same birds in NMF, 120° should be added to each value. All tests were performed without access to celestial cues.
| ring no | results of successful tests in NMF | mean direction in NMF | results of successful tests in OMF | mean direction in OMF | results of successful tests in RMF | mean direction in RMF |
|---|---|---|---|---|---|---|
| 552 | 229°, 249°, 313° | 264° | 40°, 130° | 85° | 69°, 234° | 154° |
| 528 | 260°, 240°, 310° | 270° | 270°, 0° | 315° | 0°, 110°, 99° | 75° |
| 716 | 300°, 205°, 216° | 237° | 85°, 100°, 320° | 48° | 100°, 172°, 210° | 163° |
| 625 | 151°, 140°, 210°, 258°, 001° | 191° | 170°, 330° | 250° | 159°, 328°, 60° | 59° |
| 728 | 265°, 310° | 288° | 280°, 120° | 200° | 85°, 160°, 324° | 93° |
| 809 | 220°, 310° | 265° | 330°, 280°, 200° | 273° | 155°, 110° | 132° |
| 881 | 0°, 222°, 205° | 262° | 290°, 340° | 315° | 80°, 240°, 35° | 55° |
| 689 | 40°, 270°, 210° | 263° | 98°, 169° | 134° | 310°, 91°, 60° | 34° |
| second-order mean | 256°, r = 0.89, p < 0.001 | 281°, r = 0.16, n.s. | 94°, r = 0.71, p = 0.012 |
Second-order mean circular distributions were tested for directionality by the Rayleigh test [33]. Differences in mean direction between experimental groups were analysed using the parametric Watson–Williams _F_-test in the cases where the assumptions underlying this test were fulfilled. When the concentrations of the samples to be compared were too low to use the Watson–Williams _F_-test, the non-parametric Mardia–Watson–Wheeler test was used [33].
3. Results
Garden warblers tested in NMF without access to celestial cues showed the mean orientation towards the WSW (table 1; α = 256°, 95% confidence interval (CI) = 233–280°, n = 8, r = 0.89, p < 0.001, figure 1a). This value is identical to the mean direction shown by garden warblers in nocturnal tests at the same site with full access to celestial and magnetic cues: α = 256°, 95% CI = 224–288° [38] and somewhat more westerly than the direction of autumn migration from the Courish Spit as suggested by ring recoveries: 212°, 95% CI = 204–221° (calculated from data in [39]).
Figure 1.
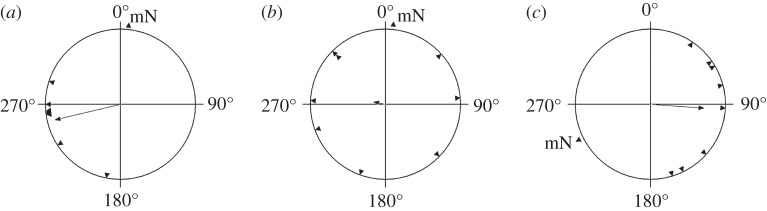
Orientation of garden warblers on the Courish Spit during autumn migration without access to celestial cues that showed directed results under all three testing conditions (n = 8). (a) Results in the natural magnetic field; (b) results in oscillating magnetic field; (c) results in the magnetic field rotated 120° anticlockwise, shown in respect to the geographical north. Each triangle at the circular diagram periphery indicates the mean orientation of one individual bird. The triangle and letters mN show the position of the magnetic north in each testing condition. Arrowheads show group mean directions and vector lengths.
The same birds tested in OMF showed no consistent orientation: α = 281°, n = 8, r = 0.16, p = 0.83 (figure 1b). The full results with 19 garden warbler in NMF and 18 birds in OMF, including birds that subsequently failed to show sufficient activity and directionality when tested in RMF, most probably owing to calendar effects, and were thus not included in the main analysis, showed the same pattern (figure 2): α = 247°, 95% CI = 231–264°, n = 19, r = 0.81, p < 0.001 in NMF and α = 236°, n = 18, r = 0.22, p = 0.41 in OMF. These two distributions were significantly different (Mardia–Watson–Wheeler test, W = 3.78, p = 0.025), apparently owing to much lower concentration of directions when RF field was applied.
Figure 2.
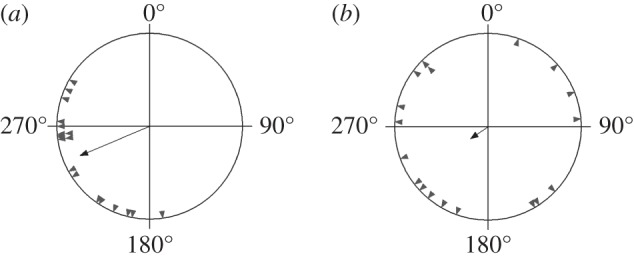
Orientation of all garden warblers that showed directed results (a) in the natural magnetic field (n = 19) and (b) in oscillating magnetic field (n = 18).
When the same birds were again tested in a stationary magnetic field rotated 120° anticlockwise, they were again significantly oriented, this time towards the east (mean direction 94°, table 1). With respect to the magnetic north, their orientation was α = 214°, 95% CI = 174–254°, n = 8, r = 0.71, p = 0.012. The difference of this direction (when taken in respect to the magnetic North) from the direction in NMF was not significant (Watson–Williams test, _F_1,14 = 4.30, p = 0.057). The southwesterly magnetic direction shown by garden warblers in RMF is consistent with autumn migratory direction of free-living birds of this species in the Eastern Baltic (212°, from [39]).
4. Discussion
Behaviour of birds in our experiments strongly suggests that in the stationary field, both the natural one and the one rotated 120° in the horizontal plane, garden warblers used the magnetic field for their orientation, because they were denied access to celestial orientation cues and showed a predictable response to the rotation of magnetic North. Our results thus indicate that (i) garden warblers can select and maintain broadly correct migratory orientation on the basis of magnetic cues alone; (ii) this ability is damaged by weak (190 nT) OMFs; (iii) this impairment is transient, i.e. when the same birds are again tested in a stationary magnetic field, they orient successfully. Thus, our results independently replicate reports of the group of R. and W. Wiltschko on the disruptive effect of RF magnetic fields on the magnetic compass of migratory songbirds [21,22,24]. It is important to stress that though our experimental protocol was essentially the same as one used in the Wiltschkos' group, there were several notable differences: we worked with a different bird species (garden warblers instead of European robins), in a different season (autumn instead of spring) and used a different type of antenna and electronic equipment. Nevertheless, the basic effect reported in [21,22,24] was reproduced.
As discussed above, the high sensitivity of the birds' magnetic compass to RF fields, found in [21,22,24] and now confirmed by us, is difficult to explain within the existing radical-pair theory. The reason is that the time needed for the RF field to change the state of an electron spin cannot be shorter than the period of rotation of the same spin in a static field of the same amplitude (assuming all the other interactions nullified). This period is determined by the field amplitude and the electron gyromagnetic ratio, which for organic radicals cannot strongly deviate from its value for free electrons (given by a combination of fundamental constants). For this simple reason, the time needed to change the spin state by applying the RF field with the amplitudes known to disrupt the birds' compass appears to be on the scale of milliseconds, exceeding by two orders of magnitude maximum electron spin lifetimes known for organic radicals. The presence of various spin–spin and spin–orbit interactions can only diminish the effect of the RF field, making the observed sensitivities even more difficult to explain.
It may be argued that we tested our birds only for a resonance effect and made no tests in an off-resonance or broadband RF oscillating field; therefore, we cannot distinguish between a specific Zeeman resonance effect of a relatively weak (190 nT) RF field and a putative effect of an off-resonance or broadband RF field of the same intensity. However, as emphasized in the Introduction, neither the resonance nor off-resonance RF field effect have so far found a satisfactory theoretical explanation. Our main point is that a RF field of roughly the same amplitude as used in the experiments of [22,24] is indeed able to disrupt the magnetic orientation of a migratory songbird, and thus it is very unlikely that the findings of aforementioned studies resulted from some systematic error or a hidden flaw of the experimental procedure. This, in its turn, means that the radical-pair theory of avian magnetoreception in its current form may need revision to account for this RF field effect.
More experimental work is needed to refine the spectral width of the observed Zeeman resonance in the sensitivity of the avian compass to RF fields [22] and to verify the reported sensitivity threshold at the exact resonance (about 15 nT). Only after obtaining these data (which may take several field seasons), one would be able to obtain a reliable experimental estimate of the spin lifetimes in the radical pair or whatever physical system is responsible for the compass magnetoreception of birds. However, already these previous results demand the renewed attention of theoreticians to the physical and chemical models of magnetoreception.
Acknowledgements
The authors are most grateful to two anonymous reviewers for their constructive criticism that helped us to improve an earlier draft. Experiments with animals followed the current legislation of Russia and institutional guidelines and were approved by the ethical committee of the Zoological Institute of Russian Academy of Sciences.
Endnote
1
An inhomogeneous broadening (e.g. owing to weak hyperfine coupling with several distant nuclei) could apparently diminish this effect by bringing RPs back to resonance; however, the inhomogeneous broadening, in fact, shifts the resonant frequency of each given RP by some random value, that is, it does the same thing as the deviation of static field in the Frankfurt experiments. The transition would have been brought back to resonance only if the two frequency shifts, internal and external, were correlated in such a manner that they exactly compensated each other, which is obviously not possible to achieve for the entire ensemble of RPs with random hyperfine couplings.
Funding statement
This study was supported by a grant from the Interdisciplinary Grant Programme of St Petersburg Research Centre of Russian Academy of Sciences and by Russian Foundation for Basic Research (grant no. 12-04-00296-a). Further financial assistance was provided by St Petersburg State University through research grants nos. 1.37.149.2014 and 1.37.159.2014.
References
- 1.Kennedy D, Norman C. 2005. What don't we know? Science 309, 78b–102b. ( 10.1126/science.309.5731.78b) [DOI] [PubMed] [Google Scholar]
- 2.Gill RE, Jr, et al. 2009. Extreme endurance flights by landbirds crossing the Pacific Ocean: ecological corridor rather than barrier? Proc. R. Soc. B 276, 447–457. ( 10.1098/rspb.2008.1142) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kramer G. 1951. Eine neue Methode zur Erforschung der Zugorientierung und die bisher damit erzielten Ergebnisse. In Proc. X Int. Ornithol. Congr., pp. 269–280. Uppsala, Sweden: Almqvist and Wiksell. [Google Scholar]
- 4.Kramer G. 1953. Die Sonnenorientierung der Vögel. _Verh. Dtsch. zool. Ges._Zool. Anzeig. Suppl. 16, 72–84. [Google Scholar]
- 5.Sauer F. 1957. Die Sternenorientierung nächtlich ziehender Grasmücken (Sylvia atricapilla, borin und curruca). Z. Tierpsychol. 14, 29–70. [Google Scholar]
- 6.Emlen ST. 1967. Migratory orientation in the indigo bunting, Passerina cyanea. Part I: evidence for use of celestial cues. Auk 84, 309–342. ( 10.2307/4083084) [DOI] [Google Scholar]
- 7.Emlen ST. 1967. Migratory orientation in the indigo bunting, Passerina cyanea. Part II: mechanism of celestial orientation. Auk 84, 463–489. ( 10.2307/4083330) [DOI] [Google Scholar]
- 8.Emlen ST. 1970. Celestial rotation: its importance in the development of migratory orientation. Science 170, 1198–1201. ( 10.1126/science.170.3963.1198) [DOI] [PubMed] [Google Scholar]
- 9.Wiltschko W, Wiltschko R. 1972. Magnetic compass of European robins. Science 176, 62–64. ( 10.1126/science.176.4030.62) [DOI] [PubMed] [Google Scholar]
- 10.Mouritsen H. 2013. The magnetic senses. In Neurosciences: from molecule to behavior: a university textbook (eds Galizia CG, Lledo P-M.), pp. 427–443. Berlin, Germany: Springer. [Google Scholar]
- 11.Kirschvink JL, Gould JL. 1981. Biogenic magnetite as a basis for magnetic field sensitivity in animals. Biosystems 13, 181–201. ( 10.1016/0303-2647(81)90060-5) [DOI] [PubMed] [Google Scholar]
- 12.Schulten K, Swenberg CE, Weller A. 1978. A biomagnetic sensory mechanism based on magnetic field modulated coherent electron spin motion. Z. Phys. Chem. 111, 1–5. ( 10.1524/zpch.1978.111.1.001) [DOI] [Google Scholar]
- 13.Ritz T, Adem S, Schulten K. 2000. A model for photoreceptor-based magnetoreception in birds. Biophys. J. 78, 707–718. ( 10.1016/S0006-3495(00)76629-X) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rodgers CT, Hore PJ. 2009. Chemical magnetoreception in birds: the radical pair mechanism. Proc. Natl Acad. Sci. USA 106, 353–360. ( 10.1073/pnas.0711968106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Turro N. 1983. Influence of nuclear spin on chemical reactions: magnetic isotope and magnetic field effects (a review). Proc. Natl Acad. Sci. USA 80, 609–621. ( 10.1073/pnas.80.2.609) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lambert N, De Liberato S, Emary C, Nori F. 2013. Radical pair model of magnetoreception with spin–orbit coupling. New J. Phys. 15, 083024 ( 10.1088/1367-2630/15/8/083024) [DOI] [Google Scholar]
- 17.Salikhov KM, Molin Yu.N, Sagdeev RZ, Buchachenko AL. 1984. Spin polarization and magnetic effects in radical reactions. Amsterdam, The Netherlands: Elsevier. [Google Scholar]
- 18.Maeda K, et al. 2012. Magnetically sensitive light-induced reactions in cryptochrome are consistent with its proposed role as a magnetoreceptor. Proc. Natl Acad. Sci. USA 109, 4774–4779. ( 10.1073/pnas.1118959109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Maeda K, Henbest KB, Cintolesi F, Kuprov I, Rodgers CT, Liddell PA, Gust D, Timmel CR, Hore PJ. 2008. Chemical compass model of avian magnetoreception. Nature 453, 387–390. ( 10.1038/nature06834) [DOI] [PubMed] [Google Scholar]
- 20.Wiltschko W, Gesson M, Wiltschko R. 2001. Magnetic compass orientation of European robins under 565 nm green light. Naturwissenschaften 88, 387–390. ( 10.1007/s001140100248) [DOI] [PubMed] [Google Scholar]
- 21.Ritz T, Thalau P, Phillips JB, Wiltschko R, Wiltschko W. 2004. Resonance effects indicate a radical-pair mechanism for avian magnetic compass. Nature 429, 177–180. ( 10.1038/nature02534) [DOI] [PubMed] [Google Scholar]
- 22.Ritz T, Wiltschko R, Hore PJ, Rodgers CT, Stapput K, Thalau P, Timmel CR, Wiltschko W. 2009. Magnetic compass of birds is based on a molecule with optimal directional sensitivity. Biophys. J. 96, 3451–3457. ( 10.1016/j.bpj.2008.11.072) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ritz T, Ahmad M, Mouritsen H, Wiltschko R, Wiltschko W. 2010. Photoreceptor-based magnetoreception: optimal design of receptor molecules, cells, and neuronal processing. J. R. Soc. Interface 7, S135–S146. ( 10.1098/rsif.2009.0456.focus) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Thalau P, Ritz T, Stapput K, Wiltschko R, Wiltschko W. 2005. Magnetic compass orientation of migratory birds in the presence of a 1.315 MHz oscillating field. Naturwissenschaften 92, 86–90. ( 10.1007/s00114-004-0595-8) [DOI] [PubMed] [Google Scholar]
- 25.Kavokin KV. 2009. The puzzle of magnetic resonance effect on the magnetic compass of migratory birds. Bioelectromagnetics 30, 402–410. ( 10.1002/bem.20485) [DOI] [PubMed] [Google Scholar]
- 26.Mouritsen H, Hore PJ. 2012. The magnetic retina: light-dependent and trigeminal magnetoreception in migratory birds. Curr. Opin. Neurobiol. 22, 343–352. ( 10.1016/j.conb.2012.01.005) [DOI] [PubMed] [Google Scholar]
- 27.Lee AA, Lau JCS, Hogben HJ, Biscup T, Kattnig DR, Hore PJ. 2014. Alternative radical pairs for cryptochrome-based magnetoreception. J. R. Soc. Interface 11, 20131063 ( 10.1098/rsif.2013.1063) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gauger EM, Rieper E, Morton JJL, Benjamin SC, Vedral V. 2011. Sustained quantum coherence and entanglement in the avian compass. Phys. Rev. Lett. 106, 040503 ( 10.1103/PhysRevLett.106.040503) [DOI] [PubMed] [Google Scholar]
- 29.Cai J, Caruso F, Plenio M. 2012. Quantum limit for avian magnetoreception: how sensitive can a chemical compass be? Phys. Rev. A 85, 040304(R) ( 10.1103/PhysRevA.85.040304) [DOI] [Google Scholar]
- 30.Dellis AT, Kominis IK. 2012. The quantum Zeno effect immunizes the avian compass against the deleterious effects of exchange and dipolar interactions. Biosystems 107, 153–157. ( 10.1016/j.biosystems.2011.11.007) [DOI] [PubMed] [Google Scholar]
- 31.Emlen ST, Emlen JT. 1966. A technique for recording orientation of captive birds. Auk 83, 361–367. ( 10.2307/4083048) [DOI] [Google Scholar]
- 32.Mouritsen H, Larsen ON. 1998. Evaluating alternative methods for the analysis of Emlen funnel data. In Compasses and orientational strategies of night migrating passerine birds. (ed. Mouritsen H.), pp. 115–124. Odense: Center of Sound Communication, Institute of Biology, Odense University. [Google Scholar]
- 33.Batschelet E. 1981. Circular statistics in biology. London, UK: Academic Press. [Google Scholar]
- 34.Beck W, Wiltschko W. 1988. Magnetic factors control the migratory direction of pied flycatchers (Ficedula hypoleuca Pallas). In Acta XIX Congr. Int. Ornithol., vol. 2 (ed. Ouellet H.), pp. 1955–1962. Ottawa, Canada: University of Ottawa Press. [Google Scholar]
- 35.Gwinner E, Wiltschko W. 1978. Endogenously controlled changes in migratory direction of the garden warbler, Sylvia borin. J. Comp. Physiol. A 125, 267–273. ( 10.1007/BF00656605) [DOI] [Google Scholar]
- 36.Fransson T, Jakobsson S, Johansson P, Kullberg C, Lind J, Vallin A. 2001. Magnetic cues trigger extensive refuelling. Nature 414, 35–36. ( 10.1038/35102115) [DOI] [PubMed] [Google Scholar]
- 37.Kishkinev D, Chernetsov N, Bolshakov CV. 2006. Migratory orientation of first-year pied flycatchers (Ficedula hypoleuca) from the Eastern Baltic. Ornithologia (Moscow) 33, 153–160 (in Russian). [Google Scholar]
- 38.Pakhomov A, Chernetsov N. 2014. Early evening activity of migratory garden warbler Sylvia borin: compass calibration activity? J. Ornithol. ( 10.1007/s10336-014-1044-x) [DOI] [Google Scholar]
- 39.Bolshakov CV, Shapoval AP, Zelenova NP. 2001. Results of bird trapping and ringing by Biological Station ‘Rybachy’ on the Courish Spit: long-distance recoveries of birds ringed in 1956–1997. Part 1. Non-passerines. Passerines (Alaudidae, Hirundinidae, Motacillidae, Bombicillidae, Troglodytidae, Prunellidae, Turdidae, Sylviidae, Regulidae, Muscicapidae, Aegithalidae). Avian Ecol. Behav. Suppl. 1, 1–126. [Google Scholar]