Mouse Taste Buds Use Serotonin as a Neurotransmitter (original) (raw)
Abstract
Synapses between gustatory receptor cells and primary sensory afferent fibers transmit the output signal from taste buds to the CNS. Several transmitter candidates have been proposed for these synapses, including serotonin (5-HT), glutamate, acetylcholine, ATP, peptides, and others, but, to date, none has been unambiguously identified. We used Chinese hamster ovary cells stably expressing 5-HT2C receptors as biodetectors to monitor 5-HT release from taste buds. When taste buds were depolarized with KCl or stimulated with bitter, sweet, or sour (acid) tastants, serotonin was released. KCl- and acid-induced 5-HT release, but not release attributable to sweet or bitter stimulation, required Ca2+ influx. In contrast, 5-HT release evoked by sweet and bitter stimulation seemed to be triggered by intracellular Ca2+ release. These experiments strongly implicate serotonin as a taste bud neurotransmitter and reveal unexpected transmitter release mechanisms.
Keywords: biosensors, taste, serotonin, transmitters, calcium imaging, phospholipase C β2
Introduction
It has long been known that serotonin stimulates gustatory sensory afferent fibers when that biogenic amine is injected into the tongue near taste buds (Esakov et al., 1983), suggesting that 5-HT may be a taste bud neurotransmitter (for review, see Nagai et al., 1996). A number of findings have subsequently supported that notion. For example, serotonin is present in a subset of taste cells in a wide variety of species (Whitear, 1989; Delay et al., 1993; Kim and Roper, 1995). In mammals, many of the taste cells that synapse with nerve fibers are serotonergic: they take up the serotonin precursor 5-hydroxytryptophan (5-HTP) and are immunopositive for 5-HT (Takeda, 1977; Kim and Roper, 1995; Yee et al., 2001). Reverse transcription-PCR data indicate that 5-HT1A and 5-HT3 receptors are expressed in taste tissue, with 5-HT1A receptors found in taste cells and 5-HT3 receptors found in sensory primary afferent fibers (Kaya et al., 2004). Finally, there is indirect evidence from autoradiographic studies showing that, when depolarized, amphibian taste cells release serotonin (Nagai et al., 1998). However, despite these findings, one of the most important of the canonical criteria for identifying synaptic neurotransmitters, namely detecting its release from stimulated synapses, has yet to be unambiguously established in taste buds for serotonin or for any other candidate. We used novel biosensor cells to detect the secretion of transmitter candidates from taste buds and report that taste stimulation elicits serotonin release.
Parts of this work have been published previously in abstract form (Huang et al., 2005).
Materials and Methods
Biosensor cells. Chinese hamster ovary (CHO) cells expressing 5-HT2C receptors (Berg et al., 1994) were seeded into 35-mm-diameter culture dishes. To produce biosensors, cells were suspended in HBSS containing 0.125% trypsin and collected in a 15 ml centrifuge tube after terminating the reaction with 2% FBS. Dispersed CHO cells were loaded with 4 μm fura-2 AM for 1 h at room temperature. An aliquot of fura-2-loaded cells was transferred to a recording chamber and viewed with an Olympus Optical (Tokyo, Japan) IX70 inverted microscope to test their responses to bath-applied 5-HT, KCl, cycloheximide, acetic acid, and saccharin. Sequential fluorescence microscopic images were recorded with a longpass emission filter (≥510 nm) with the cells excited at 340 nm, followed by 380 nm, and the ratios were calculated with Imaging Workbench version 5 software (INDEC Biosystems, Mountain View, CA) (data shown are these ratios, labeled _F_340/_F_380 in Figures 1, 2, 3).
Figure 1.
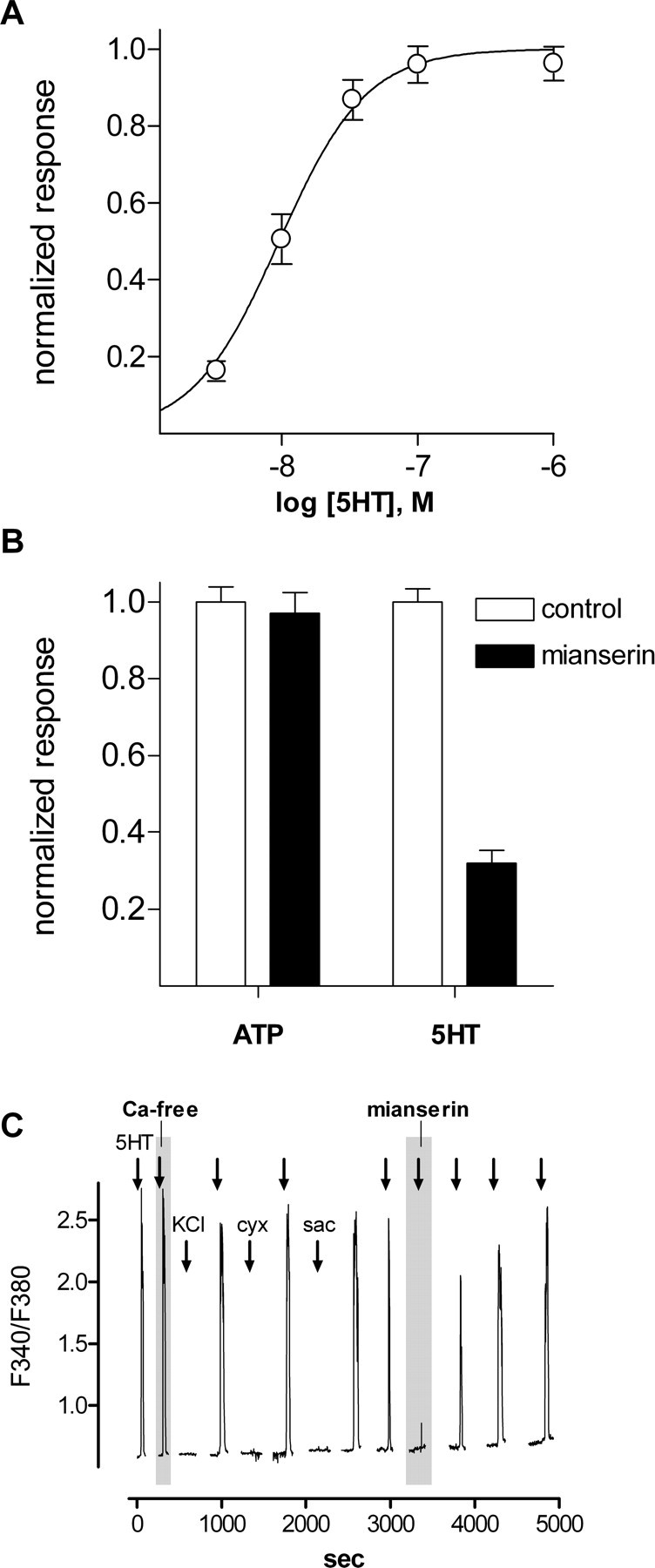
Ca2+ mobilization in CHO/5-HT2C cells elicited by serotonin and ATP. Cells were loaded with fura-2, and responses were measured as the ratio of emission to fluorescent excitation at 340-380 nm. A, Concentration-response relationships for 5-HT. Open circles indicate the mean±SEM (31 cells). B, CHO/5-HT2C cell responses evoked by 3 μm ATP and 3 nm 5-HT. Open columns, Control responses. Filled columns, Effects of mianserin (10 nm), a5-HT2C antagonist. Mianserin does not affect Ca2+ mobilization in response to ATP but reduces responses to 5-HT. Data are from a different experiment from that shown in A. ATP and 5-HT responses were measured in parallel, and responses are normalized to each agonist separately. C, Ca2+ mobilization in CHO/5-HT2C cells is elicited by repeated applications of 5-HT but not by tastants or depolarization with KCl. Sequential responses elicited by 10 nm 5-HT in the presence of 2 mm Ca2+ and in nominally 0 mm Ca2+ (shaded area labeled “Ca-free”), 50 mm KCl, 100 μm cycloheximide, and 20 mm saccharin and recovery of 5-HT responses after brief application of 10 nm mianserin (shaded area labeled “mianserin”). In this and subsequent figures, fura-2 excitation was extinguished between stimulus episodes to reduce phototoxicity and photobleaching, resulting in discontinuities in the traces. cyx, Cycloheximide; sac, saccharin.
Figure 2.
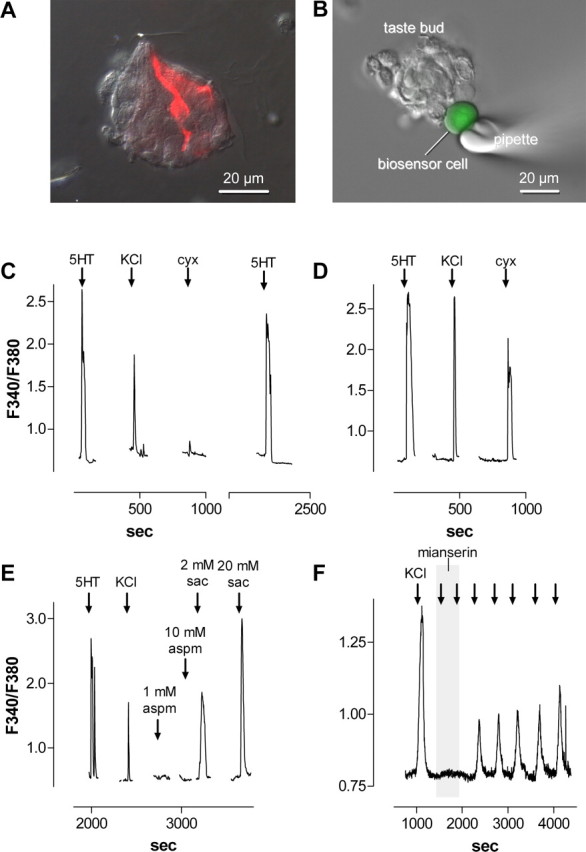
CHO/5-HT2C cells are biosensors for detecting serotonin release from isolated taste buds. A, Micrograph of a fixed, isolated taste bud immunostained for serotonin. Two immunopositive taste cells (red) are visible in this plane of focus. ANomarski differential interference contrast image was merged with an immunofluorescent micrograph for this micrograph. B, Micrograph of a fura-2-loaded (green) biosensor cell abutted against an isolated taste bud in a living preparation. A Nomarski differential interference contrast image and a fluorescence microscopy image were merged. C, Ca2+ mobilization in a biosensor cell positioned against a taste bud, as in B. The biosensor cell was initially stimulated with 3 nm 5-HT (↓) to verify its sensitivity, followed by 50 mm KCl to depolarize taste cells and 100 μm cycloheximide (cyx). D, A procedure similar to that in C was conducted on a taste bud preparation that had been pretreated with 500 μm 5-hydroxytryptophan to elevate the serotonin concentration in taste cells. Responses evoked by depolarization and taste stimulation are enhanced by this procedure. E, Saccharin (sac), but not aspartame (aspm), evoked serotonin release from taste buds. F, Mianserin (1 nm; shaded area) reversibly reduced the biosensor responses evoked by depolarizing taste buds repeatedly with 50 mm KCl (↓).
Figure 3.

Serotonin release from taste buds depends on Ca2+ influx for acid stimulation and KCl depolarization but not for taste stimulation with bitter (cycloheximide) or sweet (saccharin) compounds. A, Sequential biosensor responses from an isolated taste bud after stimulation with 50 mm KCl (↓) or 100 μm cycloheximide (cyx; ↓). The biosensor was tested for sensitivity at the beginning and end of the record with 3 nm 5-HT(↓). During the traces marked“Ca-free,” Ca2+ in the bath (2 mm) was exchanged for 3 mm Mg2+. B, An experiment similar to that shown in A, with stimulation by 20 mm sodium saccharin (sac; ↓). Note that responses elicited by KCl depolarization, but not by cycloheximide or saccharin, were eliminated by removing Ca2+ from the bathing medium. C, Serotonin release stimulated by 8 mm acetic acid, pH 5, was reduced by exchanging Mg2+ for Ca2+. D, Sequential responses from two different taste buds from a PLCβ2 null mouse. The interruption in the abscissa shows the two recordings. KCl depolarization evoked serotonin release, but neither 20 mm saccharin nor 100 μm cycloheximide stimulated transmitter release. HAc, Acetic acid.
Isolated taste buds. We removed the lingual epithelium containing taste papillae from the tongues of adult C57BL/6J female mice by injecting a mixture of 1 mg/ml collagenase A (Roche Diagnostics, Indianapolis, IN), 2.5 mg/ml dispase II (Roche Diagnostics), and 1 mg/ml trypsin inhibitor (Sigma, St. Louis, MO) directly under the epithelium surrounding taste papillae. The peeled epithelium was bathed in Ca-free solution for 30 min at room temperature, and isolated taste buds were drawn into fire-polished glass micropipettes with gentle suction. Taste buds were transferred to a shallow recording chamber that had a glass coverslip base. The coverslip was coated with Cell-Tak (BD Biosciences, San Jose, CA) to hold the taste buds firmly in place. Taste buds were perfused with Tyrode's solution (in mm): 140 NaCl, 5 KCl, 2 CaCl2, 1 MgCl2, 10 HEPES, 10 glucose, 10 Na-pyruvate, and 5 NaHCO3, pH 7.4 (310-320 Osm).
Stimulation. Taste buds were stimulated by bath perfusion of KCl (50 mm; substituted equimolar for NaCl), cycloheximide (10-100 μm), sodium saccharin (2-20 mm), aspartame (1-10 mm), and acetic acid (8 mm). All of the stimuli were made up in Tyrode's solution and applied at pH 7.2, except for acetic acid, which was applied at pH 5.
Immunohistochemistry. Isolated taste buds were fixed for 10 min in 4% paraformaldehyde in PBS. Taste buds were then rinsed three times in PBS and incubated for 2 h in PBS containing 0.3% Triton X-100, 2% normal donkey serum, and 2% bovine serum albumin. Taste buds were incubated with rabbit polyclonal anti-serotonin antibodies (1:1000; catalog #S5545; Sigma) for 60-90 min at room temperature. Thereafter, taste buds were washed three times in PBS, incubated for 1 h in Alexa Fluor 594-conjugated donkey anti-rabbit secondary antibodies (1:500; Molecular Probes, Eugene, OR), and then washed again three times in PBS.
Results
We produced biosensor cells for detecting serotonin by using CHO cells transfected with high-affinity 5-HT receptors. CHO-K1 cells stably expressing 5-HT2C receptors (Berg et al., 1994) were suspended in buffer and loaded with the Ca2+- sensitive dye fura-2. Fura-loaded CHO/5-HT2C cells responded robustly to bath-applied serotonin. The threshold for activation was ∼3 nm, and EC50 was 9 nm (Fig. 1_A_). This compares with the EC50 values of 15 and 29 nm reported previously for Ca2+ mobilization in fura-loaded CHO cells transiently transfected with 5-HT2C receptors (Akiyoshi et al., 1995, 1996). Biosensor cells respond to ATP (endogenous purinoreceptors), as well as to serotonin. Thus, we used mianserin, a 5-HT2 receptor antagonist, to differentiate responses to 5-HT from activation of endogenous (ATP) receptors (see below). Responses to 5-HT were reversibly blocked by 10 nm mianserin, but ATP responses were unaffected (Fig. 1_B_). Fura-loaded CHO/5-HT2C cells did not respond with Ca2+ signals to carbachol or glutamate up to 1 mm (data not shown). Furthermore, CHO/5-HT2C cells did not respond to depolarization with bath-applied KCl (50 mm; substituted for NaCl) nor to bath-applied taste stimuli-cycloheximide (100 μm), an aversive (bitter) taste compound for rodents, or saccharin (20 mm), a preferred (sweet) taste compound (Fig. 1_C_). The biosensor cells also did not respond to acetic acid stimulation (in humans, sour; 8 mm, pH 5; data not shown). Last, CHO/5-HT2C cells maintained their responses to bath-applied serotonin even if Ca2+ in the medium was replaced with Mg2+, consistent with the coupling of 5-HT2C receptors to intracellular Ca2+ release mechanisms (Fig. 1_C_). In short, CHO/5-HT2C cells were sensitive and reliable biosensors for serotonin, serotonergic responses could be differentiated from those produced by activating endogenous purinergic receptors by applying mianserin, and biosensor cells did not respond to taste stimuli or depolarization with KCl.
Subsequently, we isolated taste buds from vallate papillae of the mouse tongue. Serotonergic cells could be observed readily by immunostaining taste buds isolated for recording (Fig. 2_A_), verifying that their 5-HT content was maintained throughout the isolation procedure. Using fura-2-loaded taste buds, we verified that, as shown previously (Richter et al., 2004), ∼50% of taste bud cells are depolarized by bath-applied KCl (50 mm). Also, as shown previously (Bernhardt et al., 1996), cells in isolated taste buds responded to bath-applied bitter tastants (here, 10-100 μm cycloheximide) and saccharin (2-20 mm). Parenthetically, different taste cells responded to cycloheximide, saccharin, or KCl depolarization, but we did not investigate this systematically. Others have shown in more intact preparations of taste buds that many taste cells are responsive to two or more taste qualities (Gilbertson et al., 2001; Caicedo et al., 2002). Taste buds did not respond to low concentrations of bath-applied serotonin (3-10 nm).
We transferred an aliquot of suspended, fura-2-loaded CHO/5-HT2C cells into a chamber containing freshly isolated mouse taste buds adhering to the glass coverslip base of the chamber. A biosensor cell was drawn onto a fire-polished glass micropipette with gentle suction and maneuvered to an isolated taste bud (Fig. 2_B_). Mere physical contact between the biosensor cell and the taste bud did not elicit a response, nor did bath perfusion with buffer alone. However, perfusing the chamber with KCl, cycloheximide, saccharin, or acetic acid evoked rapid and repeatable biosensor responses (Figs. 2_C_-E, 3_A_-C). When a first attempt to record a response was unsuccessful, repositioning the biosensor sometimes revealed signals evoked by stimuli. To date, we have obtained successful biosensor recordings from 31 of 105 taste buds when testing KCl depolarization. Eight of those 31 taste buds also yielded biosensor responses to stimulation with cycloheximide or saccharin (one of these taste buds produced a biosensor response to cycloheximide and saccharin alike). In a separate series of experiments, 6 of 13 taste buds showed responses to acetic-acid stimulation; these same taste buds also showed biosensor responses to KCl depolarization. Responses were abolished if the biosensor cell was withdrawn a few micrometers from the isolated taste bud and returned when the biosensor was repositioned to the same location. These tests identified the taste bud as the source of the release and suggested that release sites were spatially restricted within the taste bud, consistent with the presence of only a limited number of serotonergic cells (Fig. 2_A_).
Biosensor responses were generally small but were enhanced significantly by incubating lingual epithelium for 30 min with 500 μm 5-HTP, the immediate precursor to 5-HT, before isolating taste buds (Fig. 2, compare C, D). Furthermore, responses to stimuli were reversibly blocked by 1 nm mianserin (Fig. 2_F_ shows data for KCl depolarization). Because mianserin specifically blocked serotonergic responses (Fig. 1) and because 5-HTP is a precursor for serotonin, these observations verified that the biosensor cell was detecting 5-HT released from taste buds and not responding to other compounds, such as ATP, had they been secreted. We also stimulated isolated mouse taste buds with aspartame, an artificial sweetener for humans but one that is generally not preferred by and does not evoke taste nerve responses in mice (Inoue et al., 2001). Figure 2_E_ shows that aspartame, even at 10 mm, is ineffective, validating further that the biosensor responses are produced by taste stimulation per se.
Last, we tested whether serotonin release was Ca2+ dependent. Magnesium (3 mm) was substituted for Ca2+ (2 mm) in the bath, and taste buds were stimulated with KCl and tastants, as above. In the case of KCl depolarization and acid stimulation, replacing Ca2+ with Mg2+ rapidly and reversibly blocked or significantly reduced serotonin release (Fig. 3_A,B_), consistent with influx of Ca2+ through depolarization-gated Ca channels and consistent with known synaptic mechanisms. Surprisingly, serotonin release elicited by cycloheximide (Fig. 3_A_) or saccharin (Fig. 3_B_) was not affected by replacing Ca2+ with Mg2+. Bitter and nonsugar sweeteners stimulate intracellular Ca2+ release in taste cells via a signaling cascade involving phospholipase C β2 (PLCβ2) and IP3 (Bernhardt et al., 1996; Spielman et al., 1996). Thus, a likely source of Ca2+ for transmitter release elicited by these compounds was an intracellular store. Because pharmacological interventions designed to test this notion would also affect the CHO/5-HT2C biosensor, we used a different approach to answer this question. We isolated taste buds from PLCβ2 null mutant mice (Jiang et al., 1997) and tested their ability to release serotonin after stimulation. Taste buds from PLCβ2 null mice responded to bath-applied KCl, showing a normal release of serotonin (Fig. 3_D_), as above. However, we were unable to detect serotonin release evoked by cycloheximide or saccharin from taste buds of the mutant mice, consistent with the source of Ca2+ for transmitter release being intracellular stores (via a PLCβ2 signaling cascade). Specifically, we obtained successful recordings from 16 of 56 taste buds from the knock-out mice when testing KCl depolarization; none of these taste buds responded to cycloheximide or saccharin.
Discussion
This report presents data that indicate that serotonin is one of the neurotransmitters released by taste cells in response to gustatory stimulation. Furthermore, the results suggest that certain stimuli evoke neurotransmitter release in response to Ca2+ derived from intracellular stores. The postsynaptic target(s) for the serotonin remain to be determined. Serotonin may excite primary sensory afferent fibers, as proposed long ago by a number of investigators (Takeda, 1977; Esakov et al., 1983) (for review, see Nagai et al., 1996). Serotonin may also function as a paracrine hormone and act on cells within the taste bud, as discussed by Fujita et al. (1988), Ewald and Roper (1994), and more recently by Kaya et al. (2004). Our experiments were not designed to distinguish between these possibilities, both of which remain open questions.
The approach we used to detect neurotransmitters, a sensitive biosensor, has been used previously. For example, catfish horizontal cells were used to detect glutamate release from goldfish retinal bipolar cells (Tachibana and Okada, 1991) and from lizard cone photoreceptors (Savchenko et al., 1997). Myocytes have been pressed against Chinese hamster ovary fibroblasts to detect acetylcholine release (Morimoto et al., 1995). Researchers used human embryonic kidney 293 (HEK293) cells expressing the E-prostanoid receptor EP1 to monitor the release of prostaglandin E2 from macula densa cells isolated from rabbit kidneys (Peti-Peterdi et al., 2003). That group subsequently used cultured mouse mesangial cells to detect ATP released from macula densa cells (Komlosi et al., 2004). We are not aware of any group using biosensors to identify serotonin as a transmitter.
The incidence of successful serotonin release sites (e.g., for KCl stimulation, 31 of 105 taste buds) is low, presumably attributable to the relatively few numbers of serotonergic cells present in taste buds (Fig. 2), to the necessity that the taste bud was oriented such that serotonergic cells were positioned at the periphery and accessible to the biosensor, and to the requirement that the biosensor had to be placed quite near to a release site. Movement of the biosensor of only a few micrometers from a successful recording site would immediately abolish the responses, emphasizing the spatial sensitivity for placing the biosensor. It is possible that surveying a more extensive panel of taste stimuli and alternative depolarizing agents would increase the incidence of recording serotonin release sites.
Neurotransmitter release from taste cells seems to depend on two sources of Ca2+. For depolarizing stimuli (here, KCl and acid taste stimulation) (cf. Richter et al., 2003, 2004), Ca2+ influx was necessary and sufficient to stimulate serotonin release. For bitter and sweet tastants (cycloheximide and saccharin) that stimulated G-protein-coupled receptors, transmitter release persisted in the absence of extracellular Ca2+ (substituted with Mg2+). There are three possible explanations for this. (1) Replacing Ca2+ with Mg2+ in the bath did not totally eliminate Ca2+, and residual Ca2+ in the chamber, albeit low, might have sufficed to elicit transmitter release (Piccolino and Pignatelli, 1996). (2) Transmitter release in taste buds is truly Ca2+ independent, and, for example, is mediated by Na-dependent transporters, such as that which occurs at certain retinal synapses (Schwartz, 2002). (3) Transmitter release evoked by tastants is triggered by Ca2+ from internal stores. Our finding that exchanging Ca2+ for Mg2+ totally abolished depolarization-evoked release is inconsistent with the first possibility. There is a report of the serotonin transporter SET in taste buds, but it was not believed to be related to transporter-mediated transmitter release (Ren et al., 1999). Instead, our findings with the PLCβ2 knock-out mice support the last explanation. Previous studies have reported that Ca2+ derived from intracellular stores can modulate (Emptage et al., 2001; Yang et al., 2001; Simkus and Stricker, 2002; Galante and Marty, 2003) (but see Carter et al., 2002) or possibly elicit (Llano et al., 2000) neurotransmitter release at synapses. Taste bud cells may represent an excellent model for studying transmitter release triggered by Ca2+ from intracellular stores.
Note added in proof. Hayashi et al. (2004) recently reported generating biosensor cells for ATP using pheochromocytoma 12 or HEK293 cells expressing P2X2 purinergic receptors.
Footnotes
This work was supported by National Institutes of Health-National Institute on Deafness and Other Communication Disorders Grant DC006077 (S.D.R.). We thank Dr. K. Berg for the generous gift of CHO/5-HT2C cells.
Correspondence should be addressed to Dr. Stephen Roper, Department of Physiology and Biophysics, University of Miami School of Medicine, 1600 Northwest 10th Avenue, Miami, FL 33136. E-mail: roper@miami.edu.
Y.-J. Huang's present address: Department of Anatomy, College of Medicine, Kaohsiung Medical University, Kaohsiung 807, Taiwan.
K.-S. Lu's present address: Department of Anatomy and Cell Biology, College of Medicine, National Taiwan University, Taipei 100, Taiwan.
I. Plonsky's present address: The Institute for Information Transmission Problems, Russian Academy of Science, 19 Bolshoy Karetny per., Moscow 101447, Russia.
J. E. Baur's present address: Department of Chemistry, 214 Julian Hall, Illinois State University, Normal, IL 61790. D. Wu's present address: Department of Genetics and Developmental Biology, University of Connecticut Health Center, 263 Farmington Avenue, Farmington, CT 06030.
Copyright © 2005 Society for Neuroscience 0270-6474/05/250843-05$15.00/0
References
- Akiyoshi J, Nishizono A, Yamada K, Nagayama H, Mifune K, Fujii I (1995) Rapid desensitization of serotonin 5-HT2C receptor-stimulated intracellular calcium mobilization in CHO cells transfected with cloned human 5-HT2C receptors. J Neurochem 64: 2473-2479. [DOI] [PubMed] [Google Scholar]
- Akiyoshi J, Isogawa K, Yamada K, Nagayama H, Fujii I (1996) Effects of antidepressants on intracellular Ca2+ mobilization in CHO cells transfected with the human 5-HT2C receptors. Biol Psychiatry 39: 1000-1008. [DOI] [PubMed] [Google Scholar]
- Berg KA, Clarke WP, Sailstad C, Saltzman A, Maayani S (1994) Signal transduction differences between 5-hydroxytryptamine type 2A and type 2C receptor systems. Mol Pharmacol 46: 477-484. [PubMed] [Google Scholar]
- Bernhardt SJ, Naim M, Zehavi U, Lindemann B (1996) Changes in IP3 and cytosolic Ca2+ in response to sugars and non-sugar sweeteners in transduction of sweet taste in the rat. J Physiol (Lond) 490: 325-336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caicedo A, Kim KN, Roper SD (2002) Individual mouse taste cells respond to multiple chemical stimuli. J Physiol (Lond) 544: 501-509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter AG, Vogt KE, Foster KA, Regehr WG (2002) Assessing the role of calcium-induced calcium release in short-term presynaptic plasticity at excitatory central synapses. J Neurosci 22: 21-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delay RJ, Taylor R, Roper SD (1993) Merkel-like basal cells in Necturus taste buds contain serotonin. J Comp Neurol 335: 606-613. [DOI] [PubMed] [Google Scholar]
- Emptage NJ, Reid CA, Fine A (2001) Calcium stores in hippocampal synaptic boutons mediate short-term plasticity, store-operated Ca2+ entry, and spontaneous transmitter release. Neuron 29: 197-208. [DOI] [PubMed] [Google Scholar]
- Esakov AI, Golubtsov KV, Solov'eva NA (1983) Significance of serotonin in the activity of the taste receptor apparatus of the frog Rana temporaria (in Russian). Zh Evol Biokhim Fiziol 19: 62-67. [PubMed] [Google Scholar]
- Ewald DA, Roper S (1994) Bidirectional synaptic transmission in Necturus taste buds. J Neurosci 14: 3791-3801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujita T, Kanno T, Kobayashi S (1988) The paraneuron. Tokyo: Springer.
- Galante M, Marty A (2003) Presynaptic ryanodine-sensitive calcium stores contribute to evoked neurotransmitter release at the basket cell-Purkinje cell synapse. J Neurosci 23: 11229-11234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbertson TA, Boughter Jr JD, Zhang H, Smith DV (2001) Distribution of gustatory sensitivities in rat taste cells: whole-cell responses to apical chemical stimulation. J Neurosci 21: 4931-4941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi S, Hazama A, Dutta AK, Sabirov RZ, Okada Y (2004) Detecting ATP release by a biosensor method. Sci STKE 258:p 114. [DOI] [PubMed] [Google Scholar]
- Huang YJ, Maruyama Y, Lu KS, Pereira E, Roper SD (2005) Mouse taste buds release serotonin in response to taste stimuli. Chem Senses, in press. [DOI] [PubMed]
- Inoue M, McCaughey SA, Bachmanov AA, Beauchamp GK (2001) Whole nerve chorda tympani responses to sweeteners in C57BL/6ByJ and 129P3/J mice. Chem Senses 26: 915-923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang H, Kuang Y, Wu Y, Xie W, Simon MI, Wu D (1997) Roles of phospholipase C beta2 in chemoattractant-elicited responses. Proc Natl Acad Sci USA 94: 7971-7975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaya N, Shen T, Lu SG, Zhao FL, Herness S (2004) A paracrine signaling role for serotonin in rat taste buds: expression and localization of serotonin receptor subtypes. Am J Physiol Regul Integr Comp Physiol 286: R649-R658. [DOI] [PubMed] [Google Scholar]
- Kim DJ, Roper SD (1995) Localization of serotonin in taste buds: a comparative study in four vertebrates. J Comp Neurol 353: 364-370. [DOI] [PubMed] [Google Scholar]
- Komlosi P, Peti-Peterdi J, Fuson AL, Fintha A, Rosivall L, Bell PD (2004) Macula densa basolateral ATP release is regulated by luminal [NaCl] and dietary salt intake. Am J Physiol Renal Physiol 286: F1054-F1058. [DOI] [PubMed] [Google Scholar]
- Llano I, Gonzalez J, Caputo C, Lai FA, Blayney LM, Tan YP, Marty A (2000) Presynaptic calcium stores underlie large-amplitude miniature IPSCs and spontaneous calcium transients. Nat Neurosci 3: 1256-1265. [DOI] [PubMed] [Google Scholar]
- Morimoto T, Popov S, Buckley KM, Poo MM (1995) Calcium-dependent transmitter secretion from fibroblasts: modulation by synaptotagmin I. Neuron 15: 689-696. [DOI] [PubMed] [Google Scholar]
- Nagai T, Kim DJ, Delay RJ, Roper SD (1996) Neuromodulation of transduction and signal processing in the end organs of taste. Chem Senses 21: 353-365. [DOI] [PubMed] [Google Scholar]
- Nagai T, Delay RJ, Welton J, Roper SD (1998) Uptake and release of neurotransmitter candidates, [3H]serotonin, [3H]glutamate, and [3H]gamma-aminobutyric acid, in taste buds of the mudpuppy, _Necturus maculosus._J Comp Neurol 392: 199-208. [PubMed] [Google Scholar]
- Peti-Peterdi J, Komlosi P, Fuson AL, Guan Y, Schneider A, Qi Z, Redha R, Rosivall L, Breyer MD, Bell PD (2003) Luminal NaCl delivery regulates basolateral PGE2 release from macula densa cells. J Clin Invest 112: 76-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piccolino M, Pignatelli A (1996) Calcium-independent synaptic transmission: artifact or fact? Trends Neurosci 19: 120-125. [DOI] [PubMed] [Google Scholar]
- Ren Y, Shimada K, Shirai Y, Fujimiya M, Saito N (1999) Immunocytochemical localization of serotonin and serotonin transporter (SET) in taste buds of rat. Brain Res Mol Brain Res 74: 221-224. [DOI] [PubMed] [Google Scholar]
- Richter TA, Caicedo A, Roper SD (2003) Sour taste stimuli evoke Ca2+ and pH responses in mouse taste cells. J Physiol (Lond) 547: 475-483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richter TA, Dvoryanchikov GA, Chaudhari N, Roper SD (2004) Acid-sensitive two-pore domain potassium (K2P) channels in mouse taste buds. J Neurophysiol 92: 1928-1936. [DOI] [PubMed] [Google Scholar]
- Savchenko A, Barnes S, Kramer RH (1997) Cyclic-nucleotide-gated channels mediate synaptic feedback by nitric oxide. Nature 390: 694-698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz EA (2002) Transport-mediated synapses in the retina. Physiol Rev 82: 875-891. [DOI] [PubMed] [Google Scholar]
- Simkus CR, Stricker C (2002) The contribution of intracellular calcium stores to mEPSCs recorded in layer II neurones of rat barrel cortex. J Physiol (Lond) 545: 521-535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spielman AI, Nagai H, Sunavala G, Dasso M, Breer H, Boekhoff I, Huque T, Whitney G, Brand JG (1996) Rapid kinetics of second messenger production in bitter taste. Am J Physiol 270: C926-C931. [DOI] [PubMed] [Google Scholar]
- Tachibana M, Okada T (1991) Release of endogenous excitatory amino acids from ON-type bipolar cells isolated from the goldfish retina. J Neurosci 11: 2199-2208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeda M (1977)) Uptake of 5-hydroxytryptophan by gustatory cells in the mouse taste bud. Arch Histol Jpn 40: 243-250. [DOI] [PubMed] [Google Scholar]
- Whitear M (1989) Merkel cells in lower vertebrates. Arch Histol Cytol 52: 415-422. [DOI] [PubMed] [Google Scholar]
- Yang F, He X, Feng L, Mizuno K, Liu XW, Russell J, Xiong WC, Lu B (2001) PI-3 kinase and IP3 are both necessary and sufficient to mediate NT3-induced synaptic potentiation. Nat Neurosci 4: 19-28. [DOI] [PubMed] [Google Scholar]
- Yee CL, Yang R, Bottger B, Finger TE, Kinnamon JC (2001) “Type III” cells of rat taste buds: immunohistochemical and ultrastructural studies of neuron-specific enolase, protein gene product 9.5, and serotonin. J Comp Neurol 440: 97-108. [DOI] [PubMed] [Google Scholar]