Intraflagellar Transport: The Eyes Have It (original) (raw)
The Problem: An Organelle whose Assembly Site Is Distant from the Site of Protein Synthesis
Unlike most organelles, which are surrounded by cytoplasm, the flagellum protrudes from the cell surface extending tens or even hundreds of microns into the external medium. This elongated organelle must import all the macromolecules required for its assembly, maintenance, and function including >200 polypeptides that make up the microtubular axoneme (Dutcher, 1995), all the constituents of the flagellar membrane, as well as a prodigious amount of ATP to supply the thousands of dynein motors that drive flagellar motility.
A dramatic example of the delivery of molecules into the flagellum is seen during flagellar regeneration in the biflagellate alga Chlamydomonas: flagella 10 μm long are assembled in ∼1 h. As the organelle elongates, flagellar precursors must reach the site of assembly at the distal tip (Rosenbaum and Child, 1967; Johnson and Rosenbaum, 1992), which grows farther and farther away from the site of protein synthesis. The site of tubulin addition during flagellar assembly was identified by fusing cells with half-length flagella to cells containing epitope-tagged tubulin: all the tagged tubulin incorporated into the growing flagella at their distal tips. When cells with full-length flagella lacking radial spokes were fused to wild-type cells, radial spokes from the wild-type cytoplasm entered the spokeless flagella, assembled at the distal tips of the flagella, and gradually continued assembly toward the base (Johnson and Rosenbaum, 1992). Similar results were obtained with inner dynein arms (Piperno et al., 1996). Thus, there appears to be a mechanism for transporting axonemal precursors to the distal tip of the flagellum, whether or not it is elongating.
Extensive transport also occurs into nonmotile sensory cilia of certain neuronal cells. The outer segments of retinal rods (ROS),1 for example, are modified cilia replete with the membrane-associated photoreceptor machinery for receiving and transducing light signals. These stacks of membranes are constantly being replenished: in mouse, the membranes of the ROS are completely replaced every 2 wk. The only connection between the ROS and the rod inner segment (RIS), in which synthesis of all the components occurs, is the connecting cilium composed of a membrane bound 9+0 axoneme. All the materials required for continual turnover of the ROS must pass through the connecting cilium.
The following briefly reviews recent literature that addresses how the cell rapidly mobilizes over 200 polypeptides required for flagellar assembly, and how it transports these polypeptides to the flagellar tip assembly site.
The Solution: Intraflagellar Transport of Preassembled Flagellar Complexes
Many flagellar proteins exist as complexes of multiple polypeptides, e.g., the radial spokes contain 17 polypeptides and the outer dynein arms contain >15 polypeptides (Dutcher, 1995); it would be advantageous for the cell to preassemble these axonemal structures in the cytoplasm rather than to send individual polypeptides into the flagella for assembly. In keeping with this idea, assembled radial spoke complexes have been found in the cytoplasm of Chlamydomonas (Diener, D.R., D.G. Cole, and J.L. Rosenbaum. 1996. ASCB Meeting, San Francisco. Abstract 273). Similarly, flagellar outer dynein arms from Chlamydomonas and Paramecium also preassemble in the cytoplasm (Fok et al., 1994; Fowkes and Mitchell, 1998). Because during flagellar assembly these flagellar precursor complexes assemble at the flagellar tip, a mechanism to transport large protein complexes through the flagellum was hypothesized.
Intraflagellar transport (IFT) appears to be the mechanism that moves flagellar precursors to the flagellar tip. IFT, visualized with high-resolution video-enhanced differential interference-contrast (DIC) microscopy, is a motility located between the flagellar membrane and axoneme (Kozminski et al., 1993, 1995, 1998). Particles of variable size travel to the flagellar tip (anterograde transport) at 2.0 μm/s and smaller particles return from the tip to the base (retrograde transport) at 3.5 μm/s (Kozminski et al., 1993). The movement is continuous and linear along the entire flagella. IFT is neither affected by mutations that cause immotile flagella, e.g., mutations affecting dynein arms, radial spokes, or central pair microtubules, nor does it appear to be related to other motilities associated with the flagellar membrane, e.g., gliding of whole cells on surfaces by means of their outstretched flagella or the movement of polystyrene beads on the flagellar surface (Bloodgood, 1992; Kozminski et al., 1993). IFT continues unabated in cells in which gliding or bead movement has been blocked chemically (Kozminski et al., 1993) or genetically (Kozminski, 1995). All three motitities, however, are reversibly inhibited by increasing the osmolarity with NaCl (100 mM) or sucrose (6%) (Kozminski et al., 1993).
Thin sectioning and electron microscopy indicate that the material moving beneath the flagellar membrane by IFT consists of “lollipop-shaped” particles occurring in groups of varying particle numbers, called “rafts” (Fig. 1; Kozminski et al., 1993). The rafts, initially observed by Ringo (1967) during ultrastructural analysis of the Chlamydomonas flagellum, are attached by a thin connection to the B-subfibers of the outer-doublet microtubules and to the overlying flagellar membrane (Kozminski et al., 1993). Electron microscopy of IFT particles observed by DIC in the flagellum of a single embedded cell confirmed that the particles observed by DIC are, indeed, the rafts observed by electron microscopy (Kozminski et al., 1995).
Figure 1.
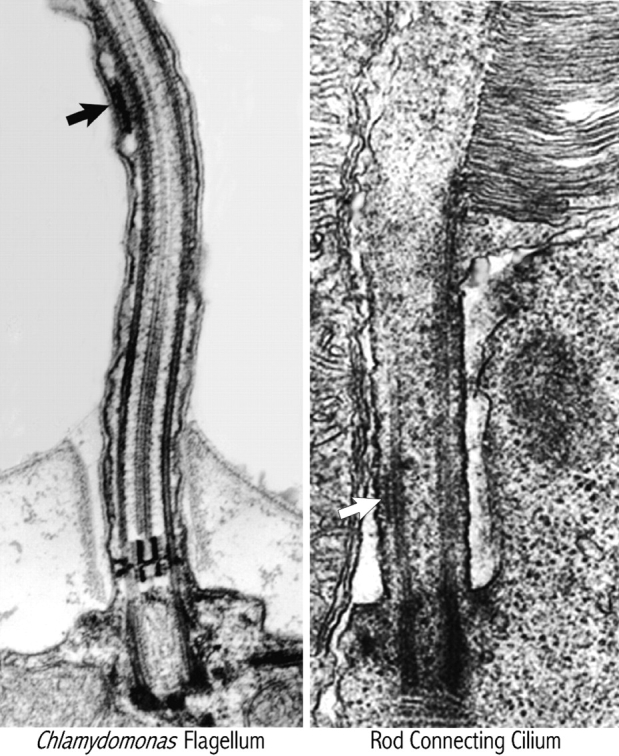
Transmission electron micrographs of a Chlamydomonas flagellum (micrograph courtesy of Karl A. Johnson) and a vertebrate rod connecting cilium (reprinted with permission from Sandborn, 1970). Note the presence of a raft (arrows) in both organelles.
Chlamydomonas flagella contain several kinesins (Bernstein et al., 1994; Fox et al., 1994; Johnson et al., 1994; Kozminski et al., 1995; Walther et al., 1994). One of these, known as FLA10 or khp1 (Walther et al., 1994), is located between the flagellar membrane and axoneme where IFT occurs (Kozminski, 1995). A temperature-sensitive mutant defective in this flagellar kinesin, fla10, cannot assemble flagella at the restrictive temperature; in fact, existing flagella shorten and disappear at the restrictive temperature (Huang et al., 1977; Adams et al., 1982; Lux and Dutcher, 1991). Just before flagellar shortening, IFT decreases and disappears in these cells; simultaneously, ∼70% of the rafts observed by electron microscopy also disappear (Kozminski et al., 1995; Vashishtha et al., 1996). Together, these results indicate that (a) the IFT particles observed by DIC microscopy are the rafts observed in thin sections by electron microscopy; (b) the movement of the rafts is dependent on FLA10; and (c) the movement of the rafts, IFT, is essential for assembly and maintenance of the flagella.
Sequence analysis reveals that FLA10 is most closely related to the kinesin-II subfamily of kinesins; some of these kinesins have been purified as heterotrimeric complexes consisting of two distinct, though related, motor subunits and one nonmotor subunit (Scholey, 1996). FLA10 is part of a similar heterotrimeric complex, FLA10 kinesin-II (Cole et al., 1998). As is typical for kinesins and their cargoes, FLA10 kinesin-II does not copurify with IFT particles; however, a small amount of FLA10 coprecipitates with immunoprecipitates of IFT particle polypeptides, suggesting a weak interaction between FLA10 kinesin-II and IFT particles (Cole et al., 1998).
The IFT particles were purified from flagellar extracts taking advantage of the fact that they are greatly decreased in flagella of fla10 cells maintained at the restrictive temperature. 15 polypeptides sedimenting at ∼16 S are reduced in flagella of fla10 cells incubated at 32°C (Piperno and Mead, 1997; Cole et al., 1998). These 15 polypeptides form two complexes: complex A, composed of 4 polypeptides; and complex B, composed of 11 polypeptides. Analysis of new mutants with defects in IFT have identified a fifth polypeptide in complex A (Piperno et al., 1998). Considering the size of the IFT particles seen in the electron microscope, there are probably multiple copies of the 16 S complexes in each single IFT particle, several of which compose the rafts.
Perhaps the strongest evidence that the 16 S particles are, indeed, the IFT particles observed in DIC, came from the work of Pazour et al. (1998). They isolated a Chlamydomonas mutant, fla14, that lacks LC8, a component of both flagellar (Piperno and Luck, 1979) and cytoplasmic dyneins (King et al., 1996). In fla14, kinesin-powered anterograde IFT is normal, but retrograde particle movement is missing (Pazour et al., 1998). The flagella of fla14 are immotile, approximately half length, and deficient in dynein arms and radial spokes. Most importantly, these flagella contain massive accumulations of the rafts, and, biochemically, contain 10–20-fold the amount of IFT particle polypeptides and FLA10 found in wild-type flagella (Pazour et al., 1998). Apparently, the rafts, composed of the 16 S particles, are brought into the flagella by the FLA10 kinesin-II, and accumulate because they cannot be moved out in the absence of retrograde IFT.
Although analysis of fla14 suggested that cytoplasmic dynein was the retrograde IFT motor, this interpretation was clouded by the fact that LC8 is also a component of myosin-V (Espindola, F.S., R.E. Cheney, S.M. King, D.M. Suter, and M.S. Mooseker. 1996. ASCB Meeting, San Francisco. Abstract 2160) and flagellar inner and outer arm dyneins (Piperno and Luck, 1979; Harrison et al., 1998). The importance of cytoplasmic dynein in IFT has now been more convincingly shown by mutations in DHC1b, a cytoplasmic dynein heavy chain. The effect of these mutations is similar to, though even more severe than, fla14: the flagella of these mutants, dhc1b, stfl-1, and stfl-2 are only 1–2 μm long and the space between the axoneme and the flagellar membrane is filled with rafts (Pazour et al., 1999; Porter et al., 1999). Therefore, IFT represents one of the only microtubule-based motility systems in which there are mutants in the motors responsible for both the anterograde and retrograde transport of defined, isolable particles.
The Cellular Localization of IFT Particles and Motors
Because FLA10 kinesin-II and IFT polypeptide antigens were obtained from isolated Chlamydomonas flagella, it was a surprise to find that their principal location in wild-type cells was not in the flagella but, rather, in the region of the flagellar basal bodies. In wild-type cells, immunofluorescent localization of IFT polypeptides and FLA10 appears to be in a tripartite or ring-like structure around the basal bodies (Vashishtha et al., 1996; Cole et al., 1998). In dhc1b the IFT particle proteins are redistributed from the peri-basal body region to the flagella (Pazour et al., 1999). Apparently, the IFT particles can enter the flagella in these retrograde transport mutants but cannot exit, leading to a buildup in the flagella and a decrease around the basal bodies. In a Chlamydomonas mutant, bld2, lacking basal bodies (Goodenough and St. Clair, 1975), the IFT particle polypeptides continue to localize around the position where the basal bodies would have been, but FLA10 does not (Cole et al., 1998). Furthermore, in fla10 cells maintained at the restrictive temperature, the IFT particle polypeptides still accumulate around the basal bodies. Therefore, the basal body region appears to be a holding area for IFT particles and the motors that move them; however, delivery and accumulation of IFT particles to this region are not dependent on FLA10 kinesin-II.
IFT in Other Motile Cilia
Several lines of evidence suggest kinesin-II is involved in ciliogenesis in a wide variety of organisms. Injection of an antibody against a motor subunit of kinesin-II into sea urchin embryos disrupts formation of cilia that normally appear in the blastula stage (Morris and Scholey, 1997). In Tetrahymena, simultaneous genetic knockout of two kinesin-II motor subunits completely blocked formation of cilia (Brown, J., C. Marsala, R. Kosoy, and J. Gaertig. 1998. ASCB Meeting, San Francisco. Abstract 173). Disruption of a kinesin-II motor subunit, KIF3B, in mice resulted in the lack of cilia in the nodal cells of 7.5-d postcoitum embryos, leading to the randomization of left–right asymmetry of the embryo (Nonaka et al., 1998), a characteristic associated with immotile cilia syndrome in humans (Afzelius, 1976). Expression of DHC1b increases during ciliogenesis in sea urchin embryos (Gibbons et al., 1994) and rat tracheal epithelial cells (Criswell et al., 1996), suggesting that this cytoplasmic dynein heavy chain plays a role in assembly of cilia in these organisms.
IFT in Nonmotile Sensory Cilia
Homologues of several Chlamydomonas IFT particle proteins have been identified in C. elegans through peptide microsequencing (Cole et al., 1998). The C. elegans mutants osm1 and osm6 have mutations in the genes encoding homologues of IFT polypeptides p172 and p52 (Cole et al., 1998), respectively. In these C. elegans mutants, the ability to sense osmotic gradients and chemoattractants is lacking. The basis for the mutant phenotype at the cellular level is that the sensory cilia found at dendritic ends of sensory neurons do not assemble properly (Perkins et al., 1986).
Interestingly, similar phenotypes are found in two additional C. elegans mutants, osm3 and che3, which have defects in a subunit of heterotrimeric kinesin-II (Shakir et al., 1993; Tabish et al., 1995) and in the cytoplasmic dynein heavy chain DHC1b (Grant, W., personal communication), respectively. Thus, IFT particle proteins and the motors that move them, kinesin-II and cytoplasmic dynein DHC1b, are essential for the assembly of neuronal nonmotile sensory cilia of C. elegans.
IFT particles and their motors also appear to be present in vertebrate cilia-containing sensory neurons. Using antibodies to KIF3A, kinesin-II has been localized by fluorescence and immunogold electron microscopies in the connecting cilium between the RIS and ROS of fish retinal rod cells (Beech et al., 1996). Rafts similar to those observed in flagella of Chlamydomonas can also be seen in the connecting cilia in the vertebrate eye (Fig. 1; Sandborn, 1970). Mouse and human expressed sequence tags with homologies to IFT particle polypeptides have also been identified (Cole, D., unpublished observations), so IFT may play a similar role in mammals. Recently, targeted knockout of a kinesin-II (KIF3A) in the mouse retina resulted in degeneration of ROS, starting at the proximal portion where new material is added (Marszalek, J.R., X. Liu, E. Roberts, D. Chui, J. Marth, D.S. Williams, and L.S.B. Goldstein. 1998. ASCB Meeting, San Francisco. Abstract 756). Thus, kinesin-powered IFT is probably present in the connecting cilium between the RIS and the ROS of vertebrates and ROS assembly and maintenance is apparently dependent on IFT. The implications of this for studies of retinal pathologies leading to blindness are clear.
Hypothesis: The Role of IFT in the Assembly and Maintenance of Flagella
One possible role of IFT in flagellar assembly is to transport flagellar precursors to the flagellar tip where assembly occurs. Flagellar proteins appear to be synthesized on mRNAs localized close to the basal bodies (Han, 1997; W. Marshall, unpublished results), and proteins that form various flagellar substructures, such as the dynein arms and radial spokes, preassemble in the cytoplasm (Fok et al., 1994; Fowkes and Mitchell, 1998; D.R. Diener, D.G. Cole, and J.L. Rosenbaum. 1996. ASCB Meeting, San Francisco. Abstract 273). The molecular motors, FLA10 kinesin-II and cytoplasmic dynein DHC1b, responsible for transporting these precursors into and out of the flagella and the IFT particle polypeptides are concentrated around the basal bodies (Vashishtha et al., 1996; Cole et al., 1998; Pazour et al., 1999) and may become associated with each other as well as with the flagellar precursors in this region. The associations between the IFT particles and the precursors are likely to be weak to facilitate release of the precursors in the flagellar compartment. The IFT particles and flagellar precursors are moved into the flagella by the anterograde motor FLA10 kinesin-II and are recycled to the cell body by cytoplasmic dynein DHC1b. How the switch is made at the flagellar tip from the anterograde to the retrograde motor remains a provocative question.
Although the above discussion has emphasized the importance of IFT in transporting flagellar axonemal precursors into the flagella, it is important to note that (a) the IFT rafts are clearly associated with the flagellar membrane as well as the B tubule of the outer doublets; (b) axonemal components, e.g., radial spokes, dynein arms, and central pair microtubules, are absent from immotile sensory cilia of C. elegans and vertebrate photoreceptors; and (c) the first phenotype that appears (in ∼30 min) in fla10 cells at the restrictive temperature is a membrane defect: the cells lose the ability to mate (Piperno et al., 1996) by use of their flagella. It is known that the flagellar mating molecules must be moved onto the flagellar surface and activated before mating (Hunnicutt et al., 1990). Therefore, IFT is almost certainly involved in flagellar membrane maintenance and function.
IFT is required for the assembly and maintenance of Chlamydomonas flagella and probably functions in a similar manner in the formation of the ciliated sensory neurons in C. elegans and other higher organisms including vertebrates (Kozminski et al., 1995; Cole et al., 1998; J.R. Marszalek, X. Liu, E. Roberts, D. Chui, J. Marth, D.S. Williams, and L.S.B. Goldstein. 1998. ASCB Meeting, San Francisco. Abstract 756). Exactly how IFT functions in the assembly and maintenance of motile cilia and nonmotile sensory cilia and what roles the individual IFT particle polypeptides play in this process are not yet known.
Acknowledgments
The authors would like to thank R. Bloodgood, J. Gaertig, W. Grant, L. Goldstein, K. Kozminski, G. Pazour, M. Porter, J. Scholey, and G. Witman for sharing unpublished data and/or for comments on the manuscript. Thanks also to K. Johnson for providing the micrograph of Chlamydomonas shown in Fig. 1.
Abbreviations used in this paper
DIC
differential interference-contrast
IFT
intraflagellar transport
RIS
rod inner segment
ROS
rod outer segment
Footnotes
Douglas G. Cole's current address is Department of Microbiology, Molecular Biology and Biochemistry, University of Idaho, Moscow, ID 83844-3052.
References
- Adams GMW, Huang B, Luck DJL. Temperature-sensitive, assembly-defective flagella mutants of Chlamydomonas reinhardtii. . Genetics. 1982;100:579–586. doi: 10.1093/genetics/100.4.579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Afzelius BA. A human syndrome caused by immotile cilia. Science. 1976;193:317–319. doi: 10.1126/science.1084576. [DOI] [PubMed] [Google Scholar]
- Beech PL, Pagh-Roehl K, Noda Y, Hirokawa N, Burnside B, Rosenbaum JL. Localization of kinesin super family proteins to the connecting cilium of fish photoreceptors. J Cell Sci. 1996;109:889–897. doi: 10.1242/jcs.109.4.889. [DOI] [PubMed] [Google Scholar]
- Bernstein M, Beech PL, Katz SG, Rosenbaum JL. A new kinesin-like protein (Klp1) localized to a single microtubule of the Chlamydomonasflagellum. J Cell Biol. 1994;125:1313–1326. doi: 10.1083/jcb.125.6.1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloodgood RA. Directed movements of ciliary and flagellar membrane components: a review. Biol Cell. 1992;76:291–301. doi: 10.1016/0248-4900(92)90431-y. [DOI] [PubMed] [Google Scholar]
- Cole DG, Diener DR, Himelblau AL, Beech PL, Fuster JC, Rosenbaum JL. Chlamydomonas kinesin-II–dependent intraflagellar transport (IFT): IFT particles contain proteins required for ciliary assembly in Caenorhabditis eleganssensory neurons. J Cell Biol. 1998;141:993–1008. doi: 10.1083/jcb.141.4.993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Criswell PS, Ostrowski LE, Asai DJ. A novel cytoplasmic dynein heavy chain: expression of DHC1b in mammalian ciliated epithelial cells. J Cell Sci. 1996;109:1891–1898. doi: 10.1242/jcs.109.7.1891. [DOI] [PubMed] [Google Scholar]
- Dutcher SK. Flagellar assembly in two hundred and fifty easy-to-follow steps. Trends Genet. 1995;11:398–404. doi: 10.1016/s0168-9525(00)89123-4. [DOI] [PubMed] [Google Scholar]
- Fok AK, Wang H, Katayama A, Aihara MS, Allen RD. 22S axonemal dynein is preassembled and functional prior to being transported to and attached on the axonemes. Cell Motil Cytoskel. 1994;29:215–224. doi: 10.1002/cm.970290304. [DOI] [PubMed] [Google Scholar]
- Fowkes ME, Mitchell DR. The role of preassembled cytoplasmic complexes in assembly of flagellar dynein subunits. Mol Biol Cell. 1998;9:2337–2347. doi: 10.1091/mbc.9.9.2337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox LA, Sawin KE, Sale WS. Kinesin-related proteins in eukaryotic flagella. J Cell Sci. 1994;107:1545–1550. doi: 10.1242/jcs.107.6.1545. [DOI] [PubMed] [Google Scholar]
- Gibbons BH, Asai DJ, Tang WJ, Hays TS, Gibbons IR. Phylogeny and expression of axonemal and cytoplasmic dynein genes in sea urchins. Mol Biol Cell. 1994;5:57–70. doi: 10.1091/mbc.5.1.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodenough UW, St. Clair HS. Bald-2: a mutant affecting the formation of doublet and triplet sets of microtubules in Chlamydomonas reinhardtii. . J Cell Biol. 1975;66:480–491. doi: 10.1083/jcb.66.3.480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han JW, Park JH, Kim M, Lee J. mRNAs for microtubule proteins are specifically colocalized during the sequential formation of basal body, flagella, and cytoskeletal microtubules in the differentiation of Naegleria gruberi. . J Cell Biol. 1997;137:871–879. doi: 10.1083/jcb.137.4.871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison A, Olds-Clarke P, King SM. Identification of the t complex–encoded cytoplasmic dynein light chain Tctex1in inner arm I1 supports the involvement of flagellar dyneins in meiotic drive. J Cell Biol. 1998;140:1137–1147. doi: 10.1083/jcb.140.5.1137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang B, Rifkin MR, Luck DJL. Temperature-sensitive mutations affecting flagellar assembly and function in Chlamydomonas reinhardtii. . J Cell Biol. 1977;72:67–85. doi: 10.1083/jcb.72.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunnicutt GR, Kosfiszer MG, Snell WJ. Cell body and flagellar agglutinins in Chlamydomonas reinhardtii: the cell body plasma membrane is a reservoir for agglutinins whose migration to the flagella is regulated by a functional barrier. J Cell Biol. 1990;111:1605–1616. doi: 10.1083/jcb.111.4.1605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson KA, Rosenbaum JL. Polarity of flagellar assembly in Chlamydomonas. . J Cell Biol. 1992;119:1605–1611. doi: 10.1083/jcb.119.6.1605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson KA, Haas MA, Rosenbaum JL. Localization of a kinesin-related protein to the central pair apparatus of the Chlamydomonas reinhardtiiflagellum. J Cell Sci. 1994;107:1551–1556. doi: 10.1242/jcs.107.6.1551. [DOI] [PubMed] [Google Scholar]
- King SM, Barbarese E, Dillman JF, III, Patel-King RS, Carson JH, Pfister KK. Brain cytoplasmic and flagellar outer arm dyneins share a highly conserved Mr 8,000 light chain. J Biol Chem. 1996;271:19358–19366. doi: 10.1074/jbc.271.32.19358. [DOI] [PubMed] [Google Scholar]
- Kozminski, K.G. 1995. Beat-independent flagellar motilities in Chlamydomonas and an analysis of the function of alpha-tubulin acetylation. Ph.D. thesis. Department of Biology, Yale University, New Haven, CT. 151 pp.
- Kozminski KG, Johnson KA, Forscher P, Rosenbaum JL. A motility in the eukaryotic flagellum unrelated to flagellar beating. Proc Natl Acad Sci USA. 1993;90:5519–5523. doi: 10.1073/pnas.90.12.5519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozminski KG, Beech PL, Rosenbaum JL. The Chlamydomonaskinesin-like protein FLA10 is involved in motility associated with the flagellar membrane. J Cell Biol. 1995;131:1517–1527. doi: 10.1083/jcb.131.6.1517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozminski, K.G., P. Forscher, and J.L. Rosenbaum. 1998. Three flagellar motilities in Chlamydomonas unrelated to flagellar beating. Cell Motil. Cytoskel. (Video Suppl. 5). 39:347–348. [PubMed]
- Lux FG, III, Dutcher SK. Genetic interactions at the FLA10 locus: suppressors and synthetic phenotypes that affect the cell cycle and flagellar function in Chlamydomonas reinhardtii. . Genetics. 1991;128:549–561. doi: 10.1093/genetics/128.3.549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris RL, Scholey JM. Heterotrimeric kinesin-II is required for the assembly of motile 9+2 ciliary axonemes on sea urchin embryos. J Cell Biol. 1997;138:1009–1022. doi: 10.1083/jcb.138.5.1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nonaka S, Tanaka Y, Okada Y, Takeda S, Harada A, Kanai Y, Kido M, Hirokawa N. Randomization of right-left asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell. 1998;95:829–837. doi: 10.1016/s0092-8674(00)81705-5. [DOI] [PubMed] [Google Scholar]
- Pazour GJ, Wilkerson CG, Witman GB. A dynein light chain is essential for retrograde particle movement in intraflagellar transport (IFT) J Cell Biol. 1998;141:979–992. doi: 10.1083/jcb.141.4.979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pazour GJ, Dickert BL, Witman GB. The DHC1b (DHC2) isoform of cytoplasmic dynein is required for flagellar assembly. J Cell Biol. 1999;144:473–481. doi: 10.1083/jcb.144.3.473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkins LA, Hedgecock EM, Thomson JN, Culotti JG. Mutant sensory cilia in the nematode Caenorhabditis elegans. . Dev Biol. 1986;117:456–487. doi: 10.1016/0012-1606(86)90314-3. [DOI] [PubMed] [Google Scholar]
- Piperno G, Luck DJL. Axonemal adenosine triphosphatases from flagella of Chlamydomonas reinhardtii. Purification of two dyneins. J Biol Chem. 1979;254:3084–3090. [PubMed] [Google Scholar]
- Piperno G, Mead K. Transport of a novel complex in the cytoplasmic matrix of Chlamydomonasflagella. Proc Natl Acad Sci USA. 1997;94:4457–4462. doi: 10.1073/pnas.94.9.4457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno G, Mead K, Henderson S. Inner dynein arms but not outer dynein arms require the activity of kinesin homologue protein KHP1FLA10 to reach the distal part of flagella in Chlamydomonas. . J Cell Biol. 1996;133:371–379. doi: 10.1083/jcb.133.2.371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piperno G, Siuda E, Henderson S, Segil M, Vaananen H, Sassaroli M. Distinct mutants of retrograde intraflagellar transport (IFT) share similar morphological and molecular defects. J Cell Biol. 1998;143:1591–1601. doi: 10.1083/jcb.143.6.1591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porter, M.E., R. Bower, J.A. Knott, P. Byrd, and W. Dentler. 1999. Cytoplasmic dynein heavy chain 1b is required for flagellar assembly in Chlamydomonas. Mol. Biol. Cell. In press. [DOI] [PMC free article] [PubMed]
- Ringo DL. Flagellar motion and fine structure of the flagellar apparatus in Chlamydomonas. . J Cell Biol. 1967;33:543–571. doi: 10.1083/jcb.33.3.543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenbaum JL, Child FM. Flagellar regeneration in protozoan flagellates. J Cell Biol. 1967;34:345–364. doi: 10.1083/jcb.34.1.345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandborn, E.B. 1970. Cells and Tissues by Light and Electron Microscopy. Vol. 1. Academic Press, New York. 366 pp.
- Scholey JM. Kinesin-II, a membrane traffic motor in axons, axonemes, and spindles. J Cell Biol. 1996;133:1–4. doi: 10.1083/jcb.133.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shakir MA, Fukushige T, Yasuda H, Miwa J, Siddiqui SS. C. elegans osm-3gene mediating osmotic avoidance behaviour encodes a kinesin-like protein. Neuroreport. 1993;4:891–894. doi: 10.1097/00001756-199307000-00013. [DOI] [PubMed] [Google Scholar]
- Tabish M, Siddiqui ZK, Nishikawa K, Siddiqui SS. Exclusive expression of C. elegans osm-3kinesin gene in chemosensory neurons open to the external environment. J Mol Biol. 1995;247:377–389. doi: 10.1006/jmbi.1994.0146. [DOI] [PubMed] [Google Scholar]
- Vashishtha M, Walther Z, Hall JL. The kinesin-homologous protein encoded by the Chlamydomonas FLA10gene is associated with basal bodies and centrioles. J Cell Sci. 1996;109:541–549. doi: 10.1242/jcs.109.3.541. [DOI] [PubMed] [Google Scholar]
- Walther Z, Vashishtha M, Hall JL. The Chlamydomonas FLA10gene encodes a novel kinesin-homologous protein. J Cell Biol. 1994;126:175–188. doi: 10.1083/jcb.126.1.175. [DOI] [PMC free article] [PubMed] [Google Scholar]