Skeletal Malformations Caused by Overexpression of Cbfa1 or Its Dominant Negative Form in Chondrocytes (original) (raw)
Abstract
During skeletogenesis, cartilage develops to either permanent cartilage that persists through life or transient cartilage that is eventually replaced by bone. However, the mechanism by which cartilage phenotype is specified remains unclarified. Core binding factor α1 (Cbfa1) is an essential transcription factor for osteoblast differentiation and bone formation and has the ability to stimulate chondrocyte maturation in vitro. To understand the roles of Cbfa1 in chondrocytes during skeletal development, we generated transgenic mice that overexpress Cbfa1 or a dominant negative (DN)-Cbfa1 in chondrocytes under the control of a type II collagen promoter/enhancer. Both types of transgenic mice displayed dwarfism and skeletal malformations, which, however, resulted from opposite cellular phenotypes. Cbfa1 overexpression caused acceleration of endochondral ossification due to precocious chondrocyte maturation, whereas overexpression of DN-Cbfa1 suppressed maturation and delayed endochondral ossification. In addition, Cbfa1 transgenic mice failed to form most of their joints and permanent cartilage entered the endochondral pathway, whereas most chondrocytes in DN-Cbfa1 transgenic mice retained a marker for permanent cartilage. These data show that temporally and spatially regulated expression of Cbfa1 in chondrocytes is required for skeletogenesis, including formation of joints, permanent cartilages, and endochondral bones.
Keywords: chondrocyte, Cbfa1, transgenic mice, endochondral ossification, permanent cartilage
Introduction
Vertebrate skeletons are constructed through the formation of bone structures at specific times and sites, a process that is achieved by intramembranous or endochondral ossification. Intramembranous bones derive from the direct differentiation of mesenchymal cells into osteoblasts, whereas endochondral bones first appear as cartilaginous structures that are then replaced with bone (Gilbert 1997). Most of the cartilage has transient phenotype and the chondrocytes proliferate, then enlarge into hypertrophic cells, and are finally replaced by osteoblasts during endochondral ossification. However, chondrocytes present in developing articular cartilage, tracheal rings, nasal septum, and intervertebral discs do not enter the maturation and endochondral processes and these cartilaginous structures thus persist throughout life as permanent cartilage (Pacifici 1995). Chondrocytes undergoing the endochondral process are responsible for formation and growth of most bones, including longitudinal bones, vertebrae, and ribs, whereas chondrocytes that organize permanent cartilage tissues exert a continuous function, including providing resilience to joints and tracheal rings and allowing normal biomechanical movements. Therefore, tight regulation of cartilage phenotype specification is very important for assembly and function of skeletal structures. However, it has not been fully understood how chondrocytes enter the endochondral process or acquire and maintain the permanent phenotype.
Core binding factor α1 (Cbfa1)/Runx2 (runt-related gene 2) is an essential transcription factor in osteoblast differentiation and bone formation (Ducy et al. 1997; Komori et al. 1997; Otto et al. 1997). _Cbfa1_-deficient mice display a complete lack of both intramembranous and endochondral ossification and show an absence of osteoblast differentiation, as well as retardation of chondrocyte differentiation (Komori et al. 1997; Otto et al. 1997; Inada et al. 1999; Kim et al. 1999). Further, Cbfa1 has the ability to stimulate chondrocyte maturation in vitro (Enomoto et al. 2000). Thus, a main function of Cbfa1 in chondrocytes in vivo appears to be promotion of their maturation and endochondral ossification. However, it may be that chondrocyte differentiation leading to endochondral ossification is largely controlled by the surrounding cells, osteoblastic and hematopoietic cells, and that a lack of Cbfa1 in chondrocytes may not be a major reason for suppression of chondrocyte maturation. Thus, the mechanism of inhibition of endochondral ossification seen in _Cbfa1_-deficient mice remains to be solved.
To examine the roles of Cbfa1 in endochondral ossification and cartilage phenotype specification, we generated transgenic mice, which overexpress Cbfa1 or a dominant negative (DN)-Cbfa1 form in chondrocytes under the control of promoter/enhancer elements of the α1 type II collagen gene (Col2a1). Cbfa1 and DN-Cbfa1 transgenic mice showed, respectively, accelerated and decelerated chondrocyte maturation and endochondral ossification. Further, permanent cartilage lost its permanent phenotype and entered into the endochondral process in Cbfa1 transgenic mice, whereas most of the chondrocytes in DN-Cbfa1 transgenic mice retained the phenotype of permanent cartilage. These data demonstrate that Cbfa1 plays an important role not only in chondrocyte maturation in the process of endochondral ossification, but also in the specification of cartilage phenotype.
Materials and Methods
Generation of Transgenic Mice
DNA fragments covering the entire coding region of the mouse type I and II Cbfa1 isoforms (Harada et al. 1999) were cloned into the NotI site of a _Col2a1_-based expression vector, which contains promoter and enhancer of mouse Col2a1 gene (sequence data available from EMBL/GenBank/DDBJ under accession number M65161, 1940–2971 nt, and 4930–5571 nt, respectively) in pNASSβ (CLONTECH Laboratories, Inc.). The fragment of the Col2a1 first intron contains tissue-specific elements (Zhou et al. 1995; Krebsbach et al. 1996). Two forms of the DN-Cbfa1, which contain the runt domain only (395-bp NcoI-HindIII fragment) and the runt domain with NH2-terminal domain of type I Cbfa1 (673-bp BamHI-HindIII fragment; BamHI is a site in the cloning vector), were cloned into the NotI site of the _Col2a1_-based expression vector. The construct inserts, including β-galactosidase with Col2a1 promoter/enhancer, were injected into the pronuclei of fertilized eggs from F1 hybrid mice (C57BL/6 × C3H). Transgenic embryos were identified by Southern blot. Transgene expression was assessed by Northern blot as described previously (Komori et al. 1997), using RNA from the head or trunk of embryonic day 12.5–18.5 (E12.5–18.5) embryos and a 32P-labeled NcoI-HindIII fragment of Cbfa1 cDNA as a probe. Filters were rehybridized with a 32P-labeled 0.85-kb fragment of mouse glyceraldehyde-3-phosphate-dehydrogenase (GAPDH).
DNA Transfections and Luciferase Assays
Type I Cbfa1, type II Cbfa1, and DN-Cbfa1 (673-bp BamHI-HindIII fragment) were subcloned into the pSG5 vector (Stratagene) as described (Harada et al. 1999) and designated pSG5-type I Cbfa1, pSG5-type II Cbfa1, and pSG5–DN-Cbfa1, respectively. Insertless pSG5 vector (pSG5) was used as a control expression vector. A pGL3 promoter vector (Promega), containing six repeats of the consensus Cbfa1 binding sites, was used as an experimental reporter vector (p6XOSE2-Luc) (Harada et al. 1999). A pRL-SV40 vector (Promega) was used as a control reporter vector. Subconfluent cultures of chick sterna chondrocytes in a 6-well plate were transfected with 0.5 μg of p6XOSE2-Luc, 0.2 μg of pRL-SV40, and 3.3 μg of a mixture of pSG5 expression vectors with LipofectAMINE 2000 reagent (GIBCO BRL). After 48 h, cells were harvested and firefly luciferase activities generated by experimental reporter, and Renilla luciferase activity generated by control reporter, were simultaneously measured by Dual-Luciferase reporter assay system (Promega). The value of reporter gene activity was calculated by dividing firefly luciferase activity by Renilla luciferase activity and the result was expressed as relative luciferase activity. Transfection and reporter gene assay were carried out according to the manufacturer's protocol.
Skeletal, Histological, and Immunohistochemical Examinations
For staining and visualization of the whole skeleton, cleared skeletons of E15.5–18.5 embryos were stained with Alizarin red–S and Alcian blue as described (Komori et al. 1997). For light microscopy, tissues from E12.5–18.5 embryos were fixed in 4% paraformaldehyde/0.1 M phosphate buffer. Sections (7 μm in thickness) were stained with hematoxylin and eosin (H-E). Selected samples were stained using von Kossa's method to identify mineralized tissues (Komori et al. 1997). For immunohistochemical analysis, the sections were pretreated with 0.2% pepsin in 0.02 N HCl for 15 min at 37°C and then incubated with rabbit anti–mouse tenascin antibody (Kalembey et al. 1997) for 16 h at 4°C. Localization of the first antibody was visualized by incubation with biotinylated anti–rabbit IgG (Vector Laboratories) for 30 min at room temperature and then with Cy3-conjugated Streptavidin (Jackson ImmunoResearch Laboratories) for 30 min at room temperature.
Detection of β-Galactosidase Activity
For the detection of β-galactosidase activity, embryos were fixed with 4% paraformaldehyde in PBS (pH 7.4) at 4°C for 20 min. The samples were then rinsed in PBS, subsequently transferred into X-gal reaction mixture (1 mg/ml 4-chloro-5-bromo-3-indolyl-β, 4 mM K4Fe[CN]63H2O, 4 mM K3Fe[CN]6, 2 mM MgCl2 in PBS), and incubated at 30°C overnight. Stained embryos were embedded in paraffin and used to generate 7-μm sections, which were counterstained with eosin.
In Situ Hybridization
Digoxigenin 11 UTP-labeled single-stranded RNA probes were prepared using a DIG RNA labeling kit (Roche Biochemica) according to the manufacturer's instructions. A 0.4-kb fragment of mouse α1(II) collagen cDNA (Komori et al. 1997), a 0.65-kb fragment of mouse α1(X) collagen cDNA (Inada et al. 1999), a 0.8-kb fragment of mouse parathyroid hormone/parathyroid hormone–related peptide (PTH/PTHrP) receptor cDNA (Inada et al. 1999), a 0.6-kb fragment of mouse Indian hedgehog (Ihh) cDNA (Inada et al. 1999), and a 0.27-kb fragment of mouse growth and differentiation factor (GDF)-5 cDNA were used to generate antisense and sense probes. Hybridization was carried out as described (Inada et al. 1999).
Cell Cultures and Virus Infection
Chick sterna chondrocytes were isolated from 17-d-old embryo (line M) sternum, infected with the RCAS retroviral vector encoding type I Cbfa1, or the runt domain with NH2-terminal domain of type I Cbfa1, and cultured in high-glucose DME containing 10% FBS and 10 μg/ml of ascorbic acid as described previously (Enomoto-Iwamoto et al. 1998; Enomoto et al. 2000). On day 5, the cultures were replated in 24-well plates or 60-mm dishes at the density of 3 × 105 per well and 106 per dish, respectively. After 5 d the cultures in 60-mm dishes were subjected to immunoblot and Northern blot analyses (Inada et al. 1999). The cultures in 24-well plates were maintained for 8 d, then treated with 3 mM β-glycerophosphate for the additional 2 d and fixed for histochemical examination (Enomoto et al. 2000). The first antibody used was a polyclonal rabbit antibody against the NH2-terminal domain of type I Cbfa1, provided by Y. Ito (Kyoto University, Kyoto, Japan). Chick type II collagen, type X collagen, and tenascin cDNAs used for probes were described previously (Pacifici et al. 1993).
Results
Skeletal Abnormalities of Cbfa1 or DN-Cbfa1 Transgenic Mice
To generate mice that overexpressed Cbfa1, we used two Cbfa1 isoforms with different NH2 termini (Fig. 1, Table ). The first one, translated from exon 2, was originally named Pebp2αA (hereafter termed type I Cbfa1; Ogawa et al. 1993); the other, translated from exon 1, was called til-1 isoform (hereafter termed type II Cbfa1; Stewart et al. 1997). Mice overexpressing type I or II Cbfa1 died at birth from respiratory failure. At E18.5, they displayed severe dwarfism, with shortened limbs, domed skull, and a protruding tongue with a shortened snout and mandible (Fig. 1 C). Most skeletal elements, including the chondrocranium, ribs, and vertebrae, showed massive mineralization with severely reduced cartilaginous portions, and the thoracic cage was narrow and bell-shaped (Fig. 2B, Fig. C, Fig. F, Fig. G, Fig. J, and Fig. K). As to be expected, age-matched wild-type mice displayed cartilaginous tissues in the joints, ventral portion of ribs, and vertebral bodies (Fig. 2A, Fig. E, Fig. I, and Fig. M). In addition, most skeletal elements in the transgenic mice were fused, except manus and pedis (Fig. 2B and Fig. C). Shoulder, elbow, hip, and knee joints were absent, and the vertebral bodies and arches were fused (Fig. 2J, Fig. K, Fig. N, and Fig. O). The overall phenotypes of type I and II Cbfa1 transgenic mice were generally similar, although type II Cbfa1 transgenic mice seemed to develop these severe abnormalities slightly earlier than type I Cbaf1 transgenic mice (Fig. 3).
Figure 1.

Generation of transgenic mice. (A) Diagrams of the DNA constructs used to generate Col2a1-type I Cbfa1, Col2a1-type II Cbfa1, and Col2a1–DN-Cbfa1 transgenic mice. DNA fragments covering the entire coding region of the mouse type I or II Cbfa1 isoforms were inserted into a _Col2a1_-based expression vector, which contains promoter and enhancer of mouse Col2a1 gene. DNA fragments encoding the runt domain only or runt domain with NH2-terminal domain of type I Cbfa1 were also inserted into the _Col2a1_-based expression vector to generate DN forms of Cbfa1. Type I, type I Cbfa1; Type II, type II Cbfa1; DN, DN Cbfa1; Pr, Col2a1 promoter; En, Col2a1 enhancer; N, NotI; H, HindIII; E, EcoRI; B, BamHI. (B) Northern blot hybridized with Cbfa1 probe. RNA was extracted from the head or trunk of E12.5–18.5 embryos, and 20 μg of total RNA was loaded per lane. Representative data using RNA from heads of E12.5 embryos are shown. Lanes of wild-type (left), type I Cbfa1, and type II Cbfa1 transgenic mice are on one filter and lanes of wild-type (right, W) and DN-Cbfa1 transgenic mice (T) are on the other filter. Arrows indicate the bands of endogenous Cbfa1 and transgenes. Hybridization with the GAPDH probe was used as internal control for the loading of equal amounts of RNA. (C) Gross appearance of wild-type, type I Cbfa1, type II Cbfa1, and DN-Cbfa1 transgenic mice at E18.5. Both type I and II Cbfa1 transgenic mice exhibit dwarfism with domed skull, short snout and mandible, protruding tongue, and short limbs and tail. DN-Cbfa1 transgenic mice exhibit dwarfism with short limbs. In DN-Cbfa1 transgenic mice, all of the presented data in this paper are from mice that contain the runt domain with NH2-terminal domain of type I Cbfa1 except skeletal examinations, in which a mouse with runt domain only is presented. Bar, 2 mm.
Table 1.
Production Frequency of Transgenic Mice
| Transgene construct | Stage of killing | Embryos obtained | Transgenic mice | Transgenic mice with phenotype |
|---|---|---|---|---|
| Embryonic day | ||||
| Cbfa1 | 12.5 | 230 (111) | 37 (15) | 13 (7) |
| 13.5 | 133 (86) | 24 (19) | 12 (8) | |
| 14.5 | 170 (122) | 35 (25) | 24 (14) | |
| 15.5 | 250 (72) | 36 (15) | 18 (10) | |
| 16.5 | 99 (57) | 8 (6) | 5 (4) | |
| 18.5 | 231 (111) | 46 (14) | 20 (6) | |
| DN-Cbfa1 | 12.5 | 38 | 8 | 5 |
| 14.5 | 23 | 5 | 3 | |
| 15.5 | 9 | 3 | 2 | |
| 16.5 | 11 | 2 | 1 | |
| 18.5 | 121 | 21 | 10 |
Figure 2.
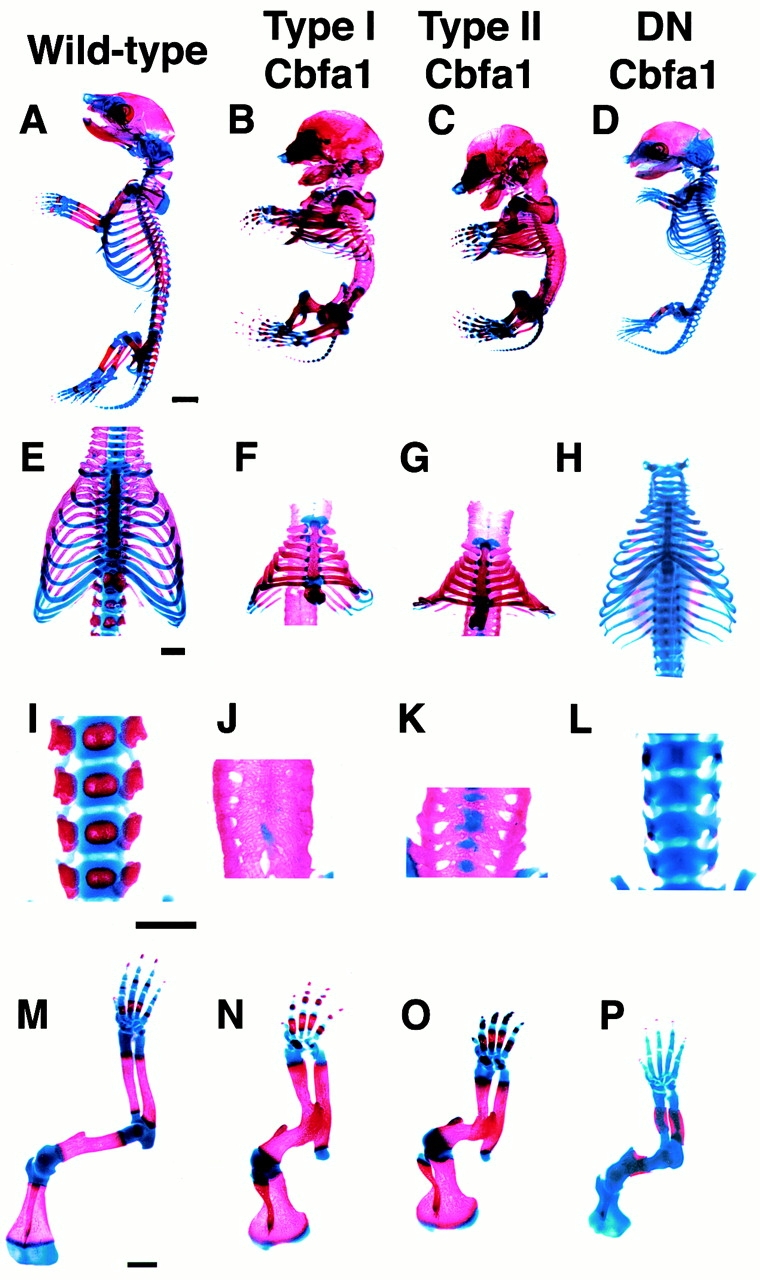
Examination of skeletal system. Skeletal examination of wild-type mice (A, E, I, and M) and type I Cbfa1 (B, F, J, and N), type II Cbfa1 (C, G, K, and O), and DN-Cbfa1 (D, H, L, and P) transgenic mice at E18.5. Calcified tissues are stained red with Alizarin red and the cartilage is stained blue with Alcian blue. Representative skeletons are shown. (A–D) Whole skeletons. In wild-type mice, cartilaginous tissues are observed in occipital bone, joints, the ventral portion of ribs, and vertebral bodies (A). In both type I and II Cbfa1 transgenic mice, most of the skeleton, including occipital bone, most of the ribs, and all of the vertebrae, is calcified (B and C). In DN-Cbfa1 transgenic mice, calcification is limited in flat bones of the head, mandible, clavicle, and long bones (D). (E–H) Thoracic cages. In wild-type mice, the ventral portion of all ribs is cartilaginous and sternum is segmentally calcified (E). In both type I and II Cbfa1 transgenic mice, the thoracic cage is small and bell-shaped, and the major portion of the ribs and sternum are calcified (F and G). In DN-Cbfa1 transgenic mice, the major portion of the ribs and the entire region of the sternum are cartilaginous (H). (I–L) Vertebral skeletons. In wild-type mice, centers of vertebral bodies and most vertebral arches are calcified (I). In both type I and II Cbfa1 transgenic mice, all of the vertebral bodies and arches are united and calcified (J and K). In DN-Cbfa1 transgenic mice, all of the vertebral bodies and arches are cartilaginous (L). (M–P) Forelimb skeletons. In wild-type mice, calcification is limited in diaphyses of long bones and the center of the scapula (M). In both type I and II Cbfa1 transgenic mice, the humerus is short and the humerus, radius, and ulna are thick and united (N and O). Shoulder joints and some carpal bones are also fused (N and O). Most of the elbow joint region is calcified and the calcified region is wider than that in wild-type mice in all long bones and scapula (N and O). In DN-Cbfa1 transgenic mice, calcification is limited around diaphyses of the humerus, radius, and ulna (P). Bars: (A–D) 2 mm; (E–P) 1 mm.
Figure 3.

Whole skeletons at E15.5. The skeletal system was also examined at E15.5 to compare type I (B) and type II Cbfa1 transgenic mice (C). (A) Wild-type mice at E15.5. Mineralization is advanced in both type I and II Cbfa1 transgenic mice in most skeletal components, including occipital bone, chondrocranium, ribs, vertebrae, and long bones, but the mineralization in type II Cbfa1 transgenic mice is more than that in type I Cbfa1 transgenic mice. The thoracic cage in type II Cbfa1 transgenic mice is smaller than that in type I Cbfa1 transgenic mice. Although fusion in joints and vertebrae is observed in both type I and II Cbfa1 transgenic mice, it is more advanced in type II Cbfa1 transgenic mice. Representative skeletons are shown. Bar, 1 mm.
To inactivate Cbfa1 function in chondrocytes, we generated additional lines of transgenic mice that express DN forms of Cbfa1 in chondrocytes under the control of the same Col2a1 promoter/enhancer elements used above (Fig. 1, Table ). We constructed cDNAs encoding the runt domain only or runt domain with NH2-terminal domain of type I Cbfa1. Both truncated forms of Cbfa1 had strong DN activity and were able to block the function of both type I and II Cbfa1 in reporter assays using osteoblast-specific elements and/or osteocalcin promoter fragments (Fig. 4; Ducy et al. 1999). In both Cbfa1 and DN-Cbfa1 transgenic mice, we did not observe apparent positive or negative regulation of endogenous Cbfa1 expression by the exogenous Cbfa1 and DN-Cbfa1 (Fig. 1 B).
Figure 4.

DN effect of the truncated form of Cbfa1, which contains the runt domain with NH2-terminal domain of type I Cbfa1. 0.5 μg of Cbfa1 target gene reporter plasmid (p6XOSE2-Luc), 0.2 μg of control reference reporter plasmid (pRL-SV40), and the indicated amounts of expression vectors were cotransfected into chick sterna chondrocytes. Relative luciferase activity was calculated as described in Materials and Methods. Values are means of four wells. Similar results were obtained in two independent experiments.
Transgenic mice overexpressing either form of DN-Cbfa1 also died soon after birth from respiratory failure and displayed dwarfism with shortened limbs, just like Cbfa1_-deficient mice (Fig. 1 C) (Komori et al. 1997; Otto et al. 1997). Staining of both types of DN-Cbfa1 transgenic mice at E18.5 with Alizarin red and Alcian blue revealed severely delayed calcification (Fig. 2D, Fig. H, Fig. L). Most skeletal elements, including ribs, occipital bone, base of skull, and vertebral bodies, were still uncalcified cartilaginous tissues, although a limited part of vertebral arches and diaphyses of limb long bones were calcified. As expected, most of the skull and clavicles, which are formed by intramembranous ossification, were clearly mineralized in the DN_-Cbfa1 transgenic mice, in contrast to the absence of calcification in the skulls of _Cbfa1_-deficient mice (Komori et al. 1997; Otto et al. 1997).
Overexpression of Cbfa1 Induced Precocious or Abnormal Endochondral Ossification
Developmental changes in cartilaginous skeletal elements were examined histologically. In E18.5 wild-type mice, nasal septum cartilage consisted of small homogenous chondrocytes (Fig. 5 A), which is typical of permanent cartilage, whereas that of Cbfa1 transgenic mice was mostly composed of hypertrophic chondrocytes and about half of it had already been replaced by bone marrow and osteoid tissues (Fig. 5 B). In wild-type mice at E18.5, the vertebral cartilage primordia just started to develop and consisted of homogenous small chondrocytes sandwiched between smaller mesenchymal cells destined to become intervertebral discs (Fig. 5 C). In Cbfa1 transgenic mice, vertebral cartilage had largely been replaced by bone, and chondrocyte maturation and bone formation were also observed in the presumed intervertebral regions (Fig. 5 D). In wild-type mice at E18.5, thyroid, cricoid, and tracheal cartilages, all of which have the permanent phenotype, consisted of small homogeneous chondrocytes (Fig. 5E and Fig. G), whereas those of Cbfa1 transgenic mice consisted of mixed populations of hypertrophic and small chondrocytes and some regions that were replaced by bone (Fig. 5F and Fig. H). Further, tracheal rings were fused in Cbfa1 transgenic mice (Fig. 5 H). These data indicate that cartilage tissues that normally develop to permanent cartilage proceeded to endochondral process in Cbfa1 transgenic mice. In the developing mandible, membranous ossification proceeds at some distance from Meckel's cartilage, the distal portion of Meckel's cartilage is replaced with fibrous tissues, and the caudal part gives rise to ossicles, such as malleus and incus, through endochondral ossification (Chung et al. 1995). However, the distal portion of Meckel's cartilage in Cbfa1 transgenic mice displayed endochondral ossification, whereas Meckel's cartilage in wild-type mice remained cartilaginous (Fig. 5, I–L). All of these findings show that chondrocyte maturation had been greatly enhanced in Cbfa1 transgenic mice, leading to abnormal behavior and development of most, if not all, cartilaginous structures.
Figure 5.
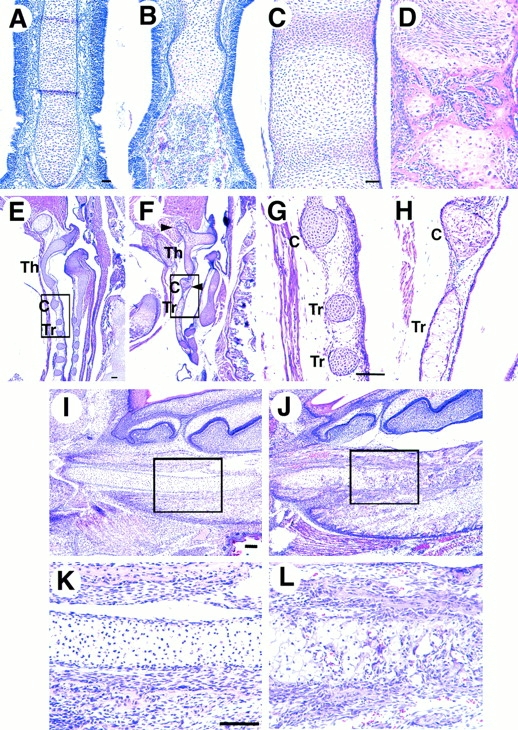
Endochondral ossification in cartilaginous tissues. (A and B) Frontal sections of the nasal septum from wild-type (A) and Cbfa1 transgenic (B) mice at E18.5. In Cbfa1 transgenic mice, most chondrocytes are hypertrophic and half of the nasal septum is replaced by bone (B). (C and D) Sagittal sections of the vertebral column from wild-type (C) and Cbfa1 transgenic (D) mice at E18.5. Most of vertebral bodies are replaced by bone, and chondrocytes in intervertebral regions are hypertrophic and partly replaced by bone in Cbfa1 transgenic mice (D). (E–H) Sagittal sections of the thyroid (Th), cricoid (C), and tracheal (Tr) cartilages from wild-type (E and G) and Cbfa1 transgenic (F and H) mice at E18.5. In Cbfa1 transgenic mice, tracheal rings are fused, large areas in thyroid, cricoid, and tracheal cartilages are occupied by hypertrophic chondrocytes, and some regions are replaced by bone (F and H). (G and H) Higher magnifications of boxed regions in E and F, respectively. Arrowheads in F show the regions that are replaced by bone. (I–L) Sections of the Meckel's cartilage from wild-type (I and K) and Cbfa1 transgenic (J and L) mice at E18.5. In wild-type mice, Meckel's cartilage is intact, but most of it is replaced by bone in Cbfa1 transgenic mice. (K and L) Higher magnifications of boxed regions in I and J, respectively. All sections were stained with H-E. Bars, 100 μm.
To establish more closely the differentiation state of chondrocytes in the transgenic mice, we focused on limb development. The Col2a1 promoter and enhancer fragments that we used were able to mimic the temporal and spatial expression of endogenous type II collagen, which is expressed in chondroprogenitor cells as well as chondrocytes, but very weakly in hypertrophic chondrocytes, as revealed by reporter activity (Fig. 6A and Fig. B; Ng et al. 1993; Zhou et al. 1995). At E12.5, when cartilage primordia begin to form in the forelimbs, we observed no significant differences in the overall size of the mesenchymal condensations in wild-type and Cbfa1 transgenic mice, but the primordia of humerus and radius and those of humerus and ulna appeared to have already fused in Cbfa1 transgenic mice (Fig. 6C and Fig. D). Thus, exogenous Cbfa1 expression in chondroprogenitor cells did not seem to affect their differentiation into chondrocytes (Fig. 6 D). In E15.5 wild-type mice, humerus, radius, and ulna were well developed; the diaphyses of these elements were undergoing replacement by bone, and cartilage tissue remained at the epiphyses and metaphyses organized in growth plates (Fig. 6 E). In these growth plates, type II collagen expression was gradually decreased toward the hypertrophic zone (Fig. 6 K) and type X collagen was conversely upregulated (Fig. 6 N). In Cbfa1 transgenic mice, these bones were fused and the growth plate was disorganized, as revealed by both the histological appearance and gene expression profile: cartilage tissues at the fused joint regions consisted of enlarged chondrocytes (Fig. 6F and Fig. I) expressing type X collagen (Fig. 6 O) and a small number of type II collagen–positive cells (Fig. 6 L). Thus, overexpression of Cbfa1 induced precocious and spatially abnormal expression of the hypertrophic phenotype. In E18.5 Cbfa1 transgenic mice, the fused forelimb skeletal elements displayed gross mineralization (Fig. 6 R), leading to the conclusion that activation of Cbfa1 signaling in immature chondrocytes promoted hypertrophy and precocious endochondral ossification.
Figure 6.
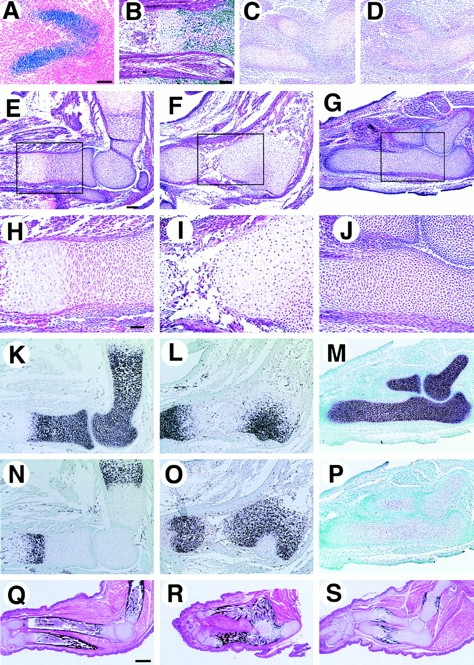
Process of endochondral ossification in limbs. The forelimb (A) and hind limb (B) from β_-_galactosidase transgenic mice and the forelimbs from wild-type (C, E, H, K, N, and Q), Cbfa1 transgenic (D, F, I, L, O, and R), and DN-Cbfal transgenic mice (G, J, M, P, and S) were examined histologically. (A and B) β-Galactosidase staining in E11.5 (A) and E16.5 (B) transgenic mice expressing β-galactosidase by Col2a1 promoter/enhancer. Staining is observed in chondroprogenitor cells in precartilaginous condensation (A). Staining is observed in chondrocytes but very weakly in hypertrophic chondrocytes and the staining is absent in osteoblasts (B). (C and D) H-E staining at E12.5. Humerus, radius, and ulna are uninterrupted in Cbfa1 transgenic mice (D). (E–J) H-E staining at E15.5. In Cbfa1 transgenic mice, humerus, radius, and ulna remain fused and the growth plate is disorganized and mainly composed of hypertrophic chondrocytes in the fused region (F and I). In DN-Cbfa1 transgenic mice, ulna remains cartilaginous and is composed of small immature chondrocytes and no vascular invasion is observed (G and J). (H–J) Higher magnifications of boxed regions in E–G. (K–P) In situ hybridization using type II collagen (K–M) and type X collagen (N–P) antisense probes at E15.5. In Cbfa1 transgenic mice, type II collagen expression is restricted in epiphyses and a small part of the fused region (L), but type X collagen is widely expressed (O). In contrast, in DN-Cbfa1 transgenic mice, type II collagen expression is observed in a whole part of humerus, radius, and ulna (M), and no type X collagen expression is detected (P). (Q–S) von Kossa and H-E double staining at E18.5. Bones of Cbfa1 transgenic mice are heavily mineralized (R), whereas mineralization in DN-Cbfa1 transgenic mice is restricted in bone collars and calcified cartilage in diaphysis of ulna (S). Bars: (A–P) 100 μm; (Q–S) 500 μm.
DN-Cbfa1 Expression Severely Inhibited Chondrocyte Maturation and Endochondral Ossification
In DN-Cbfa1 transgenic mice at E15.5, humerus, radius, and ulna remained entirely cartilaginous and were composed of immature chondrocytes expressing type II collagen, but not type X collagen (Fig. 6M). The formation of bone collars, which are formed by intramembranous ossification, was not observed (Fig. 6G and Fig. J) and it was confirmed by von Kossa's staining (data not shown). In E18.5 DN-Cbfa1 transgenic mice, bone collars of humerus, radius, and ulna appeared, but vascular invasion into the cartilage was rarely observed, indicating severe delay or blockage of endochondral ossification (Fig. 6 S). The inhibition of endochondral ossification in DN-Cbfa1 transgenic mice seemed mainly to originate from retarded maturation of chondrocytes, as indicated by the expression of type II collagen and lack of PTH/PTHrP receptor (PTHrPR), Ihh, and type X collagen in most of the diaphyses (Fig. 7). Transgenic mice overexpressing either form of DN-Cbfa1 showed similar phenotypes in all histological analyses (data not shown).
Figure 7.
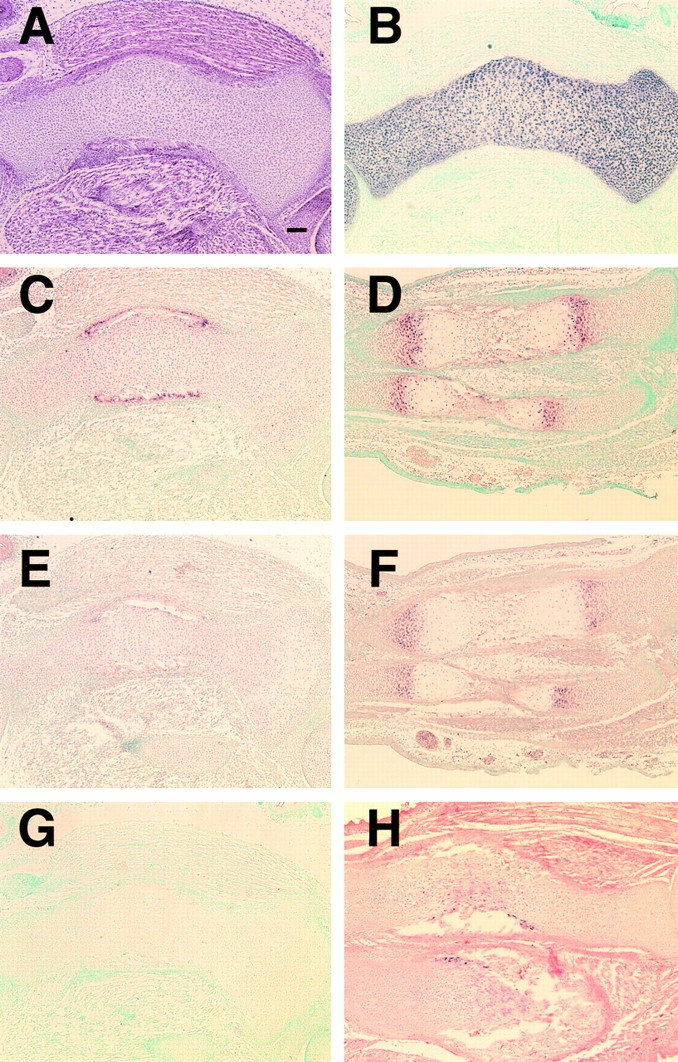
Chondrocyte maturation in DN-Cbfa1 transgenic mice. Maturational stage of chondrocytes in DN-Cbfa1 transgenic mice at E18.5 (A–C, E, G, and H) was examined with wild-type mice at E15.5 as positive control (D and F) by in situ hybridization using a type II collagen probe (B), PTH/PTHrPR probe (C and D), Ihh probe (E and F), and type X collagen probe (G and H). (A) H-E staining. In DN-Cbfa1 transgenic mice, a series of sections from the tibia is used in A–C, E, and G and a series of sections from the radius and ulna is used in H and Fig. 6 S. In wild-type mice, a series of sections from the tibia and fibula are used (D and F). In DN-Cbfa1 transgenic mice, most chondrocytes express type II collagen (B) but not PTH/PTHrPR (C), Ihh (E), or type X collagen (G). PTH/PTHrPR (D) and Ihh (F) are strongly expressed in prehypertrophic and early hypertrophic chondrocytes in wild-type mice. In DN-Cbfa1 transgenic mice, PTH/PTHrPR expression is restricted in osteoblastic cells in bone collar (C). A few type X collagen–positive cells are seen close to hematopoietic cells that invaded to inside the bone collar of radius and ulna in DN-Cbfa1 transgenic mice (H). Bar, 100 μm.
The phenotype induced by DN-Cbfa1 in chondrocytes was also studied in vitro. Chondrocytes were isolated from sterna cartilage of chick embryo and forced to express DN-Cbfa1 (Fig. 8 A). The cells retained their small size (Fig. 8 B, DN-Cbfa1), and failed to induce alkaline phosphatase activity (Fig. 8 C), matrix calcification (Fig. 8 C, Alizarin-red), and type X collagen expression (Fig. 8 D, Type X), whereas Cbfa1 induced a larger cell size and strongly stimulated expression of hypertrophic phenotype markers as reported previously (Fig. 8, B–D; Enomoto et al. 2000). The observations in Cbfa1 and DN-Cbfa1 transgenic mice, coupled with the in vitro results, clearly demonstrate that Cbfa1 plays a pivotal role in chondrocyte maturation in the process of endochondral ossification.
Figure 8.

Expression of Cbfa1 and DN-Cbfa1 in cultured chondrocytes. Chick sterna chondrocytes were infected with avian retrovirus RCAS (A) encoding type I Cbfa1 (Cbfa1) or the runt domain with NH2-terminal domain of type I Cbfa1 (DN-Cbfa1). Control cultures (Control) were infected with RCAS (A) virus encoding vector alone. (A) Immunoblot of the introduced gene products. (B) Photographs of cultures 5 d after virus infection. (C) Histochemical analysis of alkaline phosphatase activity (APase) and matrix calcification (Alizarin-red). (D) Northern blot of type II collagen (Type II), type X collagen (Type X), and tenascin (TN). Expression of DN-Cbfa1 retained the small cell size (B) and high levels of tenascin and type II collagen expression (D), whereas it inhibited the induction of alkaline phosphatase (C), matrix calcification (C), and type X collagen (D), indicating that DN-Cbfa1 disturbed chondrocyte maturation and maintained it in an early developmental state.
Activation of Cbfa1 Disturbed GDF-5 Expression and Inhibited Tenascin Expression in Joint-forming Regions, Whereas Inactivation of Cbfa1 Resulted in Broadly Retained Tenascin Expression in Cartilage Elements
Fusion of skeletal elements seen in Cbfa1 transgenic mice was accompanied with abnormal replacement of cartilage tissues by bone, suggesting that induction of endochondral ossification by Cbfa1 is a reason for their malformation. However, fusion of cartilaginous components was observed at much earlier times than when ossification began. To understand the mechanism of joint fusion seen in Cbfa1 transgenic mice, we examined the expression of GDF-5 and tenascin during limb development. GDF-5 is implicated in joint specification through its expression in prospective joints and its disruption in brachypodism mouse mutation (Storm et al. 1994; Storm and Kingsley 1996). GDF-5 expression was detected in shoulder but not in knee and elbow, and its appearance in digital rays was delayed in Cbfa1 transgenic mice (Fig. 9B, Fig. D, Fig. F, and Fig. G) compared with wild-type mice (Fig. 9A, Fig. C, and Fig. E). Therefore, Cbfa1 interrupted joint development in elbow and knee at an early stage, probably by affecting factors that determine future joint regions.
Figure 9.
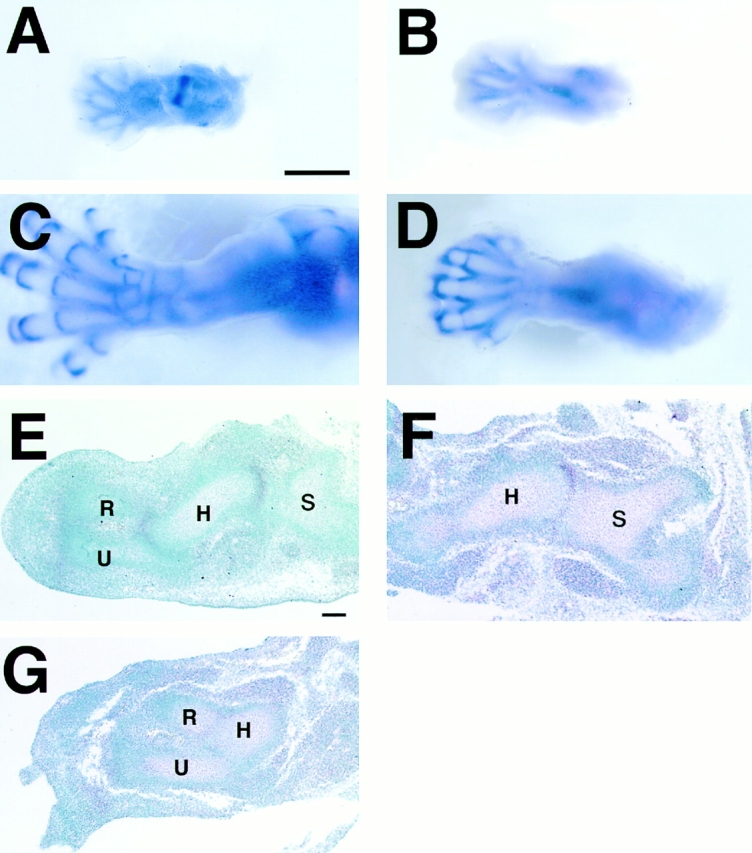
GDF-5 expression in developing limbs. (A–D) Whole-mount in situ hybridization of hind limbs from wild-type (A and C) and Cbfa1 transgenic (B and D) mice using antisense GDF-5 probe. In wild-type mice, GDF-5 expression was detected in knee and metatarsophalangeal joints at E12.5 (A). Additional stripes of expression are seen in interphalangeal joints and between the developing rows of tarsals at E13.5 (not shown) and E14.5 (C), whereas no GDF-5 expression was detected in the knee (B and D) and its expression in digital rays first appeared at E14.5 in Cbfa1 transgenic mice (D). A and B, E12.5; C and D, E14.5. In addition, GDF-5 expression in metatarsophalangeal joints is weak and no expressions between the developing tarsals are seen in Cbfa1 transgenic mice at E14.5 (D). (E–G) In situ hybridization of GDF-5 using sections from forelimbs of wild-type (E) and Cbfa1 transgenic (F and G) mice at E12.5. GDF-5 expression is clearly localized to the joint regions of the shoulder and elbow in wild-type mice (E). In Cbfa1 transgenic mice, GDF-5 expression is detected in the shoulder (F), but not in the elbow (G) joints. S, scapula; H, humerus; R, radius; U, ulna. Bars: (A–D) 1 mm; (E–G) 100 μm.
Tenascin is expressed in chondrocytes once cartilage tissue appears, but becomes limited to the articular chondrocytes as cartilage development progresses (Pacifici 1995). In wild-type mice at E15.5, several layers of chondrocytes at the edge of the epiphysis expressed tenascin, whereas chondrocytes in the presumptive joint region in Cbfa1 transgenic mice did not, suggesting that the cells had lost permanent phenotype (Fig. 10A, Fig. B, Fig. D, and Fig. E). In the shoulder of Cbfa1 transgenic mice, joint formation was observed at E12.5, but the joint was fused at E15.5 (Fig. 9 F and 10 H), indicating that Cbfa1 inhibited chondrocytes from acquiring the characteristics of articular cartilage. In contrast, many chondrocytes in DN-Cbfa1 transgenic mice expressed tenascin (Fig. 10C and Fig. F). Thus, most regions of developing cartilaginous elements in DN-Cbfa1 transgenic mice appeared to be at an early stage of cartilage development and retained the expression of a marker for permanent cartilage. The same regulation of tenascin gene expression by Cbfa1 was observed in vitro. Cbfa1 ceased tenascin expression, whereas dysfunction of Cbfa1 sustained it (Fig. 8 D).
Figure 10.
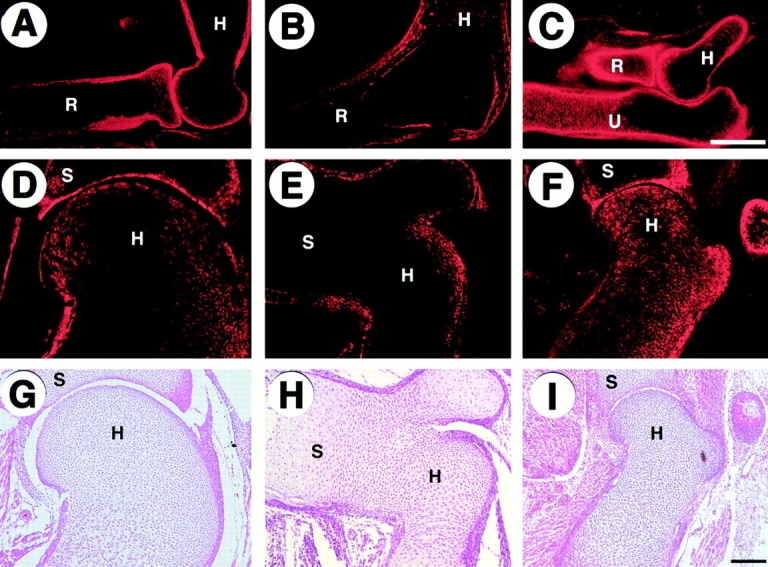
Tenascin expression in developing limbs. (A–F) Immunohistochemistry of elbow joint regions (A–C) and shoulder joint regions (D–F) from wild-type (A and D), Cbfa1 transgenic (B and E), and DN-Cbfa1 transgenic (C and F) mice at E15.5 using antitenascin antibody. Tenascin is strongly expressed along the edge of epiphysis and perichondrium and in bone of wild-type mice (A and D), but not in presumptive joint regions of Cbfa1 transgenic mice (B and E). In DN-Cbfa1 transgenic mice, tenascin is expressed more abundantly in the humerus, radius, and ulna (C and F). (G–I) H-E staining in the shoulder joint regions from wild-type (G), Cbfa1 transgenic (H), and DN-Cbfa1 transgenic (I) mice at E15.5. R, radius; U, ulna; H, humerus; S, scapula. Bars: (A–F) 300 μm; (G–I) 50 μm.
Discussion
Our results show that overexpression of Cbfa1 in growth plate chondrocytes accelerates maturation and endochondral ossification, whereas misexpression in presumptive permanent chondrocytes (such as articular chondrocytes) alters the developmental fate of the cells, resulting in the replacement of permanent cartilage by bone. Precocious or abnormal bone formation mediated by the endochondral ossification process is likely to be the cause of the malformations seen in Cbfa1 transgenic mice, including hypoplasia of the skull, shortening of long bones, fusion of vertebrae, contraction of the thoracic cage, and joint fusion. In contrast, interference with Cbfa1 function in cartilage suppresses chondrocyte maturation and matrix calcification and also delays specification of cell phenotype to articular cartilage, as determined by tenascin expression. DN-Cbfa1 transgenic mice also displayed skeletal deformities, mainly because of the loss or retardation of endochondral ossification. Cbfa1 expression is at a low level in chondrocytes at the beginning of cartilage development, is barely detectable in permanent cartilage, and is upregulated in hypertrophic chondrocytes in growth plate (Inada et al. 1999; Kim et al. 1999; our unpublished data). Therefore, it is likely that spatially and temporally regulated expression of Cbfa1 in chondrocytes is crucial for the control of skeletal formation throughout the body.
Our data clearly demonstrate that Cbfa1 plays a crucial role in chondrocyte maturation in the process of endochondral ossification and suggest that the retardation of chondrocyte maturation in _Cbfa1_-deficient mice was mainly caused by a lack of Cbfa1 in chondrocytes, not a lack of osteoblast differentiation or invasion of hematopoietic cells. However, the retardation of chondrocyte maturation observed in DN-Cbfa1 transgenic mice was severer than that seen in _Cbfa1_-deficient mice, although bone collars in diaphyses were generated and blood vessels invaded the bone collars in DN-Cbfa1 transgenic mice. In _Cbfa1_-deficient mice, the entire skeleton consisted of cartilage, most of which was composed of resting or proliferating chondrocytes, which lack the expression of PTH/PTHrPR, Ihh, and type X collagen (Inada et al. 1999; Kim et al. 1999). However, chondrocyte maturation was observed in the restricted skeleton, including tibia, fibula, radius, and ulna. Calcification occurred in these areas, and PTH/PTHrPR, Ihh, and type X collagen expression was observed in their prehypertrophic and hypertrophic chondrocytes (Inada et al. 1999; Kim et al. 1999). However, in DN-Cbfa1 transgenic mice hypertrophic chondrocytes were rare throughout the skeleton. Even in the tibia, fibula, radius, and ulna, only small numbers of cells expressed PTH/PTHrPR, Ihh, or type X collagen. As DN-Cbfa1 is expected to inhibit the expression of all runt family genes including Cbfa2/Runx1 and Cbfa3/Runx3 (Sakakura et al. 1994), runt family genes other than Cbfa1 might also be involved in chondrocyte maturation.
Ihh was considered to regulate chondrocyte differentiation via PTHrP and its receptor (Vortkamp et al. 1996; Zau et al. 1997). Consequently, Ihh suppresses the differentiation of prehypertrophic chondrocytes into hypertrophic chondrocytes. As chondrocyte maturation was arrested before the prehypertrophic stage in DN-Cbfa1 transgenic mice, the maturational disturbance cannot be explained by Ihh signaling. In DN-Cbfa1 transgenic mice, delayed bone collar formation is probably due to low Ihh expression in chondrocytes (Fig. 6G and Fig. J, and Fig. 7 E), whose level is less than the detection level of in situ hybridization, because _Ihh_-deficient mice displayed a complete lack of bone collar formation (St.-Jacques et al. 1999). Our data also indicate that osteoblast differentiation in the perichondrial region has no significant supportive effect on chondrocyte maturation at least in the absence of runt family factors in chondrocytes. Further evidence is provided by the finding that chondrocytes became hypertrophic in the absence of osteoblast differentiation in the perichondrial region of _Ihh_-deficient mice (St.-Jacques et al. 1999). However, vascular invasion seemed to support chondrocyte maturation, because a few type X collagen-positive cells appeared close to hematopoietic cells that had invaded to the inside of the bone collar in DN-Cbfa1 transgenic mice (Fig. 7 H).
Cbfa1 isoforms with different NH2 termini have been shown to have different transcriptional activities in genes of bone matrix proteins, including type I collagen and osteocalcin, in vitro (Tuji et al. 1998; Harada et al. 1999), although the functional difference remains to be clarified in vivo. We could not find functional differences between Cbfa1 isoforms in chondrocyte maturation, and both isoforms strongly stimulated chondrocyte maturation. However, type II Cbfa1 seemed to exert stronger activity in stimulating chondrocyte maturation than type I Cbfa1 (Fig. 3). This was also observed during in vitro experiments using chick primary chondrocytes (Enomoto et al. 2000). Differences in the function of the two Cbfa1 isoforms in chondrocytes need to be further investigated, especially in hypertrophic chondrocytes in which Cbfa1 and matrix protein genes related to calcification are highly expressed. Cbfa1 induced the differentiation of two lineage cells, osteoblasts and chondrocytes (Komori 2000). As the two Cbfa1 isoforms are expressed in chondrocytes as well as osteoblasts, isoform-dependent induction of each lineage differentiation is not valid. It has been shown that Ets1 works cooperatively with Cbfa1 in transactivation of the osteopontin gene (Sato et al. 1998). Although transcription factors that specifically interact with Cbfa1 in chondrocytes have not been identified, it is suggested that additional transcription factors required for cooperative work with Cbfa1 are different in chondrocyte and osteoblast differentiation.
The mechanism of specification of permanent cartilage is largely unknown. However, other investigators and we have reported that chondrocytes in permanent cartilage are able to express the terminal differentiation phenotype in vitro (Iwamoto et al. 1989; Pacifici et al. 1991; Stephens et al. 1992). In Cbfa1 transgenic mice, permanent cartilages such as articular cartilage, tracheal rings, nasal septum, and intervertebral disc underwent endochondral ossification. Taken together, it is likely that even permanent cartilage or cartilage fated to be permanent has the potential to be transient cartilage that enters endochondral pathway, and that lack of cell hypertrophy and matrix mineralization in permanent cartilage is due to negative regulation by microenvironmental cues and mechanisms, which could downregulate Cbfa1 expression. Degeneration of permanent cartilage is a feature of pathological change in osteoarthritis of the articular joints and hyperostosis of spine, such as ankylosis spondylitis or spondylosis of the spine (Sandell 1995). These diseases often displayed abnormal bone formation from cartilage, including subchondral bone sclerosis or osteophyte, associated with ectopic expression of several molecules such as type X collagen (von der Mark et al. 1992), osteopontin (Pullig et al. 2000), and matrix metalloproteinase 13 (MMP13) (Shlopov et al. 1997), which are normally specific to hypertrophic chondrocytes. Therefore, it has been proposed that the ossifying events in these diseases might be related to uncontrolled behavior of permanent chondrocytes and abnormal expression of hypertrophic phenotype. Ectopic ossification and ectopic expression of hypertrophic chondrocyte markers are characteristics of _Cbfa1_-transgenic mice. Further, Cbfa1 is able to induce the expression of osteopontin and MMP13 (Ducy et al. 1997; Komori et al. 1997; Shlopov et al. 1997; Sato et al. 1998; Harada et al. 1999; Inada et al. 1999; Jimenez et al. 1999; Porte et al. 1999). These findings suggest that Cbfa1 activation might be involved in permanent cartilage disorders. The mechanisms regulating Cbfa1 expression in chondrocytes are not yet fully clear and such information would provide important clues into cartilage development, including permanent cartilage organization, as well as the pathogenesis of osteoarthritis and spinal disorder.
Acknowledgments
We thank M. Pacifici and M. Iwasaki for critically reading this manuscript, H. Harada for Cbfa1 cDNA, K. Imanaka-Yoshida for antitenascin antibody, R. Hiraiwa for maintaining mouse colonies, and A. Kimura for secretarial assistance.
This work was supported by grants from the Ministry of Education, Science, and Culture, Japan, and the Tokyo Biochemical Research Foundation.
Footnotes
Abbreviations used in this paper: Cbfa1, core binding factor α1; Col2a1, α1 type II collagen; DN, dominant negative; GAPDH, glyceraldehyde-3-phosphate-dehydrogenase; GDF, growth and differentiation factor; H-E, hematoxylin and eosin; Ihh, Indian hedgehog; PTH, parathyroid hormone; PTHrP, PTH-related peptide; PTHrPR, PTHrP receptor.
References
- Chung K.S., Park H.H., Ting K., Takita H., Apte S.S., Kuboki Y., Nishimura I. Modulated expression of type X collagen in Meckel's cartilage with different developmental fates. Dev. Biol. 1995;170:387–396. doi: 10.1006/dbio.1995.1224. [DOI] [PubMed] [Google Scholar]
- Ducy P., Zhang R., Geoffroy V., Ridall A.L., Karsenty G. Osf2/Cbfa1a transcriptional activator of osteoblast differentiation. Cell. 1997;89:747–754. doi: 10.1016/s0092-8674(00)80257-3. [DOI] [PubMed] [Google Scholar]
- Ducy P., Starbuck M., Priemel M., Shen J., Pinero G., Geoffroy V., Amling M., Karsenty G. A Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development. Genes Dev. 1999;13:1025–1036. doi: 10.1101/gad.13.8.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enomoto H., Enomoto-Iwamoto M., Iwamoto M., Nomura S., Himeno M., Kitamura Y., Kishimoto T., Komori T. Cbfa1 is a positive regulatory factor in chondrocyte maturation. J. Biol. Chem. 2000;275:8695–8702. doi: 10.1074/jbc.275.12.8695. [DOI] [PubMed] [Google Scholar]
- Enomoto-Iwamoto M., Iwamoto M., Mukudai Y., Kawakami Y., Nohno T., Higuchi Y., Takemoto S., Ohuchi H., Noji S., Kurisu K. Bone morphogenetic protein signaling is required for maintenance of differentiated phenotype, control of proliferation, and hypertrophy in chondrocytes. J. Cell Biol. 1998;140:409–418. doi: 10.1083/jcb.140.2.409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert S.F. Developmental Biology 5th ed 1997. Sinauer Associates, Inc; Sunderland, MA: pp. 351–358 [Google Scholar]
- Harada H., Tagashira S., Fujiwara M., Ogawa S., Katsumata T., Yamaguchi A., Komori T., Nakatsuka M. Cbfa1 isoforms exert functional differences in osteoblast differentiation. J. Biol. Chem. 1999;274:6972–6978. doi: 10.1074/jbc.274.11.6972. [DOI] [PubMed] [Google Scholar]
- Inada M., Yasui T., Nomura S., Miyake S., Deguchi K., Himeno M., Sato M., Yamagiwa H., Kimura T., Yasui N. Maturational disturbance of chondrocytes in Cbfa1-deficient mice. Dev. Dyn. 1999;214:279–290. doi: 10.1002/(SICI)1097-0177(199904)214:4<279::AID-AJA1>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- Iwamoto M., Sato K., Nakashima K., Shimazu A., Kato Y. Hypertrophy and calcification of rabbit permanent chondrocytes in pelleted culturessynthesis of alkaline phosphatase and 1,25-dihydroxycholecalciferol receptor. Dev. Biol. 1989;136:500–507. doi: 10.1016/0012-1606(89)90275-3. [DOI] [PubMed] [Google Scholar]
- Jimenez M.J., Balbin M., Lopez J.M., Alvarez J., Komori T., Lopez-Otin C. Collagenase 3 is a target of Cbfa1, a transcription factor of the runt gene family involved in bone formation. Mol. Cell. Biol. 1999;19:4431–4442. doi: 10.1128/mcb.19.6.4431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalembey I., Yoshida T., Iriyama K., Sakakura T. Analysis of tenascin mRNA expression in the murine mammary gland from embryogenesis to carcinogenesisan in situ hybridization study. Int. J. Dev. Biol. 1997;41:569–573. [PubMed] [Google Scholar]
- Kim I.S., Otto F., Zabel B., Mundlos S. Regulation of chondrocyte differentiation by Cbfa1. Mech. Dev. 1999;80:159–170. doi: 10.1016/s0925-4773(98)00210-x. [DOI] [PubMed] [Google Scholar]
- Komori T. A fundamental transcription factor for bone and cartilage. Biochem. Biophys. Res. Commun. 2000;276:813–816. doi: 10.1006/bbrc.2000.3460. [DOI] [PubMed] [Google Scholar]
- Komori T., Yagi H., Nomura S., Yamaguchi A., Sasaki K., Deguchi K., Shimizu Y., Bronson R.T., Gao Y.H., Inada M. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell. 1997;89:755–764. doi: 10.1016/s0092-8674(00)80258-5. [DOI] [PubMed] [Google Scholar]
- Krebsbach P.H., Nakata K., Bernier S.M., Hatano O., Miyashita T., Rhodes C.S., Yamada Y. Identification of a minimum enhancer sequence for the type II collagen gene reveals several core sequence motifs in common with the link protein gene. J. Biol. Chem. 1996;271:4298–4303. doi: 10.1074/jbc.271.8.4298. [DOI] [PubMed] [Google Scholar]
- Ng L.J., Tam P.P., Cheah K.S. Preferential expression of alternatively spliced mRNAs encoding type II procollagen with a cysteine-rich amino-propeptide in differentiating cartilage and nonchondrogenic tissues during early mouse development. Dev. Biol. 1993;159:403–417. doi: 10.1006/dbio.1993.1251. [DOI] [PubMed] [Google Scholar]
- Ogawa E., Maruyama M., Kagoshima H., Inuzuka M., Lu J., Satake M., Shigesada K., Ito Y. PEBP2/PEA2 represents a family of transcription factors homologous to the products of the Drosophila runt gene and the human AML1 gene. Proc. Natl. Acad. Sci. USA. 1993;90:6859–6863. doi: 10.1073/pnas.90.14.6859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otto F., Thornell A.P., Crompton T., Denzel A., Gilmour K.C., Rosewell I.R., Stamp G.W., Beddington R.S., Mundlos S., Olsen B.R. Cbfa1, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell. 1997;89:765–771. doi: 10.1016/s0092-8674(00)80259-7. [DOI] [PubMed] [Google Scholar]
- Pacifici M. Tenascin-C and the development of articular cartilage. Matrix Biol. 1995;14:689–698. doi: 10.1016/s0945-053x(05)80011-3. [DOI] [PubMed] [Google Scholar]
- Pacifici M., Golden E.B., Adams S.L., Shapiro I.M. Cell hypertrophy and type X collagen synthesis in cultured articular chondrocytes. Exp. Cell Res. 1991;192:266–270. doi: 10.1016/0014-4827(91)90185-w. [DOI] [PubMed] [Google Scholar]
- Pacifici M., Iwamoto M., Golden E.B., Leatherman J.L., Lee Y.S., Chuong C.M. Tenascin is associated with articular cartilage development. Dev. Dyn. 1993;198:123–134. doi: 10.1002/aja.1001980206. [DOI] [PubMed] [Google Scholar]
- Porte D., Tuckermann J., Becker M., Baumann B., Teurich S., Higgins T., Owen M.J., Schorpp-Kistner M.P., Angel P. Both AP-1 and Cbfa1-like factors are required for the induction of interstitial collagenase by parathyroid hormone. Oncogene. 1999;18:667–678. doi: 10.1038/sj.onc.1202333. [DOI] [PubMed] [Google Scholar]
- Pullig O., Weseloh G., Gauer S., Swoboda B. Osteopontin is expressed by adult human osteoarthritic chondrocytesprotein and mRNA analysis of normal and osteoarthritic cartilage. Matrix Biol. 2000;19:245–255. doi: 10.1016/s0945-053x(00)00068-8. [DOI] [PubMed] [Google Scholar]
- Sakakura C., Yamaguchi-Iwai Y., Satake M., Bae S.C., Takahashi A., Ogawa E., Hagiwara A., Takahashi T., Murakami A., Makino K. Growth inhibition and induction of differentiation of t(8;21) acute myeloid leukemia cells by the DNA-binding domain of PEBP2 and the AML1/MTG8(ETO)-specific antisense oligonucleotide. Proc. Natl. Acad. Sci. USA. 1994;91:11723–11727. doi: 10.1073/pnas.91.24.11723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandell L.J. Molecular Biology of Collagens in Normal and Osteoarthritic Cartilage. In: Kuettner K.E., Goldberg V.M., editors. Osteoarthritic Disorders. The Academy; Rosemont, IL: 1995. pp. 131–146. [Google Scholar]
- Sato M., Morii E., Komori T., Kawahata H., Sugimoto M., Terai K., Shimizu H., Yasui T., Ogihara H., Yasui N. Transcriptional regulation of osteopontin gene in vivo by PEBP2alphaA/CBFA1 and ETS1 in the skeletal tissues. Oncogene. 1998;17:1517–1525. doi: 10.1038/sj.onc.1202064. [DOI] [PubMed] [Google Scholar]
- Shlopov B.V., Lie W.R., Mainardi C.L., Cole A.A., Chubinskaya S., Hasty K.A. Osteoarthritic lesionsinvolvement of three different collagenases. Arthritis Rheum. 1997;40:2065–2074. doi: 10.1002/art.1780401120. [DOI] [PubMed] [Google Scholar]
- Stephens M., Kwan A.P., Bayliss M.T., Archer C.W. Human articular surface chondrocytes initiate alkaline phosphatase and type X collagen synthesis in suspension culture. J. Cell Sci. 1992;103:1111–1116. doi: 10.1242/jcs.103.4.1111. [DOI] [PubMed] [Google Scholar]
- Stewart M., Terry A., Hu M., O'Hara M., Blyth K., Baxter E., Cameron E., Onions D.E., Neil J.C. Proviral insertions induce the expression of bone-specific isoforms of PEBP2alphaA (CBFA1)evidence for a new myc collaborating oncogene. Proc. Natl. Acad. Sci. USA. 1997;94:8646–8651. doi: 10.1073/pnas.94.16.8646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- St.-Jacques B., Hammerschmidt M., McMahon A.P. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999;13:2072–2086. doi: 10.1101/gad.13.16.2072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Storm E.E., Kingsley D.M. Joint patterning defects caused by single and double mutations in members of the bone morphogenetic protein (BMP) family. Development. 1996;122:3969–3979. doi: 10.1242/dev.122.12.3969. [DOI] [PubMed] [Google Scholar]
- Storm E.E., Huynh T.V., Copeland N.G., Jenkins N.A., Kingsley D.M., Lee S.J. Limb alterations in brachypodism mice due to mutations in a new member of the TGF beta-superfamily. Nature. 1994;368:639–643. doi: 10.1038/368639a0. [DOI] [PubMed] [Google Scholar]
- Tuji K., Ito Y., Noda M. Expression of the PEBP2αA/AML3/CBFA1 gene is regulated by BMP4/7 heterodimer and its overexpression suppresses type I collagen and osteocalcin gene expression in osteoblastic and nonosteoblastic mesenchymal cells. Bone. 1998;22:87–92. doi: 10.1016/s8756-3282(97)00267-6. [DOI] [PubMed] [Google Scholar]
- von der Mark K., Kirsch T., Nerlich A., Kuss A., Weseloh G., Gluckert K., Stoss H. Type X collagen synthesis in human osteoarthritic cartilage. Indication of chondrocyte hypertrophy. Arthritis Rheum. 1992;35:806–811. doi: 10.1002/art.1780350715. [DOI] [PubMed] [Google Scholar]
- Vortkamp A., Lee K., Lanske B., Segre G.V., Kronenberg H.M., Tabin C.J. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein. Science. 1996;273:613–622. doi: 10.1126/science.273.5275.613. [DOI] [PubMed] [Google Scholar]
- Zau H., Wieser R., Massagué J., Niswander L. Distinct roles of type I bone morphogenetic protein receptors in the formation and differentiation of cartilage. Genes Dev. 1997;11:2191–2203. doi: 10.1101/gad.11.17.2191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou G., Garofalo S., Mukhopadhyay K., Lefebvre V., Smith C.N., Eberspaecher H., de Crombrugghe B. A 182 bp fragment of the mouse pro alpha 1(II) collagen gene is sufficient to direct chondrocyte expression in transgenic mice. J. Cell Sci. 1995;108:3677–3684. doi: 10.1242/jcs.108.12.3677. [DOI] [PubMed] [Google Scholar]