Гибрид (биология) | это... Что такое Гибрид (биология)? (original) (raw)

Гибри́д (от лат. hibrida, hybrida — помесь) — организм (клетка), полученный вследствие скрещивания генетически различающихся форм. Понятие гибрид особенно распространено в ботанике, но применяется и в зоологии.
В промышленном и любительском цветоводстве также используется термин грекс (англ. grex). Введен Карлом Линнеем для использования биноминальной номенклатуры в классификации искусственных гибридов.
Гибриды могут быть внутривидовыми (при скрещивании различных сортов, форм, разновидностей), внутриродовыми (при скрещивании видов принадлежащих одному роду) или межродовыми (при скрещивании видов относящихся к разным родам).
В цветоводстве, гибриды первого поколения называются первичными гибридами.
Возможность искусственного получения гибридов впервые предположил немецкий учёный Р. Камерариус в 1694 году, впервые искусственную гибридизацию осуществил английский садовод Т. Фэрчайлд, скрестив в 1717 году разные виды гвоздик.
Содержание
- 1 Реципрокные гибриды
- 2 Межвидовая и межродовая гибридизация
- 3 Гибриды в научной номенклатуре
- 4 Стерильность гибридов
- 5 Разрушение гибридов
- 6 Гибриды, имеющие собственные названия
- 7 См. также
- 8 Примечания
Реципрокные гибриды
Реципрокные гибриды появляются в результате реципрокных скрещиваний — гибридизация включающая перемену пола родителей, связанных с каждым генотипом.
Реципрокные эффекты
Различия между реципрокными гибридами — реципрокные эффекты — свидетельствуют о неодинаковом вкладе мужского и женского пола в генотип потомства. Если бы потомки от отца и матери получали одинаковую генетическую информацию, то не должно было быть никаких реципрокных эффектов.
Измерение реципрокных эффектов
Для измерения реципрокных эффектов (r) можно использовать выражение:
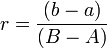
где A и B — значения признака для исходных скрещиваемых форм; a — то же самое для гибрида ♂A x ♀B; b — для реципрокного гибрида ♂B x ♀A. Положительное значение r (r > 0) будет означать «отцовский» эффект, отрицательное (r < 0) — «материнский», а абсолютная величина r (│r│) даст относительную оценку этих эффектов в единицах, равных разности значения признака для исходных форм (B — A).
Реципрокные эффекты у птиц
У кур «отцовский» эффект " наблюдался по наследованию инстинкта насиживания (r = 0.45,[1] 0.38[2] и 0.50[3]), половой скороспелости (r = 0.59[4]), яйценоскости (r = 0.32, −2.8, 1.07, 0.11, 0.46,[4] 1.14[5] и 2.71[6]), и живому весу (r = 0.30).[6]
По весу яиц наблюдался «материнский эффект» (r = −1.0).[6]
Реципрокные эффекты у млекопитающих
У свиней «отцовский» эффект " наблюдается по числу позвонков (отбор на длинное туловище) (r = 0.72[7] и 0.74[8]), длине тонкого кишечника (отбор на лучшую оплату корма), и динамике роста (отбор на скороспелость) (r = 1.8).
«Материнский эффект» наблюдался по среднему весу эмбрионов, пищеварительной системы и её частей, длине толстого кишечника и весу новорожденных поросят.[8]
У крупного рогатого скота «отцовский» эффект " наблюдался по удою молока (r = 0.07, 0.39, 0.23) и продукции молочного жира (количество жира) (r = 1.08, 1.79, 0.34).
«Материнский эффект» наблюдался по проценту жира в молоке у коров (r = −0.13, −0.19, −0.05).[5]
Теории реципрокных эффектов
«Материнский эффект»
Материнский эффект может быть обусловлен цитоплазматической наследственностью, гомогаметной конституцией и утробным развитием у млекопитающих. Различают собственно материнский эффект, когда генотип матери проявляется в фенотипе потомства. Молекулы в яйцеклетке, такие как мРНК, могут влиять на ранние стадии процесса развития. Различают также материнское наследование, при котором часть генотипа потомство получает исключительно от матери, например митохондрии и пластиды, содержащие свой собственный геном. При материнском наследовании фенотип потомства отражает его собственный генотип.
«Отцовский эффект»
Большее влияние отца на яйценоскость дочерей у кур объясняли тем, что у птиц гетерогаметным полом является самка, а гомогаметным — самец. Поэтому свою единственную X-хромосому курица получает от отца, и если яйценоскость определяется ею, то тогда все понятно.[2] Эта трактовка может объяснить хромосомный механизм явления у птиц, но для млекопитающих уже неприменима. Удивительно также то, что признаки, проявляющиеся только у женского пола (инстинкт насиживания, скороспелость и яйценоскость у курицы или удой молока и количество молочного жира у коровы), которые, казалось бы, должны передаваться матерью, тем не менее передаются больше отцом.
Межвидовая и межродовая гибридизация
Межвидовая гибридизация часто наблюдается как в природе, так и при культивировании человеком (содержании в неволе) у множества видов растений и животных. В природе в районах соприкосновения близких видов могут формироваться так называемые «гибридные зоны», где гибриды численно преобладают над родительскими формами.
Межвидовая интрогрессивная гибридизация широко распространена у дафний. В некоторых летних популяциях дафний гибриды преобладают, что затрудняет определение границ видов [1]/
Хонорик — выведенный путем селекции гибрид между тремя родительскими видами рода Mustela. Самцы хонориков стерильны, а самки фертильны.
Известный экспериментальный гибрид рафанобрассика (лат. Raphano-brassica) был получен Г. Д. Карпеченко при скрещивании редьки с капустой. Оба вида принадлежат к разным родам и имеют по 18 хромосом. Гибрид, полученный в результате удвоения числа хромосом (36), был способен к размножению, так как в процессе мейоза хромосомы редьки и капусты коньюгировали с себе подобными. Он обладал некоторыми признаками каждого из родителей и сохранял их в чистоте при размножении.[9]
Межродовые гибриды (как естественные, так и полученные селекционерами) известны также в семействах злаков, розоцветных, цитрусовых [2], орхидных и др. Так, гексаплоидный геном мягких пшениц образовался путем объединения диплоидных геномов двух предковых видов пшениц и одного вида близкого рода Aegilops.
Гибриды в научной номенклатуре
В ботанике
Гибридные таксоны растений называются нототаксонами.
- Гибридность указывается посредством знака умножения «×» или добавления префикса «notho-» к термину, обозначающему ранг таксона.
- Гибридность между таксонами обозначается посредством знака «×», помещённого между названиями этих таксонов. Названия в формуле предпочтительнее располагать в алфавитном порядке. Направление скрещивания может указываться посредством символических знаков пола (♂ и ♀).
Пример: Phalaenopsis amabilis (L.) Blume × Phalaenopsis aphrodite Rchb.f. - Гибридам между представителями двух и большего числа таксонов могут быть даны названия. В этом случае знак «×» помещается перед названием межродового гибрида или перед эпитетом в названии межвидового гибрида. Примеры:
- Cattleya ×hardyana
- × Ascocenda
- Нототаксон не может быть обозначен, если неизвестен по крайней мере один из его родительских таксонов.
- Если вместо знака «×» по каким-то причинам употребляется буква «х», то между этой буквой и эпитетом может быть сделан один буквенный пробел, если это поможет избежать неясности. Буква «х» должна быть строчной.
- Нотородовое название гибрида между двумя и более родами является либо сжатой формулой, в которой названия, принятые для родительских родов, комбинируются в одно слово, либо образовано от имени исследователя или садовода, занимавшегося этой группой. Примеры:
- × Rhynchosophrocattleya (= Rhyncholaelia × Sophronitis × Cattleya)
- × Vuylstekeara (= Cochlioda × Miltonia × Odontoglossum). Род зарегистрирован в 1911 году известным бельгийским коллекционером и селекционером орхидных Charles Vuylsteke (1844—1927).
- Таксоны, считающиеся гибридными по происхождению, не требуется обозначать как нототаксоны. Примеры:
- Гомозиготный гибридный тетраплоид наперстянки Digitalis ×mertonensis, полученный от искусственного скрещивания Digitalis grandiflora × Digitalis purpurea, может приводиться при желании, как Digitalis mertonensis.
- Rosa canina, полиплоид, который, как полагают, имеет древнее гибридное происхождение, рассматривается как вид[10].
По данным AOS начиная с января-марта 2008 года между знаком × и названием гибридного рода должен быть пробел[11].
Пример: × Rhynchosophrocattleya.
В зоологии
Стерильность гибридов
Явления стерильности гибридов неоднородны. Наблюдается изменчивость в отношении того, на какой именно стадии проявляется стерильность и каковы её генетические причины.
Нарушение сперматогенеза на стадиях, предшествующих мейозу, — непосредственная причина стерильности у самцов мула; нарушения мейоза — причина стерильности у гибридных самцов при некоторых скрещиваниях между разными видами Drosophila (например, D. pseudoobscura × D. persimilis).
К ограниченной полом стерильности и нежизнеспособности гибридов у раздельнополых животных приложимо обобщение, известное под названием правила Холдейна. Гибриды от межвидовых скрещиваний у раздельнополых животных должны состоять, во всяком случае потенциально, из гетерогаметного пола (несущего хромосомы XY) и гомогаметного (XX) пола. Правило Холдейна гласит, что в тех случаях, когда в проявлении стерильности или нежизнеспособности гибридов существуют половые различия, они наблюдаются чаще у гетерогаметного, чем у гомогаметного пола. У большинства животных, в том числе у млекопитающих и у двукрылых, гетерогаметны самцы. Из правила Холдейна имеются, однако, многочисленные исключения.
Третья стадия развития, на которой может проявляться гибридная стерильность, — это гаметофитное поколение у растений. У цветковых растений из продуктов мейоза непосредственно развиваются гаметофиты — пыльцевые зерна и зародышевые мешки, — которые содержат от двух до нескольких ядер и в которых формируются гаметы. Нежизнеспособность гаметофитов — обычная причина стерильности гибридов у цветковых растений. Мейоз завершается, но нормального развития пыльцы и зародышевых мешков не происходит.
Гибридная стерильность на генетическом уровне может быть обусловлена генными, хромосомными и цитоплазматическими причинами[12].Наиболее широко распространена и обычна генная стерильность. Неблагоприятные сочетания генов родительских типов, принадлежащих к разным видам, могут приводить и действительно приводят к цитологическим отклонениям и нарушениям развития у гибридов, что препятствует образованию гамет. Генетический анализ генной стерильности у гибридов Drosophila (D. pseudoobscura × D. persimilis, D. tnelanogaster × D. simulans и т. п.) показывает, что гены, обусловливающие стерильность, локализованы во всех или почти во всех хромосомах родительского вида [13][14].
Неблагоприятные взаимодействия между цитоплазматическими и ядерными генами также ведут к стерильности межвидовых гибридов в разных группах растений и животных[15].
Виды растений и животных часто различаются по транслокациям, инверсиям и другим перестройкам, которые в гетерозиготном состоянии вызывают полустерильность или стерильность. Степень стерильности пропорциональна числу независимых перестроек: так гетерозиготность по одной транслокации даёт 50%-ную стерильность, по двум независимым транслокациям — 75%-ную стерильность и т. д. Стерильность растений определяется гаметофитом. У гетерозигот по хромосомным перестройкам в результате мейоза образуются дочерние ядра, несущие нехватки и дупликации по определённым участкам; из таких ядер не получается функциональных пыльцевых зёрен и семязачатков. Хромосомная стерильность подобного типа очень часто встречается у межвидовых гибридов цветковых растений.
Течение мейоза у гибрида может быть нарушено либо генными факторами, либо различиями в строении хромосом. Как генная, так и хромосомная стерильность может выражаться в аберрантном течении мейоза. Но типы мейотических аберраций различны. Генная стерильность обычна у гибридов животных, а хромосомная стерильность — у гибридов растений. Генетический анализ некоторых межвидовых гибридов растений показывает, что нередко у одного гибрида наблюдается одновременно и хромосомная, и генная стерильность[12].
Разрушение гибридов
В случаях, когда некий межвидовой гибрид достаточно жизнеспособен и способен к размножению, поколения его потомков будут содержать значительную долю нежизнеспособных, субвитальных, стерильных и полустерильных особей. Эти типы представляют собой неудачные продукты рекомбинации, возникшие при межвидовой гибридизации. Такое подавление мощности и плодовитости в гибридном потомстве называют разрушением гибридов (hybrid breakdown). Разрушение гибридов — последнее звено в последовательности преград, препятствующих межвидовому обмену генами.
Разрушение гибридов неизменно обнаруживается в потомстве межвидовых гибридов у растений, где его легче наблюдать, чем при большинстве скрещиваний у животных[12].
Гибриды, имеющие собственные названия
Реципрокные гибриды
- Мул — гибрид от скрещивания осла и лошади
- Лошак — гибрид от скрещивания жеребца и ослицы
- Лигр — гибрид от скрещивания льва (Panthera leo) и тигрицы (Panthera tigris)
- Тигон — гибрид от скрещивания тигра и львицы
- Леопон — гибрид леопарда-самца и львицы-самки
- Нар — гибрид одногорбого и двугорбого верблюдов.
- Муллард — гибрид, получаемый при скрещивании селезней мускусных уток с утками породы пекинская белая, оргпингтон, руанская и белая алье.
- Хайнак (Дзо) — гибрид яка и коровы.
- Зеброид — гибрид от скрещивания зебры и домашней лошади
- Зебрул — гибрид от скрещивания зебры и осла
Гибриды в семействе Орхидные
Многие виды одного рода и даже представители различных родов легко скрещиваются между собой, образуя многочисленные гибриды, способные к дальнейшему размножению. Большинство гибридов, появившихся за последние 100 лет, создано искусственно с помощью целенаправленной селекционной работы[16].
Селекция фаленопсисов и других красивоцветущих орхидей развивается в двух направлениях: для срезки и для горшечной культуры.
Некоторые искусственные роды орхидей:
- Brassolaeliocattleya
- Vuylstekeara
- Rhynchosophrocattleya
См. также
Примечания
- ↑ Roberts E., Card L. (1933). V World Poultry Congr., 2, 353.
- ↑ 1 2 Morley F., Smith J. (1954). «Agric. Gaz. N. S. Wales» 65, N. 1, 17.
- ↑ Saeki J., Kondo K., et al. (1956). «Jpn. J. Breed.» 6, N. 1, 65.
- ↑ 1 2 Warren D. (1934). «Genetics» 19 600.
- ↑ 1 2 Дубинин Н. П., Глембоцкий Я. Л. (1967) Генетика популяций и селекция. М.: Наука с. 487, 496.
- ↑ 1 2 3 Добрынина А. Я. (1958) Реципрокные скрещивания московских кур и леггорнов. Тр. Ин-та генетики АН СССР, М, № 24, с. 307.
- ↑ Асланян М. М. (1962) Особенности наследования и эмбрионального развития поросят при скрещивании свиней крупной белой породы и шведский ландрас. Научн. докл. высш. школы, № 4, с. 179.
- ↑ 1 2 Александров Б. В. (1966) Рентгенографическое исследование варьирования и характера наследования числа позвонков при скрещивании свиней крупной белой породы и ландрас. Генетика. 2 № 7, с. 52.
- ↑ К. Вилли (1964) Биология. М. Мир. 678 с.
- ↑ Венский международный кодекс ботанической номенклатуры (2006)
- ↑ Dateline London, England – May 20, 2008. RHS Advisory Panel on Orchid Hybrid Registration (APOHR) Meeting.
- ↑ 1 2 3 Грант В. Эволюция организмов. М.: Мир, 1980. 480 с
- ↑ Dobzhansky Th. 1951. Genetics and the Origin of Species, 1st, 2d, and 3d eds. Columbia University Press, New York
- ↑ Dobzhansky Th. 1970. Genetics of the Evolutionary Process. Columbia University Press, New York
- ↑ Grun P. 1976. Cytoplasmic Genetics and Evolution. Columbia University Press, New York
- ↑ Ежек Зденек, Орхидеи. Иллюстрированная энциклопедия. Издательство: Лабиринт, 2005 г