Synergistic Effects of Interleukin-1β, Interleukin-6, and Tumor Necrosis Factor-α: Central Monoamine, Corticosterone, and Behavioral Variations (original) (raw)
Main
It is clear that interactions occur between the immune, endocrine, central, and autonomic nervous systems. Immunological manipulations (or products of an activated immune system, e.g., cytokines) may affect neuroendocrine and central neurotransmitter processes, and conversely, neuroendocrine and central neurotransmitter alterations may impact on immune activity (Blalock 1994; Dunn 1990; Rivier 1993; Rothwell et al. 1997). It has been posited that, among other things, the immune system acts like a sensory organ informing the brain of antigenic challenge (Blalock 1994) and that immune activation may be interpreted by the CNS as a stressor (Anisman and Merali 1999; Dunn 1990). Further, cytokines may be part of a regulatory loop that, by virtue of effects on CNS functioning, might influence behavioral outputs and may even contribute to the symptoms of behavioral pathologies, including mood and anxiety-related disorders (Anisman and Merali 1999). Indeed, in humans, depression was associated with variations of plasma cytokines, including interleukin-1β (IL-1β), IL-1 receptor antagonist (IL-1Ra), IL-2, soluble IL-2 receptors, IL-6, and soluble IL-6 receptors (Anisman et al. 1998; Maes 1995; Maes et al. 1995; Muller and Ackenheil 1998).
Cytokines and bacterial endotoxins, such as lipopolysaccharide (LPS), induce a constellation of apparently adaptive behavioral changes, collectively referred to as “sickness behaviors” (Dantzer et al. 1996). For instance, these agents induce fever, reduce social exploration, sexual behaviors, and food consumption (Bluthe et al. 1992; Johnson et al. 1996; Plata-Salaman 1988; Plata-Salaman et al. 1988; O'Reilly et al. 1987). In addition, endotoxins may induce anxiogenic-like effects (Lacosta et al. 1999) and disrupt responding for rewarding brain stimulation (Borowski et al. 1998), possibly reflecting anhedonic effects elicited by the immune challenge. The behavioral effects of endotoxin and cytokine treatment are paralleled by increased hypothalamic-pituitary-adrenal (HPA) activity, as reflected by increased activity of corticotropin releasing hormone (CRH) and elevated plasma ACTH and corticosterone levels (Kakucksa et al. 1993; Tilders et al. 1993). Inasmuch as cytokines elicit several effects similar to those of LPS, it has been assumed that at least some of the endotoxin effects involve IL-1β, or this cytokine acting conjointly or synergistically with IL-6 and/or TNF-α (Dunn 1992a; Ebisui et al. 1994; Long et al. 1990; Zanetti et al., 1992; Zhou et al. 1996). Indeed, IL-1β and IL-6 synergistically increased plasma corticosterone (Zhou et al. 1996) and ACTH (Matta et al. 1992; Perlstein et al. 1991), whereas IL-1β and TNF-α synergistically stimulated IL-11 through the production of prostaglandin-E2 in rheumatoid synovial fibroblasts (Mino et al. 1998). As well, IL-1β and TNF-α synergistically reduced blood glucose levels (Vogel et al. 1991), social exploration (Bluthe et al. 1994), and food intake (Plata-Salaman et al. 1996; Sonti et al. 1996; Yang et al. 1994), and increased the levels of inducible nitric oxide synthase (Kuemmerle 1998).
In addition to their peripheral actions, IL-1β, IL-6, and TNF-α exert numerous central neurochemical effects. In this respect, IL-1β increases NE turnover in the PVN, as well as other hypothalamic nuclei (Dunn 1992b; Palazzolo and Quadri, 1992; Shintani et al. 1995). Likewise, IL-1β increases the accumulation of the serotonin metabolite 5-HIAA in the hypothalamus (Dunn 1992b) as well as prefrontal cortex and hippocampus (Zalcman et al. 1994), and may alter hypothalamic DA utilization (Masana et al. 1990; Palazzolo and Quadri 1992; Shintani et al. 1993). Furthermore, IL-1β increased in vivo hippocampal serotonergic functioning (Merali et al. 1997; Song et al. 1999), and that of hypothalamic NE, 5-HT, and DA release (Shintani et al. 1995). While considerably less attention has focused on IL-6 and TNF-α, these cytokines have been shown to alter central monoamine levels, albeit to a lesser degree than that observed after IL-1β treatment (Mefford et al. 1991; Zalcman et al. 1994).
In addition to their individual effects, it is conceivable that IL-1β, IL-6, and TNF-α may synergistically provoke central neurochemical changes, and may thus also impact on behaviors associated with particular transmitter alterations. Indeed, IL-1β was found to act synergistically with a mild stressor to increase monoamine variations within the prefrontal cortex, nucleus accumbens and dorsal hippocampus (Merali et al. 1997). Moreover IL-1β plus TNF-α acted synergistically to influence neurotoxicity in mixed neuronal/glial cell cultures containing IFNγ (Jeohn et al. 1998), to stimulate mitogen-activated protein kinase (Lu et al. 1997), and along with IL-6 to provoke TNF-α mRNA expression in rat C6 glioma cells (Gayle et al. 1998). However, unlike these effects, to our knowledge, no such synergisms have been reported with respect to central monoamine functioning. The present investigation evaluated the individual and synergistic effects of IL-1β, IL-6, and TNF-α on an index of illness behavior (consumption of a highly preferred food substance, i.e., chocolate milk), plasma corticosterone, and central monoamine levels and turnover.
EXPERIMENTS 1–3
Individual Effects of IL-1β, IL-6 and TNF-α
Subjects
Male CD-1 mice, eight weeks of age were obtained from Charles River Inc, Laprairie, Quebec, Canada. After arrival at the facility, mice were housed in groups of four and acclimated to the laboratory for at least two weeks. Room temperature was maintained at 21°C, and lighting was maintained on a 12 hr light/dark cycle (lights on at 0800). One week prior to the beginning of the experiments, mice were separated into individual polypropylene cages with wire mesh lids and maintained ad libitum on pellet mouse chow (5075 Rodent diet Autoclaved; Ralston Purina) and tap water.
All procedures of the present investigation met the guidelines of the Carleton University Animal Ethics Committee, as well as the guidelines set forth by the Canadian Council on Animal Care. At all times, efforts were made to minimize the number of animals used, and to minimize animal suffering.
Procedure
In order to avoid confounding effects related to the stress of food deprivation, in the present investigation sickness was evaluated by assessing consumption of a highly preferred food substance. Prior to cytokine administration, mice were given free access to chocolate milk for one hour each day during the light phase (1000–1100 hrs). Bottles were weighed at the beginning and end of the session in order to determine consumption. Following the establishment of a steady rate of drinking (three consecutive days over which consumption varied by less than 10%), mice received an acute i.p. injection of either IL-1β, IL-6, or TNF-α (at one of several doses) or vehicle (pyrogen-free sterile saline) in three independent experiments. Consumption was measured beginning 45 min (in the case of IL-1β and IL-6) or 30 min (in the case of TNF-α) following injection. Recovery data was collected 24 hr following cytokine administration.
Recombinant human IL-1β was kindly provided by Dr. Craig Reynolds (Biological Response Modifiers Program; National Cancer Institute, Frederick, MD, USA. Produced by E.I. Dupont De Nemours; specific activity = 1.81 × 104 U/μg). Recombinant human TNF-α (specific activity = 1.1 × 105 U/ μg) and IL-6 (specific activity = 7.0 × 103 U/μg) were obtained from R&D Systems. All drugs were suspended in pyrogen-free, phosphate buffered sterile saline (0.9%). IL-1β was administered in a single dose of either 0.025, 0.05, 0.1, 0.4, or 0.8 μg (n = 10 per group) in a volume of 0.4 ml. IL-6 was administered in a single dose of 0.1, 0.2, 0.8, or 1.6 μg (n = 10 per group), whereas TNF-α was administered in a single dose of 1, 2, or 4 μg (n = 10 per group). Vehicle animals (n = 10 per group) in each experiment were injected i.p. with 0.4 ml pyrogen-free sterile saline.
Plasma Corticosterone Assay
In a parallel experiment mice received i.p. administration of either IL-1β (0.025, 0.05, or 0.1 μg), IL-6 (0.2 or 0.8 μg), TNF-α (1, 2 or 4 μg), or their respective vehicles, as described in the behavioral study (n = 10 per group). Mice were decapitated 60 min afterward and trunk blood was collected in tubes containing 10 μl EDTA. The blood was centrifuged at 3500 rpm for 15 min, and the plasma frozen and stored at −80°C. The plasma corticosterone concentrations were determined, in duplicate, by radioimmunoassay using kits obtained from ICN Biomedicals, Inc. These assays were found to yield less than 10% intra- and interassay variability.
Brain Dissection Technique
Brains were rapidly removed and frozen in isopentane and stored at −80°C until processing. The dissection of the brain was conducted in a cold chamber, with the brain resting on a stage containing dry ice. The brain was placed on a stainless steel dissecting block and razor blades were sequentially slid into adjacent slots (spaced approximately 0.5 mm apart) sectioning the brain into a series of coronal sections. Sections were mounted on glass slides and discrete nuclei were removed by micropunch using a hollow 16 or 20 gauge needle with a bevelled tip, following the mouse brain atlas of Franklin and Paxinos (1997). Tissue samples, which included the PVN, locus coeruleus, central amygdala, dorsal hippocampus, medial prefrontal cortex, and nucleus accumbens were stored at −80°C for subsequent HPLC determination of the monoamines and their metabolites.
HPLC Procedure for Analysis of Brain Amine Levels
Levels of DA, NE, and 5-HT and their respective metabolites MHPG, DOPAC, and 5-HIAA were determined by HPLC using a modification of the method of Seegal et al. (1986). Tissue punches were sonicated in a homogenizing solution comprised of 14.17g monochloracetic acid, 0.0186 g disodium EDTA, 5.0 ml methanol, and 500 ml H20. Using a waters M-6000 pump, guard column, radial compression column (5 μ, C18 reverse phase, 8 mm × 10 cm), and a three cell coulometric electrochemical detector (ESA Model 5100 A), 20 μl of the supernatant was passed through the system at a flow rate of 1.5 ml/min (1400–1600 PSI). The mobile phase used for the separation was a modification of that used by Chiueh et al. (1983). Each liter consisted of 1.3 g of heptane sulfonic acid, 0.1 g disodium EDTA, 6.5 ml triethylamine, and 35 ml acetonitrile. The mobile phase was then filtered (0.22 μm filter paper) and degassed following which the pH was adjusted to 2.5 with phosphoric acid. The area and height of the peaks was determined using a Hewlett-Packard integrator. The protein content of each sample was determined using bichinchoninic acid with a protein analysis kit (Pierce Scientific, Brockville, Ontario, Canada) and a spectrophotometer (Brinkman; PC800 colorimeter).
Statistical Analysis
Plasma corticosterone and monoamine and metabolite concentrations for each of the cytokine treatments were analyzed by one-way analyses of variance, followed by Newman-Keuls multiple comparisons (α = 0.05). The chocolate milk consumption on the day prior to cytokine treatment, immediately after, and again 24 h following cytokine administration, was analyzed by repeated measures analysis of variance.
EXPERIMENTS 4–6
Synergistic Effects of IL-1β, IL-6, and TNF-α
As indicated earlier, there is reason to expect that the pro-inflammatory cytokines may have synergistic effects with respect to behavioral and hormonal variations. Using sub-optimal doses of the cytokines based on the preceding studies, Experiments 4–6 assessed the synergistic effect of co-administration of IL-1β and IL-6, IL-1β and TNF-α, and IL-6 and TNF-α on consumption of the palatable chocolate milk solution. Parallel experiments assessed the effects of these treatments on plasma corticosterone levels, central monoamine levels and turnover.
Experimental Procedure
A total of 40 experimentally naive, male, CD-1 mice were used in each of the Experiments 4–6. The subject characteristics and the housing conditions were identical to those described in the preceding studies. In 3 independent studies, mice were assigned to four conditions wherein they received two successive i.p. injections, in a total volume of 0.4 ml. Thus, in each experiment mice received either a single cytokine treatment, two cytokine treatments or only saline treatment. The IL-1β was administered in a dose of 0.025 μg, IL-6 in a dose of 0.1 μg, and TNF-α in a dose of 1.0 μg. Behavioral testing, as described in the preceding experiments, was conducted 1 hour following drug treatment.
RESULTS
Experiments 1–3
Variations of Consumption of Palatable Diet
The administration of both IL-1β and TNF-α reduced consumption of chocolate milk (see Figures 1a and 1b), respectively). The analysis of variance indicated significant Cytokine × Sampling period interactions for IL-1β [F(10,108) = 5.43, p < .01] and for TNF-α [F(6, 72) = 5.97, p < .01]. The multiple comparisons confirmed that the 0.05, 0.1, 0.2, and 0.8 μg doses of IL-1β reduced consumption of chocolate milk relative to baseline levels and relative to that of saline treated mice. Likewise, TNF-α reduced consumption of the chocolate milk relative to saline-treated mice, but this difference only reached statistical significance at the 4.0 μg dose. Consumption in all animals tested returned to baseline levels 24 hr following treatment. Among vehicle treated mice consumption did not vary over the sampling periods.
Figure 1
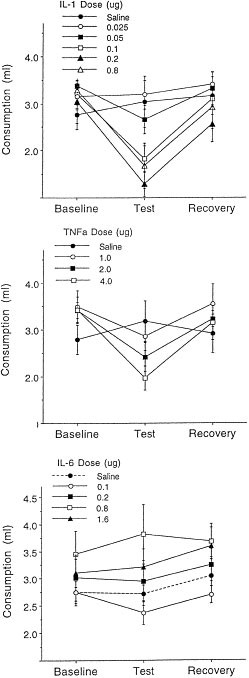
Mean (± SEM) consumption (ml) of chocolate milk among mice that received systemic IL-1β (top panel), TNF-α (middle panel), or IL-6 (lower panel) (n = 10/group). For each cytokine, mice received various doses or saline and consumption of milk was measured over a 1 hr period commencing 30 (for TNF-α) or 45 minutes (for IL-1 and IL-6) afterward. Baseline scores denote average consumption over the three days immediately preceding cytokine administration; recovery represents consumption 24 hr after the cytokine test
In contrast to the effects of IL-1β and TNF-α, systemic administration of IL-6 did not significantly affect consumption (see Figure 1c). Although a small decrease of chocolate milk intake was seen at the 0.1 or 0.2 μg doses, this decrease did not approach statistical significance. Indeed, at the higher doses tested (0.8 μg and 1.6 μg), a modest increase of consumption was detected relative to baseline levels.
Corticosterone Variations
When administered 1 hr prior to testing, IL-1β and TNF-α dose-dependently influenced plasma corticosterone levels [F(3,36) = 10.30, p < .01 and F(3,76) = 12.88, p < .01, respectively] (see Table 1) . The multiple comparisons indicated that in mice treated with 0.05 and 0.1 μg of IL-1β, corticosterone levels were significantly increased relative to saline-treated animals. Treatment with 0.025 μg of IL-1β produced a more modest, but non-significant increase in plasma corticosterone levels relative to saline-treated mice. Likewise, plasma corticosterone levels in mice treated with 4 μg of TNF-α were significantly increased when compared to mice treated with saline. The effects of the lower two doses did not differ from one other. In contrast, to the effects of IL-1β and TNF-α, plasma corticosterone levels were not affected by the IL-6 treatment.
Table 1 Plasma Corticosterone Levels (μg/dl protein) 1 hr after i.p. Cytokine Treatment
Central Monoamine Variations
One hour after IL-1β treatment, MHPG levels were influenced in the PVN [F(3,35) = 4.92, p < .05], locus coeruleus [F(3,36) = 4.28, p < .05], and central amygdala [F(3,35) = 2.75, p < .05]. Post hoc comparisons revealed elevated MHPG levels within the PVN at the 0.05 μg dose, in the locus coeruleus in response to 0.025 μg dose, and in the central amygdala after the 0.1 μg dose (see Figure 2). The levels of NE in these brain regions were not affected by the cytokine treatment. Within the central amygdala NE levels in response to the 0.025 and 0.1 μg doses of IL-1β (EquationSource math mover mixmo_ ± SEM = 21.77 ± 3.80 and 24.12 ± 4.40 ng/mg protein, respectively) were modestly reduced relative to vehicle treated mice (EquationSource math mover mixmo_ ± SEM = 31.81 ± 2.54 ng/mg), but this effect was variable and not statistically significant.
Figure 2
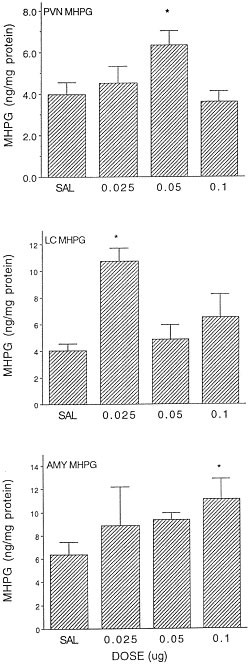
Mean (± SEM) MHPG concentrations within the PVN, locus coeruleus (LC), and central nucleus of the amygdala (AMY) 60 minutes following treatment with several doses of IL-1β (n = 8–10/group). * p < .05 relative to saline treated animals
In addition to the increased NE turnover, IL-1β altered the concentrations of 5-HT [F(3,36) = 6.51, p < .01] and its metabolite, 5-HIAA [F(3,32) = 5.72, p < .01] within the PVN. The multiple comparisons revealed that the high (0.1 μg) IL-1β dose increased both 5-HT and 5-HIAA levels within this site (see Figure 3). Additionally, in response to IL-1β treatment 5-HIAA accumulation was altered within the central amygdala [F(3,33) = 3.73, p < .05], and both 5-HT and 5-HIAA accumulation was altered in the prefrontal cortex [F(3,33) = 4.175; 5.40, p < .01, respectively]. Multiple comparisons revealed that the 0.05 and 0.1 μg doses increased the accumulation of metabolite within the central amygdala. Within the medial prefrontal cortex, an elevation of 5-HT was evident in response to the lowest dose of IL-1β (0.025 μg), whereas all three doses of IL-1β effectively elevated the metabolite levels (see Figure 3). Levels of DA and the metabolite, DOPAC, were also affected by the IL-1β treatment within the prefrontal cortex [Fs(3,32 and 3,33) = 7.84, 3.74, p < .05, respectively]. Post-hoc comparisons determined that, relative to vehicle-treated animals, increased amine and metabolite levels were evident at the 0.025 μg dose for DA (x ± SEM = 4.64 ± 0.68 vs. 9.7 ± 1.2 ng/mg protein, respectively) and at the 0.025 and 0.05 μg doses for DOPAC (x ± SEM = 3.25 ± 0.31 vs. 5.33 ± 0.44 and 5.28 ± 0.96 ng/mg protein, respectively). Unlike in the prefrontal cortex, the administration of IL-1β affected neither DA nor DOPAC within the nucleus accumbens.
Figure 3
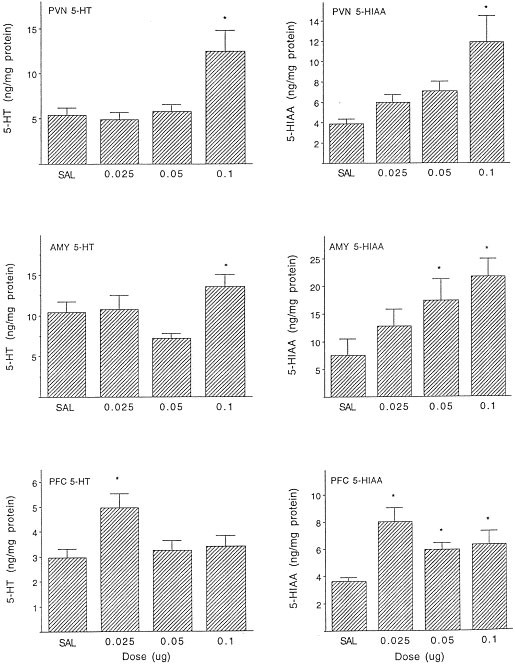
Mean (± SEM) 5-HT and 5-HIAA concentrations in the PVN, locus coeruleus (LC), and prefrontal cortex (PFC) 60 minutes following systemic IL-1β treatment (n = 8–10/group). * p < .05 vs. saline-treated animals
Treatment with TNF-α elicited region- and neurotransmitter-specific amine variations. In particular, levels of NE were not affected by the cytokine in the PVN, central amygdala, or prefrontal cortex, but were significantly reduced in the locus coeruleus [F(3,33) = 4.10, p < .05] (EquationSource math mover mixmo_ ± SEM = 19.86 ± 1.7, 13.23 ± 1.2, 13.76 ± 1.4 and 15.78 ± 1.5 ng/mg protein in the vehicle, 1.0, 2.0, and 4.0 μg doses, respectively) and dorsal hippocampus [F (3,34) = 3.55, p < .05] (EquationSource math mover mixmo_ ± SEM = 6.59 ± 0.45, 4.81 ± 0.39, 5.30 ± 0.33 and 5.15 ± 0.45 ng/mg protein, respectively) following all three doses. In contrast, levels of MHPG were not altered in any of these regions, although a modest elevation (20%) of the metabolite was apparent in the locus coeruleus.
Levels of 5-HT were altered by the TNF-α treatment within the PVN [F(3,26) = 3.52, p < .05] and prefrontal cortex [F(3,32) = 2.93, p < .05]. The multiple comparisons revealed that PVN 5-HT levels were reduced in response to each of the TNF-α doses (EquationSource math mover mixmo_ ± SEM = 19.34 ± 1.47, 11.61 ± 2.16, 11.74 ± 1.82 and 11.1 ± 2.42 ng/mg protein for vehicle, 1.0, 2.0, and 4.0 μg doses, respectively), but increased in the prefrontal cortex (vehicle = 2.58 ± 0.28, 1.0 μg = 4.17 ± 0.5, 2.0 μg = 3.7 ± 0.29 and 4 μg = 3.4 ± 0.37 ng/mg protein). In neither of these brain regions was the altered 5-HT level accompanied by variations of 5-HIAA accumulation. In the hippocampus, neither 5-HT nor 5-HIAA was affected by the cytokine. Finally, TNF-α increased nucleus accumbens DOPAC accumulation [F(3,29) = 3.71, p < .05], particularly at the 2.0 and 4.0 μg doses (vehicle = 20.24 ± 1.8, 2.0 μg = 32.08 ± 4.3 and 4.0 μg = 29.55 ± 2.1 ng/mg protein). Neither DA nor DOPAC accumulation was influenced in any of the other brain regions.
Few neurochemical effects were observed in response to systemic IL-6 treatment, although the cytokine was found to alter 5-HT [F(2,16) = 4.61, p < .05] and DA [F(2,19) = 6.33, p < .01] concentrations within the central amygdala. Multiple comparisons revealed that while 5-HT levels were raised in a dose-dependent fashion (vehicle = 5.37 ± 0.85, 1.0 μg = 7.52 ± 0.46, 0.8 μg = 13.07 ± 3.20 ng/mg protein), only the 0.1 μg IL-6 dose effectively increased amygdaloid DA levels (vehicle = 16.5 ± 2.1, 0.1 μg = 29.65 ± 3.3 ng/mg protein). In contrast to the parent amines, neither 5-HIAA nor DOPAC was affected by the cytokine treatment.
Experiments 4–6
Variations in Consumption of a Palatable Diet
Figure 4a shows the chocolate milk consumption as a function of the IL-1β and IL-6 treatments. The analysis of variance revealed a significant effect of the IL-1β × Sampling period interaction [F(2,72) = 8.257, p < .01]. Newman-Keuls multiple comparisons of the simple effects that comprised the interaction indicated that on Test day there was a modest, but significant, reduction (α = 0.05) in the amount of milk consumed by mice treated with 0.025 μg IL-1β + Vehicle, followed by an increase to baseline levels 24 hrs later. There was no reduction of consumption among vehicle treated animals. Furthermore, neither the main effect of IL-6, nor the interactions involving this variable approached significance.
Figure 4
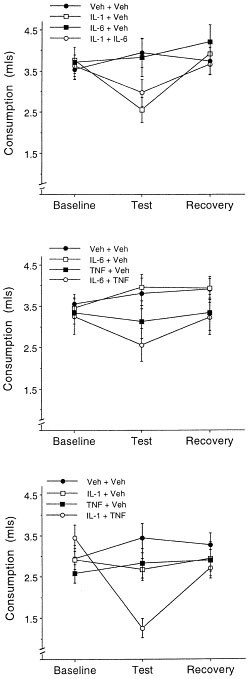
Mean (±SEM) consumption of chocolate milk among mice that received two systemic injections comprising two cytokines, a single cytokine, or no cytokine treatment. The top panel depicts the individual and combined effects of 0.025 μg IL-1 + 0.1 μg IL-6. The middle panel shows the individual and combined actions of 0.1 μg IL-6 + 1 μg TNF-α, whereas the lower panel shows the actions of 0.025 μg IL-1 + 1 μg TNF-α (n = 10/group). Consumption was measured over a 1 hr period commencing 1 hr following cytokine co-administration. Baseline scores denote average consumption over the three days immediately preceding cytokine administration; recovery represents consumption 24 hr after the cytokine test
There is reason to believe that the synergistic effects of IL-1β and IL-6 on HPA activity may be dependent on the timing of cytokine administration. The possibility thus also existed that synergism with respect to illness might be dependent on such a factor. Accordingly, two experiments assessed the conjoint effects of IL-1β and IL-6 under conditions wherein animals were tested 30 min after cytokine administration, or where the IL-1β treatment preceded that of IL-6. In the first of these studies, the procedure was identical to the preceding experiment (n = 8/group) except that testing began 30 min after cytokine co-administration. In the second experiment mice received two injections 30 min apart, and testing was conducted 1 hr after the second injection. Thus, mice received IL-1β (0.025 μg) followed by vehicle, or IL-1β followed by IL-6 (0.1 μg).
The analyses revealed that neither of these manipulations culminated in synergy between cytokine treatments. When testing commenced 30 min after their co-administration, consumption among the groups did not differ from one another or from baseline. On Test day, IL-1β provoked consumption (EquationSource math mover mixmo_ ± SEM = 3.07 ± 0.78 ml) that was moderately reduced relative to baseline (EquationSource math mover mixmo_ ± SEM = 3.46 ± 0.31), to vehicle-treated mice (EquationSource math mover mixmo_ ± SEM = 3.55 ± 0.51), and mice that received IL-6 alone (EquationSource math mover mixmo_ ± SEM = 3.25 ± 0.25). The combination of IL-1 + IL-6 (EquationSource math mover mixmo_ ± SEM = 3.18 ± 0.31) did not enhance the effects of IL-1β alone. Similarly, when IL-1β preceded IL-6 treatment by 30 min, the consumption (EquationSource math mover mixmo_ ± SEM = 2.88 ± 0.37) was no different from that of animals that received IL-1β in the absence of any other treatment (\(\overline{x}\)± SEM = 2.96 ± 0.45). In neither case was consumption different from that seen at baseline.
The combination of IL-6 (0.1 μg) + TNF-α (1 μg) did not act synergistically to reduce chocolate milk consumption (see Figure 4b). The ANOVA revealed a significant TNF-α × Sampling period interaction [F(2,72) = 5.807, p < .01], and multiple comparisons indicated that on Test day, injection of TNF-α reduced chocolate milk consumption relative to vehicle treated mice, but not in comparison to baseline consumption. Treatment with IL-6 did not influence consumption, and likewise the interaction between TNF-α and IL-6 was not significant.
In contrast to the aforementioned cytokine combinations, consumption was found to vary as a function of the IL-1β × TNF-α × Sampling period interaction [F(2,72) = 5.175, p < .01]. Multiple comparisons of the simple effects comprising this interaction confirmed that among vehicle treated animals consumption was stable over sampling periods. The low doses of IL-1β and TNF-α did not provoke statistically significant changes of consumption. However, the combination of these treatments markedly reduced consumption of chocolate milk relative to baseline scores, as well as in comparison with that of animals that received only one of the cytokines (see Figure 4c). Regardless of the treatment mice received, the consumption exhibited 24 hr afterward was comparable to the baseline scores. In effect, it appears that IL-1β + TNF-α elicited synergistic effects with respect to consumption of the palatable substance. It will be noted that at low doses the effects of both IL-1β and TNF-α were variable, as observed with respect to plasma corticosterone concentrations (see corticosterone results), likely reflecting that these doses were just at the threshold to elicit behavioral and neuroendocrine alterations. However, the combination of the treatments yielded more robust and less variable effects.
Corticosterone Variations
Administration of IL-1β increased plasma corticosterone concentrations [F(1,36) = 39.82, p < .01], whereas neither the main effect nor the interaction involving IL-6 approached statistical significance (see Figure 5a). Likewise, TNF-α increased plasma corticosterone levels [F(1, 36) = 19.79, p < .01], and this effect was not enhanced by IL-6 co-administration (see Figure 5b). In contrast, as observed in the behavioral study, the co-administration of IL-1β (0.025 μg) and TNF-α (1.0 μg) synergistically increased plasma corticosterone levels (see Figure 5c). Specifically, the analysis of variance revealed a significant IL-1β × TNF-α interaction [F(1, 36) = 4.01, p < .01], and multiple comparisons indicated that the separate administration of either IL-1β or TNF-α was not sufficient to significantly increase levels of corticosterone relative to vehicle-treated animals. However, when administered jointly, the increase of corticosterone levels was greater than the additive effects of the two treatments given alone.
Figure 5
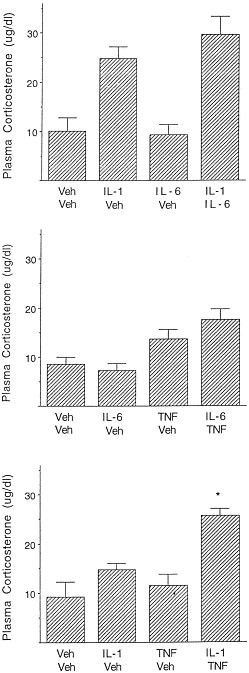
Mean (± SEM) plasma corticosterone concentrations 60 min following co-administration of two cytokines, a single cytokine, or no cytokine. The top panel depicts the individual and combined effects of 0.025 μg IL-1 + 0.1 μg IL-6. The middle panel depicts the individual and combined actions of 0.1 μg IL-6 + 1 μg TNF-α, whereas the actions of 0.025 μg IL-1 + 1 μg TNF-α are shown in the lowest panel (n = 10/group). * p < .05 relative to vehicle/vehicle treated animals. ° p < .05 relative to mice that received only a single cytokine treatment
It should be underscored, however, that in evaluating the combined effects of IL-1β and IL-6, the plasma corticosterone levels elicited by the IL-1β treatment alone were considerably higher than was observed in either the dose response study (Experiment 1) or the study evaluating the effects of IL-1β + TNF-α (Experiment 6). The relatively high level of plasma corticosterone observed in this particular study was surprising, as this dose of IL-1β ordinarily provoked plasma corticosterone concentrations in the 10–15 μg/dl range (see Experiment 1, and ensuing experiment). As indicated earlier, the 0.025 μg dose of IL-1β elicited effects that were just at threshold (see Experiment 1) and thus may have been subject to greater variability.
As indicated earlier, since IL-1β and IL-6 may synergistically influence HPA activity, depending on the timing of their administration (Perlstein et al. 1991; Zhou et al. 1996), an additional experiment assessed whether these cytokines would synergistically influence plasma corticosterone when blood was taken 30 min after treatment. The procedure was identical to that of the preceding study, except that mice were decapitated 30 min after administration of either vehicle (two successive i.p. injections), IL-1β (0.025 μg) plus vehicle, IL-6 (0.1 μg) plus vehicle, or IL-1β (0.025 μg) plus IL-6 (0.1 μg). The analysis of variance indicated that treatment with IL-1β increased corticosterone levels [F(1,28) = 8.30 p < .01]. Neither IL-6 nor the interaction between IL-1β and IL-6 approached statistical significance (vehicle = 5.57 ± 1.15; IL-1β = 13.24 ± 2.57; IL-6 = 7.02 ± 1.90; IL-1β + IL-6 = 13.95 ± 3.75).
Central Monoamine Variations
Unlike the synergistic effects observed with respect to behavioral and neuroendocrine variations, within the range of doses used, the cytokine combinations did not synergistically influence monoamine levels or turnover, and may even have provoked antagonistic effects. It will be recalled that the doses selected were relatively small and the cytokines generally induced limited effects on amine levels and turnover. Thus, from the outset it must be underscored that the absence of any synergy may be unique to the limited range of doses employed in this investigation.
In contrast to the monoamine alterations observed within the PVN following relatively high cytokine doses, neither NE nor MHPG was affected by the low doses of IL-1β, IL-6, or TNF-α administered alone or in combination. In Experiment 4, which assessed the independent and conjoint actions of IL-1β and IL-6, the level of 5-HT was reduced somewhat in animals that received IL-1β treatment alone (EquationSource math mover mixmo_ ± SEM = 7.67 ± 1.66) relative to saline treated animals (EquationSource math mover mixmo_ ± SEM = 15.33 ± 3.01). However, the treatment effects were variable and did not reach statistical significance. In the case of 5-HIAA, IL-1β was again found to provoke a modest but non-significant elevation of the metabolite concentration.
Analysis of the combined action of IL-6 and TNF-α revealed that 5-HT levels within the PVN declined in response to TNF-α administration [F(1,27) = 8.89_p_ < .01] (EquationSource math mover mixmo_ ± SEM = 11.80 ± 1.04 and 7.98 ± 0.64, for vehicle and TNF-α treated mice, respectively). It was further observed that TNF-α increased the levels of 5-HIAA, but this effect did not reach statistical significance. In response to the IL-1β and TNF-α treatments (Experiment 6), the analysis revealed that within the PVN, the level of 5-HT varied as a function of the interaction between the cytokine treatments [F(1,31) = 4.95, p < .05]. Multiple comparisons confirmed that IL-1β provoked a reduction of 5-HT, which was attenuated in animals that received the combination of IL-1β and TNF-α.
The accumulation of 5-HIAA was not influenced significantly by IL-1β administration, whereas a main effect of the TNF-α treatment was observed with respect to the metabolite levels [F(1,32) = 6.28, p < .05] (see Table 2) . A significant interaction between these treatments was not apparent with respect to 5-HIAA accumulation. However, it will be noted that while the elevation of 5-HIAA elicited by TNF-α alone did not reach statistical significance, the combination of IL-1β + TNF-α elicited 5-HIAA levels that exceeded those of the vehicle treated animals, although not those of mice that received the TNF-α alone.
Table 2 The Effect of IL-1 and TNF-α Co-administration on the Levels of 5-HT and 5-HIAA in the PVN 1 hr after Treatment
In Experiment 4 (IL-1β + IL-6), the levels of NE and MHPG within the locus coeruleus did not vary significantly as a function of the cytokine treatments, although as shown in Table 3 , IL-1β alone produced a modest increase of MHPG, coupled with a marginal decline of NE concentrations (p < .1). The analysis of the amine and metabolite levels in Experiment 5 (IL-6 + TNF-α) indicated that treatment with TNF-α alone significantly increased locus coeruleus MHPG [F(1,33) = 13.41, p < .01], whereas NE levels were unaffected. In contrast, IL-6 was uniformly without effect.
Table 3 The Effect of Cytokine Co-administration on the Levels of NE and MHPG in the Locus Coeruleus 1 hr after Treatment
Finally, in Experiment 6 (IL-1β + TNF-α), the 0.025 μg dose of IL-1β significantly elevated MHPG levels in the locus coeruleus [F(1,34) = 9.54, p < .01], whereas the effect of TNF-α was not statistically significant. Mice that received both IL-1β + TNF-α exhibited MHPG levels that exceeded that of vehicle treated animals; however, MHPG was not synergistically affected in response to the combination of the cytokine treatments (see Table 3).
The effect of the cytokines on NE activity within the central amygdala was relatively marked. While the co-administration of IL-1β + IL-6 did not influence NE levels within the central amygdala, a significant interaction between these cytokines was observed with respect to MHPG accumulation [F(1,34) = 4.65, p < .05]. Analysis of the simple effects comprising the interaction showed that IL-1β increased levels of the metabolite, whereas IL-6 was without effect. Curiously, the effects of the IL-1β treatment were attenuated in mice that had also received IL-6 administration. Analysis of the conjoint action of IL-1β and TNF-α indicated that within this region MHPG levels varied as a function of the IL-1β × TNF-α interaction [F(1,33) = 7.53, p < .05]. Analysis of the simple effects comprising the interaction revealed that both IL-1β and TNF-α administered individually increased MHPG levels. However, a further increase of MHPG was not elicited by the cytokine co-administration (see Table 4) .
Table 4 Effect of Cytokine Administration on the Levels of Biogenic Amines and Related Metabolites in the Central Nucleus of the Amygdala
The 5-HIAA accumulation within the central amygdala in the experiments assessing the synergistic actions of the cytokines was found to be unusually variable, and hence must be interpreted cautiously. Although IL-1β and IL-6 both produced almost two-fold increases of 5-HIAA accumulation, these effects were nonetheless nonsignificant (see Table 4). In the experiment where IL-6 was administered in combination with TNF-α, the IL-6 alone was found to increase levels of 5-HIAA [F(1,25) = 4.557_p_ < .05] (vehicle = 9.27 ± 1.73, and 0.1 μg = 13.71 ± 2.34 ng/mg protein), but this effect was not modified by the addition of TNF-α (16.48 ± 4.08 ng/mg protein). Furthermore TNF-α did not interact with IL-6 to alter monoamine levels or turnover in any of the other brain regions investigated. Finally, in assessing the effects of IL-1β + TNF-α, it was observed that although IL-1β elicited a modest elevation of 5-HIAA accumulation, this effect did not approach statistical significance, and was not further enhanced by co-administration of TNF-α.
DISCUSSION
Behavioral and Neuroendocrine Variations
It will be recalled that cytokines may engender a sickness profile (Kent et al. 1996), as well as neuroendocrine and central neurotransmitter changes reminiscent of those elicited by stressors (Anisman and Merali 1999; Dunn 1988, 1990). In fact, the possibility has been considered that immunological challenges and the presence of elevated circulating cytokine levels might be interpreted as a stressor (Anisman and Merali 1999; Dunn 1988). In accordance with the reports that cytokines may elicit sickness (Kent et al. 1996), systemically administered IL-1β and TNF-α suppressed the consumption of a highly palatable substance, possibly reflecting either an anorexic or an anhedonic response to cytokine administration. In contrast, IL-6 appeared to be without effect in this respect. The effects of IL-1β and TNF-α tended to be pronounced at the higher doses, but were relatively variable at the lower doses. Across experiments, for instance, the lowest dose of IL-1β (0.025 μg) generally did not influence consumption, although a relatively modest but statistically significant reduction of consumption was apparent in the experiment assessing the combined effects of IL-1β and IL-6.
Commensurate with earlier reports, both IL-1β and TNF-α increased plasma corticosterone concentrations, whereas IL-6 did not elicit such an outcome (Connor et al. 1998; Zalcman et al. 1994). At the 0.05 μg dose of IL-1β, the level of plasma corticosterone was elevated (Anisman and Merali 1999; Lacosta et al. 1998), whereas at the 0.025 μg dose, the observed effects, as in the case of the consumption studies, were more variable across experiments. Likewise, while the 1.0 μg dose of TNF-α was uniformly without effect on plasma corticosterone levels, at this low dose significant elevations of the hormone have been observed (Anisman et al. 1998; Hayley et al. 1999). It is important to recognize, however, that in the experiments assessing the synergistic effects of the cytokines, the doses used were near threshold for HPA activation, and hence the corticosterone response may have been subject to greater variability across experiments.
In assessing the potential synergistic effects of the cytokines, only a single dose of each cytokine was used based on the results of the initial studies that evaluated the dose-dependent effects of the treatments. However, as indicated by Berenbaum (1989), analysis of synergy may require multiple doses in order to assess shifts of the dose response curve. This is particularly the case with respect to cytokines, as their neurochemical actions are not necessarily linear and may be biphasic (Alonso et al. 1993; Lapchak 1992). Coupled with the fact that the cytokine doses used in the present investigation were at near-threshold levels, the results of the present investigation need to be considered cautiously, and the conclusions drawn are provisional. With respect to consumption of a favored food substance, it appeared that IL-6 did not enhance the effects of either IL-1β or TNF-α. In contrast, the co-administration of behaviorally sub-effective doses of IL-1β and TNF-α appeared to synergistically influence consumption, as previously observed with respect to feeding patterns (Plata-Salaman et al. 1996; Van der Meer et al. 1995; Yang et al. 1994).
Paralleling the behavioral effects, co-administration of IL-1β and TNF-α appeared to synergistically increase plasma corticosterone concentrations, whereas IL-6 did not augment the action of either these cytokines. While IL-1β and IL-6 synergistically influenced ACTH and corticosterone, depending on the timing of their administration (Perlstein et al. 1991; Zhou et al. 1996), such an outcome was not apparent in the present investigation. Moreover, in independent experiments there was no evidence of a synergism irrespective of whether blood was collected 30 min or 1 hr after treatment. Nevertheless, given that the effects of IL-1β on HPA activation may be augmented by IL-6 in a time-dependent fashion (Wang and Dunn 1999), the possibility cannot be excluded that a synergism would have appeared had other doses or time frames been assessed.
It is important to underscore that the present investigation involved administration recombinant human cytokines, rather than murine cytokines. Human and murine TNF-α (hTNF-α and mTNF-α) have functionally different effects in mice, as the latter excite both the p55 and the p75 receptors, whereas the former excites only the p55 receptor. It has been suggested that the differential effects on these receptors may account for the far greater lethality elicited by the murine form of TNF-α (Brouckaert et al. 1989a). It is significant that the effects of hTNF-α can be augmented by the co-administration of a “sensitizing” agent, so that the effects of this form of the cytokine will be comparable to that elicited by the mTNF-α. Among other agents, IL-1 was particularly effective in enhancing the effects of hTNF-α (Brouckaert et al. 1989b; Everaerdt et al. 1989). Thus, the possibility ought to be considered that the synergistic effects of hIL-1β and hTNF-α stem from direct or indirect activation of the p75 receptor, or actions that parallel the consequences of the conjoint activation of the p55 and p75 receptors. If this were the case, then it would be expected that synergistic effects would not be evident using the murine form of TNF-α, as both receptors will already have been activated.
Central Neurochemical Variations
The neurochemical variations induced by IL-1β, IL-6, and TNF-α were particularly intriguing in light of the suggestion that systemic stressors, such as cytokines, may influence HPA activity primarily via non-limbic mechanisms (Herman and Cullinan 1997). There have been several reports indicating that cytokine administration influences hypothalamic monoamine activity (Shintani et al. 1993), most notably within the PVN (Dunn 1988, 1992b). As well, IL-1β and TNF-α also increased amine activity at some extrahypothalamic sites (Ignatowski and Spengler 1994; Linthorst et al. 1995; Merali et al. 1997; Song et al. 1999), but few studies assessed the actions within mesolimbic regions, and none evaluated potential synergistic effects. It is clear from the present investigation, summarized in Table 5, that IL-1β and TNF-α influenced monoamine turnover in both limbic and non-limbic sites. Relatively low doses of IL-1β elevated MHPG, and 5-HIAA within the PVN, increased NE turnover at the locus coeruleus and central amygdala, 5-HT turnover within the central amygdala and prefrontal cortex, as well as DA utilization within the prefrontal cortex. In contrast, IL-1β had scant effects on hippocampal monoaminergic activity, although in vivo studies have indicated that IL-1β at higher doses enhanced 5-HT activity (Linthorst et al. 1995; Merali et al. 1997).
Table 5 Effect of Cytokine Administration on the Levels of Biogenic Amines in Various Brain Regionsa
The effects of TNF-α were less widespread than those of IL-1β, and tended to be more variable between experiments. Levels of 5-HT were affected within the PVN and prefrontal cortex, whereas NE levels were reduced within the locus coeruleus and the dorsal hippocampus. Yet, the turnover of NE within the locus coeruleus varied between experiments, being marked in the study examining the conjoint effects of TNF-α and IL-6, but absent in the dose response study and in the analysis of the combined effects of IL-1 and TNF-α. The source for these between-experiment differences is not evident, although it should be recognized that the doses of the human recombinant TNF-α employed in the present investigation were relatively modest and hence may have yielded limited effects. Finally, in contrast to IL-1β, it was observed that IL-6 had minimal effects, although it did increase 5-HT and DA levels within the central amygdala. Commensurate with the findings of Terao et al. (1993), neither IL-6 nor TNF-α significantly altered hypothalamic NE activity. However, the neurochemical effects of TNF-α may depend upon the particular hypothalamic nuclei investigated. In fact, TNF-α was shown to dose-dependently inhibit electrically stimulated NE release from axon terminals of isolated rat median eminence and may act in a modulatory capacity with respect to CRH release from this site (Elenkov et al. 1992).
The relationship between the central neurochemical and the corticosterone variations observed in the present investigation remains to be elucidated, and it is uncertain whether the amine changes were responsible for the neuroendocrine variations. Although the locus coeruleus sends only sparse projections to the PVN (Cunningham and Sawchenko 1988), it may be involved in the mediation of HPA activity (Gann et al. 1977). A wide range of stressors influence locus coeruleus activity (Abercrombie and Jacobs 1987; Weiss 1991), and it is conceivable that the HPA alterations induced by the cytokines were related to functional changes at this site. As well, NE within the PVN may contribute to HPA alterations elicited by cytokine challenge (Dunn 1988, 1992b; Whitnall 1993). Furthermore, 5-HT has important regulatory actions on HPA functioning, just as CRH release may influence forebrain 5-HT activity (Dinan 1996; Feldman et al. 1987; Korte et al. 1991). It is tempting to speculate that the IL-1β-provoked increase of NE and 5-HT turnover within the PVN, as well as the increased turnover of 5-HT elicited by TNF-α, may be related to the HPA activation induced by these cytokines.
Although IL-1β and TNF-α had clear neuroendocrine and central neurochemical effects, and appeared to act synergistically with respect to food consumption and plasma corticosterone levels, there was no evidence of a synergism with respect to central monoamine activity. Clearly, the joint actions of these cytokines on certain aspects of endocrine functioning are not necessarily translated into central monoamine changes. It may be that the cytokine doses in the present investigation were not optimal to provoke synergistic central amine changes, particularly in view of the limited IL-1β concentrations that likely reach the brain (Hopkins and Rothwell 1995). Furthermore, as indicated earlier, analysis of synergism might have been more profitably assessed using a variety of cytokine doses. Indeed, in the present investigation the greatest amine changes were not necessarily elicited by the highest cytokine doses. Furthermore, within the amygdala the elevation of MHPG associated with IL-1β were attenuated by the co-administration of IL-6.
Although the source for such an antagonistic effect cannot be determined from the present investigation, the diminished response associated with the combination of treatments may have been related to the nature of the dose response curves associated with the individual cytokines. Alternatively, it is possible that synergistic effects within the brain were limited by strict regulatory processes. For instance, treatment with a second cytokine may have promoted negative feedback effects, actions on autoreceptors, or synthesis of endogenous receptor antagonists (e.g., IL-1ra) which would have acted against the actions of IL-1β. Finally, in the present study monoamine variations were assessed in post-mortem tissues, and it is certainly likely that in vivo determinations of amine release would have provided a better index of the dynamic effects associated with cytokine treatment. In fact, synergistic effects between cytokines and stressors have been detected in vivo (Merali et al. 1997).
The finding that central monoamine activity was influenced by cytokines is consistent with the view that the immune system and the brain interact, and lends support to the potential psychological effects associated with such treatments (Maier and Watkins 1998). In this respect, it was shown that behavioral changes (e.g., locomotor activity) may correspond with in vivo amine alterations elicited by cytokines and by endotoxin administration (Linthorst et al. 1995; Linthorst and Reul 1998). In addition, such treatments may impact on anxiogenic and anhedonic processes (Borowski et al. 1998; Connor et al. 1998), both of which are related to depressive illness. Indeed, the finding that IL-1β and TNF-α influenced consumption of a palatable substance, increased HPA activity, and altered monoamine turnover in brain regions thought to be affected by stressors, is commensurate with the proposition that these cytokines may be related to depressive illness (Maes 1995). It is significant, as well, that in human trials the use of some cytokines, such as IL-2, have been found to provoke marked psychiatric and cognitive disturbances (Capuron et al. 1998; Caraceni et al. 1992; Denicoff et al. 1987) sufficiently severe to require discontinuation of treatment (Meyers and Valentine 1995). Clearly, a much better understanding of cytokine effects on central neurochemical functioning and on cognitive processes is necessary in order to optimize the efficacy and tolerability of immunotherapy.
References
- Abercrombie ED, Jacobs BL . (1987): Single-unit response of noradrenergic neurons in the locus coeruleus of freely moving cats. I. Acutely presented stressful and nonstressful stimuli. J Neurosci 9: 2837–2843
Google Scholar - Alonso R, Chaudieu I, Diorio J, Krishnamurthy A, Quirion R, Boksa P . (1993): Interleukin-2 modulates evoked release of [3H]dopamine in rat cultured mesencephalic cells. J Neurochem 61: 1284–1290
CAS PubMed Google Scholar - Anisman H, Merali Z . (1999): Cytokines and stress in relation to anxiety and anhedonia. In Dantzer R, Wollman EE, Yirmiya R (eds), Cytokines, Stress and Depression. New York, Plenum Press
Google Scholar - Anisman H, Ravindran AV, Griffiths J, Merali Z . (1998): Endocrine and cytokine correlates of major depression and dysthymia with typical or atypical features. Mol Psychiatry 4: 182–188
Google Scholar - Berenbaum MC . (1989): What is synergy? Pharmacol Rev 41: 93–141
CAS PubMed Google Scholar - Blalock JE . (1994): The syntax of immune-neuroendocrine communication. Immunol Today 15: 504–511
CAS PubMed Google Scholar - Bluthe RM, Dantzer R, Kelley KW . (1992): Effects of interleukin-1 receptor antagonist on the behavioral effects of lipopolysaccharide in the rat. Brain Res 575: 318–320
Google Scholar - Bluthe RM, Pawlowski M, Suarez S, Parnet P, Pittman Q, Kelley KW, Dantzer R . (1994): Synergy between tumor necrosis factor-α and interleukin-1 in the induction of sickness behavior in mice. Psychoneuroendocrinology 19: 197–207
CAS PubMed Google Scholar - Borowski T, Kokkinidis L, Merali Z, Anisman H . (1998): Lipopolysaccharide, central in vivo amine alterations, and anhedonia. NeuroReport 9: 3797–3802
CAS PubMed Google Scholar - Brouckaert PG, Everaerdt B, Libert C, Takahashi N, Fiers W . (1989a): Species specificity and involvement of other cytokines in endotoxin shockaction of recombinant tumor necrosis factor in mice. Agents Actions 26: 196–198
CAS PubMed Google Scholar - Brouckaert PG, Libert C, Everaerdt B, Takahashi N, Fiers W . (1989b): A role for interleukin-1 in the in vivo actions of tumor necrosis factor. Lymphok Res 8: 269–274
CAS Google Scholar - Capuron L, Ravaud A, Radat F, Dantzer R, Goodall G . (1998): Affects of interleukin-2 and alpha-interferon cytokine immunotherapy on the mood and cognitive performance of cancer patients. Neuroimmunomodulation 5: 9
Google Scholar - Caraceni A, Martini C, Belli F, Mascheroni L, Rivoltini L, Arienti F, Cascinelli N . (1992): Neuropsychological and neurophysiological assessment of the central effects of interleukin-2 administration. Eur J Cancer 29: A: 1266–1269
Google Scholar - Chiueh CC, Zukowska-Grojec Z, Kirk KL, Kopin JJ . (1983): 6-fluorocatechaolamine as a false adrenergic neurotransmitter. J Pharmac Exp Ther 225: 529–533
CAS Google Scholar - Connor TJ, Song C, Leonard BE, Merali Z, Anisman H . (1998): An assessment of the effects of central interleukin-1β, -2, -6 and tumor necrosis factor-α administration on some behavioral, neurochemical, endocrine and immune parameters in the rat. Neuroscience 84: 923–933
CAS PubMed Google Scholar - Cunningham ET Jr, Sawchenko PE . (1988): Anatomical specificity of noradrenergic inputs to the paraventricular and supraoptic nuclei of the rat hypothalamus. J Comp Neurol 274: 60–76
PubMed Google Scholar - Dantzer R, Bluthe RM, Auber A, Goodall G, Bret-Dibat JL, Kent S, Goujon E, Laye S, Parnet P, Kelley KW . (1996): Cytokine actions on behavior. In Rothwell NJ (ed), Cytokines and the Nervous System. London, Landes, pp 117–144
Google Scholar - Denicoff KD, Rubinow DR, Papa MZ, Simpson L, Seipp LA, Lotze MT, Chang AE, Rosenstein D, Rosenberg SA . (1987): The neuropsychiatric effects of treatment with interleukin-2 and lymphokine-activated killer cells. Ann Int Med 107: 293–300
CAS PubMed Google Scholar - Dinan T . (1996): Serotonin and the regulation of hypothalamic-pituitary-adrenal axis function. Life Sci 58: 1683–1694
CAS PubMed Google Scholar - Dunn AJ . (1988): Systemic interleukin-1 administration stimulates hypothalamic norepinephrine metabolism paralleling the increased plasma corticosterone. Life Sci 43: 429–435
CAS PubMed Google Scholar - Dunn AJ . (1990): Interleukin-1 as a stimulator of hormone secretion. Prog Neurol Endocrinol Immunol 3: 26–34
Google Scholar - Dunn AJ . (1992a): The role of interleukin-1 and tumor necrosis factor alpha in the neurochemical and neuroendocrine responses to endotoxin. Brain Res Bull 6: 807–812
Google Scholar - Dunn AJ . (1992b): Endotoxin-induced activation of cerebral catecholamine and serotonin metabolism: Comparison with interleukin-1. J Pharmac Exp Ther 261: 964–969
CAS Google Scholar - Ebisui O, Fukata J, Murakami N, Kobayashi H, Segawa H, Muro S, Hanaoka I, Naito Y, Masui Y, Ohmoto Y, Imura H, Nakao K . (1994): Effect of IL-1 receptor antagonist and antiserum to TNF-α on LPS-induced plasma ACTH and corticosterone rise in rats. Am J Physiol 266: E986–E992
CAS PubMed Google Scholar - Elenkov IJ, Kovacs K, Duda E, Stark E, Vizi ES . (1992): Presynaptic inhibitory effect of TNF-alpha on the release of noradrenaline in isolated median eminence. J Neuroimmunol 41: 117–120
CAS PubMed Google Scholar - Everaerdt B, Brouckaert PG, Shaw A, Fiers W . (1989): Four different interleukin-1 species sensitize to the lethal action of tumor necrosis factor. Biochem Biophys Res Commun 163: 378–385
CAS PubMed Google Scholar - Feldman S, Conforti N, Melamed E . (1987): Paraventricular nucleus serotonin mediates neurally stimulated adrenocortical secretion. Brain Res Bull 18: 165–168
CAS PubMed Google Scholar - Franklin KBJ, Paxinos G . (1997): The Mouse Brain in Stereotaxic Coordinates. San Diego, Plenum
Google Scholar - Gann DS, Ward DG, Baertschi AJ, Carlson DE, Maran JW . (1977): Neural control of ACTH release in response to hemorrage. Ann NY Acad Sci 297: 477–497
CAS PubMed Google Scholar - Gayle D, Ilyin SE, Miele ME, Plata-Salaman C . (1998): Modulation of TNF-alpha mRNA production in rat C6 glioma cells by TNF-alpha, IL-1beta, IL-6, and IFN-alpha: In vitro analysis of cytokine-cytokine interactions. Brain Res Bull 47: 231–235
CAS PubMed Google Scholar - Hayley S, Brebner K, Lacosta S, Merali Z, Anisman H . (1999): Sensitization to the effects of tumor necrosis factor-α; Neuroendocrine, central monoamine and behavioral variations. J Neurosci 19: 5654–5665
CAS PubMed Google Scholar - Herman JP, Cullinan WE . (1997): Neurocircuitry of stress: Central control of the hypothalamo-pituitary-adrenocortical axis. Trends Neurosci 20: 78–84
CAS PubMed Google Scholar - Hopkins SJ, Rothwell NJ . (1995): Cytokines and the nervous system. I. Expression and recognition. Trends Neurosci 18: 83–88
CAS PubMed Google Scholar - Ignatowski TA, Spengler RN . (1994): Tumor necrosis factor-α: Presynaptic sensitivity is modified after antidepressant drug administration. Brain Res 665: 293–299
CAS PubMed Google Scholar - Jeohn GH, Kong LY, Wilson B, Hudson P, Hong JS . (1998): Syergistic neurotoxic effects of combined treatments with cytokines in murine primary mixed neuron/glia cultures. J Neuroimmunol 85: 1–10
CAS PubMed Google Scholar - Johnson RW, Propes MJ, Shavit Y . (1996): Corticosterone modulates behavioral and metabolic effects of lipopolysaccharide. Am J Physiol 270: R192–R198
CAS PubMed Google Scholar - Kakucksa I, Qi Y, Clark BD, Lechan RM . (1993): Endotoxin-induced corticotropin-releasing hormone gene expression in the paraventricular nucleus is mediated by interleukin-1. Endocrinology 133: 815–821
Google Scholar - Kent S, Bret-Dibat JL, Kelley KW, Dantzer R . (1996): Mechanisms of sickness induced decreases in food-motivated behavior. Neurosci Biobehav Rev 20: 171–175
CAS PubMed Google Scholar - Korte SM, Van Dunn S, Bouws GA, Koolhaas JM, Bohus B . (1991): Involvement of hypothalamic serotonin in activation of the sympathoadrenomedullary system hypothalamo-pituitary-adrenocortical axis in male wistar rats. Eur J Pharmacol 197: 225–228
CAS PubMed Google Scholar - Kuemmerle JF . (1998): Synergistic regulation of NOS II expression by IL-1 beta and TNF-alpha in cultured rat colonic smooth muscle cells. Am J Physiol 274: G178–G185
CAS PubMed Google Scholar - Lacosta S, Merali Z, Anisman H . (1999): Behavioral and neurochemical consequences of lipopolysaccharide in mice: Anxiogenic-like effects. Brain Res 818: 291–303
CAS PubMed Google Scholar - Lacosta S, Merali Z, Anisman H . (1998): Influence of interleukin-1beta on exploratory behaviors, plasma ACTH, corticosterone, and central biogenic amines in mice. Psychopharmacology 137: 351–361
CAS PubMed Google Scholar - Lapchak PA . (1992): A role for IL-2 in the regulation of striatal dopaminergic function. NeuroReport 3: 165–168
CAS PubMed Google Scholar - Linthorst AC, Reul JM . (1998): Brain neurotransmission during peripheral inflammation. Ann NY Acad Sci 840: 139–152
CAS PubMed Google Scholar - Linthorst ACE, Flachskamm C, Muller-Preuss P, Holsboer F, Reul JMHM . (1995): Effect of bacterial endotoxin and interleukin-1β on hippocampal serotonergic neurotransmission, behavioral activity, and free corticosterone levels: An in vivo microdialysis study. J Neurosci 15: 2920–2934
CAS PubMed Google Scholar - Long NC, Otterness I, Kunkel SL, Vander AJ, Kluger MJ . (1990): Roles of inteleukin-1β and tumor necrosis factor in lipopolysaccharide fever in rats. Am J Physiol 259: R724–R728
CAS PubMed Google Scholar - Lu G, Beuerman RW, Zhao S, Sun G, Nguyen DH, Ma S, Kline DG . (1997): Tumor necrosis factor-alpha and interleukin-1 induce activation of MAP kinase and SAP kinase in human nueroma fibroblasts. Neurochem Intl 30: 401–410
CAS Google Scholar - Maes M . (1995): Evidence for an immune response in major depression: A review and hypothesis. Prog Neuro-Psychopharmacol Biol Psychiatry 19: 11–38
CAS Google Scholar - Maes M, Meltzer HY, Bosmans E, Bergmans R, Vandoolaeghe E, Ranjan R, Desnyder R . (1995): Increased plasma concentrations of interleukin-6, soluble interleukin-6, soluble interleukin-2 and transferrin receptor in major depression. J Affect Disord 34: 301–309
CAS PubMed Google Scholar - Maier SF, Watkins LR . (1998): Cytokines for psychologists: Implications of bidirectional immune-to-brain communication for understanding behavior, mood, and cognition. Psychol Rev 105: 83–107
CAS PubMed Google Scholar - Masana MI, Heyes MP, Mefford IN . (1990): Indomethacin prevents increased catecholamine turnover in rat brain following systemic endotoxin challenge. Prog Neuro-Psychopharmacol Biol Psychiatry 14: 609–621
CAS Google Scholar - Matta SG, Weatherbee J, Sharp BM . (1992): A central mechanism is involved in the secretion of ACTH in response to IL-6 in rats: Comparison to and interaction with IL-1β. Neuroendocrinology 56: 516–525
CAS PubMed Google Scholar - Mefford IN, Masters CF, Heyes MP, Eskay RL . (1991): Cytokine-induced activation of the neuroendocrine stress axis persists in endotoxin-tolerant mice. Brain Res 557: 327–330
CAS PubMed Google Scholar - Merali Z, Lacosta S, Anisman H . (1997): Effects of interleukin-1beta and mild stress on alterations of norepinephrine, dopamine and serotonin neurotransmission: A regional microdialysis study. Brain Res 761: 225–235
CAS PubMed Google Scholar - Meyers CA, Valentine AD . (1995): Neurological and psychiatric adverse effects of immunological therapy. CNS Drugs 3: 56–68
Google Scholar - Mino T, Sugiyama E, Taki H, Kuroda A, Yamashita N, Maruyama M, Kobayashi M . (1998): Interleukin-1alpha and tumor necrosis factor alpha syergistically stimulate prostaglandin E2-dependent production of interleukin-11 in rheumatoid synovial fibroblasts. Arthritis Rheum 41: 2004–2013
CAS PubMed Google Scholar - Muller N, Ackenheil M . (1998): Psychoneuroimmunology and the cytokine action in the CNS: Implications for psychiatric disorders. Prog Neuropsychopharmacol Biol Psychiatry 22: 1–33
CAS PubMed Google Scholar - O'Reilly B, Vander AJ, Kluger MJ . (1987): Effects of chronic infusion of lipopolysaccharide on food intake and body temperature of the rat. Physiol Behav 42: 287–291
Google Scholar - Palazzolo DL, Quadri SK . (1992): Interleukin-1 inhibits serotonin release from the hypothalamus in vitro. Life Sci 51: 1797–1802
CAS PubMed Google Scholar - Perlstein RS, Moughey EH, Jackson WE, Neta R . (1991): Interleukin-1 and interleukin-6 act synergistically to stimulate the release of adrenocorticotropic hormone in vivo. Lymphok Cytok Res 10: 141–146
CAS Google Scholar - Plata-Salaman CR . (1988): Food intake suppression by imunomodulators. Neurosci Res Comm 3: 159–165
CAS Google Scholar - Plata-Salaman CR, Oomura Y, Kai Y . (1988): Tumor necrosis factor and interleukin-1β: Suppression of food intake by direct action in the central nervous system. Brain Res 448: 106–114
CAS PubMed Google Scholar - Plata-Salaman CR, Sonti G, Borkoski JP, Wilson CD, Ffrench-Mullen JMH . (1996): Anorexia induced by chronic central administration of cytokines at estimated pathophysiological concentrations. Physiol Behav 60: 867–875
CAS PubMed Google Scholar - Rivier C . (1993): Effect of perhiperal and central cytokines on the hypothalamic-pituitary-adrenal axis of the rat. Ann NY Acad Sci 697: 97–105
CAS PubMed Google Scholar - Rothwell NJ, Allan S, Toulmond S . (1997): The role of interleukin-1 in acute neurodegeneration and stroke: pathophysiological and therapeutic implications. J Clin Invest 100: 2648–2652
CAS PubMed PubMed Central Google Scholar - Seegal RF, Bosh KO, Bush B . (1986): High performance liquid chromatography of biogenic amines and metabolites in the brain, cerebrospinal fluid, urine and plasma. J Chromatogr 377: 131–144
CAS PubMed Google Scholar - Shintani F, Kanba S, Nakaki T, Nibuya M, Kinoshita N, Suzuki E, Yagi G, Kato R, Asai M . (1993): Interleukin-1β augments release of norepinephrine, dopamine and serotonin in the rat anterior hypothalamus. J Neurosci 13: 3574–3581
CAS PubMed Google Scholar - Shintani F, Nakaki T, Kanba S, Sato K, Yagi G, Shiozawa M, Asio S, Kato R, Asai M . (1995): Involvement of interleukin-1 immobilization stress-induced increase in adrenocorticotropic hormone and in the release of hypothalamic monoamines in the rat. J Neurosci 15: 1961–1970
CAS PubMed Google Scholar - Song C, Merali Z, Anisman H . (1999): Variations of nucleus accumbens dopamine and serotonin following systemic interleukin-1, interleukin-2 or interleukin-6 treatment. Neuroscience 88: 823–836
CAS PubMed Google Scholar - Sonti G, Ilyin SE, Plata-Salaman C . (1996): Anorexia induced by cytokine interactions at pathophysiological concentrations. Am J Physiol 270: R1394–R1402
CAS PubMed Google Scholar - Terao A, Oikawa M, Saito M . (1993): Cytokine-induced change in hypothalamic norepinephrine turnover: Involvement of corticotropin-releasing hormone and prostaglandins. Brain Res 622: 257–261
CAS PubMed Google Scholar - Tilders JJH, DeRijk RH, Van Dam AM, Vincent VAM, Schotanus K, Persoons JHA . (1993): Activation of the hypothalamus-pituitary-adrenal axis by bacterial endotoxins: Routes and intermediate signals. Psychoneuroendocrinology 19: 209–232
Google Scholar - Van der Meer MJM, Sweep CGJF, Pesman GJ, Borm GF, Hermus RMM . (1995): Synergism between IL-1β and TNF-α on the activity of the pituitary adrenal axis and on food intake of rats. Am J Physiol 268: E551–E557
CAS PubMed Google Scholar - Vogel SN, Henricson BE, Neta R . (1991): Roles of interleukin and tumor necrosis factor in lipopolysaccharide-induced hypoglycemia. Brain Behav Immun 59: 2494–2498
CAS Google Scholar - Wang J, Dunn AJ . (1999): The role of interleukin-6 in the activation of the hypothalamo-pituitary-adrenocortical axis and brain indoleamines by endotoxin and interleukin-1 beta. Brain Res 815: 237–248
Google Scholar - Weiss JM . (1991): Stress induced depression: Critical neurochemical and electrophysiological changes. In Madden J (ed), Neurobiology of Learning, Emotion and Affect. New York, Raven Press
Google Scholar - Whitnall MH . (1993): Regulation of the hypothalamic corticotropin-releasing hormone neurosecretory system. Prog Neurobiol 40: 573–629
CAS PubMed Google Scholar - Yang Z, Koseki M, Meguid MM, Gleason JR, Debonis D . (1994): Synergistic effect of rhTNF-α and rhIL-1α in inducing anorexia in rats. Am J Physiol 267: R1056–R1064
CAS PubMed Google Scholar - Zalcman S, Green-Johnson JM, Murray L, Nance DM, Dyck D, Anisman H, Greenberg A . (1994): Cytokine-specific central monoamine alterations induced by interleukin (IL)-1, IL-2 and IL-6. Brain Res 643: 40–49
CAS PubMed Google Scholar - Zanetti G, Heusmann D, Gerain J, Kohler J, Abbet P, Barras C, Lucas R, Glauser M, Baumgartner J . (1992): Cytokine production after intravenous or peritoneal gram negative bacterial challenge in mice. J Immunol 6: 1890–1897
Google Scholar - Zhou D, Shanks N, Riechman SE, Liang R, Kusnecov AW, Rabin BS . (1996): Interleukin 6 modulates interleukin-1 and stress-induced activation of the hypothalamic-pituitary-adrenal axis in male rats. Neuroendocrinology 63: 227–236
CAS PubMed Google Scholar
Acknowledgements
Supported by a grant in aid of research from the Medical Research Council of Canada. H.A. is an Ontario Mental Health Senior Research Fellow. The assistance of Jerzy Kulczycki is very much appreciated.
Author information
Authors and Affiliations
- Institute of Neuroscience, Carleton University, Ottawa, Ontario, Canada
Karen Brebner MS, Shawn Hayley MS, Robert Zacharko Ph.D & Hymie Anisman Ph.D - School of Psychology and Department of Cellular and Molecular Medicine, University of Ottawa, Ottawa, Ontario, Canada
Zul Merali Ph.D
Authors
- Karen Brebner MS
- Shawn Hayley MS
- Robert Zacharko Ph.D
- Zul Merali Ph.D
- Hymie Anisman Ph.D
Rights and permissions
About this article
Cite this article
Brebner, K., Hayley, S., Zacharko, R. et al. Synergistic Effects of Interleukin-1β, Interleukin-6, and Tumor Necrosis Factor-α: Central Monoamine, Corticosterone, and Behavioral Variations.Neuropsychopharmacol 22, 566–580 (2000). https://doi.org/10.1016/S0893-133X(99)00166-9
- Received: 28 January 1999
- Revised: 10 May 1999
- Accepted: 06 December 1999
- Issue Date: 01 June 2000
- DOI: https://doi.org/10.1016/S0893-133X(99)00166-9