Aberrant CDKN1A transcriptional response associates with abnormal sensitivity to radiation treatment (original) (raw)
Main
Radiation therapy is used to treat ∼50% of all cancer patients. However, radiation therapy can cause a range of adverse normal tissue responses, which can limit therapeutic doses delivered. Early reactions principally affect high turn-over tissues, such as skin, gastrointestinal tract and bone marrow. During treatment or within a few weeks of completing a fractionated radiotherapy course, skin erythema, dry or moist desquamation of the skin, mucositis, nausea and diarrhoea are typical signs of radiation toxicity. Prediction of those patients at risk of severe reactions is difficult. Considerable efforts have been made to correlate normal tissue toxicity with cellular responses to ionising radiation (IR). However, no significant relationships have yet been found between fibroblast or lymphocyte cytotoxicity and acute (Geara et al, 1993; Brock et al, 1995; Johansen et al, 1996) or late (Russell et al, 1998; Peacock et al, 2000) normal tissue reactions to IR. However, normal cell radiosensitivity in some cases may be an important factor predictive of radiation therapy response as illustrated by West et al (2001) whose results showed that blood lymphocyte radiosensitivity (SF2) is a highly significant prognostic factor for the risk of developing late radiation morbidity. In terms of other end points, Barber et al (2000) evaluated the predictive value of lymphocyte chromosome radiosensitivity in patients receiving radiotherapy for breast cancer and concluded that these assays perform poorly in predicting normal tissue effects. Wang et al (2005) and Lopez et al (2005) both failed to find a correlation between DNA repair capacities in peripheral blood lymphocytes and acute skin reaction during radiotherapy. To date, no cellular or molecular assay has been used in clinics to predict the severity of radiotherapy reactions. It is not clear if this is due to the absence of a suitable assay or the lack of a suitable indicator cell/tissue.
It is likely that individual radiosensitivity has a heritable component as demonstrated in syndromes such as ataxia telangiectasia (AT), Nijmegen breakage syndrome and Fanconi's anaemia (Andreassen, 2005). Some evidence suggests that gene expression patterns are abnormal in radiosensitive conditions such as AT (Watts et al, 2002); heterozygosity for mutation in ATM (ataxia telangiectasia mutated) may occur in 1% of individuals and has been associated with sensitivity to IR in vitro (Smilenov et al, 2001). Furthermore, gene expression can be affected by radiation exposure and some of these responses have a heritable component (Correa and Cheung, 2004). It is, therefore, reasonable to postulate that genetic variation contributes to determining the severity of response by irradiated tissues in the body and that this variation is reflected in gene expression patterns. Rapid and simple predictive assays of radiation response in easily accessible cells, for example lymphocytes, would bring clinical benefits (lymphocytes are an ideal cell population as they can be obtained by a minimally invasive method combining simplicity and rapidity as well as reliability, also repeat sampling is easy). Using multigene classifiers, earlier work showed that lymphoblastoid cell lines from patients with acute radiation toxicity have abnormal transcriptional responses to radiation (Rieger et al, 2004). Svensson et al (2006) also reported that changes of expression in a specific set of genes after in vitro irradiation of stimulated peripheral lymphocytes can, to some extent, successfully predict severe late reaction status. Using subcutaneous fibroblasts from breast cancer patients, Rodningen et al (2007) identified a set of 18 radiation-responsive genes, which may provide a predictive assay for late normal tissue reactions after radiotherapy. The amount of information available on gene expression responses to radiation has been increasing considerably in recent years (Kruse and Stewart, 2007). These studies of altered gene expression have been useful for elucidating the molecular mechanisms underlying cellular radiation response and a few have been able to identify genes as potential indicators of severe reactions to radiotherapy treatment. Surprisingly, in these studies, the most pronounced radiation-responsive genes (Rieger and Chu, 2004), which show high variation in expression between individuals, do not seem to be predictive of radiation toxicity. Those, which have been identified as informative, are associated with various pathways and differ between studies thus complicating the interpretation of the data; this remains a challenge, particularly at the level of individual gene expression. Our approach was to re-examine, in a rigorous quantitative manner, the expression response of a fewer number of genes associated with relevant pathways and previously identified as radiation responsive, in an attempt to correlate expression levels with normal tissue reaction to IR.
Exposure of cells to IR induces a large range of DNA alterations and results in complex biological responses. The DNA-damage response (DDR) network mediates DNA repair, cell cycle checkpoints and/or apoptosis. In response to DNA double-strand breaks, the ATM gene with its regulator the MRN (Mre11-Rad50-NBS1) complex are activated (Lee and Paull, 2005). Ataxia telangiectasia mutated protein is auto-phosphorylated at Ser-1981 in response to DNA damage, which causes dissociation of the inactive dimers (Bakkenist and Kastan, 2003). Ataxia telangiectasia mutated kinase is essential for activation of cell cycle checkpoints and DNA repair in response to IR. It acts upstream of p53 by activating p53 protein through phosphorylation of Ser-15 (Banin et al, 1998; Canman et al, 1998) leading to p53 stabilisation.
p53 protein is also activated through a number of other post-translational modifications, including phosphorylations, acetylations and methylation (Toledo and Wahl, 2006). After DNA damage induced by IR, cells enter either cell-cycle arrest or apoptosis, depending on which of the pathways is predominant in the specific cell type and environment. p53 functions as a crucial transcription factor in a well established response through direct protein binding to target gene promoter elements. Although cell-cycle arrest depends on the ability of p53 to induce the transcription of genes such as CDKN1A and GADD45, apoptosis depends on induction of a distinct class of genes including BBC3 also called PUMA (p53 upregulated modulator of apoptosis) (Yu et al, 2003). p53 also negatively regulates the expression of cell-cycle regulator genes such as CYCB1 (Badie et al, 2000; Imbriano et al, 2005). In this study, we speculated that patients overreacting to radiotherapy treatment may have abnormal transcriptional responses in one or more of the genes involved specifically in DDR pathways. Among a wide range of candidate genes (Rieger and Chu, 2004), CDKN1A, GADD45A, BBC3 and CCNB1 were selected for the present work.
It has been established that there are significant correlations between SNPs (single nucleotide polymorphisms) in genes related to the biological response to radiation injury (e.g. genes involved in DNA repair (BRCA1 and 2, XRCC1)) or DNA damage signalling (e.g. ATM) and the risk of radiation-induced normal tissue reaction (Andreassen et al, 2003). Very recently, it has been shown that chromosomal radiosensitivity can be influenced by genetic polymorphisms and that SNPs in nonhomologous end-joining genes may be associated with breast cancer risk (Willems et al, 2008). These genetic variations can affect gene transcription, the stability of the mRNA, the protein structure or protein–protein interactions. Of particular interest are the nonsynonymous SNPs that lead to amino-acid changes in the translated protein, therefore, having the potential to alter protein function and contribute to variations in response between patients. It has been suggested that genetic polymorphisms in TP53 might affect some of its functions. Among the TP53 variants, the common TP53 Arg72Pro has been shown to differ biochemically and biologically leading to different levels of apoptotis (Thomas et al, 1999) and exerting different effects on cell cycle progression (Pim and Banks, 2004). CDKN1A is transcriptionally activated by TP53 and both genes play a direct role in G1/S checkpoint control in response to IR. Earlier work suggested that there is a possible combined effect of polymorphisms in the two genes. An association between the risk of acute skin toxicity and TP53 72Pro carriers in those with the CDKN1A 31Ser genotype in a subset of normal weight patients treated with radiotherapy for breast cancer has been shown (Tan et al, 2006). Therefore, to examine the relationship between normal tissue radiosensitivity and polymorphisms in key genes, we also genotyped our samples for these two non-synonymous SNPs TP53 codon 72 Arg/Pro G>C and CDKN1A codon 31 Ser/Arg C>A, previously associated with radiation sensitivity (Alsbeih et al, 2007).
Materials and methods
Samples
Lymphocyte samples, separated on Histopaque-1077 (Sigma Aldrich, Poole, Dorset, UK) and frozen within 24 h of blood collection from 22 sporadic breast cancer patients 30–72 years old treated at the Christie Hospital in Manchester, were obtained between 1993 and 1996 after local excision but before radiotherapy, in a prospective study of acute skin reactions (Barber et al, 2000). None of the patients had had a mastectomy or had received chemotherapy, but most of the patients involved in the study were taking tamoxifen. The previously published work demonstrated no significant relationship between chromosomal radiosensitivity, tamoxifen intake, menopausal status, age or smoking history (Barber et al, 2000). Consent was obtained from the patients and South Manchester Medical Research Ethics Committee approved the study. Radiotherapy was delivered with a tangent pair of fields at a prescribed dose of 40 Gy to the mid-plane. Measurements of skin reactions were made on 201 patients, of which 13 showed severe reactions (sufficient to warrant premature termination radiotherapy, or where most of the breast had moist desquamation), the remainder having normal reactions. The mean age of the patients was 52.6 (normal reactors (NRs)) and 53.6 (severe reactors (SRs)). Lymphocytes for all patients were screened for homozygous or heterozygous ATM mutations but none were detected (Appleby et al, 1997).
Lymphocytes separated from blood samples from two healthy female donors (PH4B and JM1) were used to produce disease-free control T-cell lines; in addition, an AT T-cell line (AT58) was used, originally obtained from Dr C Arlett, University of Sussex (Cole and Arlett, 1994).
Cell growth
T-lymphocyte cultures were prepared using the method described previously. Briefly, frozen stocks were thawed and cultured in 10 ml of stimulating medium (SR10) at 2–3 × 105 cells per ml. Cultures were incubated for about 4 days at 37°C, 5% CO2 atmosphere, then disaggregated and counted daily, maintaining cell density between 0.2 and 1 × 106 cells per ml.
Irradiation of lymphocyte cultures
Cells were seeded at a density of 4 × 105 cells per ml in GR10 medium and irradiated with 2 Gy using a Siemens Stabilipan Therapy X-ray set (output 14 mA 250 kVp, 0.7 Gy min−1) at the MRC Harwell, then incubated in GR10 medium at 37°C and 5% CO2 for 2 h before processing.
RNA extraction and reverse transcription
Cells (2 × 106) were collected for each sample point by centrifugation, re-suspended in RNA Later (Ambion, Applied Biosystems, Foster City, CA, USA) and stored at −80°C. Total RNA was prepared by using RNAqueous®-4PCR (Ambion) kit. Reverse transcriptase reactions were performed with 1–1.5 _μ_g of total RNA (High Capacity cDNA Archive Kit, Applied Biosystems).
Quantitative real-time PCR
Real-time PCR was performed using iQ5 thermocyclers (Bio-Rad, Hercules, CA, USA). All reactions including no-template controls were run in triplicate using primer and probe sets for target genes (Table 1). FAM, HEX and Texas Red were used as fluorochrome reporters for the hydrolysis probes analysed in multiplexed reactions.
Table 1 PCR primers and probes for quantitative analysis of gene expression and primers used for PCR amplification and DNA sequencing of SNPs
Data were collected and analysed by iQ5 Detection System software. Gene target _C_t (cycle threshold) values were normalised to a Hypoxanthine–Guanine phosphoribosyltransferase 1 human (HPRT1) internal control. _C_t values were converted to transcript quantity using standard curves obtained by serial dilution of PCR-amplified DNA fragments of each gene or cDNA. The linear dynamic range of the standard curves covering five orders of magnitude (serial dilution from 4.10−2 to 1024.10−7) gave typical PCR efficiencies >90% for each gene with _R_2>0.95. Relative gene expression levels after irradiation were similarly obtained for comparison with preirradiation controls.
DNA extraction, amplification, sequencing and data analysis
The selected SNPs and PCR primers utilised are listed in Table 1. DNA was extracted using QIAamp DNA Blood Midi Kit (Qiagen, Hamburg, Germany). Relevant segments of DNA were amplified by thermal cycling, directly sequenced using the DYEnamic ET Dye Terminator Cycle Sequencing Kit (GE Heathcare, Piscataway, NJ, USA) and run on a MegaBase 1000 sequencer (Applied Biosystems). Sequencing results were aligned to the corresponding reference sequence and SNPs were genotyped using SeqManII sequence analysis software (DNASTAR Inc., Madison, WI, USA).
Analysis of variance (ANOVA) was used to compare the means of gene expression values between normals and overreactors. When possible, a nested ANOVA method was used to calculate the variance between experiments and the variance between groups and to draw comparisons.
The association between severity of acute reactions to radiotherapy and SNPs genotype was determined from odds ratios, confidence intervals and significance level or, where not applicable, Fisher's Exact tests. Statistical analyses used Epi Info database and statistics software for public health professionals (Centers for Disease Control and Prevention, Atlanta, GA, USA).
Results
To identify suitable genes, develop assays and establish their sensitivity, tests were carried out comparing responses of cultured T-lymphocytes from normal controls and an AT case. Cells from AT patients are defective in the initiation of cell-cycle checkpoints following DNA damage, as ATM kinase activity is critical for the appropriate initiation of signalling pathways. Results obtained for three genes CCNB1, CDKN1A and BBC3 from a healthy control (PH4b), a breast cancer patient with normal therapy reaction (NR 11) and an AT (AT58) case are presented in Figure 1. Both the disease-free control PH4b and NR 11 have strong and similar radiation-induced decreases in cyclin B1 expression, 2.9- and 3-fold, respectively. AT58 cells showed only a marginal decrease of cyclin B1 mRNA of 1.2-fold. Both the cell-cycle arrest promoter CDKN1A and the proapoptotic BBC3 in PH4b and NR 11 cells were strongly upregulated; AT58 responses were significantly less.
Figure 1
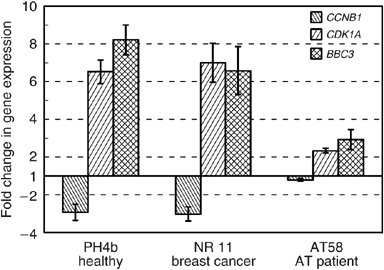
Quantitative PCR analysis of gene expression in cultured lymphocytes 2 h after 2 Gy X-rays from a healthy donor (PH4B), one breast cancer case with normal therapy reaction (NR 11) and an AT case (AT58). Values were normalised using HPRT as standard. The figures represent the ratio of level of expression after irradiation divided by the mock-treated cell expression levels. The mean value of 2–3 experiments reproduced at least twice each with three reactions are shown (error bars, s.e.).
To examine the relationship between acute skin reactions and gene activation/repression after irradiation in CCNB1, CDKN1, BBC3 and GADD45A (a well-characterised radiation-responsive gene; Grace et al, 2002), T-lymphocytes from 22 breast cancer patients were analysed. The NR and SR patient groups were comparable in size (11), sex, age and tumour status. Individuals showed a considerable range in baseline expression for the four genes (Table 2), although NR and SR groups were statistically indistinguishable.
Table 2 Expression of genes in NR and SR samples
After irradiation, there was a wide range of responses in terms of either repression (CCNB1) or induction (GADD45A, CDKN1A, BBC3) as shown in Figure 2. Statistical analysis of results is shown in Table 2, no statistical differences were found between NR and SR responses for GADD45A (_P_=0.98), CCNB1 (_P_=0.55) and BBC3 (_P_=0.55). By contrast, radiation-induced increases in CDKN1 expression were significantly less in SR than NR (P<0.0021). Nested ANOVA tests confirmed that replicate experiments performed consistently without significant differences between experiments (_P_=0.308). In Figure 3, the relative CDKN1A expression for all 22 patients is presented sorted in terms of increasing magnitude of response. There is an overlap between NR and SR and it is not possible to predict the normal tissue response on an individual basis for all samples. However, using a cutoff of seven-fold increase, reaction status in ∼91% (20/22) would be correctly identified with only two false negatives and no false positives.
Figure 2
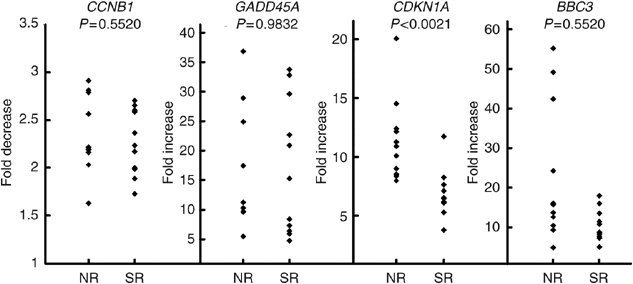
Gene expression analysis of CCNB1, GADD45A, CDKN1A and BBC3 by quantitative PCR in 22 cultures of cultured T-lymphocytes from female breast cancer patients (11 SR and 11 NR). Values were standardised to HPRT expression level. The gene expression level ratio of irradiated/mock-treated cell is presented. The mean values of triplicate experiments, each with three reactions, are shown.
Figure 3
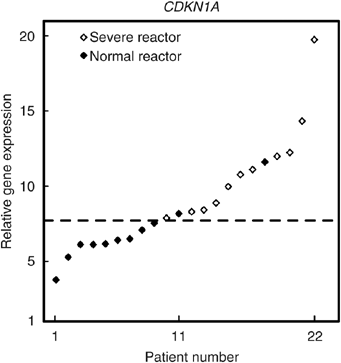
Gene expression analysis by quantitative PCR in 22 cultures of T-lymphocytes from female breast cancer patients (11 SR and 11 NR). The results presented show the cells response 2 h after 2 Gy X-rays and have been sorted into ascending magnitude of response (fold increase in CDKN1A gene expression). Values were standardised to HPRT expression level. The gene expression level ratio of irradiated/mock-treated cell is presented. The mean value of triplicate experiments, each with three reactions, are shown. The dotted line indicates the retrospectively defined cutoff for predicting radiation toxicity, with two false negatives and no false positive. Filled diamonds: SR, empty diamonds: NR
To determine whether the CDKN1 response differences between NR and SR could be related to polymorphisms in TP53 codon 72 or CDKN1A codon 31, sequencing was performed on DNAs from lymphocyte cultures of 12 SR patients as described in Materials and Methods plus two additional SR cultures. No significant differences in frequency were found between groups for either polymorphism (Table 3).
Table 3 Genotype and allelic frequencies of two polymorphisms assessed in 25 breast cancer patients treated with radiotherapy, 11 normal reactors (NR) and 14 severe reactors (SR)
Discussion
In this study, QRT-PCR assays have been used to assess the predictive value of four genes for acute skin reactions to radiotherapy. The transcripts levels of CCNB1, CDKN1A, BBC3 and GADD45, all identified as radiation responsive in previous gene expression array studies (Tusher et al, 2001; Jen and Cheung, 2003) and recognised as biomarkers of radiation exposure, have been analysed. This study, therefore, addresses the hypothesis that expression differences in genes involved in the DDR network associate with severity of normal tissue radiation toxicity. Expression of these genes was analysed in a set of breast cancer patients with differences in sensitivity to radiation treatment.
Specific genetic variation among populations contributes appreciably to differences in gene expression phenotypes (Morley et al, 2004; Spielman et al, 2007). Gene expression signatures highlighting the molecular characteristics of individual patients are already useful for personalised cancer therapy (Garman et al, 2007). In this study, it was speculated that intrinsic radiosensitivity differences in most or all tissues in the body are reflected in the expression of genes influencing radiation sensitivity.
The DDR network senses genotoxic stress and coordinates a response, which includes activation of transcription, cell cycle checkpoints, apoptosis, senescence and DNA repair processes. This coordination is essential for cell survival.
Results comparing AT and normal cells are in good agreement with knowledge of the ATM DNA-damage-dependent signal transduction pathway. Unlike the Rieger et al (2004) study where microarray data on lymphoblastoid cells failed to predict toxicity in subjects with defined genetic defects like AT, the present results with CCNB1, CDKN1A and BBC3 clearly demonstrated differential responses between a healthy donor and an AT case.
Of the genes studied, only CDKN1A expression allowed discrimination of NR from SR on average and in ∼91% of individuals. Although CDKN1A is a well-known damage response gene, it was not previously identified in array-based screens for genes predictive of early (Rieger et al, 2004) or late (Svensson et al, 2006) toxicity. CDKN1A is cyclin-dependent kinase inhibitor-1A also referred to as p21; this gene codes for a protein, which inhibits cyclin kinase activity; it is tightly regulated at the transcriptional level by p53 protein and serves as the effector of TP53 cell cycle control. CDKN1A is well known as a biological indicator of IR exposure in humans (Amundson et al, 2003). It is, therefore, not entirely surprising that CDKN1A expression level may be predictive of radiation toxicity; a plausible biological explanation exists in that knockout mouse studies demonstrate CDKN1A, which plays a role in protecting intestinal epithelial cells from radiation-induced apoptosis (Wang et al, 1997). Loss of CDKN1A in a context of ATM deficiency also leads to accelerated kinetics of acute radiation toxicity in vivo, principally because of effects on the gut epithelium, suggesting that CDKN1A has a crucial role in acute normal tissue response to IR (Wang et al, 1997). Amundson et al (2004) showed that in vivo patterns of stress-gene induction by IR in blood are similar to those observed ex vivo. They demonstrated that CDKN1A is induced in vivo in humans undergoing total body irradiation and was one of the most promising biomarkers that also showed interindividual variation in response. The expression level of one gene in lymphocytes in response to radiation cannot alone explain the response of complex tissues; the vascular system or cytokines involved in the inflammation process must contribute as well. Nevertheless, our results support the concept that gene expression differences, perhaps reflecting underlying genetic variants, seem to be associated with radiation toxicity.
At least 40 polymorphisms of CDKN1A have been identified (Li et al, 2005) of which 35 are intronic. Only seven have an allele frequency >10%, but they are ∼1.3 kb upstream of exon 2. Among the common SNPs, CDKN1A C98A is found in exon 2 and causes a non-synonymous serine-to-arginine substitution at codon 31. This SNP is located in a highly conserved region of the gene (Chedid et al, 1994) and, as it is likely to change the properties of the protein, it is the most widely studied of the CDKN1A SNPs.
In our group of patients, CDKN1A gene expression neither associated with the CDKN1A C31A SNP nor with the common SNP in TP53 (G72C). Larger studies would be needed to confirm these results. However, our results are in line with those obtained by Tan et al (2006) who examined the impact of these two SNPs on radiation response using the end point of acute skin toxicity after radiotherapy for breast cancer, no clear-cut association was found. In an in vitro study, Alsbeih et al (2007) described a significant association between cellular clonogenic radiosensitivity and TP53 G72C but not CDKN1A C31A. The discrepancy between these two sets of data may be due to the presence in vivo of cofactors specific to each patient that could mask subtle SNP associations.
There are other polymorphic SNPs in CDKN1A, which could be more relevant to the rate of gene expression, for example, some SNPs in the first intron containing the promoter with the p53 binding site. These SNPs have wide range of frequency (∼0.001–0.48) in the population and could influence the rate of gene expression by modifying the binding affinity of transcription factors. Therefore, screening of the whole CDKN1A genetic region would be necessary to determine the influence of all these SNP variations on gene expression and toxicity to radiation therapy.
To summarise, the data presented here support the hypothesis that at least some breast cancer patients, who develop severe reactions to radiotherapy, have an intrinsic radiosensitivity that can be identified in peripheral blood lymphocytes by quantifying gene expression response to IR and that CDKN1A is a surrogate marker for early effects of radiation therapy. Confirmation and extension of these results may enable development of simple clinical tests to predict the likely level of radiation toxicity and to individualise patient treatment.
Accession codes
Accessions
GenBank/EMBL/DDBJ
Change history
16 November 2011
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
- Alsbeih G, Al-Harbi N, Al-Buhairi M, Al-Hadyan K, Al-Hamed M (2007) Association between TP53 codon 72 single-nucleotide polymorphism and radiation sensitivity of human fibroblasts. Radiat Res 167: 535–540
Article CAS Google Scholar - Amundson SA, Grace MB, McLeland CB, Epperly MW, Yeager A, Zhan Q, Greenberger JS, Fornace Jr AJ (2004) Human in vivo radiation-induced biomarkers: gene expression changes in radiotherapy patients. Cancer Res 64: 6368–6371
Article CAS Google Scholar - Amundson SA, Lee RA, Koch-Paiz CA, Bittner ML, Meltzer P, Trent JM, Fornace Jr AJ (2003) Differential responses of stress genes to low dose-rate gamma irradiation. Mol Cancer Res 1: 445–452
CAS PubMed Google Scholar - Andreassen CN (2005) Can risk of radiotherapy-induced normal tissue complications be predicted from genetic profiles? Acta Oncol 44: 801–815
Article Google Scholar - Andreassen CN, Alsner J, Overgaard M, Overgaard J (2003) Prediction of normal tissue radiosensitivity from polymorphisms in candidate genes. Radiother Oncol 69: 127–135
Article CAS Google Scholar - Appleby JM, Barber JB, Levine E, Varley JM, Taylor AM, Stankovic T, Heighway J, Warren C, Scott D (1997) Absence of mutations in the ATM gene in breast cancer patients with severe responses to radiotherapy. Br J Cancer 76: 1546–1549
Article CAS Google Scholar - Badie C, Itzhaki JE, Sullivan MJ, Carpenter AJ, Porter AC (2000) Repression of CDK1 and other genes with CDE and CHR promoter elements during DNA damage-induced G(2)/M arrest in human cells. Mol Cell Biol 20: 2358–2366
Article CAS Google Scholar - Bakkenist CJ, Kastan MB (2003) DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 421: 499–506
Article CAS Google Scholar - Banin S, Moyal L, Shieh S, Taya Y, Anderson CW, Chessa L, Smorodinsky NI, Prives C, Reiss Y, Shiloh Y, Ziv Y (1998) Enhanced phosphorylation of p53 by ATM in response to DNA damage. Science 281: 1674–1677
Article CAS Google Scholar - Barber JB, Burrill W, Spreadborough AR, Levine E, Warren C, Kiltie AE, Roberts SA, Scott D (2000) Relationship between in vitro chromosomal radiosensitivity of peripheral blood lymphocytes and the expression of normal tissue damage following radiotherapy for breast cancer. Radiother Oncol 55: 179–186
Article CAS Google Scholar - Brock WA, Tucker SL, Geara FB, Turesson I, Wike J, Nyman J, Peters LJ (1995) Fibroblast radiosensitivity vs acute and late normal skin responses in patients treated for breast cancer. Int J Radiat Oncol Biol Phys 32: 1371–1379
Article CAS Google Scholar - Canman CE, Lim DS, Cimprich KA, Taya Y, Tamai K, Sakaguchi K, Appella E, Kastan MB, Siliciano JD (1998) Activation of the ATM kinase by ionizing radiation and phosphorylation of p53. Science 281: 1677–1679
Article CAS Google Scholar - Chedid M, Michieli P, Lengel C, Huppi K, Givol D (1994) A single nucleotide substitution at codon 31 (Ser/Arg) defines a polymorphism in a highly conserved region of the p53-inducible gene WAF1/CIP1. Oncogene 9: 3021–3024
CAS PubMed Google Scholar - Cole J, Arlett CF (1994) Cloning efficiency and spontaneous mutant frequency in circulating T-lymphocytes in ataxia-telangiectasia patients. Int J Radiat Biol 66: S123–S131
Article CAS Google Scholar - Correa CR, Cheung VG (2004) Genetic variation in radiation-induced expression phenotypes. Am J Hum Genet 75: 885–890
Article CAS Google Scholar - Garman KS, Nevins JR, Potti A (2007) Genomic strategies for personalized cancer therapy. Hum Mol Genet 16 (Spec No. 2): R226–R232
Article CAS Google Scholar - Geara FB, Peters LJ, Ang KK, Wike JL, Brock WA (1993) Prospective comparison of in vitro normal cell radiosensitivity and normal tissue reactions in radiotherapy patients. Int J Radiat Oncol Biol Phys 27: 1173–1179
Article CAS Google Scholar - Grace MB, McLeland CB, Blakely WF (2002) Real-time quantitative RT-PCR assay of GADD45 gene expression changes as a biomarker for radiation biodosimetry. Int J Radiat Biol 78: 1011–1021
Article CAS Google Scholar - Imbriano C, Gurtner A, Cocchiarella F, Di Agostino S, Basile V, Gostissa M, Dobbelstein M, Del Sal G, Piaggio G, Mantovani R (2005) Direct p53 transcriptional repression: in vivo analysis of CCAAT-containing G2/M promoters. Mol Cell Biol 25: 3737–3751
Article CAS Google Scholar - Jen KY, Cheung VG (2003) Transcriptional response of lymphoblastoid cells to ionizing radiation. Genome Res 13: 2092–2100
Article CAS Google Scholar - Johansen J, Bentzen SM, Overgaard J, Overgaard M (1996) Relationship between the in vitro radiosensitivity of skin fibroblasts and the expression of subcutaneous fibrosis, telangiectasia, and skin erythema after radiotherapy. Radiother Oncol 40: 101–109
Article CAS Google Scholar - Kruse JJ, Stewart FA (2007) Gene expression arrays as a tool to unravel mechanisms of normal tissue radiation injury and prediction of response. World J Gastroenterol 13: 2669–2674
Article CAS Google Scholar - Lee JH, Paull TT (2005) ATM activation by DNA double-strand breaks through the Mre11-Rad50-Nbs1 complex. Science 308: 551–554
Article CAS Google Scholar - Li G, Liu Z, Sturgis EM, Shi Q, Chamberlain RM, Spitz MR, Wei Q (2005) Genetic polymorphisms of p21 are associated with risk of squamous cell carcinoma of the head and neck. Carcinogenesis 26: 1596–1602
Article CAS Google Scholar - Lopez E, Guerrero R, Nunez MI, del Moral R, Villalobos M, Martinez-Galan J, Valenzuela MT, Munoz-Gamez JA, Oliver FJ, Martin-Oliva D, Ruiz de Almodovar JM (2005) Early and late skin reactions to radiotherapy for breast cancer and their correlation with radiation-induced DNA damage in lymphocytes. Breast Cancer Res 7: R690–R698
Article CAS Google Scholar - Morley M, Molony CM, Weber TM, Devlin JL, Ewens KG, Spielman RS, Cheung VG (2004) Genetic analysis of genome-wide variation in human gene expression. Nature 430: 743–747
Article CAS Google Scholar - Peacock J, Ashton A, Bliss J, Bush C, Eady J, Jackson C, Owen R, Regan J, Yarnold J (2000) Cellular radiosensitivity and complication risk after curative radiotherapy. Radiother Oncol 55: 173–178
Article CAS Google Scholar - Pim D, Banks L (2004) p53 polymorphic variants at codon 72 exert different effects on cell cycle progression. Int J Cancer 108: 196–199
Article CAS Google Scholar - Rieger KE, Chu G (2004) Portrait of transcriptional responses to ultraviolet and ionizing radiation in human cells. Nucleic Acids Res 32: 4786–4803
Article CAS Google Scholar - Rieger KE, Hong WJ, Tusher VG, Tang J, Tibshirani R, Chu G (2004) Toxicity from radiation therapy associated with abnormal transcriptional responses to DNA damage. Proc Natl Acad Sci USA 101: 6635–6640
Article CAS Google Scholar - Rodningen OK, Borresen-Dale AL, Alsner J, Hastie T, Overgaard J (2007) Radiation-induced gene expression in human subcutaneous fibroblasts is predictive of radiation-induced fibrosis. Radiother Oncol 86: 314–320
Article Google Scholar - Russell NS, Grummels A, Hart AA, Smolders IJ, Borger J, Bartelink H, Begg AC (1998) Low predictive value of intrinsic fibroblast radiosensitivity for fibrosis development following radiotherapy for breast cancer. Int J Radiat Biol 73: 661–670
Article CAS Google Scholar - Smilenov LB, Brenner DJ, Hall EJ (2001) Modest increased sensitivity to radiation oncogenesis in ATM heterozygous vs wild-type mammalian cells. Cancer Res 61: 5710–5713
CAS PubMed Google Scholar - Spielman RS, Bastone LA, Burdick JT, Morley M, Ewens WJ, Cheung VG (2007) Common genetic variants account for differences in gene expression among ethnic groups. Nat Genet 39: 226–231
Article CAS Google Scholar - Svensson JP, Stalpers LJ, Esveldt-van Lange RE, Franken NA, Haveman J, Klein B, Turesson I, Vrieling H, Giphart-Gassler M (2006) Analysis of gene expression using gene sets discriminates cancer patients with and without late radiation toxicity. PLoS Med 3: e422
Article Google Scholar - Tan XL, Popanda O, Ambrosone CB, Kropp S, Helmbold I, von Fournier D, Haase W, Sautter-Bihl ML, Wenz F, Schmezer P, Chang-Claude J (2006) Association between TP53 and p21 genetic polymorphisms and acute side effects of radiotherapy in breast cancer patients. Breast Cancer Res Treat 97: 255–262
Article CAS Google Scholar - Thomas M, Kalita A, Labrecque S, Pim D, Banks L, Matlashewski G (1999) Two polymorphic variants of wild-type p53 differ biochemically and biologically. Mol Cell Biol 19: 1092–1100
Article CAS Google Scholar - Toledo F, Wahl GM (2006) Regulating the p53 pathway: in vitro hypotheses, in vivo veritas. Nat Rev Cancer 6: 909–923
Article CAS Google Scholar - Tusher VG, Tibshirani R, Chu G (2001) Significance analysis of microarrays applied to the ionizing radiation response. Proc Natl Acad Sci USA 98: 5116–5121
Article CAS Google Scholar - Wang WD, Chen ZT, Li DZ, Cao ZH, Sun SL, Pu P, Chen XP (2005) Correlation between DNA repair capacity in lymphocytes and acute side effects to skin during radiotherapy in nasopharyngeal cancer patients. Clin Cancer Res 11: 5140–5145
Article CAS Google Scholar - Wang YA, Elson A, Leder P (1997) Loss of p21 increases sensitivity to ionizing radiation and delays the onset of lymphoma in atm-deficient mice. Proc Natl Acad Sci USA 94: 14590–14595
Article CAS Google Scholar - Watts JA, Morley M, Burdick JT, Fiori JL, Ewens WJ, Spielman RS, Cheung VG (2002) Gene expression phenotype in heterozygous carriers of ataxia telangiectasia. Am J Hum Genet 71: 791–800
Article Google Scholar - West CM, Davidson SE, Elyan SA, Valentine H, Roberts SA, Swindell R, Hunter RD (2001) Lymphocyte radiosensitivity is a significant prognostic factor for morbidity in carcinoma of the cervix. Int J Radiat Oncol Biol Phys 51: 10–15
Article CAS Google Scholar - Willems P, Claes K, Baeyens A, Vandersickel V, Werbrouck J, De Ruyck K, Poppe B, Van den Broecke R, Makar A, Marras E, Perletti G, Thierens H, Vral A (2008) Polymorphisms in nonhomologous end-joining genes associated with breast cancer risk and chromosomal radiosensitivity. Genes Chromosomes Cancer 47: 137–148
Article CAS Google Scholar - Yu J, Wang Z, Kinzler KW, Vogelstein B, Zhang L (2003) PUMA mediates the apoptotic response to p53 in colorectal cancer cells. Proc Natl Acad Sci USA 100: 1931–1936
Article CAS Google Scholar
Acknowledgements
We thank Wei Zhang for statistical analysis of QRT-PCR results, Ms Najla Al-Harbi for technical assistance in SNPs determination, Dr Colin Arlett for the AT cells, Dr Mike O’Donovan for T-cell culture advice and Richard Doull (MRC Harwell) for irradiations.
Author information
Author notes
- D Scott: Retired.
Authors and Affiliations
- Radiation Effects Department, Radiation Protection Division, Health Protection Agency, Centre for Radiation Chemical and Environmental Hazards, Chilton, Didcot, Oxfordshire, OX11 ORQ, UK
C Badie, S Dziwura, C Raffy, T Tsigani, J Moody, P Finnon & S Bouffler - Biomedical Physics Department, Radiation Biology Laboratory, King Faisal Specialist Hospital & Research Centre, Riyadh, 11211, Saudi Arabia
G Alsbeih - Clinical Oncology Department, Christie Hospital NHS Foundation Trust, Wilmslow Road, Manchester, M20 4BX, UK
E Levine - Cancer Genetics Department, Paterson Institute for Cancer Research, Christie Hospital NHS Foundation Trust, Manchester, M20 4BX, UK
D Scott
Authors
- C Badie
You can also search for this author inPubMed Google Scholar - S Dziwura
You can also search for this author inPubMed Google Scholar - C Raffy
You can also search for this author inPubMed Google Scholar - T Tsigani
You can also search for this author inPubMed Google Scholar - G Alsbeih
You can also search for this author inPubMed Google Scholar - J Moody
You can also search for this author inPubMed Google Scholar - P Finnon
You can also search for this author inPubMed Google Scholar - E Levine
You can also search for this author inPubMed Google Scholar - D Scott
You can also search for this author inPubMed Google Scholar - S Bouffler
You can also search for this author inPubMed Google Scholar
Corresponding author
Correspondence toC Badie.
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Badie, C., Dziwura, S., Raffy, C. et al. Aberrant CDKN1A transcriptional response associates with abnormal sensitivity to radiation treatment.Br J Cancer 98, 1845–1851 (2008). https://doi.org/10.1038/sj.bjc.6604381
- Received: 14 February 2008
- Revised: 07 April 2008
- Accepted: 07 April 2008
- Published: 20 May 2008
- Issue Date: 03 June 2008
- DOI: https://doi.org/10.1038/sj.bjc.6604381