The Role of the Dorsomedial Prefrontal Cortex, Basolateral Amygdala, and Dorsal Hippocampus in Contextual Reinstatement of Cocaine Seeking in Rats (original) (raw)
INTRODUCTION
Relapse is a major impediment in the treatment of cocaine dependence. Drug-associated environments, discrete conditioned stimuli (CSs, eg, paraphernalia), and exposure to cocaine elicit craving and relapse in drug users (Jaffe et al, 1989; Ehrman et al, 1992; Foltin and Haney, 2000). Similarly, cocaine-paired contextual stimuli (Rauhut et al, 2000; Alleweireldt et al, 2001; Crombag et al, 2002), explicit CSs (de Wit and Stewart, 1981), and cocaine priming injections (Gerber and Stretch, 1975) elicit cocaine-seeking behavior, an index of incentive motivation for cocaine in rats.
Cocaine-seeking behavior elicited by drug-paired contextual stimuli, explicit CSs, and cocaine appears to be identical; however, different cognitive mechanisms likely mediate the motivational effects of these stimuli. During drug self-administration training, explicit CSs are presented response contingently and consistently paired with a drug reinforcer. Thus, explicit CSs acquire the ability to elicit drug seeking by signaling imminent drug reinforcement or by acting as conditioned reinforcers. In contrast, contexts are comprised of static multimodal background stimuli, which constitute a setting where response–drug and/or CS–drug associations form. Consequently, contexts elicit drug seeking primarily by functioning as ‘occasion setters,’ discriminative stimuli (SD) which signal response-contingent drug availability (Gordon and Klein, 1994). Lastly, cocaine is thought to produce drug seeking by directly stimulating the brain reward system (for review see, Shalev et al, 2002).
Given the putative differences in the informational content of drug-paired contexts, explicit CSs, and cocaine, separate brain systems likely mediate their motivational effects. Previous research indicates that the dorsomedial prefrontal cortex (dmPFC), basolateral amygdala (BLA), and hippocampus exhibit neuronal activation concomitant with cocaine-seeking behavior in a cocaine-paired environment (Neisewander et al, 2000), and lesions of these structures impair place conditioning, a task dependent on context-reward learning (Tzschentke and Schmidt, 1999; Ferbinteanu and McDonald, 2001; Fuchs et al, 2002b; Meyers et al, 2003). BLA or dmPFC inactivation also impairs explicit CS-induced reinstatement of cocaine seeking (McLaughlin and See, 2003), a behavior based on explicit CS-reward associations (Kruzich and See, 2001). Furthermore, dmPFC inactivation impairs, whereas BLA inactivation fails to alter, cocaine-primed reinstatement of cocaine-seeking behavior (McFarland and Kalivas, 2001). Unlike the BLA and dmPFC, the dorsal hippocampus (DH) does not play a critical role in explicit CS–US associations according to the fear conditioning literature (Kim and Fanselow, 1992); however, its involvement in explicit CS-induced and cocaine-primed reinstatement of cocaine-seeking behavior has yet to be examined. Importantly, it remains to be investigated whether the functional integrity of these structures is necessary for contextual reinstatement of cocaine-seeking behavior.
The primary objective of the present study was to test the hypothesis that distinct, albeit likely overlapping, neural circuitries mediate the ability of contextual SDs vs explicit CSs and cocaine to elicit cocaine-seeking behavior. To this end, following drug self-administration training and extinction, the dmPFC, BLA, or DH were functionally inactivated using the sodium channel blocker, tetrodotoxin, and resulting alterations in cocaine-seeking behavior were assessed in a cocaine-paired context or a non-cocaine-paired, alternate context. Effects of DH inactivation on explicit CS- and cocaine-induced reinstatement were also assessed. It was postulated that BLA or dmPFC inactivation would impair contextual reinstatement of cocaine-seeking behavior and DH inactivation would selectively impair contextual, but not explicit CS- or cocaine-induced, reinstatement.
MATERIALS AND METHODS
Animals
Male Sprague–Dawley rats (Charles-River, _N_=87), weighing 300–325 g at the start of the experiment, were individually housed in a temperature and humidity controlled vivarium on a reversed light–dark cycle. Rats were maintained on 20–25 g of rat chow per day, with water available ad libitum. The housing and treatment of the rats followed the guidelines of the ‘Guide for the Care and Use of Laboratory Rats’ (Institute of Laboratory Animal Resources on Life Sciences, National Research Council, 1996). Rats were given 5 days for adaptation before the start of the experiment.
Food Training
Rats were trained to lever press on a fixed ratio (FR) 1 schedule of food reinforcement (45 mg pellets; Noyes, Lancaster, NH) in sound-attenuated operant conditioning chambers (30 × 20 × 24 cm high; Med Associates Inc., St Albans, VT) during a 16-h overnight food training session. The chambers were equipped with two retractable levers, a stimulus light above each lever, a food pellet dispenser between the levers, a house light on the wall opposite to the levers, and a speaker connected to a tone generator (ANL-926, Med Associates). During the session, the stimuli subsequently used for contextual or explicit cue conditioning were not presented. In the standard operant conditioning chamber, each lever press on the active lever resulted in delivery of a food pellet only. Lever presses on the inactive lever had no programmed consequences. Following food training, food pellet dispensers were removed from the chambers.
Surgery
At 48 h after food training, rats were anesthetized using ketamine hydrochloride (66.6 mg/kg, IP; Fort Dodge Animal Health, Fort Dodge, IA) and xylazine (1.3 mg/kg, IP; Bayer, Shawnee Mission, KS) followed by equithesin (1 ml/kg of a solution of 9.72 mg/ml pentobarbital sodium, 42.5 mg/ml chloral hydrate, and 21.3 mg/ml magnesium sulfate heptahydrate dissolved in a 44% propylene glycol, 10% ethanol solution; IP). Chronic indwelling catheters were constructed using a bent steel cannula with a screw-type connector (Plastics One, Roanoke, VA), SILASTIC tubing (10 cm, i.d. 0.64 mm, o.d. 1.19 mm, Dow Corning, Midland, MI), Marlex mesh, and cranioplastic cement, as described previously (See et al, 2001). The end of the catheter was inserted into the right jugular vein and was secured to surrounding tissue with suture. The catheter ran subcutaneously and exited on the rat's back, posterior to the shoulder blades.
Immediately after the catheter surgery, the rats were placed into a stereotaxic instrument (Stoelting, Wood Dale, IL). They were implanted with bilateral stainless-steel guide cannulae (26 gauge, Plastics One) aimed at the BLA, dmPFC, DH or their respective cortical anatomical control brain regions, the barrel field region of the somatosensory cortex (SSbf), the ventromedial prefrontal cortex (vmPFC), or the trunk region of the somatosensory cortex (SStr), using standard stereotaxic procedures (BLA: −2.5 mm AP, ±5.0 mm ML, −6.6 mm DV; dmPFC: +3.0 mm AP, ±0.6 mm ML, −1.4 mm DV; DH: cannulae angled at 15°: −3.1 mm AP, ±3.1 mm ML, −2.1 mm DV; SSbf: −2.5 mm AP, ±5.0 mm ML, −2.3 mm DV; vmPFC: +3.0 mm AP, ±0.6 mm ML, −4.2 mm DV; SStr, cannulae angled at 15°: −3.1 mm AP, ±3.1 mm ML, −1.0 mm DV, relative to bregma). These anatomical control brain regions are in the same anterior–posterior and medial–lateral planes as the target brain regions and are adjacent to the target brain regions with the exception of the SSbf. The SSbf was chosen over more ventral brain regions as the anatomical control region for the BLA in order to minimize the inactivation of the external capsule. Three small screws and cranioplastic cement secured the guide cannulae to the skull. Stylets (Plastics One) were placed into the guide cannulae and catheter to prevent occlusion. To extend catheter patency, the catheters were flushed twice daily for 5 days following surgery with 0.1 ml of an antibiotic solution of cefazolin (10.0 mg/ml, Schein Pharmaceutical, Florham Park, NJ) dissolved in heparinized saline (70 U/ml; Elkins-Sinn, Cherry Hill, NJ). Thereafter, catheters were flushed with 0.1 ml heparinized saline (10 U/ml) prior to each self-administration session, and with 0.1 ml of the cefazolin solution and 0.1 ml of heparinized saline (70 U/ml) after each session. Catheter patency was periodically verified by infusing 0.08–0.10 ml of methohexital sodium (20 mg/ml, IV; Eli Lilly and Co., Indianapolis, IN), which produces a rapid loss of muscle tone only when administered intravenously.
Self-Administration
Self-administration training was conducted during 2-h sessions on 10 consecutive days during the rats' dark cycle. Rats were trained to press a lever according to an FR 1 schedule of cocaine reinforcement (cocaine hydrochloride; 0.20 mg/0.05 ml; National Institute on Drug Abuse, Research Triangle Park, NC) with a 40-s time out period following each reinforcer. The catheters were connected to liquid swivels (Instech, Plymouth Meeting, PA) via polyethylene 20 tubing (Dow Corning Co., Midland, MI) that was encased in steel spring leashes (Plastics One). The swivels were suspended above the operant conditioning chamber and were connected to infusion pumps (Model PHM-100, Med Associates) with Tygon tubing (Performance Plastics, Akron, OH). Data collection and reinforcer delivery were controlled using Schedule Manager for Windows software version 2.09 (Med Associates).
Intracranial Infusions
For intracranial infusions, stainless-steel injection cannulae (33 gauge, Plastics One) were inserted to a depth of 1 mm (dmPFC, DH, vmPFC, SStr) or 2 mm (BLA, SSbf) below the tip of the guide cannulae immediately prior to placement into the chamber. The injection cannulae were connected to 10-μl Hamilton syringes (Hamilton Co., Reno, NV) that were mounted on an infusion pump (Harvard Apparatus, South Natick, MA). TTX (5.0 ng/side) or phosphate-buffered saline vehicle (pH=7.0 for both) were infused bilaterally at a volume of 0.5 μl/hemisphere over 2 min. The injection cannulae were left in place for 1 min prior to and after the infusion. TTX acts by inhibiting the generation and propagation of action potentials (for a critical review of this and other temporary inactivation procedures, see Martin and Ghez, 1999). Metabolic activation and other physiological studies have shown that the mean inactivated tissue radius after infusion of 5 ng/0.5 μl of TTX is ∼0.7–0.8 mm, and the maximal effect of TTX lasts for no less than 120 min, then disappears exponentially within 24 h (Cahill et al, 1987; Zhuravin and Bures, 1991; Ambrogi Lorenzini et al, 1995). Thus, subjects can be used as their own controls. Using an infusion protocol and coordinates identical to those utilized in the present experiments, we have been able to establish functional dissociation between the dmPFC and vmPFC (McLaughlin and See, 2003), as well as between the BLA and the SSbf (McLaughlin and See, 2003) and central nucleus of the amygdala (Kruzich and See, 2001), suggesting that TTX administration resulted in selective inactivation of the target structures in the present study.
Experiment 1: Effects of BLA, dmPFC, DH, SSbf, vmPFC, or SStr Inactivation on Contextual Reinstatement of Cocaine-Seeking Behavior
Previous studies using the renewal procedure have demonstrated that Pavlovian and operant conditioned responses to an explicit CS can be reinstated when a subject is returned to the conditioning context after extinction in another context, suggesting that context acts as an occasion setter (Bouton and Bolles, 1979; Bouton and Swartzentruber, 1986; Bouton and Ricker, 1994; Goddard, 1999; Nakajima et al, 2000). Similarly, it has been shown that re-exposure to a drug-associated context is sufficient to reinstate drug-seeking behavior in the response-contingent presence (Crombag et al, 2002; Crombag and Shaham, 2002) or absence (Rauhut et al, 2000; Alleweireldt et al, 2001) of a drug-paired explicit CS. However, the neural substrates that mediate context-induced reinstatement of drug seeking have not been identified. Thus, Experiment 1 examined the involvement of the BLA, dmPFC, and DH in context-induced reinstatement of cocaine seeking.
Stimulus presentation during self-administration training
Rats (_N_=53) were trained to self-administer cocaine in operant conditioning chambers during 2-h daily sessions. The chambers contained one of two distinctly different sets of visual, auditory, olfactory, and tactile contextual stimuli (see Table 1). The contextual stimuli were presented throughout each session independent of responding. Active lever presses resulted in a 2-s activation of the infusion pump only. After each infusion, responses on the active lever were counted but had no consequences during the 40-s time-out period. During the sessions, responses on the inactive lever had no programmed consequences, but were recorded.
Table 1 Contextual Stimuli Used in Experiment 1
Extinction
Rats underwent 2-h extinction sessions on at least 10 consecutive days, during which responses on either lever had no programmed consequences. Extinction sessions were conducted either in the self-administration context (alternate context group; _N_=27) or in the alternate context (cocaine context group; _N_=52, see Table 2). Assignment of the BLA-, dmPFC-, and DH-cannulated rats to the alternate and cocaine context groups was counterbalanced based on cocaine intake during self-administration, whereas in the follow-up experiments, all SSbf-, vmPFC-, and SStr-cannulated rats received extinction training in the alternate context. To examine the effects of the intracranial infusion procedure on responding, rats received a sham intracranial infusion prior to placement into the chamber on extinction day 7. During the sham procedure, injection cannulae were bilaterally inserted into the rat's guide cannulae and were left in place for 4 min, but fluid was not infused through the infusion cannulae. Extinction sessions were terminated when the rats reached the criterion for extinction (ie, ⩽25 active lever responses/session on 2 consecutive days).
Table 2 Contextual Stimulus Presentation During the Three Phases of Experiment 1
Contextual reinstatement test
On the test days, rats received bilateral intracranial microinfusions of TTX (5.0 ng/0.5 μl/side). or phosphate-buffered saline vehicle (0.5 μl/side) over 2 min immediately before reinstatement testing. Rats that had received microinfusions into the BLA, dmPFC, or DH were then placed into the self-administration context (cocaine context group) or an alternate context (alternate context group, see Table 2) for a 2-h test session. All rats that had received microinfusions into the anatomical control brain regions (ie SSbf, vmPFC, or SStr) were placed into the self-administration context on the reinstatement test days. This design resulted in the following groups: BLA cocaine context (_N_=9), BLA alternate context (_N_=9), dmPFC cocaine context (_N_=7), dmPFC alternate context (_N_=10), DH cocaine context (_N_=10), DH alternate context (_N_=8), SSbf cocaine context (_N_=8), vmPFC cocaine context (_N_=10), SStr cocaine context (_N_=8). The order of TTX and vehicle infusions was counterbalanced across two test sessions based on the history of cocaine intake during self-administration. During testing, lever presses were recorded but had no scheduled consequences. Between the two test sessions, the rats underwent additional extinction sessions in the self-administration context (alternate context group) or the alternate context (cocaine context group) until they reached the criterion for extinction described above.
Locomotor activity test
Studies from our laboratory (McLaughlin and See, 2003) have demonstrated that TTX microinfusions into the BLA do not significantly alter general activity during a 1-h locomotor activity test, suggesting that general motor deficits do not account for the effects of BLA inactivation on reinstatement. To further assess possible motor impairing effects of intracranial TTX infusions, we examined the effects of TTX infused into the dmPFC, DH, vmPFC, SStr, or SSbf on general activity.
Four days following the last reinstatement test, locomotor activity was measured in novel Plexiglas chambers (40.8 cm W × 40.8 cm L × 37.5 cm H) equipped with an array of eight photodetectors and corresponding light sources that emitted photobeams 8 cm apart at 4.5 cm above the chamber floor. A computerized activity system (San Diego Instruments, San Diego, CA) recorded the number of times neighboring photobeams were broken consecutively by a rat moving in the chamber during a 1-h test session. Prior to placement into the apparatus, rats received bilateral intracranial infusions of TTX or vehicle into the dmPFC, DH, vmPFC, SStr, or SSbf using the procedures described above. Assignment to TTX vs vehicle infusions was randomized. Data from nine additional rats with correct cannula placement (vmPFC, _N_=2; SSbf, _N_=4; SStr, _N_=3) was included only in the locomotion study because of technical problems that disrupted contextual conditioning. The resulting group sizes for the locomotor study were as follows: dmPFC vehicle (_N_=5), dmPFC TTX (_N_=5), DH vehicle (_N_=6), DH TTX (_N_=5), vmPFC vehicle (_N_=6), vmPFC TTX (_N_=6), SSbf vehicle (_N_=6), SSbf TTX (_N_=6), SStr vehicle (_N_=6), and SStr TTX (_N_=6).
Experiment 2: Effects of Dorsal Hippocampus Inactivation on Explicit CS-Induced and Cocaine-Primed Reinstatement of Cocaine Seeking
Previous research from our laboratory has demonstrated that inactivation of the dmPFC or BLA impairs explicit CS-induced reinstatement of extinguished cocaine-seeking behavior (McLaughlin and See, 2003). Furthermore, inactivation of the dmPFC, but not of the BLA, abolishes cocaine-primed reinstatement of extinguished cocaine-seeking behavior (McFarland and Kalivas, 2001). It has not been examined, however, whether the functional integrity of the DH is necessary for explicit CS-induced or cocaine-induced relapse to cocaine seeking. In order to facilitate comparison of the neural circuitries that mediate various forms of relapse to drug seeking, Experiment 2 examined the effects of DH inactivation on explicit CS-induced and cocaine-primed reinstatement of extinguished cocaine-seeking behavior.
Stimulus presentation during self-administration training
Rats (_N_=8) were trained to self-administer cocaine in standard operant conditioning chambers during 2-h daily sessions. The house light was illuminated throughout each session. Lever presses on the active lever resulted in a 2-s activation of the infusion pump and a 5-s presentation of a CS complex, consisting of activation of the white stimulus light above the active lever and the tone generator (78 dB, 2kHz). After each infusion, responses on the active lever were counted but had no consequences during the 40-s time-out period. During the sessions, responses on the inactive lever were recorded but had no programmed consequences.
Extinction
Rats underwent 2-h extinction sessions on at least 10 consecutive days, during which responses on either lever were recorded but had no programmed consequences. Extinction sessions were terminated when the rats reached the criterion for extinction (ie, ⩽25 active lever responses/session on 2 consecutive days). To examine the effects of the intracranial infusion procedure on responding, rats received a sham intracranial infusion prior to placement into the chamber on extinction day 7, using the sham procedure described above.
Explicit CS-induced reinstatement test
Two reinstatement tests were conducted to examine the effects of DH inactivation on the ability of the light-tone CS complex to reinstate responding. On the test days, rats first received infusions of TTX or vehicle into the DH. Rats were then placed into the chambers for a 2-h session, during which responses resulted in 5-s presentations of the light-tone CSs in the absence of cocaine reinforcement. Each CS presentation was followed by a 40-s time-out period during which responses had no programmed consequences. The order of TTX vs vehicle infusions was counterbalanced across the two test days based on cocaine intake. Prior to the second test day, rats underwent additional extinction sessions until they exhibited ⩽25 active lever responses per session on 2 consecutive days.
Cocaine-primed reinstatement test
Following the second explicit CS-induced reinstatement test, rats underwent additional extinction sessions until they reached the extinction criterion described above. Subsequently, two tests were conducted to examine the effects of DH inactivation on the ability of a cocaine priming injection to reinstate responding. On the test days, rats first received TTX or vehicle infusions into the DH and a cocaine priming injection (10 mg/kg, i.p.). Rats were then placed into the chambers for a 2-h extinction session, during which the light-tone CSs were not presented and responses had no programmed consequences. The 10 mg/kg priming dose of cocaine was selected based on previous studies showing that it produces maximal reinstatement of cocaine seeking (Cornish et al, 1999). The order of TTX vs vehicle infusions was counterbalanced across the two test days based on cocaine intake, and was either the same or reversed as that during the explicit CS-induced reinstatement tests. Prior to the second cocaine reinstatement test day, rats underwent additional extinction sessions until they reached the extinction criterion.
Histology
Rats were transcardially perfused with phosphate-buffered saline and 10% formaldehyde solution. The brains were dissected out and were stored in 10% formaldehyde solution until sectioning. Brains were sectioned in the coronal plane at a thickness of 75 μm on a vibratome (Technical Products International, Inc., St Louis, MO). Sections containing cannula tracks were mounted on gelatin-coated slides and were stained with cresyl violet (Kodak, Rochester, NY). The sections were examined under a light microscope to determine cannula placement. The most ventral point of each cannula track was mapped onto schematics of the appropriate plates from the rat brain atlas of Paxinos and Watson (1997).
Data analysis
Mixed-factor and repeated measures analyses of variance (ANOVA) were used to analyze responses on the active and inactive levers with lever (active, inactive) and day (self-administration, extinction, TTX reinstatement day, vehicle reinstatement day) as within-subjects factors, and group (alternate context group, cocaine-paired context group) as a between-subjects factor, where appropriate. Statistically significant interaction effects were further investigated using Tukey HSD post hoc tests. Planned one-tail _t_-tests were used to compare responding during the explicit CS- and cocaine-induced reinstatement tests to responding during self-administration training and to compare responding during the cocaine-induced reinstatement tests to the extinction baseline.
RESULTS
Histology
Representative photomicrographs and/or schematics illustrating the distribution of injection cannula placements in the brains of rats from Experiments 1 and 2 are included in Figures 1 and 2, respectively. The most ventral point of the injection cannula tracks was located within the target region bilaterally for all rats whose data were included in data analysis and in the _N_s reported above. The target regions were defined as follows: dmPFC, the Cg 1 region and the Cg 1/Cg 2 transition area of the medial prefrontal cortex; vmPFC, the ventral Cg 2, ventromedial orbitofrontal, and infralimbic areas of the ventromedial prefrontal cortex, BLA, the lateral/basolateral nuclei of the amygdala; SSbf, the barrel fields region of the somatosensory cortex that overlies the basolateral amygdala; DH, the dorsal hippocampus; SStr, the trunk region of the somatosensory cortex that overlies the dorsal hippocampus. The vmPFC, SSbf, and SStr were selected as anatomical controls regions because they are either adjacent and/or dorsal (ie, in the path of TTX diffusion) relative to the brain regions of primary interest (ie, dmPFC, BLA, and DH, respectively). High-power microscopy of the infused area did not reveal any abnormalities in neuronal integrity.
Figure 1
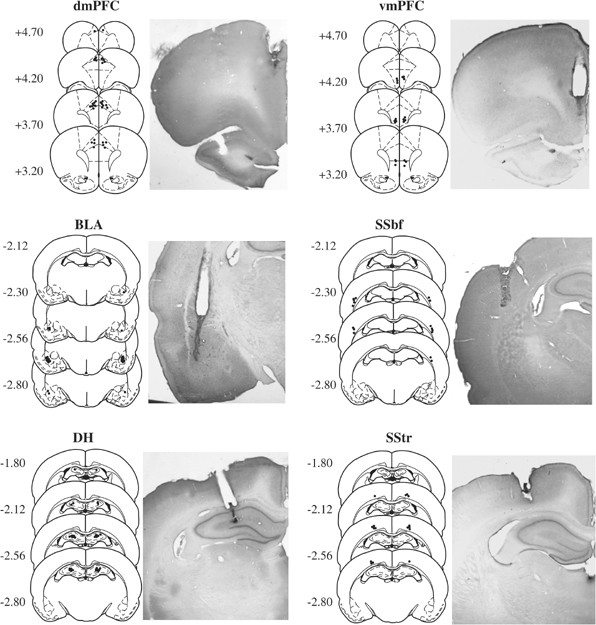
Microinfusion cannula placement as verified on cresyl violet-stained sections and representative photomicrographs of each brain region of interest (left) and its respective anatomical control brain region (right) in Experiment 1. The symbols represent the most ventral point of the infusion cannula tract for each rat on coronal sections based on the atlas of Paxinos and Watson (1997). Numbers indicate the distance from bregma in millimeters.
Figure 2
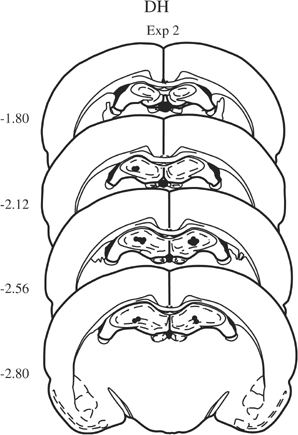
Microinfusion cannula placement within the DH as verified on cresyl violet-stained sections in Experiment 2. The symbols represent the most ventral point of the infusion cannula tract for each rat on coronal sections based on the atlas of Paxinos and Watson (1997). Numbers indicate the distance from bregma in millimeters.
Experiment 1
Self-administration and extinction
Rats exhibited stable responding on the active lever during the last 7 self-administration days with a within-subject variability of less than 10% in daily cocaine intake. There was no difference in cocaine intake between the groups with guide cannulae aimed at the dmPFC, BLA, DH, vmPFC, SSbf, or SStr (F(5,73)=0.46, _P_=0.80). The mean daily cocaine intake (±SEM) was 31.81±1.3 infusions (approx. 20.05±1.10 mg/kg/session).
Responding declined upon removal of cocaine reinforcement and extinguished to criterion (⩽25 responses/day per two consecutive days) prior to reinstatement testing (Figure 3) independent of the context in which extinction training occurred (data not shown). The effects of context on extinction training were examined in DH-, BLA-, and dmPFC-cannulated rats. SStr-, SSbf-, or vmPFC-cannulated rats were not included in these analyses because none of these subjects underwent extinction training in the previously cocaine-paired context. Separate 2 × 7 mixed factor ANOVAs of active lever presses indicated that responding declined across extinction sessions (F(6,90–96)=15.00–16.87, P<0.0001). Extinction responding was not significantly altered by whether the rats underwent extinction training in the previously cocaine-paired or in the alternate context (context main effect: F(1,15–16)=0.31–1.47, _P_=0.59–0.24; context by day interaction effect: F(6,90–96)=0.85–1.35, _P_=0.53–0.24). There was also no effect of context on the total number of days needed to reach the extinction criterion prior to the first or second reinstatement tests (t(15–16)=0.284–1.865, _P_=0.08–0.78). The mean number of days (±SEM) needed for rats to reach the extinction criterion was 11.58±0.38 and 2.68±0.23 days prior to the first and second reinstatement test days, respectively. Furthermore, sham intracranial infusions on extinction day 7 did not alter extinguished responding (data not shown).
Figure 3
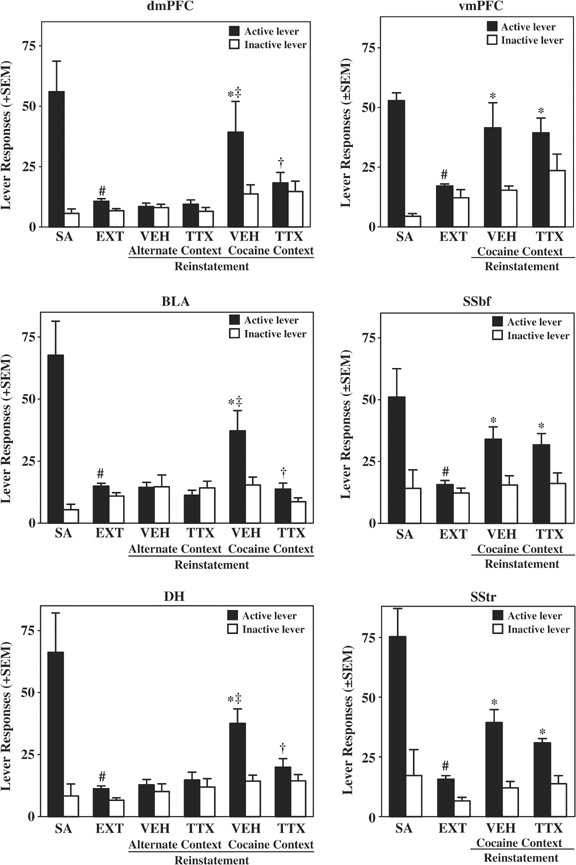
Responses on the active and inactive levers (±SEM) during self-administration (SA; average of the last 3 days), extinction (EXT; average of the last extinction day preceding the TTX reinstatement test day and the last extinction day preceding the vehicle reinstatement test day), and contextual reinstatement testing. On the test day, rats received intracranial pretreatment with TTX or vehicle and were then exposed to the previously cocaine-paired context (_N_=7–10/brain region) or the alternate context (_N_=8–10/brain region) in the absence of cocaine reinforcement or response-contingent stimulus presentation. Symbols represent significant difference relative to responding during self-administration (#, ANOVA day simple main effects test, P<0.05), during extinction (*, Tukey test, P<0.05; SSbf and vmPFC, _t-_test, P<0.05), upon exposure to the alternate context (‡, Tukey test, P<0.05), and relative to vehicle pretreatment (†, Tukey test, P<0.05).
Contextual reinstatement of extinguished cocaine-seeking behavior
Exposure to the cocaine-paired context reinstated extinguished cocaine-seeking behavior, whereas exposure to the alternate context failed to reinstate cocaine-seeking behavior. Furthermore, TTX inactivation of the BLA, dmPFC, or DH disrupted the ability of the cocaine-paired context to reinstate extinguished cocaine-seeking behavior (Figure 3 left), whereas inactivation of the adjacent SSbf, vmPFC, or SStr failed to alter responding (Figure 3 right). There was no difference in lever responding on the extinction days that immediately preceding the TTX and vehicle reinstatement test days; therefore, extinction data were collapsed across these time points (extinction, henceforth).
For the BLA, dmPFC, and DH, separate 3 × 2 × 2 mixed factor ANOVAs of lever presses indicated statistically significant three-way day by group by lever interaction effects (BLA, F(2,32)=5.46, P<0.01; dmPFC, F(2,30)=6.37, P<0.005; DH, F(2,32)=7.62, P<0.005). Subsequent pairwise comparisons revealed that in vehicle-pretreated rats, exposure to the cocaine-paired context produced a significant increase in responding on the active lever relative to extinction (Tukey test, P<0.05) and relative to the inactive lever (Tukey test, P<0.05). Furthermore, exposure to the cocaine-paired context elicited more responding on the active lever relative to exposure to the alternate context (Tukey test, P<0.05). Exposure to the alternate context failed to elicit an increase in responding on the active lever relative to the extinction baseline or to responding on the inactive lever. Pretreatment with TTX into the BLA, dmPFC, or DH blocked reinstatement of responding in the cocaine-paired context (Tukey test, P<0.05), whereas it did not alter responding in the alternate context. Pairwise comparisons of responses on the inactive lever did not reveal any significant day or group effects.
For the SSbf and vmPFC, separate 3 × 2 mixed factor ANOVAs of lever presses indicated significant main effects of day (SSbf: F(2,14)=5.66, P<0.05; vmPFC, F(2,18)=4.09, _P_<0.5) and lever (SSbf: F(1,7)=21.01, _P_<0.005; vmPFC, F(1,9)=12.68, _P_<0.01), but no interaction effects. However, planned comparisons indicated that exposure to the cocaine-paired context produced a significant increase in responding on the active lever relative to extinction (SSbf: _t_(7)=3.54, _P_<0.01; vmPFC, _t_(9)=2.26, _P_=0.05). Furthermore, pretreatment with TTX into the SSbf or vmPFC failed to alter reinstatement of responding in the cocaine-paired context relative to saline pretreatment (SSbf: _t_(7)=0.25, _P_=0.81; vmPFC, _t_(9)=0.15, _P_=0.88). Planned comparisons of responses on the inactive lever did not reveal any significant day effects. For the SStr, the 3 × 2 mixed factor ANOVA of lever presses indicated a statistically significant two-way day by lever interaction effect (SStr: F(2,14)=1.58, _P_<0.05). Subsequent pairwise comparisons revealed that in vehicle-pretreated rats, exposure to the cocaine-paired context produced a significant increase in responding on the active lever relative to extinction (Tukey test, _P_<0.05) and relative to the inactive lever (Tukey test, _P_<0.05). However, pretreatment with TTX into the SStr failed to alter the reinstatement of responding in the cocaine-paired context (Tukey test, _P_>0.05). Pairwise comparisons of responses on the inactive lever did not reveal any significant day effects.
Experiment 2
Self-administration and extinction
Similar to Experiment 1, rats exhibited stable responding on the active lever during the last 7 self-administration days with a within-subject variability of less than 10% in daily cocaine intake. The mean daily cocaine intake (±SEM) was 32.63±2.21 infusions (approx. 20.88±1.42 mg/kg/session). There was no difference in cocaine intake between the explicit cue-trained rats of Experiment 2 and the DH group of Experiment 1 (t(24)=0.21, _P_>0.5). Responding declined upon removal of cocaine reinforcement (F(6,42)=9.01, P<0.0001; Tukey test, P<0.05) and extinguished to criterion prior to reinstatement testing (Figure 4). Furthermore, sham infusions into the DH on extinction day 7 did not reinstate extinguished responding.
Figure 4
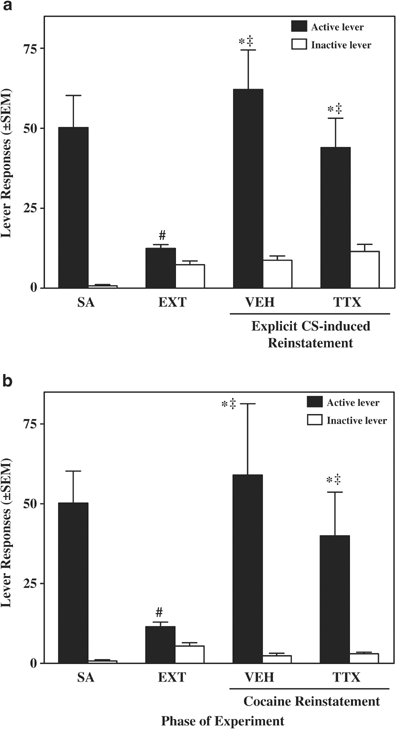
Responses on the active and inactive levers (±SEM) during self-administration (SA; average of the last 3 days), extinction (EXT; average of the last extinction day preceding the TTX reinstatement test day and the last extinction day preceding the vehicle reinstatement test day), and reinstatement testing. On the explicit CS-induced reinstatement test day (a), rats (_N_=8) received an infusion of TTX or vehicle into the DH, and responses on the active lever resulted in presentations of the light-tone stimulus complex in the absence of cocaine reinforcement. On the cocaine reinstatement test day (b), rats (_N_=8) received an infusion TTX or vehicle into the DH and a systemic infusion of cocaine (10 mg/kg, i.p.); responses had no programmed consequences. Symbols represent significant difference relative to responding during self-administration (#, ANOVA day simple main effects test, P<0.05), during extinction (*a: Tukey test, P<0.05; b: planned _t_-test, P<0.5), and relative to the inactive lever (‡, a: Tukey test, P<0.05; b: planned _t_-test, P<0.5).
Explicit cue-induced reinstatement of extinguished cocaine-seeking behavior
Response-contingent presentation of the light-tone CSs elicited robust reinstatement of responding on the active lever (Figure 4). Furthermore, a comparison of active lever responding exhibited by rats that received vehicle infusions into the DH in Experiments 1 and 2 revealed that the time course of lever pressing was similar in response to the light-tone CSs vs the cocaine-paired context, even though response-contingent presentation of the light-tone CSs elicited more responding overall (data not shown).
TTX inactivation of the DH failed to alter the ability of the light-tone CSs to reinstate responding (Figure 4a). There was no significant difference in responding during the extinction days that immediately preceded the vehicle and TTX reinstatement test days; therefore, extinction data were collapsed across these time points (extinction, henceforth). The 3 × 2 repeated measures ANOVA of lever presses during the reinstatement test days and extinction indicated a significant day by lever interaction effect (F(2,14)=9.11, P<0.005), and significant main effects of day (F(2,14)=11.09, _P_<0.001) and lever (F(1,7)=22.36, _P_<0.005). Subsequent pairwise comparisons revealed that response-contingent presentation of the light-tone CSs following vehicle or TTX infusions into the DH produced a significant increase in responding on the active lever relative to responding during extinction (Tukey test, _P_<0.05) and relative to the inactive lever (Tukey test, _P_<0.05). Thus, TTX pretreatment infused into the DH failed to alter the reinstatement in responding on the active lever elicited by the light-tone CSs relative to vehicle (Tukey test, _P_>0.5). Furthermore, TTX pretreatment infused into the DH failed to alter responding on the active lever during the reinstatement test relative to responding during self-administration (t(7)=0.37, _P_>0.05). Pairwise comparisons of responses on the inactive lever did not reveal a day effect. The effects of intra-DH infusions of TTX were not examined on extinction responding in the absence of cue presentation. However, given that inactivation of the DH failed to alter locomotion (see below) and operant responding on the inactive lever (see above), it is unlikely that the observed lack of an effect on active lever responding was due to nonspecific effects on general activity.
Cocaine-primed reinstatement of extinguished cocaine-seeking behavior
TTX inactivation of the DH failed to alter the ability of the cocaine priming injection to reinstate responding (Figure 4b). Cocaine-primed reinstatement of responding was not due to the stress of intraperitoneal drug administration since intraperitoneal administration of saline alone or in combination with intracranial administration of saline fails to reinstate responding (Neisewander et al, 2000; Fuchs et al, 2002a; Fuchs and See, 2002; Ledford et al, 2003).
There was no significant difference in responding during the extinction days that immediately preceded the vehicle and TTX reinstatement test days; therefore, extinction data were collapsed across these time points (extinction, henceforth). The 3 × 2 repeated measures ANOVA of lever presses during the reinstatement test days and extinction indicated a significant main effect of lever (F(1,14)=17.326, P<0.005) and a trend for a day by lever interaction effect (F(2,14)=2.64, _P_=0.10), but no day main effect (F(2,14)=1.89, _P_=0.19). Subsequent planned comparisons indicated that cocaine priming following vehicle or TTX infusions into the DH produced a significant increase in responding on the active lever relative to responding during extinction (Vehicle, one-tail _t_(7)=2.07, _P_<0.05; TTX, one-tail _t_(7)=2.13, _P_<0.05) and relative to the inactive lever (Vehicle, _t_(7)=2.53, _P_<0.05; TTX, _t_(7)=2.51, _P_<0.05). Thus, TTX pretreatment infused into the DH failed to alter the reinstatement in responding on the active lever elicited by cocaine priming relative to vehicle (_t_(7)=0.67, _P_>0.05). Furthermore, TTX pretreatment infused into the DH failed to alter responding on the active lever during the reinstatement test relative to responding during self-administration (t(7)=0.59, _P_>0.05). Given that inactivation of the DH failed to alter locomotion (see below) and operant responding on the inactive lever (see above), it is unlikely that the observed lack of an effect on active lever responding was due to nonspecific effects on general activity.
Locomotor activity
TTX infused into the dmPFC, DH, vmPFC, SStr, or SSbf did not alter locomotor activity (Figure 5), similar to findings obtained following infusion of TTX into the BLA (McLaughlin and See, 2003). Separate 2 × 6 mixed factor ANOVAs of photobeam breaks indicated significant main effects of time (dmPFC, F(5,40)=39.89,. P<0.0001; DH, F(5,45)=43.58, P<0.0001; vmPFC, F(5,50)=36.02, P<0.0001; SStr, F(5,45)=36.47, P<0.0001; SSbf, F(5,50)=21.78, P<0.0001), but no significant main effects of group or time by group interaction effects. Thus, locomotor activity gradually decreased across the 1-h test. TTX pretreatment infused into the dmPFC, DH, vmPFC, SSbf, or SStr failed to significantly alter locomotor activity relative to vehicle pretreatment, although TTX pretreatment infused into the SStr produced a nonsignificant trend toward a decrease in locomotion (ANOVA treatment main effect, F(1,9)=3.69, _P_=0.08).
Figure 5
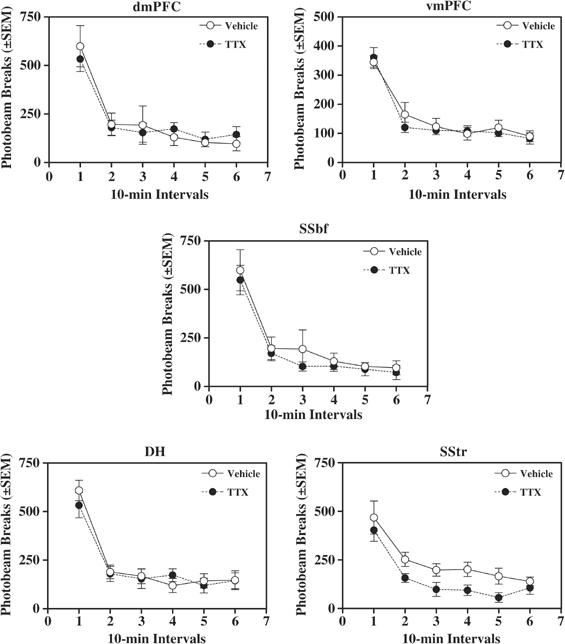
Locomotor activity during a 1-h test session following intracranial pretreatment with TTX (_N_=5–6/brain region) or vehicle (_N_=5–6/brain region). An automated photocell system recorded the number of photobeam interruptions elicited by a rat moving in the chamber. TTX infused into the dmPFC, DH, vmPFC, SStr, or SSbf failed to alter this measure of locomotor activity, similar to findings obtained following infusion of TTX into the BLA (McLaughlin and See, 2003).
DISCUSSION
Contextual Reinstatement of Cocaine-Seeking Behavior
The present study is first to map the neural circuitry of contextual reinstatement of cocaine-seeking behavior and to compare this circuitry to the neural substrates of explicit CS- and cocaine-induced cocaine-seeking behavior. Rats were initially trained to self-administer cocaine in a distinct environmental context, then underwent extinction training in the previously cocaine-paired context or in an alternate context. Consistent with previous research (Crombag et al, 2002), the magnitudes of extinction responding and the rates of extinction exhibited by the groups that underwent extinction training in the previously cocaine-paired vs alternate contexts were similar. However, presentation of the cocaine-paired contextual stimuli selectively reinstated extinguished cocaine-seeking behavior. Reinstatement was not due to dishabituation precipitated by a change in testing environment, since presentation of the alternate context failed to alter responding. TTX-induced inactivation of the dmPFC, BLA, or DH inhibited contextual reinstatement of cocaine seeking, while inactivation of the DH failed to alter explicit CS-induced and cocaine-primed reinstatement.
Before comparing the neural substrates of cocaine-seeking behavior elicited by contexts vs explicit CSs based on the above observations, we must consider that cocaine-associated contexts vs explicit CSs varied in several respects in the present study as well as in the natural environment of human drug users. First, these manipulations differed in sensory demand and complexity. A more appropriate comparison between contextual SDs and CSs of identical sensory demand and complexity could not be made because our preliminary studies indicated that a context consisting of only auditory and visual stimuli was insufficient to significantly reinstate cocaine-seeking behavior in control rats unlike the light-tone CSs presented response contingently. Therefore, the complexity of the context was increased in an attempt to facilitate context-reward learning by increasing stimulus salience (Rescorla and Wagner, 1972). Conversely, the olfactory and tactile stimuli could not be included as components of a multimodal explicit CS complex because of technical difficulties associated with their response-contingent presentation. Second, contexts and explicit CSs were administered using different modes of stimulus presentation. Contexts were presented independent of responding, whereas explicit CSs had to be presented response-contingently because passive presentation of these stimuli results in little or no cocaine-seeking behavior (Tran-Nguyen et al, 1998; Grimm et al, 2000). These differences in stimulus presentation are important because they likely determine the relationship of these stimuli to the US. Cocaine-paired stimuli presented response-contingently can act as conditioned reinforcers (ie, stimuli that elicit responding with the goal of obtaining further cue presentations) or conditioned incentive motivational stimuli (ie, stimuli that elicit responding with the goal of obtaining the US) (Mackintosh, 1974; Markou et al, 1993). In contrast, contexts engender conditioned behaviors primarily by acting as occasion setters, or SDs that retrieve response-US and/or explicit CS–US associations learned in those contexts (Bouton and Swartzentruber, 1986; Gordon and Klein, 1994; Robinson and Berridge, 2003). Thus, they signal drug availability contingent upon responding and likely act as conditioned incentive motivational stimuli as opposed to conditioned reinforcers. Based on these differences, it has been hypothesized that SDs, such as contexts, initiate relapse to drug seeking whereas explicit CSs are critical to the maintenance of drug seeking (Di Ciano and Everitt, 2003).
Emerging neurochemical and behavioral findings support the idea that SDs and explicit CSs act through separate mechanisms (See et al, 1999; Ito et al, 2000; Di Ciano and Everitt, 2003; Bossert et al, 2004). In contrast to previous studies utilizing the renewal model and discriminative stimulus training procedures, in which explicit CS–US pairings occurred in the presence of contextual or discrete SDs (Weiss et al, 2000; Ciccocioppo et al, 2001; Crombag et al, 2002; Crombag and Shaham, 2002; Yun and Fields, 2003), explicit CSs in the present study were not programmed to occur in the drug-paired context allowing isolation of the motivational effects of the context from those of explicit CSs. Under these conditions, strong context–response–US associations formed in the drug-paired context, enabling the context to reinstate cocaine-seeking behavior. The finding that DH inactivation selectively abolished contextual reinstatement of extinguished cocaine-seeking behavior is consistent with the argument that context acted as an occasion setter or ‘background stimulus’ as opposed to a CS paired directly with cocaine (see below).
Contributions of the DH, BLA, and dmPFC to Cocaine-Seeking Behavior
Functional inactivation of the dmPFC or BLA abolished the ability of the cocaine-paired context to elicit cocaine-seeking behavior (Figure 3), similar to the effects of dmPFC or BLA inactivation on explicit CS-induced reinstatement of cocaine-seeking behavior (McLaughlin and See, 2003). In contrast, inactivation of the DH abolished contextual reinstatement (Figure 3), but failed to alter explicit CS- or cocaine-induced reinstatement, of cocaine-seeking behavior (Figure 4). It is unlikely that the TTX-induced impairments in cocaine-seeking behavior were due to nonspecific performance deficits, since TTX impaired operant responding in a lever- and stimulus-specific manner and failed to alter locomotor activity (Figure 5, McLaughlin and See, 2003). Furthermore, the effects of TTX were anatomically specific, since TTX failed to impair responding when administered to brain regions adjacent and/or dorsal to the target brain regions (ie, vmPFC, SSbf, SStr). Overall, the findings support the hypothesis that unique but partially overlapping brain systems mediate the motivational effects of drug contexts vs explicit CSs on cocaine-seeking behavior. Furthermore, the DH, BLA, and dmPFC appear to contribute to cocaine-seeking behavior with varying degree of specificity regarding the type of manipulation that is used to trigger relapse (see below).
The present study suggests that the functional integrity of the DH is necessary for expression of context-induced, but not explicit CS- or cocaine-induced, cocaine-seeking behavior. The differential effects of DH inactivation on contextual vs explicit CS-induced reinstatement were not likely due to differences in stimulus modality, since the DH receives rich multimodal sensory input from sensory cortical association areas (Moser and Moser, 1998). Instead, these effects likely reflect the differential involvement of the DH in various forms of associative learning. The DH is not necessary for explicit CS–US or ‘foreground’ contextual conditioning since post-training DH lesions or inactivation fail to impair conditioned fear both to explicit (Kim and Fanselow, 1992; Maren et al, 1997) and contextual CSs paired directly with shock (Phillips and LeDoux, 1994), and fail to impair contextual discrimination performance (Good and Honey, 1991). In contrast, the integrity of the DH is necessary for ‘background’ contextual fear conditioning, since post-training DH lesions abolish conditioned fear to a context that previously served as a background to CS-shock pairings and impair context-dependent latent inhibition to an explicit CS (Kim and Fanselow, 1992; Phillips and LeDoux, 1994; Maren et al, 1997; Holt and Maren, 1999). The DH interacts with subcortical brain regions via its output structure, the dorsal subiculum (Jay et al, 1992; Ferbinteanu and McDonald, 2001). Thus, it is somewhat surprising that dorsal subiculum inactivation fails to alter cocaine-seeking behavior elicited by a combination of a light-tone contextual SD and an explicit CS, or by a combination of these stimuli and a cocaine priming injection (Black et al, 2004). Cocaine seeking is expected to be maintained, in part, by the SDs under these testing conditions; thus, these findings appear to be inconsistent with those of Experiment 1. However, it is possible that a bimodal SD does not exert significant control over cocaine seeking in the presence of a more salient explicit CS or a cocaine priming injection and, under these conditions, cocaine-seeking behavior does not depend on the functional integrity of the DH as shown in Experiment 2. Alternatively, cocaine-seeking behavior is perhaps controlled by the SD, but the DH mediates contextual reinstatement via its cortically projecting output region, the lateral entorhinal cortex, as opposed to the dorsal subiculum (Jay et al, 1992; Ferbinteanu and McDonald, 2001). Overall, these and the present findings suggest that the DH does not appear to mediate the incentive motivational effects of explicit cocaine-paired CSs or cocaine itself. However, consistent with its theorized role in spatial and configular associations (Sutherland et al, 1989; Sutherland and McDonald, 1990; Rolls, 2000), the DH mediates expression of cocaine-seeking behavior elicited by contextual background stimuli and/or occasion setters. Future studies will need to elucidate the involvement of other regions of the hippocampal formation in context-induced cocaine seeking.
Previous studies suggest that the functional integrity of the BLA is necessary for the maintenance of stimulus-reward associations that underlies the ability of cocaine-paired stimuli to elicit goal-directed behaviors. Consistent with this characterization, rostral BLA lesions or inactivation also abolish expression of cocaine-seeking behavior elicited by discrete cocaine-predictive SDs (Kantak et al, 2002) or explicit CSs (Meil and See, 1997; Grimm and See, 2000; McLaughlin and See, 2003) and spatial place conditioning (White and McDonald, 1993). Similarly, post-training BLA lesions abolish expression of conditioned fear to a shock-paired context as well as to an explicit CS (Maren, 1998; 2001), suggesting that the integrity of the rostral BLA is necessary for the expression of previously established context-US as well as discrete CS–US associations. However, caudal BLA lesions or inactivation fail to abolish cocaine-seeking behavior elicited by cocaine-predictive stimuli (Fuchs et al, 2002b; Kantak et al, 2002) and impair extinction of cocaine place conditioning (Fuchs et al, 2002b; but see Hiroi and White, 1991). In the present study, BLA inactivation abolished expression of contextual reinstatement of extinguished cocaine-seeking, similar to the effects of rostral BLA inactivation on explicit CS-induced reinstatement. Alternatively, BLA inactivation may have abolished contextual reinstatement by impairing the rat's ability to solve this task by treating components of the context as a constellation of CSs. However, the idea that rats simply processed the context as a collection of CSs is inconsistent with the finding that DH inactivation had a differential effect on contextual vs explicit CS-induced reinstatement of cocaine seeking. Thus, collectively, the above findings suggest that the contribution of the BLA to cocaine-seeking behavior is subregion specific, and less specialized than that of the DH. Unlike the DH, portions of the BLA are involved in determining the incentive motivational significance of both cocaine-paired contextual and explicit CSs.
Inactivation of the dmPFC, which targeted the anterior cingulate and dorsal prelimbic cortices, abolished contextual reinstatement of cocaine-seeking behavior. This finding adds to a growing literature indicating that the functional integrity of the dmPFC is necessary for cocaine-seeking behavior. For instance, impairment of dmPFC function disrupts stimulus control over operant responding on second-order schedules of cocaine reinforcement (Weissenborn et al, 1997) and attenuates explicit CS-induced reinstatement of cocaine-seeking behavior (McLaughlin and See, 2003). Additionally, the dmPFC exhibits neural activation in response to both cocaine-paired stimuli and cocaine priming injections (Brown et al, 1992; Neisewander et al, 2000; Ciccocioppo et al, 2001) and the functional integrity of this structure is necessary for cocaine-primed reinstatement of cocaine-seeking behavior (McFarland and Kalivas, 2001), unlike the BLA or DH (Figure 3b; Grimm and See, 2000; Neisewander et al, 2000; McFarland and Kalivas, 2001, but see Kantak et al, 2002; Yun and Fields, 2003). Thus, among the three structures examined, the dmPFC is the most generalized in its involvement in cocaine-seeking behavior which is consistent with its theorized role in executive functions, including attentional processes and response selection (Janer and Pardo, 1991; Devinsky et al, 1995; Bolla et al, 1998). Specifically, the dmPFC is thought to modulate the salience and motivational significance of stimuli by regulating attention to the sensory input that enters the BLA and hippocampus (Kolb, 1984; Rosenkranz and Grace, 2001) and by mediating BLA input to the nucleus accumbens (Jackson and Moghaddam, 2001), respectively. Furthermore, it has been theorized that the dmPFC initiates cocaine-seeking behavior in response to motivational stimuli, including cocaine-paired contextual SDs, explicit CSs, or cocaine, via its extensive projections to the spinal cord and the basal ganglia (Devinsky et al, 1995; Bolla et al, 1998), which may mediate the compulsive, automated aspect of cocaine-seeking behavior (McFarland and Kalivas, 2001).
Contextual reinstatement of cocaine-seeking behavior likely involves interaction of the DH, BLA, dmPFC, as well as sensory and motor structures, because following the inactivation of any of these three structures, functional integrity of the remaining two brain regions was insufficient to maintain context-induced reinstatement. However, a series of functional disconnection experiments will need to be conducted in order to determine the exact functional connectivity between these brain regions. Nevertheless, the main finding of the present study is that distinct, but partially overlapping, neural substrates mediate the motivational effects of cocaine-paired contexts, cocaine-paired explicit CSs, and cocaine; therefore, the development of pharmacotherapies that selectively target the neural substrates of context-induced relapse may be possible. Future research will need to elucidate the neuropharmacological mechanism of contextual reinstatement within the DH, BLA, and dmPFC in order to provide further information for treatment development for drug dependence.