Further delineation of the SATB2 phenotype (original) (raw)
Introduction
The special AT-rich sequence-binding protein 2 (SATB2) gene (OMIM *608148, first described as KIAA1034),1 is located in a relatively gene-poor region on chromosome 2q33.1. It encodes a protein of 733 amino acids, which shows a high degree of evolutionary conservation.2 The SATB2 protein is involved in chromatin remodeling and transcriptional regulation through its ability to bind AT-rich DNA sequences known as matrix attachment regions (MAR).3, 4 Depending on the locus, it can either repress or activate transcription of specific genes.5
Chromosomal translocations disrupting SATB2 have been associated with cleft palate (CP) and learning difficulties (OMIM #119540).6, 7, 8, 9 SATB2 is deleted in the contiguous 2q32q33 deletion syndrome, which is characterized by intellectual disability (ID), feeding difficulties with growth retardation, dysmorphic features, thin and sparse hair, cleft or high-arched palate, tooth abnormalities and behavioral problems.10, 11, 12, 13, 14, 15, 16 Subsequently, smaller deletions of 2q33.1 with similar phenotypes have been described (all of them encompassing SATB2), suggesting that haploinsufficiency of this gene might be causative for the core phenotype of the 2q32q33 microdeletion syndrome (see Figure 1).10, 17
Figure 1
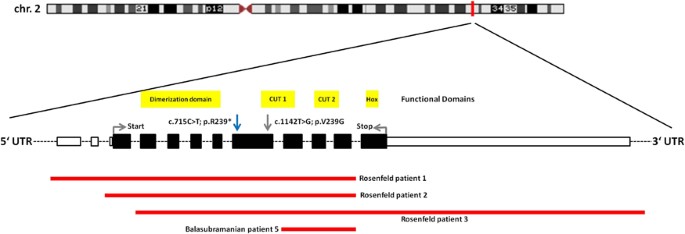
Synopsis of the patients carrying microdeletions restricted to SATB2 published so far.10, 17 Extent and position of the deletions are derived from the respective original articles. Exons are shown in real relative size (according to NM_001172517.1), whereas intronic areas are standardized and sized down. Larger deletions encompassing neighboring genes and translocations disrupting SATB2 are not shown. A comprehensive overview of larger deletions encompassing the SATB2 locus can be found in Balasubramanian et al.10 (cave: new genes have been annotated to the 2q33.1 region, which are not listed in that publication). Only one de novo nonsense mutation has been described before,18 affecting the exact same nucleotide position as the one described in this report (blue arrow). In addition, a probably pathogenic missense mutation has been reported (grey arrow).28 Indicated in yellow are the functional domains of SATB2, according to the Uniprot database. There are a dimerization domain, two CUT domains and one homeodomain (Hox), which constitute crucial DNA-binding motifs. A full color version of this figure is available at the European Journal of Human Genetics journal online.
Only one patient carrying a nonsense mutation in SATB2 has been reported in the literature to date.18 This individual was identified by screening of 59 patients with craniofacial dysmorphisms with or without ID. Recently, a patient carrying a missense mutation in SATB2 which was judged as probably damaging was identified in screening a cohort of patients with ID for de novo variants.
Here, we report and characterize a second patient carrying the same de novo nonsense mutation in SATB2 as the patient described previously (c.715C>T; p.R239*). The girl displays a specific pattern of craniofacial malformations and severe speech impairment. We propose the delineation of a clinically recognizable syndrome – the _SATB2_-associated syndrome (SAS), which is likely to be underdiagnosed. This report should enable a targeted diagnostic approach in similar patients.
Clinical report
The index patient is a 3-year-old girl, the first child of healthy non-consanguineous parents of central European descent. She was born after an uneventful pregnancy by vaginal delivery at 42 weeks gestation. APGAR scores were 9/10/10, and the pH value of the umbilical cord was 7.34. Birth measurements were within normal limits (length 52 cm (50th centile), weight 3320 g (25th–50th centile), head circumference 35 cm (50th centile)). Physical examination after birth showed Pierre Robin sequence with micrognathia and cleft soft and hard palate. Apart from apnea attacks, a weak suck and mild hypotonia, the clinical course was unremarkable during the first months. There were no feeding difficulties, and growth was between the 50th and 25th centile. CP was surgically corrected at the age of 8 months. The girl continued to display muscular hypotonia. At the age of about 12 months, the parents noticed stagnation and then mild regression of her general development. Walking was achieved at 22 months. Her gait was unsteady and atactic and she has not developed precise pincer grip to date. The most noticeable point with regard to development was the severely delayed speech: at the age of 3 years, she was able to pronounce a few syllables, but no words, and her receptive abilities were very limited. Assessment by the Munich developmental test battery at the age of 34 months showed that her skills for the different subtests were between 13 and 19 months. She had severe sleeping disturbances and showed restlessness/hyperactivity, as well as recurrent episodes of temper tantrums. An MRI scan of the brain at the age of 24 months did not show any abnormalities. Dentition started at the beginning of the second year. The teeth are of abnormal shape and positioning.
On examination at the age of 2 11/12 years (see Figure 2), weight was 12 kg (10th centile), height 94.5 cm (25th centile) and head circumference 49 cm (40th centile). The girl showed facial hypotonia with salivation, a broad forehead and microretrognathia. Palpebral fissures were downslanting and she had an upturned nose with a broad tip and a long and poorly modulated philtrum. She had irregularly shaped and crowded teeth.
Figure 2
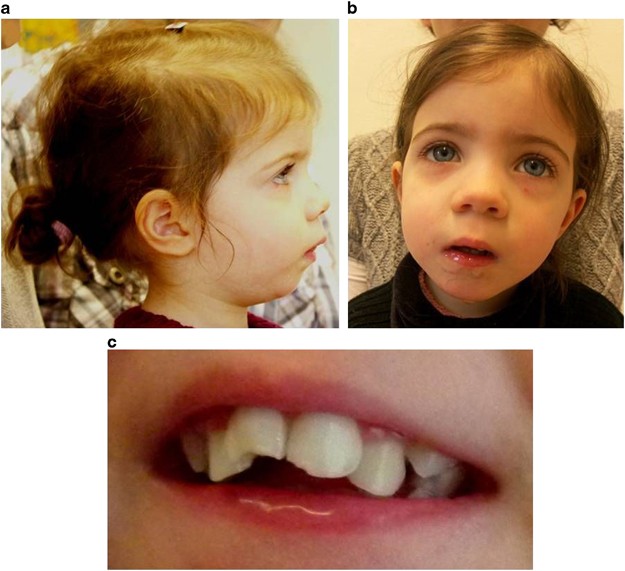
(a, b) Facial features of the patient at the age of 2 9/12 years. Note the hypotonic face with pronounced salivation. The girl displays hypertelorism, downslanting palpebral fissures, long eyelashes and a slightly upturned nose with a broad tip. She has micrognathia, a long and poorly modulated philtrum, and the ears are low set and dorsally rotated. (c) Irregular shape and positioning of the teeth. Consent of the parents for publishing the pictures was obtained.
Genetic testing including conventional chromosomal analysis, chromosomal microarray analysis and targeted molecular testing (analysis for Angelman syndrome, Rett syndrome and Fragile X syndrome) returned normal. After obtaining written consent of the parents (according to the German law on genetic testing), the patient was included into diagnostic trio-exome sequencing.
Materials and methods
DNA isolation
Genomic DNA was isolated from blood samples using the QIAamp DNA Mini Kit (Qiagen NV, Hilden, The Netherlands; cat. no. 51306) following the manufacturer’s instructions.
Exome sequencing
The exomes of the patient and her parents were sequenced using the following protocol: DNA was enriched using the SureSelect 52-Mb exome kit (Agilent, Böblingen, Germany) according to the manufacturer’s instructions. Sequencing was performed on a SOLiD 5500xl sequencer (Life Technologies, Darmstadt, Germany). In average, 130 million reads with a length of 75 bp were produced per exome. All raw reads were corrected using the spectral alignment error correction algorithm (SAET) from LifeScope and were mapped with LifeScope 2.5.1 (LifeTechnologies) against the human reference genome (hg19 from UCSC). This yielded an average coverage of 139 for the enriched target regions.
Data analysis
Variant calling – including small insertions and deletions as well as single-nucleotide variants (SNVs) – was performed by LifeScope 2.5.1 (LifeTechnologies), and samtools mpileup 0.1.18 with bcftools and vcfutils.pl.19 We used high sensitive configurations to reduce false negatives at the cost of an increased false-positive mutation rate. Calls also found in dbSNP (Database of Single-Nucleotide Polymorphisms (Bethesda, MD, USA), National Library of Medicine. Build ID: 137, http://www.ncbi.nlm.nih.gov/SNP/) or the Exome Variant Server database (NHLBI GO Exome Sequencing Project (ESP), Seattle, WA, USA; http://evs.gs.washington.edu/EVS/) with an allele frequency >5% were removed. In addition, frequently observed variants in an in-house database produced from the same sequencing technology and enrichment kit were removed (>15%, minimum number of references: 21).
Transcript and peptide alterations were annotated with NGS-SNP v1.01020 using the ENSEMBL v69 database.21 Only variants potentially changing the protein sequence were used for further analysis; intronic, UTR and synonymous mutations were removed.
Trio analysis was conducted to further filter the variant list. The remaining SNVs and insertions or deletions of the index patient were used to check for three cases: index patient (1) is compound heterozygous and each parent carries only one heterozygous mutation; (2) has a homozygous mutation and parents are heterozygous; and (3) has a de novo mutation. All three cases were automatically computed by a self-developed tool (unpublished).
Confirmation by Sanger sequencing
The variant in SATB2 (according to NM_001172517.1, see Results section) was confirmed by Sanger sequencing on an ABI 3730XL sequencer (Life Technologies) following standard procedures. Sequencing was done using the BigDye Terminator v3.1 Cycle Sequencing Kit (Life Technologies). Primers used for amplification and sequencing: _SATB2_-E08F: 5′-AGAGGATTTAGAATGGTCAGTGGTTTCTGTC-3′; _SATB2_-E08R: 5′-GGTCCTGAGTTGCATGTTGGGTTC-3′.
Confirmation in fibroblasts
In order to test the possibility of mosaicism, we performed skin biopsies in the index patient and her parents and established a fibroblast cell culture according to standard procedures. DNA extracted from these samples was analyzed by Sanger sequencing for the variant in SATB2 identified in lymphocytes.
RNA sequencing
RNA of fibroblast cultures of the index patient and her parents was isolated using the RNeasy Mini Kit from Qiagen NV (Venlo, The Netherlands; cat. no. 74014) according to the manufacturer’s protocol.
cDNA synthesis was done using the Transcriptor High Fidelity cDNA synthesis Kit from Roche Diagnostics (Rotkreuz, Schweiz, cat. no. 05 081 955 001) according to the manufacturer’s protocol, using 300 ng of RNA in each case.
RT-PCR was performed with Promega GoTaq Polymerase (cat. no. M3175) using 3 mM MgCl2, Q-solution from Qiagen (betain), and a primer concentration of 0.1 pmol/μl.
Sequencing was done with the BigDye Terminator v3.1 Cycle Sequencing Kit.
The primers used for amplification and sequencing of SATB2 were as follows: _SATB2_-F: 5′-GGAGCAGTGGAACCATGCCACAGT-3′; _SATB2_-R: 5′-CTGTGGTGAATTTGGCTGTGAGGAG-3′.
Results
Trio-exome analysis in the girl and her parents was performed generating 130 million reads per exome. On average, a coverage of 139 was achieved for the enriched target regions. A total of 3615 potential mutations remained in the index patient after population and protein alteration filtering. The analysis of the trio-exome resulted in 49 mutations (18 genes) in the category ‘compound heterozygous mutations’, eight homozygous mutations and 536 ‘_de novo_’ mutations (for definitions, see Materials and Methods section). Identified ‘_de novo_’ mutations consisted mainly of sequencing and calling errors because of the sensitive calling approach.
In addition, all variant calls were filtered by a list of 517 genes known to be associated with ID, leaving zero mutations for ‘compound heterozygous’, one mutation for ‘homozygous’ and 13 ‘_de novo_’ mutations. These variants were manually inspected and the homozygous call, as well as 12 of the potential de novo mutations, turned out to be false positives.
In summary, only one de novo mutation in a gene associated with ID/mental retardation – the SATB2 gene – was detected and confirmed by Sanger sequencing.
The heterozygous point mutation c.715C>T; p.R239* (rs137853127) identified in lymphocytes was also present in the patient’s, but absent in her parent’s, fibroblast DNA. RNA sequencing of fibroblast mRNA in the affected girl clearly showed the presence of the mutation on RNA level and excluded the mutation in both parents (r.1128c>u; for an overview, see Supplementary Material, Supplementary Figure 3).
Discussion
ID comprises a group of phenotypically and genetically highly heterogeneous entities and represents a major diagnostic challenge in clinical genetics. Trio-exome sequencing has been shown to be a powerful tool in detecting pathogenic mutations in known ID genes and in identifying novel ID genes. Here, we present a patient carrying a de novo SATB2 point mutation identified by trio-exome sequencing. It is the second patient with a nonsense mutation in this gene reported so far.
SATB2 is located on chromosome 2q33.1 and the NCBI database lists three different transcripts, which all encode the same 733-amino-acid protein.1 It is a DNA-binding protein that is expressed in vertebrates in the brain, branchial arch derivatives, at sites of bone formation and in various other tissues including kidney and gut.22, 23, 24 SATB2 was first reported as a CP gene in two individuals carrying chromosomal translocations on 2q32-q33, with the breakpoints mapped to SATB2.6 Interestingly, these two individuals did not display pronounced dysmorphisms or ID, and were therefore classified as ‘non-syndromic’ CP patients. The role of SATB2 as a CP gene was further supported by its spatial and temporal expression in the developing vertebrate palate (see below). However, screening of several large cohorts of patients with isolated CP did not reveal pathogenic sequence variants in SATB2,25 questioning its role as an isolated CP gene.
Subsequent to the publication of the translocation patients, several papers reported patients carrying heterozygous deletions of varying extent, encompassing the 2q32q33 or 2q33.1 regions (see Figure 1).10, 11, 12, 13, 14, 15, 16, 17 These deletions, ranging from 20 Mb to 200 kb approximately and all including SATB2, were associated with growth retardation, severe ID, dysmorphic features and CP or high-arched palate. As would be predicted from the variation in the extent of deletion and the specific gene content, the patients – although presenting a similar core phenotype – exhibited marked heterogeneity in some of their clinical features.
On the basis of the patient reported in this paper and the previous patient carrying the same nonsense SATB2 mutation,18 together with a patient with a small intragenic duplication26 and four patients carrying deletions restricted to SATB2,10, 17 we propose a novel clinical entity termed SAS. SAS is characterized by severe ID with no or only limited speech development, behavioral problems such as restlessness, aggression and sleep disturbances, abnormalities in craniofacial patterning such as micrognathia, cleft or high-arched palate, and problems with teeth/dentition (oligodontia and/or misshaped and crowded teeth). The patients also show subtle dysmorphisms including a hypotonic face and downslanting palpebral fissures (see Table 1 and Figure 2). To our knowledge, this clinical constellation of SAS is distinct from any clinical entity described so far.
Table 1 Comparison of clinical features of the patients carrying a microdeletion,10, 17 microduplication26 or de novo nonsense mutation18 only affecting the SATB2 gene
Leoyklang et al,18 who published the first patient with a point mutation in SATB2 back in 2007, proposed that chromosomal deletions associated with haploinsufficiency of SATB2 cause a less severe phenotype than the nonsense mutation detected in their patient. They suggested that this might be explained by a dominant-negative effect of the truncated or altered SATB2 protein. In our patient, we could detect the mutation at RNA level, indicating that the RNA is stable enough not to undergo immediate nonsense-mediated decay. A very recently published follow-up study of Leoyklang et al.27 including immunoblot analyses further supports the hypothesis of a possible dominant-negative effect of the truncated SATB2 protein.
Future functional studies of patients with different mutations will hopefully provide deeper insight into the role of normal and altered SATB2 protein during human development. Interestingly, a patient carrying a missense mutation in SATB2 (c.1142T>G, p.V381G) was recently reported in a large exome sequencing study in individuals with severe sporadic non-syndromic ID. Although the pathogenic role of this specific mutation is not yet clearly established, it seems as if this patient has a mild SAS phenotype.28
It also remains to be elucidated, whether the fact that the patient described in this report has the exactly same point mutation as the first patient described in 2007 is due to coincidence or whether there is an underlying pathogenic reason favoring this nucleotide change as a ‘hot spot’ region.
SATB2 encodes a protein that specifically binds nuclear MARs and thereby has an important role in transcriptional regulation by remodeling chromatin structure. Studies of murine Satb2 show that it is highly expressed in the developing upper and lower jaw including the palatal shelves and teeth buds, as well as in specific areas of the cortex and at sites of bone formation. Satb2 knock out mice on different backgrounds have been generated to study craniofacial patterning and osteoblast differentiation.22, 24 Heterozygous Satb2 +/− mice have a phenotype similar to that of SAS patients, including CP, micrognathia, premaxillary and nasocapsular hypoplasia, and variable incisor hypodontia or adontia. Homozygote Satb2 −/− knock out mice are born with exacerbated skeletal malformations and often die perinatally. It has been shown that Satb2 interacts directly with and enhances activity of Runx2 and Atf4, two transcription factors regulating osteoblast formation.29, 30 Furthermore, Satb2 represses the expression of several Hox genes including Hoxa2, an inhibitor of bone formation and regulator of branchial arch patterning.22 These studies, together with zebra fish studies,31, 32 have established the crucial role of Satb2 in skeletogenesis, influencing such different processes as patterning, osteoblast differentiation and matrix formation. With respect to humans, it is noteworthy that the first described patient carrying a de novo SATB2 nonsense mutation, who was 36 years old at the time of reporting, suffered from unexplained very severe osteoporosis.18
Interestingly, variants in an adjacent gene, FONG, have been linked to increased risk of osteoporosis in a Japanese genome-wide association study.33 It remains to be elucidated whether there is interaction (or relation, such as co-expression) between SATB2 and FONG.
Apart from its well-established role in craniofacial patterning and skeletogenesis, Satb2 is expressed in the developing cortex and has an important role in determining the identity of neurons during cortical development.24, 34, 35 In the mammalian cerebral cortex, neurons can be divided into two major classes: cortico-cortical projection neurons, which are concentrated in the upper layers of the cortex, and sub-cortical projection neurons, which are found in the deep layers. Early progenitor cells in the ventricular zone give rise to deep layer neurons while progenitors in the subventricular zone give rise to upper layer neurons. Expression of Satb2 is required in upper layer neurons for the formation of axonal projections that connect the two cerebral hemispheres. In Satb2 mutants, upper layer neurons fail to migrate to superficial layers and do not contribute to the corpus callosum, but erroneously to the corticospinal tract, which is normally populated by axons of deep layer neurons.24, 32 In addition to Satb2, projection identities of cortical neurons are orchestrated by three further genes, Fezf2, Ctip2 and Tbr1.33 These four genes interact with each other, and studies in double mutants led to the hypothesis that cortical neurons undergo a complex genetic switch by mutual repression to produce subcortical or callosal projections. Beyond regulating neuronal projections, Satb2 has also been shown to contribute to a network of genetic repression and derepression in the developing neocortex and thereby to regulate the expression of two disease-related genes, Auts2 (autism susceptibility gene 2) and Bhlhb5 (mutated in hereditary spastic paraplegia).36
In light of these studies, it is surprising that SAS patients do not exhibit agenesis or hypogenesis of corpus callosum or more subtle malformations of the cortical architecture that would be detectable by MRI (see Table 1). However, the crucial role of Satb2 in neuronal connectivity very likely explains the severe ID with pronounced impairment of speech development and the seizure disorder in the first patient carrying a nonsense mutation in SATB2.
In addition to SATB2, numerous other chromatin remodeling genes have been implicated in human neurodevelopmental disorders such as ID, autism spectrum disorder and schizophrenia. Examples include CBP and p300 (Rubinstein–Taybi syndrome), CHD7 (Charge syndrome), ATRX (X-linked alpha-thalassemia mental retardation syndrome) and genes of the BAF complex (Coffin–Siris and Kleefstra syndrome).37 Interestingly, mutations in several of the chromatin remodeling genes implicated in neurodevelopmental disorders are also involved in human cancer, which is also true for SATB2. In colorectal cancer, downregulation of SATB2 is associated with metastasis and poor prognosis.38, 39 Also in laryngeal carcinoma, decreased SATB2 expression is associated with a poorer outcome.40 In bone and soft tissue tumors, SATB2 has been proposed to be a novel marker for bone osteoblastic differentiation in benign and malignant mesenchymal tumours.41 A single study showed an association between elevated SATB2 mRNA levels and worse outcome in breast cancer patients.42
In conclusion, this report confirms SATB2 as an essential gene for craniofacial patterning and cognitive development in humans and leads to delineation of the SAS. Children with ID and no or severely impaired speech, cleft or high-arched palate, micrognathia and teeth abnormalities should be analyzed for deletions or point mutations in SATB2. The identification of further patients with SAS will help to establish the specific role(s) of this gene in human development, determine the long-term prognosis and possibly to identify strategies for therapy.
References
- Kikuno R, Nagase T, Ishikawa K et al: Prediction of the coding sequences of unidentified human genes. XIV. The complete sequences of 100 new cDNA clones from brain which code for large proteins in vitro. DNA Res 1999; 6: 197–205.
Article CAS Google Scholar - Sheehan-Rooney K, Palinkasova B, Eberhart JK, Dixon MJ : A cross-species analysis of Satb2 expression suggests deep conservation across vertebrate lineages. Dev Dyn 2010; 239: 3481–3491.
Article CAS Google Scholar - Britanova O, Akopov S, Lukyanov S, Gruss P, Tarabykin V : Novel transcription factor Satb2 interacts with matrix attachment region DNA elements in a tissue-specific manner and demonstrates cell-type-dependent expression in the developing mouse CNS. Eur J Neurosci 2005; 21: 658–668.
Article Google Scholar - Szemes M, Gyorgy A, Paweletz C, Dobi A, Agoston DV : Isolation and characterization of SATB2, a novel AT-rich DNA binding protein expressed in development- and cell-specific manner in the rat brain. Neurochem Res 2006; 31: 237–246.
Article CAS Google Scholar - Gyorgy AB, Szemes M, de Juan Romero C, Tarabykin V, Agoston DV : SATB2 interacts with chromatin-remodeling molecules in differentiating cortical neurons. Eur J Neurosci 2008; 27: 865–873.
Article Google Scholar - FitzPatrick DR, Carr IM, McLaren L et al: Identification of SATB2 as the cleft palate gene on 2q32-q33. Hum Mol Genet 2003; 12: 2491–2501.
Article CAS Google Scholar - Brewer CM, Leek JP, Green AJ et al: A locus for isolated cleft palate, located on human chromosome 2q32. Am J Hum Genet 1999; 65: 387–396.
Article CAS Google Scholar - Tegay DH, Chan KK, Leung L et al: Toriello-Carey syndrome in a patient with a de novo balanced translocation [46,XY,t(2;14)(q33;q22)] interrupting SATB2. Clin Genet 2009; 75: 259–264.
Article CAS Google Scholar - Talkowski ME, Rosenfeld JA, Blumenthal I et al: Sequencing chromosomal abnormalities reveals neurodevelopmental loci that confer risk across diagnostic boundaries. Cell 2012; 149: 525–537.
Article CAS Google Scholar - Balasubramanian M, Smith K, Basel-Vanagaite L et al: Case series: 2q33.1 microdeletion syndrome—further delineation of the phenotype. J Med Genet 2011; 48: 290–298.
Article CAS Google Scholar - Van Buggenhout G, Van Ravenswaaij-Arts C, Mc Maas N et al: The del(2)(q32.2q33) deletion syndrome defined by clinical and molecular characterization of four patients. Eur J Med Genet 2005; 48: 276–289.
Article CAS Google Scholar - de Ravel TJ, Balikova I, Thiry P, Vermeesch JR, Frijns JP : Another patient with a de novo deletion further delineates the 2q33.1 microdeletion syndrome. Eur J Med Genet 2009; 52: 120–122.
Article Google Scholar - Urquhart J, Black GC, Clayton-Smith J : 4.5 Mb microdeletion in chromosome band 2q33.1 associated with learning disability and cleft palate. Eur J Med Genet 2009; 52: 454–457.
Article Google Scholar - Ferreira SI, Matoso E, Venancio M, Saraiva J, Melo JB, Carreira IM : Critical region in 2q31.2q32.3 deletion syndrome: report of two phenotypically distinct patients, one with an additional deletion in Alagille syndrome region. Mol Cytogenet 2012; 5: 25.
Article CAS Google Scholar - Mencarelli MA, Caselli R, Pescucci C et al: Clinical and molecular characterization of a patient with a 2q31.2-32.3 deletion identified by array-CGH. Am J Med Genet A 2007; 143A: 858–865.
Article CAS Google Scholar - Cocchella A, Malacarne M, Forzano F et al: The refinement of the critical region for the 2q31.2q32.3 deletion syndrome indicates candidate genes for mental retardation and speech impairment. Am J Med Genet B Neuropsychiatr Genet 2010; 153B: 1342–1346.
Article Google Scholar - Rosenfeld JA, Ballif BC, Lucas A et al: Small deletions of SATB2 cause some of the clinical features of the 2q33.1 microdeletion syndrome. PLoS One 2009; 4: e6568.
Article Google Scholar - Leoyklang P, Suphapeetiporn K, Siriwan P et al: Heterozygous nonsense mutation SATB2 associated with cleft palate, osteoporosis, and cognitive defects. Hum Mutat 2007; 28: 732–738.
Article CAS Google Scholar - Li H, Handsaker B, Wysoker A et al: The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009; 25: 2078–2079.
Article Google Scholar - Grant JR, Arantes AS, Liao X, Stothard P : In-depth annotation of SNPs arising from resequencing projects using NGS-SNP. Bioinformatics 2011; 27: 2300–2301.
Article CAS Google Scholar - Flicek P, Amode MR, Barrell D et al: Ensembl 2012. Nucleic Acids Res 2012; 40: D84–D90.
Article CAS Google Scholar - Dobreva G, Chahrour M, Dautzenberg M et al: SATB2 is a multifunctional determinant of craniofacial patterning and osteoblast differentiation. Cell 2006; 125: 971–986.
Article CAS Google Scholar - Huang Y, Song NN, Lan W et al: Expression of transcription factor Satb2 in adult mouse brain. Anat Rec (Hoboken) 2013; 296: 452–461.
Article CAS Google Scholar - Britanova O, de Juan Romero C, Cheung A et al: Satb2 is a postmitotic determinant for upper-layer neuron specification in the neocortex. Neuron 2008; 57: 378–392.
Article CAS Google Scholar - Vieira AR, Avila JR, Daack-Hirsch S et al: Medical sequencing of candidate genes for nonsyndromic cleft lip and palate. PLoS Genet 2005; 1: e64.
Article Google Scholar - Kaiser AS, Maas B, Wolff A et al: Girl with developmental delay, nearly absent speech and oligodontia: first case of an intragenic SATB2 gene duplication. Medizinische Genetik 2013; 25: 125.
Google Scholar - Leoyklang P, Suphapeetiporn K, Srichomthong C et al: Disorders with similar clinical phenotypes reveal underlying genetic interaction: SATB2 acts as an activator of the UPF3B gene. Hum Genet 2013; 132: 1383–1393.
Article CAS Google Scholar - Rauch A, Wieczorek D, Graf E et al: Range of genetic mutations associated with severe non-syndromic sporadic intellectual disability: an exome sequencing study. Lancet 2012; 380: 1674–1682.
Article CAS Google Scholar - Deng Y, Wu S, Zhou H et al: Effects of a miR-31, Runx2 and Satb2 regulatory loop on the osteogenic differentiation of bone marrow mesenchymal stem cells. Stem Cells Dev 2013; 22: 2278–2286.
Article CAS Google Scholar - Hassan MQ, Gordon JA, Beloti MM et al: A network connecting Runx2, SATB2, and the miR-23a∼27a∼24-2 cluster regulates the osteoblast differentiation program. Proc Natl Acad Sci USA 2010; 107: 19879–19884.
Article CAS Google Scholar - Sheehan-Rooney K, Swartz ME, Lovely CB, Dixon MJ, Eberhart JK : Bmp and Shh signaling mediate the expression of satb2 in the pharyngeal arches. PLoS One 2013; 8: e59533.
Article CAS Google Scholar - Fish JL, Villmoare B, Kobernick K et al: Satb2, modularity, and the evolvability of the vertebrate jaw. Evol Dev 2011; 13: 549–564.
Article CAS Google Scholar - Kou I, Takahashi A, Urano T et al: Common variants in a novel gene, FONG on chromosome 2q33.1 confer risk of osteoporosis in Japanese. PLoS One 2011; 6: e19641.
Article CAS Google Scholar - Baranek C, Dittrich M, Parthasarathy S et al: Protooncogene Ski cooperates with the chromatin-remodeling factor Satb2 in specifying callosal neurons. Proc Natl Acad Sci USA 2012; 109: 3546–3551.
Article CAS Google Scholar - Alcamo EA, Chirivella L, Dautzenberg M et al: Satb2 regulates callosal projection neuron identity in the developing cerebral cortex. Neuron 2008; 57: 364–377.
Article CAS Google Scholar - Srinivasan K, Leone DP, Bateson RK et al: A network of genetic repression and derepression specifies projection fates in the developing neocortex. Proc Natl Acad Sci USA 2012; 109: 19071–19078.
Article CAS Google Scholar - Ronan JL, Wu W, Crabtree GR : From neural development to cognition: unexpected roles for chromatin. Nat Rev Genet 2013; 14: 347–359.
Article CAS Google Scholar - Wang S, Zhou J, Wang XY et al: Down-regulated expression of SATB2 is associated with metastasis and poor prognosis in colorectal cancer. J Pathol 2009; 219: 114–122.
Article CAS Google Scholar - Eberhard J, Gaber A, Wangefjord S et al: A cohort study of the prognostic and treatment predictive value of SATB2 expression in colorectal cancer. Br J Cancer 2012; 106: 931–938.
Article CAS Google Scholar - Liu TR, Xu LH, Yang AK et al: Decreased expression of SATB2: a novel independent prognostic marker of worse outcome in laryngeal carcinoma patients. PLoS One 2012; 7: e40704.
Article CAS Google Scholar - Conner JR, Hornick JL : SATB2 is a novel marker of osteoblastic differentiation in bone and soft tissue tumours. Histopathology 2013; 63: 36–49.
Article Google Scholar - Patani N, Jiang W, Mansel R, Newbold R, Mokbel K : The mRNA expression of SATB1 and SATB2 in human breast cancer. Cancer Cell Int 2009; 9: 18.
Article Google Scholar
Acknowledgements
We thank the parents of the girl for their consent and their positive attitude toward publishing. We also thank Erika Selo for her help in establishing the fibroblast cultures. We thank Miriam Döcker for helpful suggestions on the manuscript.
Author information
Author notes
- Saskia Biskup and Deborah Bartholdi: These authors contributed equally to this work and share senior authorship.
Authors and Affiliations
- Institute of Clinical Genetics, Klinikum Stuttgart, Stuttgart, Germany
Dennis Döcker, Christiane Spaich, Saskia Biskup & Deborah Bartholdi - CeGaT GmbH, Tübingen, Germany
Max Schubach, Moritz Menzel & Saskia Biskup - Hertie Institute for Clinical Brain Research, Tübingen, Germany
Marita Munz & Saskia Biskup
Authors
- Dennis Döcker
You can also search for this author inPubMed Google Scholar - Max Schubach
You can also search for this author inPubMed Google Scholar - Moritz Menzel
You can also search for this author inPubMed Google Scholar - Marita Munz
You can also search for this author inPubMed Google Scholar - Christiane Spaich
You can also search for this author inPubMed Google Scholar - Saskia Biskup
You can also search for this author inPubMed Google Scholar - Deborah Bartholdi
You can also search for this author inPubMed Google Scholar
Corresponding author
Correspondence toDeborah Bartholdi.
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on European Journal of Human Genetics website
Supplementary information
Rights and permissions
About this article
Cite this article
Döcker, D., Schubach, M., Menzel, M. et al. Further delineation of the SATB2 phenotype.Eur J Hum Genet 22, 1034–1039 (2014). https://doi.org/10.1038/ejhg.2013.280
- Received: 17 July 2013
- Revised: 18 October 2013
- Accepted: 30 October 2013
- Published: 04 December 2013
- Issue Date: August 2014
- DOI: https://doi.org/10.1038/ejhg.2013.280