Lymphatic reprogramming of blood vascular endothelium by Kaposi sarcoma–associated herpesvirus (original) (raw)
Kaposi sarcoma is the most frequently occurring malignant tumor in individuals infected with the human immunodeficiency (HIV) virus and also occurs in HIV-negative immunosuppressed individuals. Kaposi sarcoma mainly affects the skin and forms lesions of various types, including early inflammatory and patch stage lesions, and tumors with a predominant population of spindle cells. Infection with Kaposi sarcoma–associated herpesvirus (KSHV, also known as human herpesvirus-8) is essential for the formation of Kaposi sarcoma tumors1,2. Both latent viral genes, such as latency-associated nuclear antigen (LANA), and lytic viral genes, such as viral G-protein-coupled receptor, have been implicated in KSHV-mediated tumorigenesis3. In Kaposi sarcoma lesions, cells infected with KSHV characteristically appear spindle-shaped and are associated with slit-like spaces that sometimes contain red blood cells. Kaposi sarcoma is considered to be a neoplasm of KSHV-infected lymphatic endothelium, due to the morphological characteristics of the tumor cells and the expression of several lymphatic lineage–specific proteins, including VEGFR-3 and podoplanin4,5,6,7.
The homeobox gene PROX1 is a master gene that controls lymphatic vessel development and differentiation8, and ectopic expression of PROX1 in differentiated blood vascular endothelial cells leads to lymphatic endothelial reprogramming of these cells9,10. Because KSHV can infect blood vascular endothelium, such as human umbilical vein endothelial cells, in vitro, we wondered whether KSHV infection might result in lymphatic reprogramming of blood vascular endothelium, potentially involving upregulation of PROX1.
We first characterized the lineage-specific gene expression of cultured human lymphatic endothelial cells (LECs) versus blood vascular endothelial cells (BECs)11 by Affymetrix HU133A gene arrays. We found that LECs, but not BECs, expressed PROX1, XLKD1 (encoding the hyaluronan receptor LYVE-1) and a number of other lymphatic lineage–specific genes (Table 1). We then infected human dermal microvascular endothelial cells (HDMECs) with KSHV and carried out two independent transcriptional profiling studies 7 d later. Efficient KSHV infection was confirmed by high levels of expression of LANA mRNA in infected HDMECs. We found that 3–7% of infected HDMECs were ORF59-positive and 1.5–3% were K8.1 positive, in agreement with previous results12,13,14. This indicates that <10% of the cells were undergoing lytic reactivation.
Table 1 Genes whose expression was higher in LECs than in BECs and was upregulated in KSHV-infected endothelial cells
Infection of HDMECs with KSHV resulted in significant upregulation (P < 0.002) of ∼70% of the previously identified lymphatic lineage–specific genes that were upregulated by a factor of at least 3 in LECs compared with BECs (Table 1). KSHV-induced genes included those encoding PROX1, LYVE-1, reelin, follistatin, desmoplakin and leptin receptor. These results concur with the finding that KSHV infection induced a comprehensive reprogramming of BECs to adopt an LEC phenotype15, although no significant upregulation of PROX1 mRNA was detected in that report, probably owing to differences in cell isolation and culture conditions and in the time points investigated.
Whereas TGFA was reported to be downregulated 5 weeks after KSHV infection13, we found strong induction of TGFA 7 d after infection. Induction of lymphatic lineage–specific genes was also detected after KSHV infection of human umbilical vein endothelial cells that are of blood vascular origin (data not shown). Among lymphatic lineage–specific genes that were not significantly induced by KSHV infection were those encoding macrophage mannose receptor and intestinal trefoil factor. Conversely, several blood vascular–specific genes, such as those encoding chemokine receptor CXCR4, placental growth factor, CD44, laminin, VEGF-C, neuropilin-1 and intercellular adhesion molecule-1, were strongly downregulated after KSHV infection. In agreement with a previous study12, KSHV infection induced expression of cKit, which is involved in Kaposi sarcoma spindle cell transformation, by a factor of 52.7.
The transcriptional profiling results were confirmed by quantitative real-time RT-PCR (Supplementary Methods online) for 35 of the 36 genes shown in Table 1. Expression of PROX1 and XLKD1 was ∼8 times higher after KSHV infection, and a less-pronounced induction of the genes encoding the lymphatic markers VEGFR-3, podoplanin and secondary lymphoid chemokine was also observed (Fig. 1a). Induction of VEGFR-3 and podoplanin was comparable to that caused by transfection of BECs with PROX1 (ref. 9,10). Northern-blot analyses confirmed the strong upregulation of PROX1 mRNA after KSHV infection of BECs (by a factor of 6.0) to levels that were close to those observed in primary LECs (9.3 times higher than in BECs).
Figure 1: KSHV infection of BECs induces expression of the lymphatic lineage–specific genes PROX1 and XLKD1.
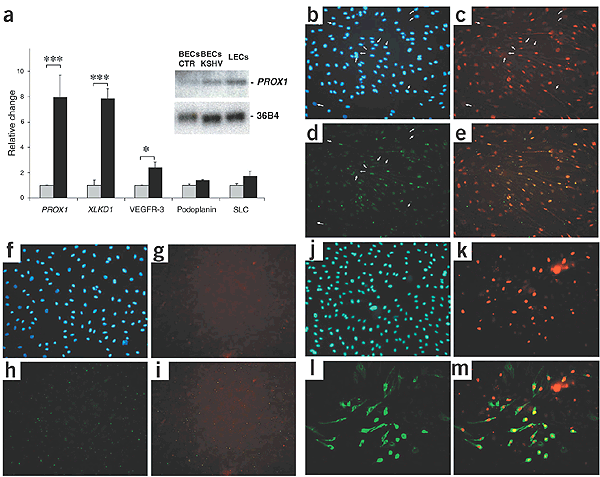
(a) Quantitative real-time TaqMan RT-PCR analysis showed that PROX1 and XLKD1 were induced by a factor of ∼8 in BECs infected with KSHV at 7 d after infection (filled bars) as compared with uninfected BECs (open bars). Expression of the lymphatic markers VEGFR-3, podoplanin and secondary lymphoid chemokine (SLC) was induced to a lesser extent. Inset, Northern-blot analysis confirmed the induction of PROX1 expression after KSHV infection of BECs, to levels that were close to those observed in primary LECs. BECs CTR indicates uninfected BECs. Ribosome-associated gene 36B4 served as loading control. (b–e) Seven days after infection, KSHV-infected BECs expressed LANA (red, c). More than 90% of all LANA-positive BECs also expressed PROX1 (green, d), whereas no LANA-negative BECs did (e; merge of c and d). Arrows indicate LANA-negative, PROX1-negative cells. (f–i) No expression of LANA (g) or PROX1 (h) was detected in uninfected BECs (i; merge of g and h. (j–m) KSHV infection upregulated LYVE-1 expression in BECs, as confirmed by colabeling for LANA (red, k) and LYVE-1 (green, l) in >70% of LANA-positive BECs. LANA-negative cells did not express LYVE-1 (m). Hoechst nuclear stain is shown in blue (b,f,j).
We next infected cultured BECs with KSHV and assessed their expression of PROX1 by immunostaining. Seven days after infection, KSHV-infected BECs, but not uninfected cells, expressed LANA (Fig. 1c,g). More than 90% of all LANA-positive BECs also expressed PROX1, whereas no LANA-negative cells did; uninfected BECs did not express PROX1 (Fig. 1d,e,h,i). KSHV infection also upregulated expression of LYVE-1 protein in >70% of LANA-positive BECs, whereas LANA-negative cells did not express LYVE-1 (Fig. 1j–m). We next investigated whether LANA might be directly involved in the KSHV-induced upregulation of PROX1 expression. Retroviral transduction of BECs with LANA induced expression of PROX1 by a factor of 1.93 (± 0.14), whereas transduction with two other latency-associated KSHV genes (v-flip and cyclin) did not affect PROX1 expression, as assessed by quantitative real-time RT-PCR.
Transfection of KSHV-infected BECs with PROX1 siRNA partially inhibited the inductive effect of KSHV infection on the expression of LYVE-1, CD36, c-maf and several other lymphatic-specific proteins (Table 1), indicating that PROX1 has an important role in the lymphatic reprogramming of BECs by KSHV. The fact that PROX1 knockdown did not completely prevent KSHV-induced lymphatic reprogramming of BECs suggests that additional transcriptional regulators might be involved. The finding that PROX1 was downregulated after infection of LECs by KSHV, associated with a transcriptional shift towards a BEC phenotype15, further supports the idea that PROX1 has an important role in mediating the effects of KSHV on endothelial reprogramming.
We next investigated whether PROX1 and LYVE-1 were also expressed by Kaposi sarcoma tumor cells in situ. In all cases examined, Kaposi sarcoma tumor cells, but not normal blood vascular endothelium, expressed PROX1 and LYVE-1 (Supplementary Fig. 1 online), as well as the panvascular marker CD31. PROX1 was expressed by >90% of Kaposi sarcoma tumor cells that expressed LANA (Supplementary Fig. 1 online). In contrast, Kaposi sarcoma tumor cells did not express the blood vascular markers laminin and VEGF-C, which were downregulated after KSHV infection in vitro. Lesional Kaposi sarcoma tumor cells maintained expression of the blood vessel–specific proteins endoglin and type IV collagen (Supplementary Fig. 1 online), however, whose expression was also not affected by KSHV infection of HDMECs.
Our results show that KSHV infection activates a developmental genetic program specifying lymphatic endothelial cell fate, including upregulation of PROX1 that is essential for the embryonic development of the mammalian lymphatic system8.
GEO accession numbers. GSM21935, GSM21961, GSM21971, GSM21972.
Note: Supplementary information is available on the Nature Genetics website.