Angiotensin II AT1 Receptor Blockade Ameliorates Brain Inflammation (original) (raw)
INTRODUCTION
Brain inflammation is an essential mechanism to restore homeostasis in response to stress, infection, or injury (Yong and Rivest, 2009). Circulating proinflammatory cytokines and hormones, bacterial endotoxins and products of neuronal injury activate transcription factors, setting in motion inflammatory cascades with participation of brain parenchymal microglia and blood-derived infiltrating macrophages (Licinio and Wong, 1997). Although a well-regulated central inflammatory reaction contributes to reestablish homeostasis, exaggerated responses lead to chronic inflammation and neuronal damage (Dantzer et al, 2008). Such excessive activation of immune responses increases the risk of disease in vulnerable individuals, and is an important contributing factor underlying the pathophysiology of many psychiatric and neurological diseases. These include major depression, schizophrenia, autism, post-traumatic stress disorder, and neurodegenerative disorders, such as Alzheimer's and Parkinson's disease, amyotrophic lateral sclerosis, multiple sclerosis, HIV-associated dementia, and traumatic brain injury (Fassbender et al, 2004; Hope et al, 2009; Miller et al, 2009; Rivest, 2009).
Because drugs used in peripheral inflammatory conditions are not fully effective in the brain (Nimmo and Vink, 2009), the search for novel, safe, and effective central anti-inflammatory drugs is of major interest. Recent studies have focused on a neuropeptide, angiotensin II (Ang II) initially discovered as a circulating prohypertensive hormone. Ang II was later recognized as a pleiotropic factor locally formed in most tissues, including the brain (Saavedra, 1992). Most of the effects of Ang II are dependent on the stimulation of its AT1 receptors (Saavedra et al, 2006), and AT1 receptor overactivity promotes peripheral vascular and tissue inflammation (Savoia and Schiffrin, 2007). AT1 receptor blockers (ARBs) not only antagonize the Ang II-mediated vasoconstriction but also reverse peripheral inflammation in hypertension, atherosclerosis, and diabetes (Savoia and Schiffrin, 2007). For this reason ARBs, compounds with excellent safety margins, are commonly used not only to regulate blood pressure, but also for their capacity to protect end organs, thus decreasing heart failure and renal damage (Savoia and Schiffrin, 2007).
We previously used the centrally acting ARB candesartan (Nishimura et al, 2000), a compound blocking central AT1 receptor stimulation by Ang II when administered systemically (Seltzer et al, 2004). In spontaneously hypertensive rats (SHRs), a rodent model of genetic hypertension with overactive brain AT1 receptors, behavioral and cognitive deficits, hypersensitive to stress and to brain inflammation (Kvetnansky et al, 1979; Saavedra, 1992; Sirén et al, 1992; Russell, 2003), candesartan reversed cerebrovascular inflammation and brain ischemia (Ando et al, 2004; Zhou et al, 2005). The effects of candesartan were not restricted to hypertensive models, because the ARB also reduced stroke-related inflammation in normotensive rats (Schmerbach et al, 2008).
We hypothesized that excessive AT1 receptor activation may participate in other types of inflammatory responses in the brain, and we postulated a general anti-inflammatory effect of ARBs, unrelated to their blood pressure reducing effect. To test our hypothesis, we first selected a well-characterized model of inflammation, the activation of the innate immune response in the periphery and in the brain by a systemic administration of the bacterial endotoxin lipopolysaccharide (LPS) (Quan and Banks, 2007). AT1 receptor blockade significantly decreased endotoxin-induced inflammation both in vivo in peripheral tissues of normotensive rats (Sánchez-Lemus et al, 2008, 2009a, 2009b), and in vitro in circulating monocytes from normotensive human volunteers (Larrayoz et al, 2009). These studies demonstrated that ARBs could be considered potent anti-inflammatory agents by effects unrelated to their anti-hypertensive properties.
To establish whether ARBs can exert central anti-inflammatory activity, in this study we determined the effect of AT1 receptor blockade on the LPS-induced brain inflammatory response. This study extends our preliminary results reporting anti-inflammatory effects of candesartan in the cingulate cortex (Benicky et al, 2009). To investigate whether anti-inflammatory effects observed in vivo were dependent on cross-system interactions, we included in vitro assays using LPS target cells. We studied cerebellar granule cells, a highly homogeneous population of neurons (Gao et al, 1995), cerebral microvascular endothelial cells (Nakagawa et al, 2009), and cortical microglial cells (Giulian and Baker, 1986). We show here that AT1 receptor inhibition significantly and broadly decreases the brain immune response in vivo as well as the inflammatory responses in all LPS target cells studied. In addition, candesartan ameliorates LPS-induced sickness behavior, the acute anorexic effects and body weight loss (Konsman and Dantzer, 2001) and anxiety-like behavior in untreated and LPS-treated rats. These studies show that ARBs are potent central anti-inflammatory compounds, an important property of translational value.
MATERIALS AND METHODS
Detailed methods can be found in Supplementary Materials and Methods section.
Animals
Nine-week-old male Wistar Hannover (WH) rats and SHR (Taconic Farms, Germantown, NY) were used for in vivo experiments. Three-week-old male WH rats were used for isolation of brain microvascular endothelial cells. Eight-day-old and one-day-old Sprague Dawley male and female pups (Taconic Farms) were used for isolation of cerebellar granule cells and cortical microglia, respectively. The National Institute of Mental Health Animal Care and Use Committee (Bethesda, MD) approved all procedures.
In Vivo Studies
Short-term Ang II AT1 receptor blockade
WH rats received daily subcutaneous injections of vehicle or candesartan (CV-11974, Astra-Zeneca, Mölndal, Sweden) at the dose of 1 mg/kg per day. This dose was similar to that used in the treatment of human cardiovascular disorders (Weinberg et al, 2004). The animals were injected intraperitoneally with 50 μg/kg LPS (Escherichia coli serotype 055:B5; Sigma-Aldrich, St Louis, MO) or saline on day 3 and were killed 3 h later by decapitation. Trunk blood was used for plasma preparation and brains were dissected, frozen in isopentane on dry ice, and stored at −80 °C until used.
Behavioral studies
For anorexia determination, separate groups of individually housed WH rats were subjected to short-term Ang II AT1 receptor blockade followed by intraperitoneal injection of 50 μg/kg LPS as described above. Animals were euthanized 2 days after LPS injection. Body weight and food consumption were monitored daily for the duration of experiment. For anxiety determination, SHR were treated as above. At 3 h after LPS injection, the rats were tested on an elevated plus maze as described previously (Saavedra et al, 2006). The time spent and the number of entries into the open and closed arms were recorded during 5 min.
Sustained Ang II AT1 receptor blockade
WH rats were implanted subcutaneously with osmotic minipumps (model 2002, Alza Scientific Products, Palo Alto, CA) containing candesartan (1 mg/kg per day) or vehicle. On day 14, the animals were injected intraperitoneally with 50 μg/kg LPS or saline and were killed 3 h later by decapitation. Trunk blood was used for plasma preparation and brains were dissected, frozen in isopentane on dry ice and stored at −80 °C until used. For immunohistochemical studies, separate groups of animals treated as above were anesthetized 3 h after LPS or saline injection and transcardially perfused with 4% buffered formaldehyde. The brains were dissected, post-fixed overnight, cryoprotected by 30% sucrose, frozen in isopentane on dry ice, and stored at −80 °C until used.
In Vitro Studies
Cerebellar granule cell culture
Cells were isolated from 8-day-old rat pups as described previously (Gao et al, 1995). The isolated cells were plated at a density of 1.2 × 106 cells/ml and cultured for 6–14 days. Before the experiment, the cells were starved overnight in plain medium and subsequently treated for 4 h with 500 ng/ml LPS with or without 2 h pre-treatment with 5 μM candesartan. The dose of candesartan used was similar to those necessary to reduce Ang II effects in cell cultures (Seeger et al, 2001). Medium was saved and cells were lysed for RNA isolation as described below.
Brain microvascular endothelial cell culture
Cultures were prepared as described previously (Nakagawa et al, 2009). The cells were used after the first passage when they reached 80% confluence. Before the experiment, cells were starved overnight in a serum-free medium followed by treatment with 100 ng/ml LPS for 4 h with or without 2 h pre-incubation with 10 μM candesartan. Medium was saved and cells were lysed for RNA isolation as described below.
Cortical microglia cell culture
Cultures were prepared as described (Giulian and Baker, 1986). Initially prepared mixed glial cells were cultured for 10 days and subsequently used for microglia preparation. Purified microglial cells were allowed to grow for 24 h, starved overnight in a serum-free medium, and subsequently treated for 4 h with 100 ng/ml LPS with or without 2 h pre-incubation with 10 μM candesartan. Medium was saved and cells were lysed for RNA isolation as described below.
Determination of Cytokine Levels
Cytokines in plasma and medium were measured by commercial enzyme-linked immunosorbent assay kits from BioSource (Camarillo, CA) for interleukin 6 (IL-6) and tumor necrosis factor-α (TNF-α), and from R&D Systems (Minneapolis, MN) for IL-1_β_ and IL-10.
Microdissection of Brain Structures
Coronal sections (300 μm) of fresh-frozen brains were cut on a cryostat, mounted on slides, and used for punch microdissection as follows (positions are relative to bregma): three sections between −1.3 and −2.2 mm for the paraventricular nucleus (PVN) and the central nucleus of the amygdala; two sections between −0.7 and −1.3 mm for the subfornical organ (SFO); three sections between −2.2 and −3.1 mm for the CA1 subdivision of the hippocampus; and three sections between 3.7 and 2.9 mm for the ventromedial prefrontal cortex (Paxinos and Watson, 1998). Punch microdissection was performed under stereomicroscope control using Harris Uni-Core microdissection needles of 0.75 mm diameter for the PVN and 1 mm for other brain structures. The individual punches dissected from the same nucleus were pooled and immediately homogenized for RNA isolation as described below. Tissues from each rat were studied individually.
Determination of mRNA Expression By Real-Time PCR
RNA was isolated by TRIzol reagent (Invitrogen, Carlsbad, CA) and purified using the RNeasy Mini kit (Qiagen, Valencia, CA). Total RNA (0.15 μg from brain tissue; 1 μg from cell lysates) was reversely transcribed into cDNA and quantified by real-time PCR as previously described (Sánchez-Lemus et al, 2009b). Primers used in the study are listed in Supplementary Table S1. Target cDNA from individual samples was normalized to GAPDH mRNA and expressed as a fold change relative to the vehicle-saline group.
Ang II Receptor Autoradiography
Binding of [125I] Sarcosine1-Ang II (ARC, St Louis, MO) to 16 μm coronal sections of fresh frozen brains was performed as described previously (Tsutsumi and Saavedra, 1991).
Localization and Quantification of mRNA Expression By In Situ Hybridization
Coronal sections (16 μm) of fresh frozen brains were cut on a cryostat, mounted on positively charged slides, dried for 5 min at 50 °C, and stored at −80 °C until used. In situ hybridization of AT1A receptor mRNA was performed using [35S]-labeled antisense and sense riboprobes corresponding to the 368 bp Pal1 fragment from the 3′-non-coding region of the rat AT1A cDNA as described previously (Jőhren et al, 1995).
In situ hybridization of c-Fos and IL-1_β_ mRNA was performed using [35S]-labeled oligonucleotide probes. Antisense probes for the rat IL-1_β_, 5′-CCACGGGCAAGACATAG GTAGCTGCCACAGCTTCTCCACAGCCAC-3′ (Kim et al, 2004), and c-Fos, 5′-GCAGCGG GAGGATGACGCCTCGTAGTCCGCGTTGAAACCCGAGAA-3′ (Vaisanen et al, 2004), and their corresponding sense probes were 3′ labeled with [_α_-35S]-ATP (NEN/Perkin Elmer, Waltham, MA) to a specific activity of 3–4 × 108 c.p.m./μg. The sections were fixed in 4% formaldehyde, acetylated, delipidated, and exposed to hybridization buffer containing 2 × 107 c.p.m./ml sense or antisense probe. After hybridization for 18 h at 40 °C, the sections were washed, dehydrated, and exposed to Biomax MR film. The optical densities of autoradiograms were quantified by comparison with [14C]-microscale standards (Tsutsumi and Saavedra, 1991) and expressed as nCi/g tissue.
Immunohistochemical Localization and Quantification of c-Fos and Microglia
Coronal sections (16 μm) from fixed brains were cut on a cryostat, mounted to positively charged slides, vacuum-dried, and stored at −80 °C until used. The sections were immersed into ice-cold acetone for 15 min, air dried, and pre-incubated 3 × 5 min in Tris-buffered saline (20 mM Tris, 136 mM NaCl, pH 7.4) containing 0.2% Triton X-100. After quenching of intracellular peroxidase and blocking of endogenous biotin, the sections were blocked with 10% of normal goat serum and exposed to primary antibody overnight at 4 °C. Rabbit polyclonal antibody against rat c-Fos (Ab-5, 1:10 000, Calbiochem, La Jolla, CA), or mouse monoclonal antibody against rat CD11b/c (Ox-42, 1:300, BD Pharmingen, San Jose, CA) were used as primary antibodies. After washing, the sections were exposed to goat biotin-conjugated antibody against rabbit or mouse IgG (1:1000, Jackson ImmunoResearch, West Grove, PA) followed by incubation with avidin and biotin-conjugated horseradish peroxidase H. The sections were exposed to DAB reagent (Vector Laboratories, Burlingame, CA), counter-stained with hematoxylin, dehydrated in ethanol, cleared with xylene, and coverslipped with mounting medium.
Data acquisition and quantification
For data acquisition and quantification, sections were examined by an investigator blinded to the protocol used and the identity of the slides. Images were acquired by Axiocam digital camera (Zeiss, Jena, Germany) connected to a Zeiss Axioskop light microscope under constant illumination. Images were processed and analyzed using Image J software (Ruifrok and Johnston, 2001). Induction of c-Fos protein was expressed as the number of c-Fos-positive cells per PVN section. Microglia activation was expressed as a proportion of Ox-42-positive area to the total area of the PVN. The increase in the proportion of the area occupied by microglia was used as a measure of activation (Milligan et al, 1991; Popovich et al, 1997).
Statistical Analysis
Results are expressed as means±SEM for groups of 5–10 animals measured individually. Results from cell culture experiments are expressed as means±SEM of three independent cell preparations. To address the differences among all individual groups, we analyzed the data by one-way ANOVA followed by the post hoc Newman–Keuls’ test for all determinations with significant omnibus ANOVA. Differences were considered significant at P<0.05.
RESULTS
Effects of LPS and an ARB on brain Ang II AT1A mRNA expression and AT1 receptor binding. When rats were studied at 3 h after LPS administration, the endotoxin upregulated AT1A receptor mRNA expression in the hypothalamic PVN but not in the SFO, and did not alter AT1 receptor binding in either structure (Supplementary Figure S1). Candesartan significantly blocked AT1 receptor binding in the PVN and SFO in both vehicle- and in LPS-treated rats (Supplementary Figure S1).
Ang II AT1 Receptor Blockade Decreases LPS-Induced Release of Proinflammatory Cytokines to the Circulation
The basal cytokine levels in plasma were below the limit of detection in vehicle-saline control rats and were not changed by AT1 receptor blockade (Figure 1). LPS markedly elevated plasma levels of the proinflammatory cytokines TNF-α, IL-6 and IL-1_β_, and the anti-inflammatory cytokine IL-10 (Figure 1). Both short term and sustained AT1 receptor blockade significantly reduced LPS-induced plasma levels of proinflammatory cytokines, but did not affect IL-10 concentrations (Figure 1).
Figure 1
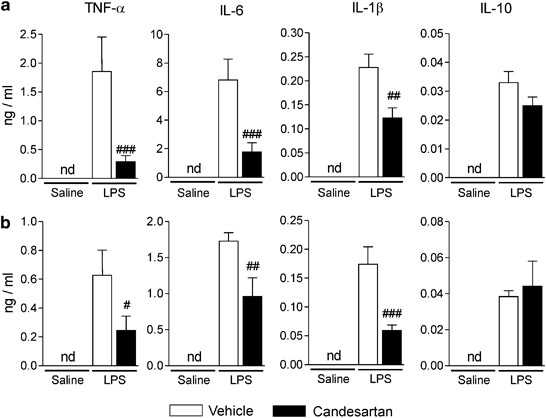
Selective decrease of LPS-induced proinflammatory cytokine release to the circulation by angiotensin II AT1 receptor blockade. Short-term (a) and sustained (b) candesartan treatments were followed by LPS or saline injection as described in Materials and Methods. The levels of cytokines in plasma 3 h after LPS or saline injection were detected by specific ELISA. Data are expressed as means±SEM (_n_=5–16). #P<0.05, ##P<0.01, ###P<0.001 compared with vehicle-LPS group; nd, non-detectable.
Ang II AT1 Receptor Blockade Reduces LPS-Induced Proinflammatory Cytokine mRNA in the Brain
LPS produced a widespread increase in TNF-α, IL-1_β_, and IL-6 mRNA expression in the PVN, SFO, ventromedial prefrontal cortex, central nucleus of the amygdala, and the CA1 subdivision of the hippocampus (Figure 2). The effects of LPS were significantly reduced by short-term AT1 receptor blockade in all areas examined (Figure 2).
Figure 2
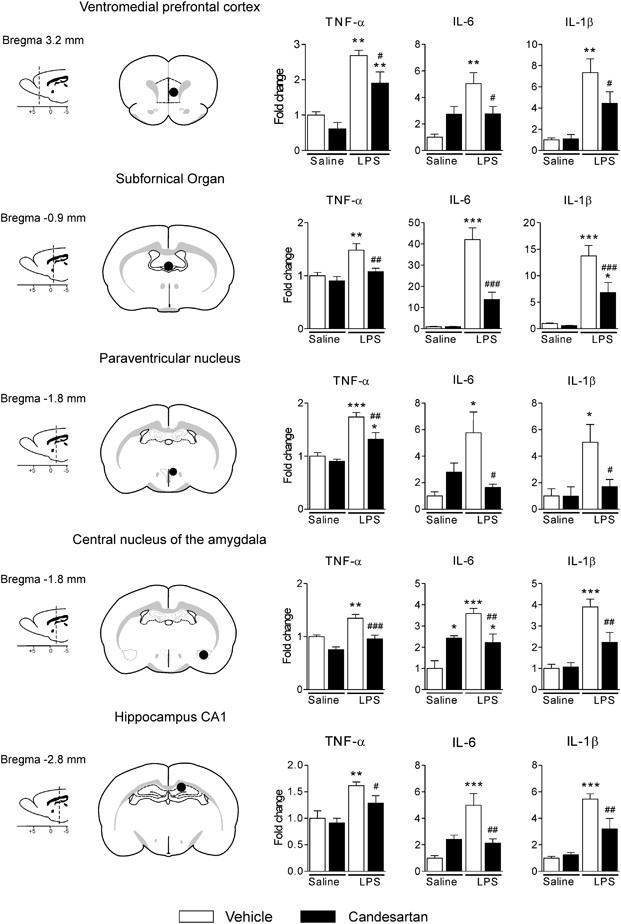
Widespread decrease of LPS-induced proinflammatory cytokine mRNA expression in brain by angiotensin II AT1 receptor blockade. Short-term treatment with candesartan was followed by LPS injection as described in Materials and Methods. Expression of mRNAs of proinflammatory cytokines was detected by real-time PCR in brain structures isolated by punch microdissection from 300 μm coronal brain sections. The scheme on the left illustrates the distance from bregma and the punch area (black circle) in the corresponding coronal sections. The expression of all genes was normalized to the level of GAPDH mRNA. Data are expressed as a fold change relative to vehicle-saline group. Data represent mean±SEM (_n_=5–8). *P<0.05, **P<0.01, ***P<0.001 compared with vehicle-saline group; #P<0.05, ##P<0.01, ###P<0.001 compared with vehicle-LPS group.
Ang II AT1 Receptor Blockade Diminishes LPS-Induced mRNA Expression of Brain Proinflammatory Factors
Short-term candesartan pretreatment significantly reduced LPS-induced increase in mRNA expression of multiple proinflammatory factors in the PVN and the SFO (Figure 3 and Supplementary Figure S2). These include: (a) the LPS recognition molecule CD14; (b) the immediate early genes c-Fos and FosB, and the inhibitor of _κ_B (I_κ_B-α), representing the activation of the nuclear factor-_κ_B (NF-_κ_B); (c) inflammatory factor receptors, TNF-α R1, TNF-α R2, IL-1 R1, IL-6R, the IL-1R antagonist, and the prostaglandin E2 (PGE2) receptor EP4; (d) the nitric oxide (NO)-producing isoenzyme inducible NO synthase (iNOS) and the PGE2-producing enzyme cyclooxygenase 2 (COX-2); (e) the adhesion molecules intercellular adhesion molecule-1 (ICAM-1) and the vascular cell adhesion molecule-1 (VCAM-1); and (f) the microglia activation marker CD40 (Figure 3 and Supplementary Figure S2). For the most part, the responses to LPS and AT1 receptor blockade were similar in the PVN and SFO, with the ARB significantly decreasing expression of LPS-induced inflammatory factor mRNA expression. A few exceptions are noted in Supplementary Figure S2.
Figure 3
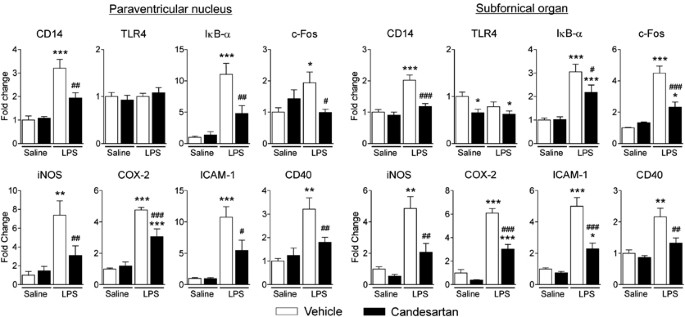
Decrease of LPS-induced inflammatory factor mRNA expression in PVN and SFO by angiotensin II AT1 receptor blockade. Short-term candesartan treatment was followed by LPS injection as described in Materials and Methods. Expression of mRNAs of proinflammatory factors and mediators was detected in microdissected PVN and SFO by real-time PCR and was normalized to the level of GAPDH mRNA. Results are expressed as a fold change relative to vehicle-saline group. Data represent mean±SEM (_n_=5–8). *P<0.05, **P<0.01, ***P<0.001, compared with vehicle-saline group; #P<0.05, ##P<0.01, ##P<0.001, compared with vehicle-LPS group.
Sustained candesartan pretreatment yielded results similar to those obtained with short-term pretreatment for all factors examined in the PVN (Supplementary Figure S3). In addition, LPS upregulated, and short-term candesartan treatment prevented, c-Fos mRNA in the PVN, SFO, and choroid plexus, and IL-1_β_ mRNA in the SFO and choroid plexus visualized by in situ hybridization. IL-1_β_ mRNA was not detected in the PVN (Figure 4a–d).
Figure 4
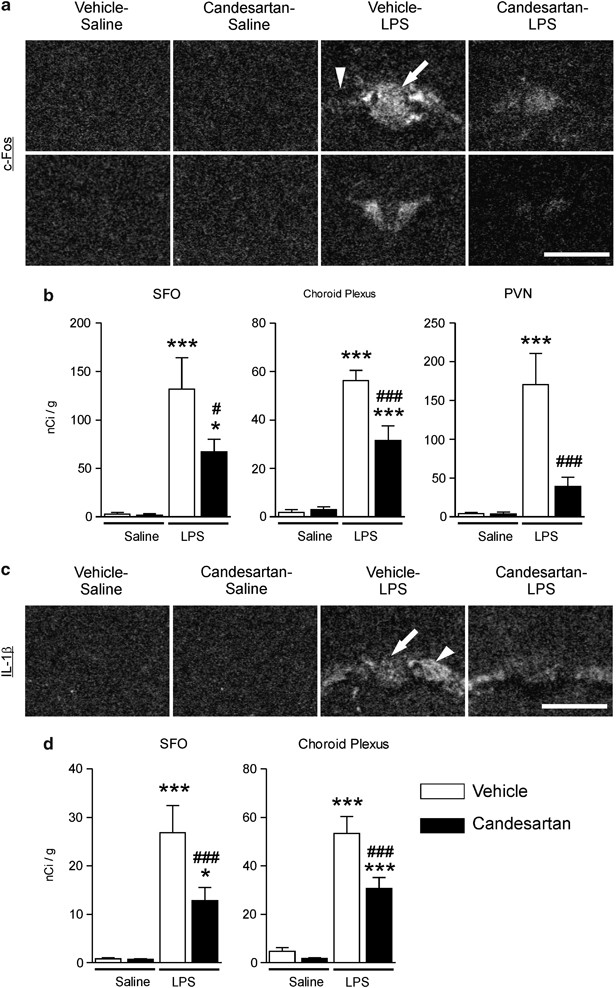
Angiotensin II AT1 receptor blockade attenuates LPS-induced expression of c-Fos mRNA in PVN, SFO, and choroid plexus and IL-1_β_ mRNA in SFO and choroid plexus. Short-term candesartan and LPS were administered as described in Materials and Methods. Expression of c-Fos and IL-1_β_ mRNAs was analyzed by in situ hybridization (a) Representative dark field images of film autoradiograms illustrating the expression of c-Fos mRNA in SFO (top) and PVN (bottom). (b) Quantitative analysis of c-Fos mRNA in SFO and PVN. (c) Representative dark field images of film autoradiograms illustrating the expression of IL-1_β_ mRNA in SFO and choroid plexus. (d) Quantitative analysis of IL-1_β_ mRNA in SFO and choroid plexus. Data are expressed as means±SEM (_n_=7–9). *P<0.05, ***P<0.001 compared with vehicle-saline group, #P<0.05, ###P<0.001 compared with vehicle-LPS group. Scale bar represents 1 mm. Arrow points to SFO; arrowhead indicates adjacent choroid plexus.
Ang II AT1 Receptor Blockade Lessens LPS-Induced c-Fos Protein Expression and Microglia Activation
Long-term ARB pretreatment suppressed, in the PVN, LPS-induced increase in the number of cells expressing c-Fos (Figure 5a and c) and microglial activation detected by OX-42 immunostaining (Figure 5b and d).
Figure 5
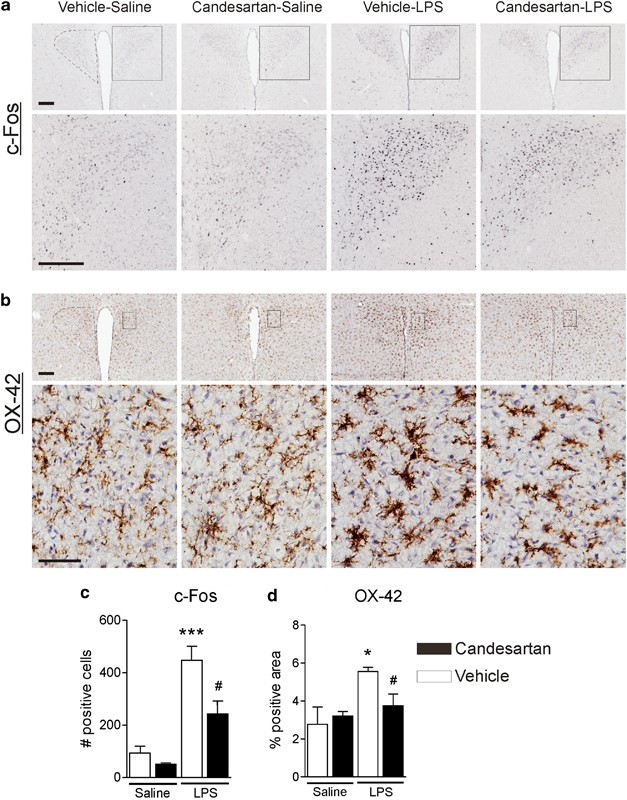
Angiotensin II AT1 receptor blockade suppresses LPS-stimulated c-Fos induction and microglia activation in the PVN. Rats were treated with candesartan for 2 weeks followed by LPS administration as described in Materials and Methods. (a) Representative photomicrographs of c-Fos immunoreactivity in PVN. Rectangle area on the top image is magnified in the image below. Scale bar represents 200 μm. Dashed line outlines the area of PVN. (c) Quantitative analysis of c-Fos immunoreactivity in the PVN. Results are expressed as a number of c-Fos-positive cells per PVN section. (b) Representative photomicrographs of microglia in PVN stained with OX-42 antibody. Dashed line in the upper image outlines the area of PVN. Rectangle area in the upper image is magnified below. Scale bars represent 200 μm in the upper image and 50 μm in the lower image. (d) Quantitative analysis of microglia activation in PVN. An increase in the proportion of area positively stained with OX-42 antibody was used as a measure of microglia activation and is expressed as a percent of total area of PVN per section. Data are shown as means±SEM (_n_=5–8). *P<0.05, ***P<0.001 compared with vehicle-saline group; #P<0.05 compared with vehicle-LPS group.
AT1 Receptor Blockade Decreases LPS-Induced Stimulation of Proinflammatory Factors in Cultured Cerebellar Granule Cells
In primary cultures of cerebellar granule cells, candesartan suppressed LPS-induced upregulation of TNF-α, IL-1_β_, and IL-6, AT1A receptor, CD14, and I_κ_B-α mRNA (Figure 6a). LPS elicited an ∼100-fold increase in TNF-α release into the cultured medium, dose dependently inhibited by candesartan (Figure 6a).
Figure 6
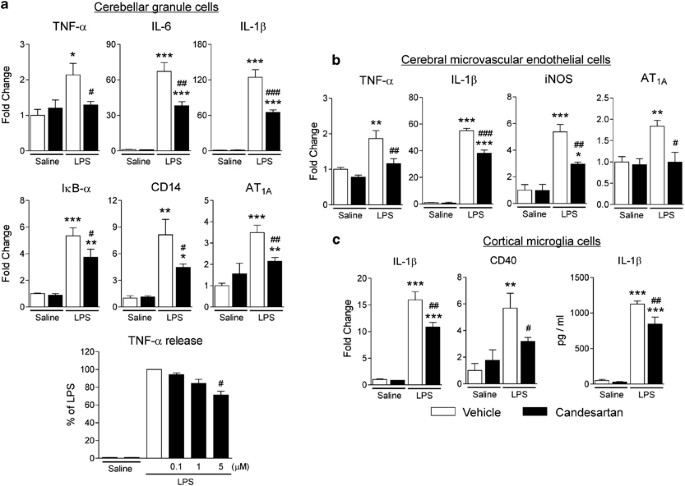
Angiotensin II AT1 receptor blockade decreases LPS-induced inflammation markers in cerebellar granule cells (a), cerebral microvascular endothelial cells (b) and cortical microglia in culture (c). Cells were isolated and cultured as described in Materials and Methods. Expression of mRNAs was detected by real-time PCR and normalized to the level of GAPDH mRNA. Results are expressed as a fold change relative to vehicle-saline group. The release of TNF-α and IL-1_β_ was determined by ELISA in the incubation media as described in Materials and Methods. Data are presented as means±SEM of three independent experiments performed in duplicates. *P<0.05, **P<0.01, ***P<0.001 compared with vehicle-saline group; #P<0.05, ##P<0.01, ###P<0.001 compared with vehicle-LPS group.
LPS Increases AT1A Receptor Transcription and AT1 Receptor Blockade Diminishes LPS-Induced iNOS and Cytokine Expression in Cultured Cerebral Microvascular Endothelial Cells
In primary cultures of rat cerebrovascular endothelial cells, LPS induced expression of AT1A receptor mRNA (Figure 6b). Incubation in the presence of candesartan decreased the LPS-induced expression of TNF-α, IL-1_β_, and iNOS mRNA (Figure 6b).
AT1 Receptor Blockade Decreases LPS-Induced Stimulation of Proinflammatory Factors in Cultured Microglial Cells
In primary cultures of cortical microglia, candesartan reduced LPS-induced upregulation of IL-1_β_ and CD40 mRNA (Figure 6c). LPS induced ∼20-fold increase in IL-1_β_ release into the cultured medium; the effect that was significantly reduced by candesartan (Figure 6c).
AT1 Receptor Blockade Reduces the Acute LPS-Induced Anorexic Effect and Body Weight Loss
Administration of LPS produced an acute anorexic effect and decrease in body weight (Figure 7a). These effects were significantly reduced by AT1 receptor blockade (Figure 7a).
Figure 7
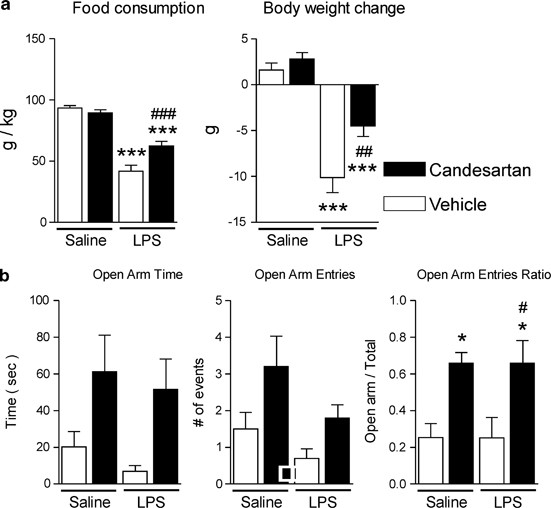
Candesartan ameliorates LPS-induced anorexia and weight loss in Wistar Hannover rats, and decreases anxiety-like behavior in SHRs. (a) Short-term candesartan and LPS were administered as described in Materials and Methods. Food consumption and body weight change were measured the day after LPS injection (_n_=8–10). (b) Short-term candesartan and LPS were administered to SHRs as described in Materials and Methods. At 3 h after LPS injection, the rats were tested on an elevated plus maze for 5 min as described in Materials and Methods (_n_=9–10). Data are expressed as means±SEM. *P<0.05, ***P<0.001 compared with vehicle-saline group, #P<0.05,##P<0.01, ###P<0.001 compared with vehicle-LPS group.
AT1 Receptor Blockade Ameliorates Anxiety in SHR
Anxiety-like behavior was determined by testing on an elevated plus maze. The time spent in the open arms and the number of entries into the open arms were not significantly affected by candesartan pre-treatment, although the consistent tendency toward the higher scores in the candesartan-treated groups was observed in both parameters (Figure 7b). The proportion of the open arm entries relative to the total number of entries was significantly increased by candesartan (Figure 7b) indicative of reduced anxiety.
DISCUSSION
Our results demonstrate widespread anti-inflammatory effects of Ang II AT1 receptor blockade in the periphery and brain. Blockade of brain AT1 receptors was achieved by systemic administration of candesartan, an ARB with central AT1 receptor blocking activity (present results; Nishimura et al, 2000; Sánchez-Lemus et al, 2009b) administered at a dose without major effects on systolic blood pressure in normotensive rats (Sánchez-Lemus et al, 2008). At this dose, candesartan prevented the consequences of not only of peripheral but also central Ang II-induced AT1 receptor stimulation (Seltzer et al, 2004). The anti-inflammatory effects of AT1 receptor blockade occurred both after short-term and sustained ARB pretreatment, indicating that the effects are not lost over time. At the early time point studied, LPS enhanced AT1A receptor transcription in the PVN but not in the SFO (present results; Sánchez-Lemus et al, 2009b) and in cultured cerebral microvascular endothelial and cerebellar granule cells.
AT1 receptor blockade reduced, as a consequence of effects on peripheral macrophages (Sánchez-Lemus et al, 2008, 2009a, 2009b), the acute induction and release to the circulation of the major proinflammatory cytokines with direct actions in the brain without changing levels of the anti-inflammatory cytokine IL-10 (Maynard and Weaver, 2008). Candesartan decreased the LPS-induced formation and release of the proinflammatory hormone aldosterone and proinflammatory cytokines while preserving endotoxin-induced release of anti-inflammatory corticosterone (Sánchez-Lemus et al, 2008). This indicates that the overall peripheral effect of ARB pretreatment is to shift the balance in favor of anti-inflammatory factors in the circulation during the initial phase of the inflammatory response.
The ARB significantly reduced the inflammation-induced mRNA expression of proinflammatory cytokines and their receptors in all brain areas examined, a widespread effect not only restricted to previously identified inflammation target areas, such as the PVN and SFO, but also present in brain structures expressing AT1 receptors (Jőhren et al, 1995; Lenkei et al, 1998) and involved in the regulation of emotion, cognition, and memory, such as the ventromedial prefrontal cortex, central nucleus of the amygdala, and CA1 subdivision of the hippocampus (McEwen, 2008).
AT1 receptor blockade reduced inflammation-induced activation of transcription factors regulating expression of multiple inflammatory genes (Licinio and Wong, 1997; Quan and Banks, 2007), including c-Fos and FosB, members of the activator protein-1 (AP-1) transcription family, and NF-_κ_B, visualized by I_κ_B-α induction, which parallels NF-_κ_B activation (Beg et al, 1993; Figure 8). This explains the inhibition not only of proinflammatory cytokines but also of enzymes, such as iNOS and COX-2, regulating production of inflammatory signals NO and prostanoids, such as PGE2 (Figure 8). Our results are consistent with the established role of NF-_κ_B as a major mediator of LPS effects in the brain (Pan et al, 2010), contradicting earlier observations reporting that intracerebral administration of LPS did not activate NF-_κ_B or AP-1 in the brain (Watanabe et al, 2006).
Figure 8
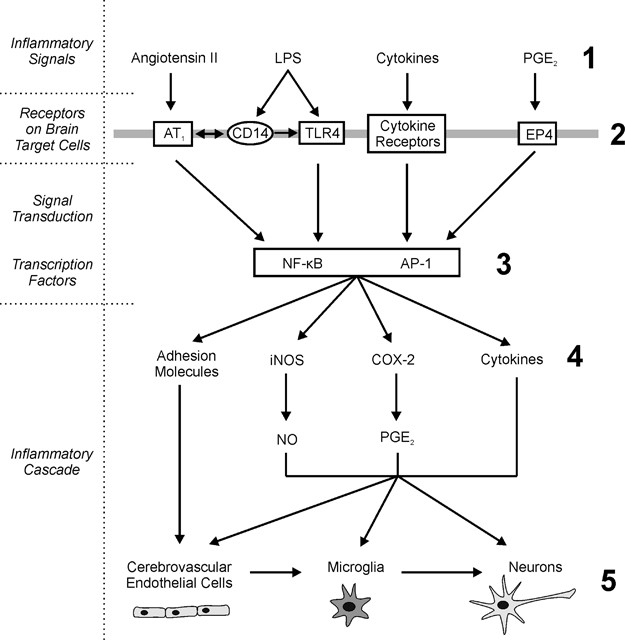
Angiotensin II and inflammatory signal pathways in the brain. AT1 receptor activity is an important factor during brain inflammation. Angiotensin II and inflammatory stimuli interact, enhancing several common downstream signaling pathways. ARBs reduce inflammatory induction of peripheral cytokines and their release to the circulation (1). Inflammatory signals (1) induce transcription of angiotensin II AT1, CD14, cytokine, and EP4 receptors in brain target cells (2). Angiotensin II AT1 receptor blockade eliminates angiotensin II-stimulated downstream signaling and represses downstream transcription factors NF-_κ_B and AP-1 (3). This limits inflammation-induced increase in brain inflammatory cascades: proinflammatory cytokines, cytokine and prostanoid receptors, inducible enzymes leading to PGE2 and NO overproduction and adhesion molecules (4). In turn, this reduces microglia activation and neuronal injury in brain parenchyma (5) limiting the brain inflammatory reaction.
Inhibition of gene expression of the PGE2 receptor EP4 and many cytokine receptors reveals that the ARB limits not only the production of inflammatory mediators but also the capacity of the cellular targets to respond to inflammation. These results suggest the possibility that ARBs may also protect brain cells from other kinds of injury. For example, the EP4 receptor has been linked to production of amyloid β peptides characteristic of Alzheimer's disease (Hoshino et al, 2009), suggesting that its repression may be therapeutically beneficial.
ARBs exert brain anti-inflammatory effects by a number of parallel mechanisms (Figure 8). By decreasing circulating proinflammatory cytokines (present results; Sánchez-Lemus et al, 2008, 2009a, 2009b), transcription of cytokine receptors, TLR4 (Dasu et al, 2009), and in particular CD14 (present results) in target areas, ARBs may reduce the capacity of the brain to recognize and respond to inflammatory factors evoked by the innate immune response. Our data suggest that the potent proinflammatory effects of Ang II in the periphery (Savoia and Schiffrin, 2007) and the brain (Zhou et al, 2005) involve sharing signal transduction pathways with inflammatory stimuli (Savoia and Schiffrin, 2007). Inflammation enhances brain AT1 receptor transcription in the PVN and directly in cultured cerebellar granule cells, suggesting a clear link between inflammatory stimuli and Ang II. A reduction of Ang II-induced inflammatory cascades is an additional mechanism explaining the overall central anti-inflammatory effects of AT1 receptor blockade.
Although the systemic administration of candesartan studied here effectively blocks both peripheral and brain AT1 receptors and the effects of centrally administered Ang II (Nishimura et al, 2000; Seltzer et al, 2004), we cannot conclusively determine the relative role of central and peripheral AT1 receptors in our model. To demonstrate that the changes observed in the brain may not exclusively depend on cross-system interactions, we studied the effects of LPS and AT1 receptor blockade at the cellular level. We used primary cultures of isolated brain neurons, microglia, and endothelial cells, primary targets for circulating inflammatory cytokines and LPS (Quan and Banks, 2007). AT1 receptor blockade exerted direct anti-inflammatory effects in a cerebellar cell culture highly enriched in granule cell neurons (Gao et al, 1995) and constitutively expressing AT1A receptors, CD14, and TLR4. In addition, AT1 receptor blockade reduced inflammation in cultured cerebral microvascular endothelial cells. These cells are involved in inflammatory leukocyte infiltration into the brain (Zhou et al, 2006; Man et al, 2007). Blockade of AT1 receptors in cerebrovascular endothelial cells (Zhou et al, 2006) may reduce Ang II-mediated increase in blood brain barrier permeability and peripheral proinflammatory cell migration to the vasculature or brain parenchyma, explaining the beneficial effects of ARBs ameliorating brain inflammation in hypertension and diabetes (Fleegal-DeMotta et al, 2009). This anti-inflammatory activity is most likely sequentially replicated in adjacent neighboring microglia and neuronal cells. Stimulation of inflammatory cascades at endothelial target cells is followed by activation of microglia (Figure 8), the central resident immune cells (Henry et al, 2009), leading to induction of additional inflammatory cascades and ultimately producing, if unchecked, glial and neuronal damage (Henry et al, 2009). In particular, microglia activation increases LPS-induced neurotoxicity and consequently, the reduction of the proinflammatory microglia response enhances neuronal survival (Liu et al, 2010). We demonstrate here that the ARB candesartan inhibited the microglia activation marker CD40 and IL-1_β_ mRNA expression and release in an enriched microglia cells culture. This is consistent with our finding of decreased microglia activation in vivo as a result of systemic candesartan administration, and it is in agreement with the previous observation of inhibition of inflammation-induced microglia activation, in vitro, by the ARB losartan (Miyoshi et al, 2008). Reduction of microglia activation might be another mechanism of ARBs promoting neuroprotection.
We report here direct anti-inflammatory effects of candesartan in LPS brain target cells. Our results do not exclude a major influence of peripheral anti-inflammatory effects in the candesartan-induced amelioration of brain inflammation. However, the in vitro experiments demonstrate that candesartan is able to directly and significantly reduce inflammation in LPS target cells in the brain. This suggests that direct central effects of candesartan may have a role in the amelioration of brain inflammation, which follows systemic administration of the ARB.
The central anti-inflammatory effects of the ARB have behavioral correlates as we demonstrate here. Candesartan reduced the acute sickness behavior produced by LPS, in particular its anorexic effect leading to acute weight loss (Konsman and Dantzer, 2001). In addition, candesartan ameliorated anxiety-like behavior in SHR, a strain with known hypersensitivity to stress and to LPS administration (Kvetnansky et al, 1979; Sirén et al, 1992). These effects were observed in both control and LPS-treated animals, indicating general anxiolytic properties that may be independent on the effect of LPS. Anxiolytic effects have been previously reported in normotensive rats exposed to prolonged treatment with candesartan (Saavedra et al, 2006), and both normotensive and hypertensive rats after acute treatment with another ARB, losartan (Srinivasan et al, 2003). Acute anxiolytic effects observed 5 min after intracerebroventricular injection of losartan (Kulakowska et al, 1996) further point to the direct involvement of brain Ang II AT1 receptors in these behavioral changes.
In conclusion, our results demonstrate that direct brain inflammation, and peripheral inflammation affecting the brain are, to a significant degree, dependent on full AT1 receptor function. ARBs limit, but do not eliminate the response of the brain to inflammatory signals. This is an important finding, as responding to inflammation is necessary to maintain and restore homeostasis, and pathological conditions may further develop as a consequence of unrestricted inflammation (Dantzer et al, 2008).
This report provides a basis to support recent findings in clinically relevant neuroinflammatory models. For example, brain RAS is upregulated in an animal model of multiple sclerosis, the experimental autoimmune encephalomyelitis, and ARBs ameliorate the characteristic paralysis and infiltration of inflammatory TH1 and TH17 cells in this model (Platten et al, 2009). Further evidence indicates that ARBs protect the brain from injuries promoting parenchymal inflammation and neuronal damage, such as irradiation-induced brain inflammation (Jenrow et al, 2010) and inflammation-induced dopaminergic neuronal death (Mertens et al, 2010).
Our results have major clinical significance. Excessive, uncontrolled inflammation is recognized as a major factor contributing to many psychiatric and neurological diseases, including but not limited to major depression and anxiety disorders, bipolar illness, schizophrenia, and neurodegenerative diseases such as Parkinson's and Alzheimer's disease (Dantzer et al, 2008; Doorduin et al, 2009; Glass et al, 2010; Fassbender et al, 2004; Hope et al, 2009; Miller et al, 2009). ARBs are compounds with excellent margin of safety, successfully used for years to treat not only increased systemic blood pressure but also to ameliorate the profound inflammatory component of peripheral cardiovascular disease and diabetes (Savoia and Schiffrin, 2007). We have previously found that ARBs reverse the cerebrovascular inflammation characteristic of genetic hypertension in an animal model (Ando et al, 2004; Zhou et al, 2005), and we demonstrate here that these compounds are very effective anti-inflammatory agents in the brain parenchyma of normotensive rats.
Our proposal to test ARBs for the therapy of inflammatory diseases of the brain is sustained by substantial clinical evidence indicating that ARBs are neuroprotective in humans (Anderson, 2010). ARBs prevent and ameliorate stroke in hypertensive patients to a greater degree than other similarly potent anti-hypertensive medications, in part as a consequence of effects beyond blood pressure control (Anderson, 2010). ARBs protect cognition both in hypertensive patients and in the elderly, significantly reducing the incidence and progression of Alzheimer's disease (Li et al, 2010). Other studies reported improvement in the quality of life, and amelioration of depression and stress (Phillips and de Oliveira, 2008). ARBs may represent a novel and safe approach for the treatment of acute and chronic neuropsychiatric disorders of increasing prevalence and socioeconomic impact where inflammation has a significant role.
References
- Anderson C (2010). More direct evidence of potential neuroprotective benefits of Angiotensin receptor blockers. Hypertension 28: 429.
Article CAS Google Scholar - Ando H, Zhou J, Macova M, Imboden H, Saavedra JM (2004). Angiotensin II AT1 receptor blockade reverses pathological hypertrophy and inflammation in brain microvessels of spontaneously hypertensive rats. Stroke 35: 1726–1731.
Article CAS Google Scholar - Beg AA, Finco TS, Nantermet PV, Baldwin Jr AS (1993). Tumor necrosis factor and interleukin-1 lead to phosphorylation and loss of IκBα: a mechanism for NF-κB activation. Mol Cell Biol 13: 3301–3310.
Article CAS Google Scholar - Benicky J, Sánchez-Lemus E, Pavel J, Saavedra JM (2009). Anti-inflammatory effects of angiotensin receptor blockers in the brain and the periphery. Cell Mol Neurobiol 29: 781–792.
Article CAS Google Scholar - Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW (2008). From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci 9: 46–57.
Article CAS Google Scholar - Dasu MR, Riosvelasco AC, Jialal I (2009). Candesartan inhibits toll-like receptor expression and activity both in vitro and in vivo. Atherosclerosis 202: 76–83.
Article CAS Google Scholar - Doorduin J, de Vries EF, Willemsen AT, de Groot JC, Dierckx RA, Klein HC (2009). Neuroinflammation in schizophrenia-related psychosis: a PET study. J Nucl Med 50: 1801–1807.
Article Google Scholar - Fassbender K, Walter S, Kühl S, Landmann R, Ishii K, Bertsch T et al (2004). The LPS receptor (CD14) links innate immunity with Alzheimer's disease. FASEB J 18: 203–205.
Article CAS Google Scholar - Fleegal-DeMotta MA, Doghu S, Banks WA (2009). Angiotensin II modulates BBB permeability via activation of the AT(1) receptor in brain endothelial cells. J Cereb Blood Flow Metab 29: 640–647.
Article CAS Google Scholar - Gao X-M, Margolis RL, Leeds P, Hough C, Post RM, Chuang DM (1995). Carbamazepine induction of apoptosis in cultured cerebellar neurons: effects of N-methyl-D-aspartate, aurintricarboxylic acid and cycloheximide. Brain Res 703: 63–71.
Article CAS Google Scholar - Glass CK, Saijo K, Winner B, Marchetto MC, Gage FH (2010). Mechanisms underlying inflammation in neurodegeneration. Cell 140: 918–934.
Article CAS Google Scholar - Giulian D, Baker TJ (1986). Characterization of ameboid microglia isolated from developing mammalian brain. J Neurosci 6: 2163–2178.
Article CAS Google Scholar - Henry CJ, Huang Y, Wynne AM, Godbout JP (2009). Peripheral lipopolysaccharide (LPS) challenge promotes microglial hyperactivity in aged mice that is associated with exaggerated induction of both pro-inflammatory IL-1b and anti-inflammatory IL-10 cytokines. Brain Behav Immun 23: 309–317.
Article CAS Google Scholar - Hope S, Melle I, Aukrust P, Steen NE, Birkenaes AB, Lorentzen S et al (2009). Similar immune profile in bipolar disorder and schizophrenia: selective increase in soluble tumor necrosis factor receptor I and von Willebrand factor. Bipolar Disord 11: 726–734.
Article CAS Google Scholar - Hoshino T, Namba T, Takehara M, Nakaya T, Sugimoto Y, Araki W et al (2009). Prostaglandin E2 stimulates the production of amyloid-beta peptides through internalization of the EP4 receptor. J Biol Chem 284: 18493–18502.
Article CAS Google Scholar - Jenrow KA, Brown SL, Liu J, Kolozsvary A, Lapanowski K, Kim JH (2010). Ramipril mitigates radiation-induced impairment of neurogenesis in the rat dentate gyrus. Radiat Oncol 5: 6.
Article Google Scholar - Jőhren O, Inagami T, Saavedra JM (1995). AT1A, AT1B, and AT2 angiotensin II receptor subtype gene expression in rat brain. Neuroreport 6: 2549–2552.
Article Google Scholar - Kim YS, Honkaniemi J, Sharp FR, Tauber MG (2004). Expression of proinflammatory cytokines tumor necrosis factor-α and interleukin-1β in the brain during experimental group B streptococcal meningitis. Brain Res Mol Brain Res 128: 95–102.
Article CAS Google Scholar - Konsman JP, Dantzer R (2001). How the immune and nervous system interact during disease-associated anorexia. Nutrition 17: 664–668.
Article CAS Google Scholar - Kulakowska A, Karwowska W, Wisniewski K, Braszko JJ (1996). Losartan influences behavioural effects of angiotensin II in rats. Pharmacol Res 34: 109–115.
Article CAS Google Scholar - Kvetnansky R, McCarty R, Thoa NB, Lake CR, Kopin IJ (1979). Sympatho-adrenal responses of spontaneously hypertensive rats to immobilization stress. Am J Physiol 236: H457–H462.
CAS PubMed Google Scholar - Larrayoz IM, Pang T, Benicky J, Pavel J, Sánchez-Lemus E, Saavedra JM (2009). Candesartan reduces the innate immune response to lipopolysaccharide in human monocytes. J Hypertens 27: 2365–2376.
Article CAS Google Scholar - Lenkei Z, Palkovits M, Corvol P, Llorens-Cortes C (1998). Distribution of angiotensin type-1 receptor messenger RNA expression in the adult rat brain. Neuroscience 82: 827–841.
Article CAS Google Scholar - Li NC, Lee A, Whitmer RA, Kivipelto M, Lawler E, Kazis LE et al (2010). Use of angiotensin receptor blockers and risk of dementia in a predominantly male population: prospective cohort analysis. BMJ 340: b5465.
Article Google Scholar - Licinio J, Wong M-L (1997). Pathways and mechanisms for cytokine signaling of the central nervous system. J Clin Invest 100: 2941–2947.
Article CAS Google Scholar - Liu Y, Lo YC, Qian L, Crews FT, Wilson B, Chen HL et al (2010). Verapamil protects dopaminergic neuron damage through a novel anti-inflammatory mechanism by inhibition of microglial activation. Neuropharmacology, doi:10.1016/j.neuropharm2010.10.002 (in press).
- Man S, Ubogu EE, Ransohoff RM (2007). Inflammatory cell migration into the central nervous system: a few new twists on an old tale. Brain Pathol 17: 243–250.
Article CAS Google Scholar - Maynard CL, Weaver CT (2008). Diversity in the contribution of interleukin-10 to T-cell-mediated immune regulation. Immunol Rev 226: 219–233.
Article CAS Google Scholar - McEwen BS (2008). Central effects of stress hormones in health and disease: understanding the protective and damaging effects of stress and stress mediators. Eur J Pharmacol 583: 174–185.
Article CAS Google Scholar - Mertens B, Vanderheyden P, Michotte Y, Sarre S (2010). The role of the central renin-angiotensin system in Parkinson's disease. J Renin Angiotensin Aldosterone Syst 11: 49–56.
Article CAS Google Scholar - Miller AH, Maletic V, Raison CL (2009). Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol Psych 65: 732–741.
Article CAS Google Scholar - Milligan CE, Cunningham TJ, Levitt P (1991). Differential immunochemical markers reveal the normal distribution of brain macrophages and microglia in the developing rat brain. J Comp Neurol 314: 125–135.
Article CAS Google Scholar - Miyoshi M, Miyano K, Moriyama N, Taniguchi M, Watanabe T (2008). Angiotensin type 1 receptor antagonist inhibits lipopolysaccharide-induced stimulation of rat microglial cells by suppressing nuclear factor jB and activator protein-1 activation. Eur J Neurosci 27: 343–351.
Article Google Scholar - Nakagawa S, Deli MA, Kawaguchi H, Schimizudani T, Shimono T, Kittel A et al (2009). A new blood-brain barrier model using primary rat brain endothelial cells, pericytes and astrocytes. Neurochem Int 54: 253–263.
Article CAS Google Scholar - Nimmo AJ, Vink R (2009). Recent patents in CNS drug discovery: the management of inflammation in the central nervous system. Recent Pat CNS Drug Discov 4: 86–95.
Article CAS Google Scholar - Nishimura Y, Ito T, Hoe K, Saavedra JM (2000). Chronic peripheral administration of the angiotensin II AT(1) receptor antagonist candesartan blocks brain AT(1) receptors. Brain Res 871: 29–38.
Article CAS Google Scholar - Pan W, Yu C, Hsuchou H, Kastin AJ (2010). The role of cerebral vascular NFkappaB in LPS-induced inflammation: differential regulation of efflux transporter and transporting cytokine receptors. Cell Physiol Biochem 25: 623–630.
Article CAS Google Scholar - Paxinos G, Watson C (1998). The Rat Brain in Stereotaxic Coordinates, 4th edn. Academic Press: New York.
Google Scholar - Phillips MI, de Oliveira EM (2008). Brain renin angiotensin in disease. J Mol Med 86: 715–722.
Article CAS Google Scholar - Platten M, Youssef S, Hur EM, Ho PP, Han MH, Lanz TV et al (2009). Blocking angiotensin-converting enzyme induces potent regulatory T cells and modulates TH1- and TH17-mediated autoimmunity. Proc Nat Acad Sci USA 106: 14948–14953.
Article CAS Google Scholar - Popovich PG, Wei P, Stokes BT (1997). Cellular inflammatory response after spinal cord injury in Sprague-Dawley and Lewis rats. J Comp Neurol 377: 443–464.
Article CAS Google Scholar - Quan N, Banks WA (2007). Brain-immune communication pathways. Brain Behav Immun 21: 727–735.
Article CAS Google Scholar - Rivest S (2009). Regulation of innate immune responses in the brain. Nat Rev Immunol 9: 429–439.
Article CAS Google Scholar - Ruifrok AC, Johnston DA (2001). Quantification of histochemical staining by color deconvolution. Anal Quant Cytol Histol 23: 291–299.
CAS PubMed Google Scholar - Russell VA (2003). Dopamine hypofunction possibly results from a defect in glutamate-stimulated release of dopamine in the nucleus accumbens shell of a rat model for attention deficit hyperactivity disorder—the spontaneously hypertensive rat. Neurosci Biobehav Rev 27: 671–682.
Article CAS Google Scholar - Saavedra JM (1992). Brain and pituitary angiotensin. Endocr Rev 13: 329–380.
Article CAS Google Scholar - Saavedra JM, Armando I, Bregonzio C, Juorio A, Macova M, Pavel J et al (2006). A centrally acting, anxiolytic angiotensin II AT1 receptor antagonist prevents the isolation stress-induced decrease in cortical CRF1 receptor and benzodiazepine binding. Neuropsychopharm 31: 1123–1134.
Article CAS Google Scholar - Sánchez-Lemus E, Benicky J, Pavel J, Larrayoz IM, Zhou J, Baliova M et al (2009a). Angiotensin II AT1 blockade reduces the lipopolysaccharide-induced innate immune response in rat spleen. Am J Physiol Regul Integr Comp Physiol 296: R1376–R1384.
Article Google Scholar - Sánchez-Lemus E, Benicky J, Pavel J, Saavedra JM (2009b). In vivo angiotensin II AT(1) receptor blockade selectively inhibits LPS-induced innate immune response and ACTH release in rat pituitary gland. Brain Behav Immun 23: 945–957.
Article Google Scholar - Sánchez-Lemus E, Murakami Y, Larrayoz-Roldan IM, Moughamian AJ, Pavel J, Nishioku T et al (2008). Angiotensin II AT1 receptor blockade decreases lipopolysaccharide-induced inflammation in the rat adrenal gland. Endocrinology 149: 5177–5188.
Article Google Scholar - Savoia C, Schiffrin EL (2007). Vascular inflammation in hypertension and diabetes: molecular mechanisms and therapeutic interventions. Clin Sci 112: 375–384.
Article CAS Google Scholar - Schmerbach K, Schefe JH, Krikov M, Müller S, Villringer A, Kintscher U et al (2008). Comparison between single and combined treatment with candesartan and pioglitazone following transient focal ischemia in rat brain. Brain Res 1208: 225–233.
Article CAS Google Scholar - Seeger H, Lippert C, Wallwiener D, Mueck AO (2001). Valsartan and candesartan can inhibit deteriorating effects of angiotensin II on coronary endothelial function. J Renin Angiotensin Aldosterone Syst 2: 141–143.
Article CAS Google Scholar - Seltzer A, Bregonzio C, Armando I, Baiardi G, Saavedra JM (2004). Oral administration of an AT1 receptor antagonist prevents the central effects of angiotensin II in spontaneously hypertensive rats. Brain Res 1028: 9–18.
Article CAS Google Scholar - Sirén AL, Heldman E, Doron D, Lysko PG, Yue TL, Liu Y et al. (1992). Release of proinflammatory and prothrombotic mediators in the brain and peripheral circulation in spontaneously hypertensive and normotensive Wistar-Kyoto rats. Stroke 23: 1643–1650.
Article Google Scholar - Srinivasan J, Suresh B, Ramanathan M (2003). Differential anxiolytic effect of enalapril and losartan in normotensive and renal hypertensive rats. Physiol Behavior 78: 585–591.
Article CAS Google Scholar - Tsutsumi K, Saavedra JM (1991). Characterization and development of angiotensin II receptor subtypes (AT1 and AT2) in rat brain. Am J Physiol 261: R209–R216.
CAS PubMed Google Scholar - Vaisanen J, Ihalainen J, Tanila H, Castrén E (2004). Effects of NMDA-receptor antagonist treatment on c-fos expression in rat brain areas implicated in schizophrenia. Cell Mol Neurobiol 24: 769–780.
Article Google Scholar - Watanabe K, Taniguchi M, Miyoshi M, Shimizu H, Imoto T, Sato K et al (2006). Effects of central injection of angiotensin-converting-enzyme inhibitor and Angiotensin type 1 receptor antagonist on the brain NF-kappaB and AP-1 activities of rats given LPS. Peptides 27: 1538–1546.
Article CAS Google Scholar - Weinberg AJ, Zappe DH, Ashton AA, Weinberg MS (2004). Safety and tolerability of high-dose angiotensin receptor blocker therapy in patients with chronic kidney disease: a pilot study. Am J Nephrol 24: 340–345.
Article CAS Google Scholar - Yong VW, Rivest S (2009). Taking advantage of the systemic immune system to cure brain diseases. Neuron 64: 55–60.
Article CAS Google Scholar - Zhou J, Ando H, Macova M, Dou J, Saavedra JM (2005). Angiotensin II AT1 receptor blockade abolishes brain microvascular inflammation and heat shock protein responses in hypertensive rats. J Cereb Blood Flow Metab 25: 878–886.
Article CAS Google Scholar - Zhou J, Pavel J, Macova M, Yu ZX, Imboden H, Ge L et al (2006). AT1 receptor blockade regulates the local angiotensin II system in cerebral microvessels from spontaneously hypertensive rats. Stroke 37: 1271–1276.
Article CAS Google Scholar
Acknowledgements
This study was supported by the Division of Intramural Research Programs, National Institute of Mental Health, Department of Health and Human Services, USA. Dr Honda was supported by the Japan Society for the Promotion of Science Research Fellowship for Japanese Biomedical and Behavioral Researchers at NIH. We thank Astra-Zeneca, Mőlndal, Sweden, for the supply of candesartan, and to Andrés Negro Vilar MD, PhD, for his helpful comments and suggestions during the preparation of the manuscript.
Author information
Author notes
- Julius Benicky, Enrique Sánchez-Lemus and Masaru Honda: These authors contributed equally to this work.
Authors and Affiliations
- Division of Intramural Research Programs, Department of Health and Human Services, Section on Pharmacology, National Institute of Mental Health, National Institutes of Health, Bethesda, MD, USA
Julius Benicky, Enrique Sánchez-Lemus, Masaru Honda, Tao Pang, Martina Orecna, Juan Wang & Juan M Saavedra - Division of Intramural Research Programs, Department of Health and Human Services, Molecular Neurobiology Section, National Institute of Mental Health, National Institutes of Health, Bethesda, MD, USA
Yan Leng & De-Maw Chuang
Authors
- Julius Benicky
- Enrique Sánchez-Lemus
- Masaru Honda
- Tao Pang
- Martina Orecna
- Juan Wang
- Yan Leng
- De-Maw Chuang
- Juan M Saavedra
Corresponding author
Correspondence toJulius Benicky.
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
Supplementary information
PowerPoint slides
Rights and permissions
About this article
Cite this article
Benicky, J., Sánchez-Lemus, E., Honda, M. et al. Angiotensin II AT1 Receptor Blockade Ameliorates Brain Inflammation.Neuropsychopharmacol 36, 857–870 (2011). https://doi.org/10.1038/npp.2010.225
- Received: 12 August 2010
- Revised: 28 October 2010
- Accepted: 12 November 2010
- Published: 08 December 2010
- Issue Date: March 2011
- DOI: https://doi.org/10.1038/npp.2010.225