Serological and molecular detection of Neospora caninum and Toxoplasma gondii in human umbilical cord blood and placental tissue samples (original) (raw)
Abstract
Neosporosis primarily affects cattle and dogs and is not currently considered a zoonotic disease. Toxoplasmosis is a zoonosis with a worldwide distribution that is asymptomatic in most cases, but when acquired during pregnancy, it can have serious consequences. The seropositivity rates determined by the indirect fluorescent antibody test for Neospora caninum (N. caninum) and Toxoplasma gondii (T. gondii) were 24.3% (49 samples) and 26.8% (54 samples), respectively. PCR positivity for N. caninum was observed in two samples of cord blood (1%) using the Nc5 and ITS1 gene, positivity for T. gondii was observed in 16 samples using the primer for the B1 gene (5.5% positivity in cord blood and 2.5% positivity in placental tissue). None of the samples showed structures characteristic of tissue cysts or inflammatory infiltrate on histopathology. Significant associations were observed only between N. caninum seropositivity and the presence of domestic animals (p = 0.039) and presence of dogs (p = 0.038) and between T. gondii seropositivity and basic sanitation (p = 0.04). This study obtained important findings regarding the seroprevalence and molecular detection of N. caninum and T. gondii in pregnant women; however, more studies are necessary to establish a correlation between risk factors and infection.
Similar content being viewed by others
Introduction
Neospora caninum is an obligate intracellular parasite belonging to the phylum Apicomplexa and was first identified in 1984 in the central nervous system and skeletal muscle of dogs in Norway1. N. caninum has a wide range of hosts2,[3](/articles/s41598-020-65991-1#ref-CR3 "Dubey, J. P. & Lindsay, D. S. A review of Neospora caninum and neosporosis. Vet. Parasitol. 67, 1–59, https://doi.org/10.1016/s0304-4017(96)01035-7
(1996)."), but neosporosis is a disease that primarily affects cattle and dogs, and canids are definitive hosts. The forms of infection are essentially the same as those of toxoplasmosis, occurring horizontally in herbivores via intake of water or foods contaminated by oocysts and in carnivores via ingestion of tissues infected with tachyzoites or tissue cysts. Vertical transmission may also occur, and _N. caninum_ is very efficiently transplacentally transmitted in cattle, which may cause abortion[2](/articles/s41598-020-65991-1#ref-CR2 "Dubey, J. P., Schares, G. & Ortega-Mora, L. M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 20, 323–67, 10.1128%2FCMR.00031-06 (2007).") or birth of infected and asymptomatic calves[2](/articles/s41598-020-65991-1#ref-CR2 "Dubey, J. P., Schares, G. & Ortega-Mora, L. M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 20, 323–67, 10.1128%2FCMR.00031-06 (2007)."),[3](/articles/s41598-020-65991-1#ref-CR3 "Dubey, J. P. & Lindsay, D. S. A review of Neospora caninum and neosporosis. Vet. Parasitol. 67, 1–59,
https://doi.org/10.1016/s0304-4017(96)01035-7
(1996).").Toxoplasma gondii can infect all warm-blooded animals, including mammals, birds, and humans[4](/articles/s41598-020-65991-1#ref-CR4 "Tenter, A. M., Heckeroth, A. R. & Weiss, L. M. Toxoplasma gondii: from animals to humans. Int. J. Parasitol. 30, 1217–58, https://doi.org/10.1016/S0020-7519(00)00124-7
(2000)."). Toxoplasmosis is an infection caused by the parasite _T. gondii_ and may be congenital or acquired[5](/articles/s41598-020-65991-1#ref-CR5 "Dubey, J. P. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 28, 1019–24,
https://doi.org/10.1016/S0020-7519(98)00023-X
(1998)."). Intake of oocysts present in the environment and consumption of undercooked meat infected with tissue cysts are the two main forms of transmission in acquired infection[5](/articles/s41598-020-65991-1#ref-CR5 "Dubey, J. P. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 28, 1019–24,
https://doi.org/10.1016/S0020-7519(98)00023-X
(1998)."),[6](/articles/s41598-020-65991-1#ref-CR6 "Dubey, J. P., Lago, E. G., Gennari, S. M., Su, C. & Jones, J. L. Toxoplasmosis in humans and animals in Brazil: high prevalence, high burden of disease, and epidemiology. Parasitol. 139, 1375–1424,
https://doi.org/10.1017/S0031182012000765
(2012)."). Congenital transmission occurs after primary infection during pregnancy[7](/articles/s41598-020-65991-1#ref-CR7 "Lindsay, D. S. & Dubey, J. P. Toxoplasma gondii: the changing paradigm of congenital toxoplasmosis. Parasitol. 138, 1829–31,
https://doi.org/10.1017/S0031182011001478
(2011).").The infection in most cases is asymptomatic, the mother develops temporary parasitemia. However, focal lesions can develop in the placenta, and the fetus may be infected. Slightly diminished vision is characteristic of mild disease, whereas severely all children may present with retinochoroiditis, hydrocephalus, seizures and intracerebral calcification[8](/articles/s41598-020-65991-1#ref-CR8 "Hill, D. & Dubey, J. P. Toxoplasma gondii: transmission, diagnosis and prevention. Clin. Microbiol. Infect. 8, 634–40, https://doi.org/10.1046/j.1469-0691.2002.00485.x
(2002).").The diseases caused by T. gondii and N. caninum have similar characteristics, such as neurological conditions and reproductive pathologies, due to the morphological, genetic and immunological similarities of the two parasites[9](/articles/s41598-020-65991-1#ref-CR9 "Hemphill, A., Fuchs, N., Sonda, S. & Hehl, A. The antigenic composition of Neospora caninum. Int. J. Parasitol. 29, 1175–88, https://doi.org/10.1016/S0020-7519(99)00085-5
(1999)."),[10](/articles/s41598-020-65991-1#ref-CR10 "Barratt, J. L. N., Harkness, J., Marriott, D., Ellis, J. T. & Stark, D. Importance of nonenteric protozoan infections in immunocompromised people. Clin. Microbiol. Rev. 23, 795–836, 10.1128%2FCMR.00001-10 (2010).").The pathological, immunological and epidemiological aspects of neosporosis in human pregnancies are still unknown, since viable N. caninum has not been isolated from human tissues so far. However, knowing that this parasite has a wide range of intermediate hosts2,[3](/articles/s41598-020-65991-1#ref-CR3 "Dubey, J. P. & Lindsay, D. S. A review of Neospora caninum and neosporosis. Vet. Parasitol. 67, 1–59, https://doi.org/10.1016/s0304-4017(96)01035-7
(1996)."), the possibility of human infection should not be ruled out. If there is a possibility of vertical transmission in humans, we believe that the evolution and severity of the infection is dependent on the mother’s gestational age and the virulence of the strain causing the infection, as occurs in other animal species[11](/articles/s41598-020-65991-1#ref-CR11 "Buxton, D., McAllister, M. M. & Dubey, J. P. The comparative pathogenesis of neosporosis. Trends Parasitol. 18, 546–52,
https://doi.org/10.1016/S1471-4922(02)02414-5
(2002)."),[12](/articles/s41598-020-65991-1#ref-CR12 "Al-Qassab, S. E., Reichel, M. P. & Ellis, J. T. On the biological and genetic diversity in Neospora caninum. Diversity. 2, 411–38,
https://doi.org/10.3390/d2030411
(2010).").Anti-N. caninum antibodies have been reported in humans in several studies13,[14](#ref-CR14 "Ibrahim, H. M. et al. Prevalence of Neospora caninum and Toxoplasma gondii antibodies in northern Egypt. Am. J. Trop. Med. hyg. 80, 263–67, https://doi.org/10.4269/ajtmh.2009.80.263
(2009)."),[15](#ref-CR15 "Tranas, J., Heinzen, R. A., Weiss, L. M. & McAllister, M. M. Serological evidence of human infection with the protozoan Neospora caninum. Clin. Diagn. Lab. Immunol. 6, 765–67 (1999)."),[16](/articles/s41598-020-65991-1#ref-CR16 "Oshiro, L. M. et al. Neospora caninum and Toxoplasma gondii serodiagnosis in human immunodeficiency virus carriers. Rev. Soc. Bras. Med. Trop. 48, 568–72,
https://doi.org/10.1590/0037-8682-0151-2015
(2015)."), and its zoonotic potential is still uncertain. Studies conducted with human placental tissue and umbilical cord blood for detecting _N. caninum_ remain scarce in the literature. Therefore, the objective of this study was the molecular and serological detection of _N. caninum_ and/or _T. gondii_ in blood samples from the umbilical cords and placental tissues of pregnant women.Results
The pregnant women who participated in the study had a mean age of 27.5 ± 6.022 years and were at a gestational age of 39 ± 1.4 weeks, and 13.9% of the women did not have appropriate prenatal care (28/201) as recommended by the competent organs of Brazil, which recommends six or more prenatal visits.
Serology
Of the 201 samples analyzed, 24.3% were positive for IgG anti-N. caninum antibodies (Table 1), and no sample was positive for IgM antibodies. For T. gondii, 26.8% of the samples were positive for the presence of IgG antibodies (Table 2), and no sample was positive for IgM antibodies. Of all samples analyzed, 8.4% presented seropositivity for both parasites. Western blot positive samples corroborate IFAT results, showing reactivity with 29 kDa protein (Supplementary Fig. S1). Positive samples for N. caninum in PCR, despite not being positive by IFAT, were western blot positive.
Table 1 IFAT for IgG anti-N. caninum antibodies in cord serum.
Table 2 IFAT for IgG anti_-T. gondii_ antibodies in cord serum.
Statistical analyses of the N. caninum data showed significant associations (p < 0.05) between seropositivity and the presence of domestic animals and the presence of dogs. For T. gondii, a significant association (p < 0.05) between seropositivity and basic sanitation (Table 3) was observed.
Table 3 Seroprevalence of IgG antibodies against N. caninum and T. gondii associated with risk factors for infection.
Molecular biology
Of the 201 umbilical cord blood samples analyzed, two samples (1%) were Nc5 PCR-positive for N. caninum and these same samples were positive for ITS1 (GenBank: MN731361), but no sample of the placenta was positive.
These two Nc5 gene samples were sent for sequencing, and both shared 100% similarity with each other and 100% homology for N. caninum Liverpool (GenBank: LN714476). The highest homology (98–99%) was obtained for N. caninum with strains from other countries (Fig. 1). ITS1 gene samples shared 100% homology for N. caninum Liverpool (GenBank: U16159) after sequencing.
Figure 1
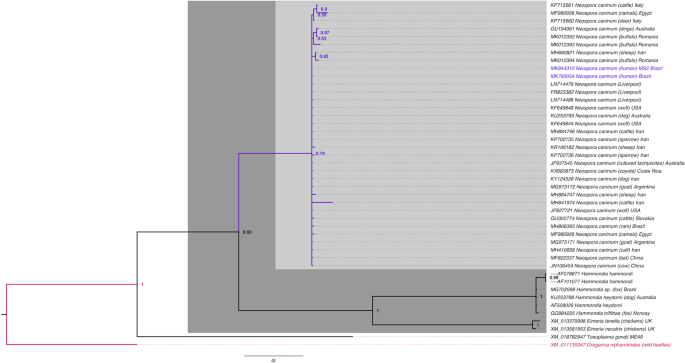
Phylogenetic tree of Neospora caninum (GenBank: MK790054; MK944312). Evolutionary history was inferred from the Bayesian Inference tree with the probability scores for the Nc5 gene. Bar, 0.2 changes per nucleotide position. The sample sequences obtained in this study are indicated in blue.
For the B1 gene of T. gondii, 16 samples presented bands compatible with the positive control in the PCR results, with 5.5% positivity in cord blood and 2.5% positivity in placental tissue. Detailed information on PCR positive samples are presented in the Supplementary Table S1.
Histopathological analysis
Histopathological analyses of 75 placenta samples (selected from among the samples showing PCR positivity and serological positive, prioritizing higher serological titles for both parasites) stained with hematoxylin-eosin were performed. These samples showed no structures characteristic of tissue cysts or inflammatory infiltrate.
Discussion
Changes in the maternal immune status occur during pregnancy to maintain fetal survival, and this immunosuppression may leave pregnant woman more prone to infections[17](/articles/s41598-020-65991-1#ref-CR17 "Yip, L., McCluskey, J. & Sinclair, R. Immunological aspects of pregnancy. Clin. Dermatol. 24, 84–7, https://doi.org/10.1016/j.clindermatol.2005.10.022
(2006)."),[18](/articles/s41598-020-65991-1#ref-CR18 "Weetman, A. P. The immunology of pregnancy. Thyroid. 9, 643–46,
https://doi.org/10.1089/thy.1999.9.643
(1999)."). Under healthy conditions, these infections are typically kept under control during pregnancy; however, the immature immune system of the fetus leaves it vulnerable to parasites that are able to cross the uteroplacental barrier[10](/articles/s41598-020-65991-1#ref-CR10 "Barratt, J. L. N., Harkness, J., Marriott, D., Ellis, J. T. & Stark, D. Importance of nonenteric protozoan infections in immunocompromised people. Clin. Microbiol. Rev. 23, 795–836, 10.1128%2FCMR.00001-10 (2010)."). The transplacental hematogenic route is the most common route of maternal-fetal parasite transmission[19](/articles/s41598-020-65991-1#ref-CR19 "Carlier, Y., Truyens, C., Deloron, P. & Peyron, F. Congenital parasitic infections: a review. Acta. Trop. 121, 55–70,
https://doi.org/10.1016/j.actatropica.2011.10.018
(2012)."). _T. gondii_ and other parasites in the phylum Apicomplexa actively penetrate their host cells _in vitro_, and this process is also possible _in vivo_[20](/articles/s41598-020-65991-1#ref-CR20 "Dobrowolski, J. M. & Sibley, L. D. Toxoplasma invasion of mammalian cells is powered by the actin cytoskeleton of the parasite. Cell. 84, 933–39,
https://doi.org/10.1016/s0092-8674(00)81071-5
(1996).").In the present study, 24.3% seropositivity for anti-N. caninum antibodies was found, suggesting human exposure to the parasite. The seropositivity rate was higher in the present study than the rate of 5% seropositivity found by Lobato et al.13, in 91 cord blood samples. Studies by Ibrahim et al.[14](/articles/s41598-020-65991-1#ref-CR14 "Ibrahim, H. M. et al. Prevalence of Neospora caninum and Toxoplasma gondii antibodies in northern Egypt. Am. J. Trop. Med. hyg. 80, 263–67, https://doi.org/10.4269/ajtmh.2009.80.263
(2009)."), found a 7.92% (8/101) seroprevalence among pregnant women for _N. caninum_, Tranas _et al_.[15](/articles/s41598-020-65991-1#ref-CR15 "Tranas, J., Heinzen, R. A., Weiss, L. M. & McAllister, M. M. Serological evidence of human infection with the protozoan Neospora caninum. Clin. Diagn. Lab. Immunol. 6, 765–67 (1999)."), found 6.7% (69/1,029) seropositivity in blood bank samples, and Oshiro _et al_.[16](/articles/s41598-020-65991-1#ref-CR16 "Oshiro, L. M. et al. Neospora caninum and Toxoplasma gondii serodiagnosis in human immunodeficiency virus carriers. Rev. Soc. Bras. Med. Trop. 48, 568–72,
https://doi.org/10.1590/0037-8682-0151-2015
(2015)."), found 26.1% (81/310) positivity in HIV-positive patients. The variations in the seropositivity rates found in several studies may be attributed to the study populations and the climatic and environmental factors of each region, as some authors have reported an association between climatic factors and risk factors for _N. caninum_ infection in cattle[2](/articles/s41598-020-65991-1#ref-CR2 "Dubey, J. P., Schares, G. & Ortega-Mora, L. M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 20, 323–67, 10.1128%2FCMR.00031-06 (2007)."),[21](/articles/s41598-020-65991-1#ref-CR21 "Rinaldi, L. et al. Neospora caninum in pastured cattle: determination of climatic, environmental, farm management and individual animal risk factors using remote sensing and geographical information systems. Vet. Parasitol. 128, 219–30,
https://doi.org/10.1016/j.vetpar.2004.12.011
(2005)."),[22](/articles/s41598-020-65991-1#ref-CR22 "Wouda, W., Bartels, C. J. M. & Moen, A. R. Characteristics of Neospora caninum-associated abortion storms in dairy herds in The Netherlands (1995 to1997). Theriogenology. 52, 233–45,
https://doi.org/10.1016/s0093-691x(99)00125-9
(1999)."). The sporulation and survival of coccidial oocysts in the environment may be favored by temperature and humidity[2](/articles/s41598-020-65991-1#ref-CR2 "Dubey, J. P., Schares, G. & Ortega-Mora, L. M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 20, 323–67, 10.1128%2FCMR.00031-06 (2007).").Of all the samples tested, only 8.4% presented concomitant seropositivity to T. gondii. In the literature, there have been reports of seropositivity for both parasites: Ibrahim et al.[14](/articles/s41598-020-65991-1#ref-CR14 "Ibrahim, H. M. et al. Prevalence of Neospora caninum and Toxoplasma gondii antibodies in northern Egypt. Am. J. Trop. Med. hyg. 80, 263–67, https://doi.org/10.4269/ajtmh.2009.80.263
(2009)."), reported 5.94% positivity, and Oshiro _et al_.[16](/articles/s41598-020-65991-1#ref-CR16 "Oshiro, L. M. et al. Neospora caninum and Toxoplasma gondii serodiagnosis in human immunodeficiency virus carriers. Rev. Soc. Bras. Med. Trop. 48, 568–72,
https://doi.org/10.1590/0037-8682-0151-2015
(2015)."), reported 25.2% positivity. However, the extent of _N. caninum_ and _T. gondii_ coinfection in humans is still unknown. To decrease the possibility of cross reaction, in the present study was used a serological cut off point of 1:50 and only fluorescent reactions along the periphery of the parasite were considered positive.In a study by Paré et al.[23](/articles/s41598-020-65991-1#ref-CR23 "Paré, J., Hietala, S. K. & Thurmond, M. C. Interpretation of an indirect fluorescent antibody test for diagnosis of Neospora sp. infection in cattle. J. Vet. Diagn. Investig. 7, 273–75, https://doi.org/10.1177/104063879500700310
(1995)."), complete peripheral fluorescence of the parasite was considered a positive response and apical fluorescence a negative response because the conservation of antigens in the apical organelles of a variety of Apicomplexa parasites may be responsible for cross-reactivity[24](/articles/s41598-020-65991-1#ref-CR24 "Taylor, D. W., Evans, C. B., Aley, S. B., Barta, J. R. & Danforth, H. D. Identification of an apically located antigen that is conserved in sporozoan parasites. J. Protozool. 37, 540–45,
https://doi.org/10.1111/j.1550-7408.1990.tb01262.x
(1990)."). Dilutions equal to or greater than 1:50 in the IFAT may be considered appropriate to avoid cross-reactivity between _N. caninum_ and _T. gondii_ in serum samples from some hosts[13](/articles/s41598-020-65991-1#ref-CR13 "Lobato, J. et al. Detection of immunoglobulin G antibodies to Neospora caninum in humans: high seropositivity rates in patients who are infected by human immunodeficiency virus or have neurological disorders. Clin. Vaccine. Immunol. 13, 84–9 (2006). 10.1128%2FCVI.13.1.84-89.2006."),[25](/articles/s41598-020-65991-1#ref-CR25 "Silva, D. A., Lobato, J., Mineo, T. W. & Mineo, J. R. Evaluation of serological tests for the diagnosis of Neospora caninum infection in dogs: optimization of cut off titers and inhibition studies of crossreactivity with Toxoplasma gondii. Vet. Parasitol. 143, 234–44,
https://doi.org/10.1016/j.vetpar.2006.08.028
(2007).").IFAT positive samples and molecular biology for N. caninum demonstrated western blot reactivity for rNcSRS2 (Nc-p43) surface antigen which is immunodominant, highly immunogenic, well conserved26 and does not cross react with T. gondii27.
The two positive PCR samples for N. caninum were not IFAT positive, but showed weak reactivity by western blot, demonstrating that western blot can be used as a complementary serological method in the diagnosis of neosporosis. In the literature there are reports of positive PCR samples that tested negative in serology tests in studies carried out with dogs[28](/articles/s41598-020-65991-1#ref-CR28 "Pena, H. F. D. J. et al. Isolation and molecular detection of Neospora caninum from naturally infected sheep from Brazil. Vet. Parasitol. 147, 61–66, https://doi.org/10.1016/j.vetpar.2007.03.002
(2007)."),[29](/articles/s41598-020-65991-1#ref-CR29 "Lindsay, D. S., Dubey, J. P. & Duncan, R. B. Confirmation that the dog is a definitive host for Neospora caninum. Vet. Parasitol. 82, 327–33,
https://doi.org/10.1016/S0304-4017(99)00054-0
(1999)."), and in a study carried out with bovines, in tests with aborted fetal tissues, the mother tested negative for _N. caninum_ by IFAT and ELISA and positive by PCR, with these samples showing poor reactivity on a western blot test[30](/articles/s41598-020-65991-1#ref-CR30 "Sager, H. et al. A Swiss case–control study to assess Neospora caninum-associated bovine abortions by PCR, histopathology and serology. Vet. Parasitol. 102, 1–15,
https://doi.org/10.1016/S0304-4017(01)00524-6
(2001).").Therefore, explanations for this fact can be attributed to the inability of some individuals to synthesize detectable antibodies against N. caninum due to acquired or innate immunotolerance[30](/articles/s41598-020-65991-1#ref-CR30 "Sager, H. et al. A Swiss case–control study to assess Neospora caninum-associated bovine abortions by PCR, histopathology and serology. Vet. Parasitol. 102, 1–15, https://doi.org/10.1016/S0304-4017(01)00524-6
(2001)."), or also to the previous chronic infection with antibodies not detectable in the 1:50 dilution[28](/articles/s41598-020-65991-1#ref-CR28 "Pena, H. F. D. J. et al. Isolation and molecular detection of Neospora caninum from naturally infected sheep from Brazil. Vet. Parasitol. 147, 61–66,
https://doi.org/10.1016/j.vetpar.2007.03.002
(2007)."). In studies carried out with mice, it has been demonstrated the appearance of IgM antibodies after 7 days of infection by _N. caninum_ and the production of IgG antibodies after 14 days of infection[31](/articles/s41598-020-65991-1#ref-CR31 "Teixeira, L. et al. Characterization of the B-cell immune response elicited in BALB/c mice challenged with Neospora caninum tachyzoites. Immunol. 116, 38–52,
https://doi.org/10.1111/j.1365-2567.2005.02195.x
(2005)."). This reinforces the possibility of the infection being acquired at the end of pregnancy with the mother still seronegative at delivery, as with _T. gondii_ infections[32](/articles/s41598-020-65991-1#ref-CR32 "Sensini, A. Toxoplasma gondii infection in pregnancy: opportunities and pitfalls of serological diagnosis. Clin. Microbiol. Infect. 12, 504–12,
https://doi.org/10.1111/j.1469-0691.2006.01444.x
(2006).").Samples were considered positive when Nc5 and ITS1 were positive. Of the 201 cord blood samples and 201 placental tissue samples analyzed, two cord blood samples showed PCR positivity for N. caninum using primers for the Nc5 and ITS1 region, and these samples were negative for T. gondii. After sequencing for Nc5 gene (GenBank: MK790054; MK944312), the samples demonstrated 98%-100% identity with several strains in the database, and for ITS1 gene (GenBank: MN731361) shared 100% homology for N. caninum Liverpool, suggesting that these sequences really represented samples of N. caninum. The phylogenetic tree showed a cluster of N. caninum among samples from around the world and different hosts.
Nc5 sequences were used to construct the phylogenetic tree, because unlike ITS1, the Nc5 gene is highly specific and excludes other species from the Toxoplasmatinae subfamily[33](/articles/s41598-020-65991-1#ref-CR33 "Yamage, M., Flechtner, O. & Gottstein, B. Neospora caninum: specific oligonucleotide primers for the detection of brain “cyst” DNA of experimentally infected nude mice by the polymerase chain reaction (PCR). J. Parasitol. 82, 272–79, https://doi.org/10.2307/3284160
(1996)."), which strengthens the molecular diagnosis of the present study.The positivity found for the Nc5 and ITS1 genes corroborate literature data. The use of nested-PCR methods directed to the Nc5 and ITS1 genes to detect N. caninum DNA may increase sensitivity and detection rate[34](#ref-CR34 "Gui, B. Z. et al. First report of Neospora caninum infection in pigs in China. Transbound. Emerg. Dis. 00, 1–4, https://doi.org/10.1111/tbed.13358
(2019)."),[35](#ref-CR35 "Wang, X. et al. Detection of Neospora caninum DNA by polymerase chain reaction in bats from Southern China. Parasitol. Int. 67, 389–91,
https://doi.org/10.1016/j.parint.2018.03.002
(2018)."),[36](/articles/s41598-020-65991-1#ref-CR36 "Salehi, N., Gottstein, B. & Haddadzadeh, H. R. Genetic diversity of bovine Neospora caninum determined by microsatellite markers. Parasitol. Int. 64, 357–61,
https://doi.org/10.1016/j.parint.2015.05.005
(2015).").The present study found positive molecular biology results for two umbilical cord blood samples but not for the corresponding placental samples. Because this is the first report of N. caninum in human samples, further studies are needed to clarify these findings. In studies with cows experimentally inoculated at different stages of pregnancy, some authors have reported that histopathological changes are less frequent at more advanced stages of pregnancy, suggesting that gestational age influences the outcome of placental infection[37](/articles/s41598-020-65991-1#ref-CR37 "Ogino, H. et al. Neosporosis in the aborted fetus and newborn calf. J. Comp. Pathol. 107, 231–37, https://doi.org/10.1016/0021-9975(92)90039-w
(1992)."),[38](/articles/s41598-020-65991-1#ref-CR38 "Macaldowie, C. et al. Placental pathology associated with fetal death in cattle inoculated with Neospora caninum by two different routes in early pregnancy. J. Comp. Pathol. 131, 142–56,
https://doi.org/10.1016/j.jcpa.2004.02.005
(2004).").In an experimental study conducted by Ho et al.39, with pregnant monkeys (Macaca mulatta), the sporadic and inconsistent distribution of N. caninum in tissues other than those from the central nervous system was proposed to be a manifestation of constant dissemination of a small number of parasites into the bloodstream.
Human neosporosis is still an uncertain issue, despite serological evidence of human exposure, primarily in immunocompromised populations10,13,15. Considering the high efficiency and prevalence of vertical transmission of N. caninum in cattle[40](/articles/s41598-020-65991-1#ref-CR40 "Davison, H. C., Otter, A. & Trees, A. J. Estimation of vertical and horizontal transmission parameters of Neospora caninum infections in dairy cattle. Int. J. Parasitol. 29, 1683–89, https://doi.org/10.1016/s0020-7519(99)00129-0
(1999).") and its close relationship with _T. gondii_, the possibility of Neospora posing a risk to human pregnancy should not be ruled out. Experimental studies with nonhuman primates indicated susceptibility to transplacental infection, and fetal lesions caused by _N. caninum_ infection were similar to those induced by _T. gondii_ infection[41](/articles/s41598-020-65991-1#ref-CR41 "Barr, B. C., Conrad, P. A., Sverlow, K. W., Tarantal, A. F. & Hendrickx, A. G. Experimental fetal and transplacental Neospora infection in the nonhuman primate. Lab. Invest. 71, 236–42 (1994)."). An _in vitro_ study has shown that human trophoblasts and cervical cells are readily infected by _N. caninum_, although they show differences in susceptibility to infection, cytokine production and cell viability[42](/articles/s41598-020-65991-1#ref-CR42 "Carvalho, J. V. et al. Differential susceptibility of human trophoblastic (BeWo) and uterine cervical (HeLa) cells to Neospora caninum infection. Int. J. Parasitol. 40, 1629–37,
https://doi.org/10.1016/j.ijpara.2010.06.010
(2010).").In this study, a significant association between seropositivity for N. caninum and the presence of animals as well as the presence of dogs was observed. Canids are known to be the definitive, exclusive hosts of N. caninum2. Some authors have reported that the presence of dogs on rural properties may be related to an increased likelihood of infection in cattle, thus highlighting the role of dogs in the epidemiological chain of neosporosis in farm animals[43](#ref-CR43 "Schares, G. et al. Potential risk factors for bovine Neospora caninum infection in Germany are not under the control of the farmers. Parasitol. 129, 301–9, https://doi.org/10.1017/s0031182004005700
(2004)."),[44](#ref-CR44 "Guimarães, J. S. Jr., Souza, S. L. P., Bergamaschi, D. P. & Gennari, S. M. Prevalence of Neospora caninum antibodies and factors associated with their presence in dairy cattle of the north of Paraná state, Brazil. Vet. Parasitol. 124, 1–8,
https://doi.org/10.1016/j.vetpar.2004.07.002
(2004)."),[45](/articles/s41598-020-65991-1#ref-CR45 "Wouda, W., Dijkstra, T., Kramer, A. M., van Maanen, C. & Brinkhof, J. M. Seroepidemiological evidence for a relationship between Neospora caninum infections in dogs and cattle. Int. J. Parasitol. 29, 1677–82,
https://doi.org/10.1016/S0020-7519(99)00105-8
(1999)."). Since dogs are definitive hosts and excrete oocysts in feces, the potential for human exposure to _N. caninum_ is high[14](/articles/s41598-020-65991-1#ref-CR14 "Ibrahim, H. M. et al. Prevalence of Neospora caninum and Toxoplasma gondii antibodies in northern Egypt. Am. J. Trop. Med. hyg. 80, 263–67,
https://doi.org/10.4269/ajtmh.2009.80.263
(2009)."). The presence of dogs may be related to _N. caninum_ seropositivity in the analyzed pregnant women, but additional studies in this area are necessary to establish this correlation.In the present study, 26.8% seropositivity for anti-T. gondii IgG antibodies was observed. Being 73.1% were seronegative for the presence of IgG antibodies, and 100% were negative for IgM. According to Villard et al.[46](/articles/s41598-020-65991-1#ref-CR46 "Villard, O. et al. Serological diagnosis of Toxoplasma gondii infection: recommendations from the French National Reference Center for Toxoplasmosis. Diagn. Microbiol. Infect. Dis. 84, 22–33, https://doi.org/10.1016/j.diagmicrobio.2015.09.009
(2016)."), the presence of specific IgG and the absence of IgM antibodies are indications of previous infection, however, the infection can be acquired at the end of gestation, with the mother still being seronegative at birth[32](/articles/s41598-020-65991-1#ref-CR32 "Sensini, A. Toxoplasma gondii infection in pregnancy: opportunities and pitfalls of serological diagnosis. Clin. Microbiol. Infect. 12, 504–12,
https://doi.org/10.1111/j.1469-0691.2006.01444.x
(2006)."). The prevalence of IgG antibodies among pregnant women in Brazil is variable, and it can reach 63.03%[47](#ref-CR47 "da Silva, M. G., Vinaud, M. C. & de Castro, A. M. Prevalence of toxoplasmosis in pregnant women and vertical transmission of Toxoplasma gondii in patients from basic units of health from Gurupi, Tocantins, Brazil, from 2012 to 2014. PLoS One 10, e0141700,
https://doi.org/10.1371/journal.pone.0141700
(2015)."),[48](#ref-CR48 "Lopes-Mori, F. M. et al. Gestational toxoplasmosis in Paraná State, Brazil: prevalence of IgG antibodies and associated risk factors. Braz. J. Infect. Dis. 17, 405–9,
https://doi.org/10.1016/j.bjid.2012.12.003
(2013)."),[49](/articles/s41598-020-65991-1#ref-CR49 "Reis, M. M., Tessaro, M. M. & D’Azevedo, P. A. Serologic profile of toxoplasmosis in pregnant women from a public hospital in Porto Alegre. Rev. Bras. Ginecol. Obstet. 28, 158–64,
https://doi.org/10.1590/S0100-72032006000300004
(2006).").A significant association was found between T. gondii seropositivity and basic sanitation (access to sewage and treated water) with p = 0.04, but no significant associations were found with other risk factors. According to Silva et al.[50](/articles/s41598-020-65991-1#ref-CR50 "da Silva, M. G., Câmara, J. T., Vinaud, M. C. & de Castro, A. M. Epidemiological factors associated with seropositivity for toxoplasmosis in pregnant women from Gurupi, State of Tocantins, Brazil. Rev. Soc. Bras. Med. Trop. 47, 469–75, https://doi.org/10.1590/0037-8682-0127-2014
(2014)."), a lack of basic sanitation is associated with risk factors for _T. gondii_ infection, with low socioeconomic level, low educational level, older age, soil management and contact with cats being considered more important risk factors in pregnant women in Brazil[6](/articles/s41598-020-65991-1#ref-CR6 "Dubey, J. P., Lago, E. G., Gennari, S. M., Su, C. & Jones, J. L. Toxoplasmosis in humans and animals in Brazil: high prevalence, high burden of disease, and epidemiology. Parasitol. 139, 1375–1424,
https://doi.org/10.1017/S0031182012000765
(2012)."). In the analyzed samples, there was no significant correlation between older age and serological positivity, perhaps because the pregnant women composing the study group were young.The use of PCR analysis in the determination of intrauterine T. gondii infection allows early diagnosis and avoids the use of invasive procedures for the fetus[51](/articles/s41598-020-65991-1#ref-CR51 "Montoya, J. G. Laboratory diagnosis of Toxoplasma gondii infection and toxoplasmosis. J. Infect. Dis. 185, S73–S82, https://doi.org/10.1086/338827
(2002)."). In this study, we observed 5.5% positivity in cord blood and 2.5% positivity in placental tissue for the B1 gene, even with the exclusion of acute infection confirmed by serology. Postnatal screening may be associated with the detection of these parasites in amniotic fluid, the placenta and cord or neonate serum and may be a management strategy complementary to prenatal diagnosis[46](/articles/s41598-020-65991-1#ref-CR46 "Villard, O. et al. Serological diagnosis of Toxoplasma gondii infection: recommendations from the French National Reference Center for Toxoplasmosis. Diagn. Microbiol. Infect. Dis. 84, 22–33,
https://doi.org/10.1016/j.diagmicrobio.2015.09.009
(2016).").The B1 gene has approximately 35 copies and is highly conserved in all strains[52](/articles/s41598-020-65991-1#ref-CR52 "Cazenave, J., Cheyrou, A., Blouin, P., Johnson, A. M. & Begueret, J. Use of polymerase chain reaction to detect Toxoplasma. J. Clin. Pathol. 44, 1037, https://doi.org/10.1136/jcp.44.12.1037-a
(1991)."). According to Jones _et al_.[53](/articles/s41598-020-65991-1#ref-CR53 "Jones, C. D., Okhravi, N., Adamson, P., Tasker, S. & Lightman, S. Comparison of PCR detection methods for B1, P30, and 18S rDNA genes of T. gondii in aqueous humor. Invest. Ophthalmol. Vis. Sci. 41, 634–44 (2000)."), primers for the B1 gene have higher specificity because they do not amplify DNA from a variety of bacterial and fungal species and because, even in the presence of increasing amounts of human DNA, the sensitivity of the reaction remains unchanged; it is able to detect 50 femtograms (corresponding to a single organism) of _T. gondii_ DNA.In conclusion, the seroprevalence of N. caninum can be indicative of parasite exposure, and the presence of dogs may be associated with seropositivity. Additional studies are needed to clarify possible risk factors related to N. caninum. The PCR DNA detection results indicate that the role of N. caninum in human pregnancy still needs to be elucidated in order to determine the extent and importance of human exposure, given that the parasite has thus far not been isolated from human tissues. These findings may contribute to implementation of diagnostic tests in routine prenatal screening. The seroprevalence for T. gondii in pregnant women found in the present study was low compared with that found in other regions of Brazil, and lack of basic sanitation represented an important risk factor. However, seronegativity may indicate susceptibility to infection.
Methods
Ethics statement
The study was approved by the Ethics Committee for Research Involving Human Beings of the Federal University of Mato Grosso do Sul (UFMS) on 03 November 2016, document number 1.804.047. All included patients accepted the conditions of the study and signed the free informed consent form. All methods were carried out in accordance with relevant guidelines and regulations.
Sample collection
This study is an analytical cross-sectional study. Between January and May 2017, a total of 201 cord blood and placental tissue samples were collected from pregnant women admitted to the delivery room and surgical center of Cândido Mariano Maternity Hospital, located in Campo Grande, Mato Grosso do Sul, Brazil.
Immediately after delivery, umbilical cord blood was collected in a vacutainer tube containing K2 EDTA for molecular analysis and a clot activator tube for serological analysis. Placental fragments weighing 1–2 grams were collected from the fetal (or chorionic) and maternal ends of the placental hilus for molecular and histological analyses[54](/articles/s41598-020-65991-1#ref-CR54 "Duarte, P. O. et al. Neonatal sepsis: evaluation of risk factors and histopathological examination of placentas after delivery. Biosci. J. 35, 629–39, https://doi.org/10.14393/BJ-v35n2a20198-41814
(2019).").Data were collected from the patients’ charts and from a form completed by the patients that evaluated the following variables: age, gestational age, number of prenatal consultations, problems in previous pregnancies, and living conditions and habits (consumption of raw or undercooked meat; work or leisure activities involving soil; domestic animal raising; presence of cats and/or dogs in the home; and presence of basic sanitation/access to sewage collection or treated water).
The patients included in the study were healthy pregnant women with a normal pregnancy who were in initial labor and admitted to the same maternity sector.
Serology
IFAT
The indirect fluorescent antibody test (IFAT) for the detection of anti-N. caninum antibodies was performed using an Imunoteste Neospora (RIFI) commercial diagnostic Kit (Imunodot diagnósticos, Jaboticabal-SP, Brazil) following the manufacturer’s recommendations with adaptations. Previously established positive and negative human serum controls provided by Oshiro et al., were used[16](/articles/s41598-020-65991-1#ref-CR16 "Oshiro, L. M. et al. Neospora caninum and Toxoplasma gondii serodiagnosis in human immunodeficiency virus carriers. Rev. Soc. Bras. Med. Trop. 48, 568–72, https://doi.org/10.1590/0037-8682-0151-2015
(2015)."). Samples were considered positive at a dilution of 1:50.The IFAT for the detection of anti-T. gondii antibodies was performed using an Imuno-Con Toxoplasmose kit (WAMA Diagnóstica, São Carlos-SP, Brazil) following the manufacturer’s recommendations. Samples were considered positive at a dilution of 1:64.
For both serological tests of the 201 samples, human anti-IgG and anti-IgM fluorescence conjugate at 1:100 dilution (conjugated with fluorescein isothiocyanate; Sigma-Aldrich, St. Louis, Missouri, USA) were used. The slides were observed using a fluorescence-equipped microscope (Axioskop- Carl Zeiss, Germany) (epi-lighting system) with a 40× objective.
Fluorescent reactions along the periphery of the parasite were considered positive. In the negative reactions, the parasites on the slide did not show fluorescence, or the fluorescence was located at only one end, characterized as polar coloration or an apical reaction. Samples with peripheral fluorescence of total tachyzoites were considered positive[23](/articles/s41598-020-65991-1#ref-CR23 "Paré, J., Hietala, S. K. & Thurmond, M. C. Interpretation of an indirect fluorescent antibody test for diagnosis of Neospora sp. infection in cattle. J. Vet. Diagn. Investig. 7, 273–75, https://doi.org/10.1177/104063879500700310
(1995).").Western blot
N. caninum rNcSRS2 partial recombinant sequence (Nc-p43)55 protein was separated on 12% SDS-PAGE gel and transferred to PVDV membrane (GE Healthcare, UK) at 25 mA overnight (Supplementary Protocols S1).
Molecular biology
DNA isolation
Approximately 300 microliters (µl) of cord blood from each sample (201 total) and 50 milligrams of placental tissue from each sample (201 total) were subjected to DNA extraction using a protocol adapted from Regitano and Coutinho56 (Supplementary Protocols S1).
Samples were quantified via spectrophotometry (NanoDrop ND-1000, Uniscience) and diluted to 100 nanograms for PCR. The viability of the samples and DNA quality were evaluated using primers for the human β-globin gene as described by Bauer et al.[57](/articles/s41598-020-65991-1#ref-CR57 "Bauer, H. M. et al. Genital human papillomavirus infection in female university students as determined by a PCR-based method. JAMA. 265, 472–77, https://doi.org/10.1001/jama.1991.03460040048027
(1991).").PCR for neospora caninum and toxoplasma gondii
For detection of N. caninum, the primers NP21 and NP4 were used for the primary amplification and primers NP7 and NP4 were used in the secondary reactions to target the Nc5 gene as described by Yamage et al.[33](/articles/s41598-020-65991-1#ref-CR33 "Yamage, M., Flechtner, O. & Gottstein, B. Neospora caninum: specific oligonucleotide primers for the detection of brain “cyst” DNA of experimentally infected nude mice by the polymerase chain reaction (PCR). J. Parasitol. 82, 272–79, https://doi.org/10.2307/3284160
(1996)."). Primers for internal transcribed spacer (ITS1) region was used out with four oligonucleotides as described by Buxton _et al_.[58](#ref-CR58 "Buxton, D. et al. The pathogenesis of experimental neosporosis in pregnant sheep. J. Comp. Pathol. 118, 267–79,
https://doi.org/10.1016/s0021-9975(07)80003-x
(1998)."),[59](#ref-CR59 "Bartley, P. M. et al. Detection of Neospora caninum DNA in cases of bovine and ovine abortion in the South-West of Scotland. Parasitol. 146, 979–82,
https://doi.org/10.1017/S0031182019000301
(2019)."),[60](/articles/s41598-020-65991-1#ref-CR60 "Hecker, Y. P. et al. Ovine abortion by Neospora caninum: first case reported in Argentina. Acta Parasitol. 1–6,
https://doi.org/10.2478/s11686-019-00106-z
(2019).") (Supplementary Protocols [S1](/articles/s41598-020-65991-1#MOESM1)).For detection of T. gondii, was used primer to perform simple PCR targeting the repetitive and conserved B1 gene61, a nested PCR was also performed using N2-C2 primers, which amplified a 97-bp product of the B1 gene[62](/articles/s41598-020-65991-1#ref-CR62 "Spalding, S. M., Amendoeira, M. R. R., Coelho, J. M. C. & Angel, S. O. Otimização da reação de polimerase em cadeia para detecção de Toxoplasma gondii em sangue venoso e placenta de gestantes. J. Bras. Med. Patol. Lab. 38, 105–10, https://doi.org/10.1590/S1676-24442002000200006
(2002).") (Supplementary Protocols [S1](/articles/s41598-020-65991-1#MOESM1)).Negative (ultrapure water) and positive (N. caninum NC‐1 strain and T. gondii RH strain) controls were included with all PCR reactions. To increase the sensitivity of the assay, each DNA sample was tested in triplicate.
The final product was visualized on a 1.5% agarose gel stained with ethidium bromide (EtBr).
Samples yielding an expected PCR product size for N. caninum were purified using a PureLink quick gel extraction kit (Invitrogen, Carlsbad, CA) and DNA-sequenced at René Rachou Research Center (Oswaldo Cruz Foundation-FIOCRUZ) in an automatic sequencer (ABI Prism 3730XL Genetic Analyzer, Applied Biosystems, EUA) with a 48-capillary DNA analysis system.
Phylogenetic tree construction
Using the BLASTn program, sequences available from GenBank was aligned with the sequence of the Nc5 gene (GenBank: MK790054; MK944312). The Mega 6.0 program[63](/articles/s41598-020-65991-1#ref-CR63 "Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 30, 2725–29, https://doi.org/10.1093/molbev/mst197
(2013).") was used to align the sequences taken from GenBank and construct a database that contained all similar sequences obtained from the analysis. Using the MrBayes 3.2.6 program was performed a Bayesian phylogenetic analysis for the Nc5 gene and the results were plotted using the FigTree 1.4.2 program[64](#ref-CR64 "Ronquist, F. & Huelsenbeck, J. P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 19, 1572–74,
https://doi.org/10.1093/bioinformatics/btg180
(2003)."),[65](#ref-CR65 "Tree Bio. FigTree [online]. London: Tree Bio; 2016 [cited 2019 November 10]. Available from,
http://tree.bio.ed.ac.uk/software/figtree/
."),[66](/articles/s41598-020-65991-1#ref-CR66 "Csordas, B. G. et al. New insights from molecular characterization of the tick Rhipicephalus (Boophilus) microplus in Brazil. Rev. Bras. Parasitol. Vet. 25, 317–26,
https://doi.org/10.1590/S1984-29612016053
(2016).").The topology of the tree was used to generate a 50% majority rule consensus, with the percentage of samples recovering any particular clade representing the posterior probability of a clade (1 = 100%). No manual editing of the tree was performed. The Gregarina niphandrodes (GenBank: XM_011135347) dataset was used as the outgroup in the phylogenetic tree.
Histopathological analysis
Fragments of placental samples weighing 1 to 2 grams collected during delivery were immediately fixed in 10% buffered formalin for 24 hours, processed (xylol alcohol), embedded in paraffin, sliced into a final thickness of 5 µm and placed on a slide for hematoxylin-eosin staining. The slides were visualized at 400X magnification to examine placental morphology.
Statistical analysis
The collected data were tabulated and analyzed using the statistical software IBM SPSS Statistics version 20 (Inc., Chicago, Illinois, USA). The χ2 test, Fisher’s exact test and odds ratios were used to assess associations between the variables (consumption of raw/undercooked meat, work or leisure activities involving soil, domestic animals, cat, dog, basic sanitation) and the serology results. p values less than 0.05 were considered statistically significant.
References
- Bjerkfis, I., Mohn, S. F. & Presthus, J. Unidentified cyst-forming sporozoon causing encephalomyelitis and myositis in dogs. Z. Parasitenkd. 70, 271–74 (1984).
Article Google Scholar - Dubey, J. P., Schares, G. & Ortega-Mora, L. M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 20, 323–67, 10.1128%2FCMR.00031-06 (2007).
- Dubey, J. P. & Lindsay, D. S. A review of Neospora caninum and neosporosis. Vet. Parasitol. 67, 1–59, https://doi.org/10.1016/s0304-4017(96)01035-7 (1996).
Article CAS PubMed Google Scholar - Tenter, A. M., Heckeroth, A. R. & Weiss, L. M. Toxoplasma gondii: from animals to humans. Int. J. Parasitol. 30, 1217–58, https://doi.org/10.1016/S0020-7519(00)00124-7 (2000).
Article CAS PubMed PubMed Central Google Scholar - Dubey, J. P. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 28, 1019–24, https://doi.org/10.1016/S0020-7519(98)00023-X (1998).
Article CAS PubMed Google Scholar - Dubey, J. P., Lago, E. G., Gennari, S. M., Su, C. & Jones, J. L. Toxoplasmosis in humans and animals in Brazil: high prevalence, high burden of disease, and epidemiology. Parasitol. 139, 1375–1424, https://doi.org/10.1017/S0031182012000765 (2012).
Article CAS Google Scholar - Lindsay, D. S. & Dubey, J. P. Toxoplasma gondii: the changing paradigm of congenital toxoplasmosis. Parasitol. 138, 1829–31, https://doi.org/10.1017/S0031182011001478 (2011).
Article CAS Google Scholar - Hill, D. & Dubey, J. P. Toxoplasma gondii: transmission, diagnosis and prevention. Clin. Microbiol. Infect. 8, 634–40, https://doi.org/10.1046/j.1469-0691.2002.00485.x (2002).
Article CAS PubMed Google Scholar - Hemphill, A., Fuchs, N., Sonda, S. & Hehl, A. The antigenic composition of Neospora caninum. Int. J. Parasitol. 29, 1175–88, https://doi.org/10.1016/S0020-7519(99)00085-5 (1999).
Article CAS PubMed Google Scholar - Barratt, J. L. N., Harkness, J., Marriott, D., Ellis, J. T. & Stark, D. Importance of nonenteric protozoan infections in immunocompromised people. Clin. Microbiol. Rev. 23, 795–836, 10.1128%2FCMR.00001-10 (2010).
- Buxton, D., McAllister, M. M. & Dubey, J. P. The comparative pathogenesis of neosporosis. Trends Parasitol. 18, 546–52, https://doi.org/10.1016/S1471-4922(02)02414-5 (2002).
Article CAS PubMed Google Scholar - Al-Qassab, S. E., Reichel, M. P. & Ellis, J. T. On the biological and genetic diversity in Neospora caninum. Diversity. 2, 411–38, https://doi.org/10.3390/d2030411 (2010).
Article CAS Google Scholar - Lobato, J. et al. Detection of immunoglobulin G antibodies to Neospora caninum in humans: high seropositivity rates in patients who are infected by human immunodeficiency virus or have neurological disorders. Clin. Vaccine. Immunol. 13, 84–9 (2006). 10.1128%2FCVI.13.1.84-89.2006.
Article CAS Google Scholar - Ibrahim, H. M. et al. Prevalence of Neospora caninum and Toxoplasma gondii antibodies in northern Egypt. Am. J. Trop. Med. hyg. 80, 263–67, https://doi.org/10.4269/ajtmh.2009.80.263 (2009).
Article PubMed Google Scholar - Tranas, J., Heinzen, R. A., Weiss, L. M. & McAllister, M. M. Serological evidence of human infection with the protozoan Neospora caninum. Clin. Diagn. Lab. Immunol. 6, 765–67 (1999).
Article CAS Google Scholar - Oshiro, L. M. et al. Neospora caninum and Toxoplasma gondii serodiagnosis in human immunodeficiency virus carriers. Rev. Soc. Bras. Med. Trop. 48, 568–72, https://doi.org/10.1590/0037-8682-0151-2015 (2015).
Article PubMed Google Scholar - Yip, L., McCluskey, J. & Sinclair, R. Immunological aspects of pregnancy. Clin. Dermatol. 24, 84–7, https://doi.org/10.1016/j.clindermatol.2005.10.022 (2006).
Article PubMed Google Scholar - Weetman, A. P. The immunology of pregnancy. Thyroid. 9, 643–46, https://doi.org/10.1089/thy.1999.9.643 (1999).
Article CAS PubMed Google Scholar - Carlier, Y., Truyens, C., Deloron, P. & Peyron, F. Congenital parasitic infections: a review. Acta. Trop. 121, 55–70, https://doi.org/10.1016/j.actatropica.2011.10.018 (2012).
Article PubMed Google Scholar - Dobrowolski, J. M. & Sibley, L. D. Toxoplasma invasion of mammalian cells is powered by the actin cytoskeleton of the parasite. Cell. 84, 933–39, https://doi.org/10.1016/s0092-8674(00)81071-5 (1996).
Article CAS PubMed Google Scholar - Rinaldi, L. et al. Neospora caninum in pastured cattle: determination of climatic, environmental, farm management and individual animal risk factors using remote sensing and geographical information systems. Vet. Parasitol. 128, 219–30, https://doi.org/10.1016/j.vetpar.2004.12.011 (2005).
Article CAS PubMed Google Scholar - Wouda, W., Bartels, C. J. M. & Moen, A. R. Characteristics of Neospora caninum-associated abortion storms in dairy herds in The Netherlands (1995 to1997). Theriogenology. 52, 233–45, https://doi.org/10.1016/s0093-691x(99)00125-9 (1999).
Article CAS PubMed Google Scholar - Paré, J., Hietala, S. K. & Thurmond, M. C. Interpretation of an indirect fluorescent antibody test for diagnosis of Neospora sp. infection in cattle. J. Vet. Diagn. Investig. 7, 273–75, https://doi.org/10.1177/104063879500700310 (1995).
Article Google Scholar - Taylor, D. W., Evans, C. B., Aley, S. B., Barta, J. R. & Danforth, H. D. Identification of an apically located antigen that is conserved in sporozoan parasites. J. Protozool. 37, 540–45, https://doi.org/10.1111/j.1550-7408.1990.tb01262.x (1990).
Article CAS PubMed Google Scholar - Silva, D. A., Lobato, J., Mineo, T. W. & Mineo, J. R. Evaluation of serological tests for the diagnosis of Neospora caninum infection in dogs: optimization of cut off titers and inhibition studies of crossreactivity with Toxoplasma gondii. Vet. Parasitol. 143, 234–44, https://doi.org/10.1016/j.vetpar.2006.08.028 (2007).
Article PubMed Google Scholar - Howe, D. K., Crawford, A. C., Lindsay, D. & Sibley, L. D. The p29 and p35 immunodominant antigens of Neospora caninum tachyzoites are homologous to the family of surface antigens of Toxoplasma gondii. Infect. Immun. 66, 5322–28 (1998).
Article CAS Google Scholar - Hemphill, A. Subcellular localization and functional characterization of Nc-p43, a major Neospora caninum tachyzoite surface protein. Infect. Immun. 64, 4279–87 (1996).
Article CAS Google Scholar - Pena, H. F. D. J. et al. Isolation and molecular detection of Neospora caninum from naturally infected sheep from Brazil. Vet. Parasitol. 147, 61–66, https://doi.org/10.1016/j.vetpar.2007.03.002 (2007).
Article CAS PubMed Google Scholar - Lindsay, D. S., Dubey, J. P. & Duncan, R. B. Confirmation that the dog is a definitive host for Neospora caninum. Vet. Parasitol. 82, 327–33, https://doi.org/10.1016/S0304-4017(99)00054-0 (1999).
Article CAS PubMed Google Scholar - Sager, H. et al. A Swiss case–control study to assess Neospora caninum-associated bovine abortions by PCR, histopathology and serology. Vet. Parasitol. 102, 1–15, https://doi.org/10.1016/S0304-4017(01)00524-6 (2001).
Article CAS PubMed Google Scholar - Teixeira, L. et al. Characterization of the B-cell immune response elicited in BALB/c mice challenged with Neospora caninum tachyzoites. Immunol. 116, 38–52, https://doi.org/10.1111/j.1365-2567.2005.02195.x (2005).
Article CAS Google Scholar - Sensini, A. Toxoplasma gondii infection in pregnancy: opportunities and pitfalls of serological diagnosis. Clin. Microbiol. Infect. 12, 504–12, https://doi.org/10.1111/j.1469-0691.2006.01444.x (2006).
Article CAS PubMed Google Scholar - Yamage, M., Flechtner, O. & Gottstein, B. Neospora caninum: specific oligonucleotide primers for the detection of brain “cyst” DNA of experimentally infected nude mice by the polymerase chain reaction (PCR). J. Parasitol. 82, 272–79, https://doi.org/10.2307/3284160 (1996).
Article CAS PubMed Google Scholar - Gui, B. Z. et al. First report of Neospora caninum infection in pigs in China. Transbound. Emerg. Dis. 00, 1–4, https://doi.org/10.1111/tbed.13358 (2019).
Article CAS Google Scholar - Wang, X. et al. Detection of Neospora caninum DNA by polymerase chain reaction in bats from Southern China. Parasitol. Int. 67, 389–91, https://doi.org/10.1016/j.parint.2018.03.002 (2018).
Article CAS PubMed PubMed Central Google Scholar - Salehi, N., Gottstein, B. & Haddadzadeh, H. R. Genetic diversity of bovine Neospora caninum determined by microsatellite markers. Parasitol. Int. 64, 357–61, https://doi.org/10.1016/j.parint.2015.05.005 (2015).
Article CAS PubMed Google Scholar - Ogino, H. et al. Neosporosis in the aborted fetus and newborn calf. J. Comp. Pathol. 107, 231–37, https://doi.org/10.1016/0021-9975(92)90039-w (1992).
Article CAS PubMed Google Scholar - Macaldowie, C. et al. Placental pathology associated with fetal death in cattle inoculated with Neospora caninum by two different routes in early pregnancy. J. Comp. Pathol. 131, 142–56, https://doi.org/10.1016/j.jcpa.2004.02.005 (2004).
Article CAS PubMed Google Scholar - Ho, M. S. et al. Detection of Neospora from tissues of experimentally infected rhesus macaques by PCR and specific DNA probe hybridization. J. Clin. Microbiol. 35, 1740–45 (1997).
Article CAS Google Scholar - Davison, H. C., Otter, A. & Trees, A. J. Estimation of vertical and horizontal transmission parameters of Neospora caninum infections in dairy cattle. Int. J. Parasitol. 29, 1683–89, https://doi.org/10.1016/s0020-7519(99)00129-0 (1999).
Article CAS PubMed Google Scholar - Barr, B. C., Conrad, P. A., Sverlow, K. W., Tarantal, A. F. & Hendrickx, A. G. Experimental fetal and transplacental Neospora infection in the nonhuman primate. Lab. Invest. 71, 236–42 (1994).
CAS PubMed Google Scholar - Carvalho, J. V. et al. Differential susceptibility of human trophoblastic (BeWo) and uterine cervical (HeLa) cells to Neospora caninum infection. Int. J. Parasitol. 40, 1629–37, https://doi.org/10.1016/j.ijpara.2010.06.010 (2010).
Article CAS PubMed Google Scholar - Schares, G. et al. Potential risk factors for bovine Neospora caninum infection in Germany are not under the control of the farmers. Parasitol. 129, 301–9, https://doi.org/10.1017/s0031182004005700 (2004).
Article CAS Google Scholar - Guimarães, J. S. Jr., Souza, S. L. P., Bergamaschi, D. P. & Gennari, S. M. Prevalence of Neospora caninum antibodies and factors associated with their presence in dairy cattle of the north of Paraná state, Brazil. Vet. Parasitol. 124, 1–8, https://doi.org/10.1016/j.vetpar.2004.07.002 (2004).
Article PubMed Google Scholar - Wouda, W., Dijkstra, T., Kramer, A. M., van Maanen, C. & Brinkhof, J. M. Seroepidemiological evidence for a relationship between Neospora caninum infections in dogs and cattle. Int. J. Parasitol. 29, 1677–82, https://doi.org/10.1016/S0020-7519(99)00105-8 (1999).
Article CAS PubMed Google Scholar - Villard, O. et al. Serological diagnosis of Toxoplasma gondii infection: recommendations from the French National Reference Center for Toxoplasmosis. Diagn. Microbiol. Infect. Dis. 84, 22–33, https://doi.org/10.1016/j.diagmicrobio.2015.09.009 (2016).
Article CAS PubMed Google Scholar - da Silva, M. G., Vinaud, M. C. & de Castro, A. M. Prevalence of toxoplasmosis in pregnant women and vertical transmission of Toxoplasma gondii in patients from basic units of health from Gurupi, Tocantins, Brazil, from 2012 to 2014. PLoS One 10, e0141700, https://doi.org/10.1371/journal.pone.0141700 (2015).
Article CAS Google Scholar - Lopes-Mori, F. M. et al. Gestational toxoplasmosis in Paraná State, Brazil: prevalence of IgG antibodies and associated risk factors. Braz. J. Infect. Dis. 17, 405–9, https://doi.org/10.1016/j.bjid.2012.12.003 (2013).
Article ADS PubMed Google Scholar - Reis, M. M., Tessaro, M. M. & D’Azevedo, P. A. Serologic profile of toxoplasmosis in pregnant women from a public hospital in Porto Alegre. Rev. Bras. Ginecol. Obstet. 28, 158–64, https://doi.org/10.1590/S0100-72032006000300004 (2006).
Article Google Scholar - da Silva, M. G., Câmara, J. T., Vinaud, M. C. & de Castro, A. M. Epidemiological factors associated with seropositivity for toxoplasmosis in pregnant women from Gurupi, State of Tocantins, Brazil. Rev. Soc. Bras. Med. Trop. 47, 469–75, https://doi.org/10.1590/0037-8682-0127-2014 (2014).
Article PubMed Google Scholar - Montoya, J. G. Laboratory diagnosis of Toxoplasma gondii infection and toxoplasmosis. J. Infect. Dis. 185, S73–S82, https://doi.org/10.1086/338827 (2002).
Article PubMed Google Scholar - Cazenave, J., Cheyrou, A., Blouin, P., Johnson, A. M. & Begueret, J. Use of polymerase chain reaction to detect Toxoplasma. J. Clin. Pathol. 44, 1037, https://doi.org/10.1136/jcp.44.12.1037-a (1991).
Article CAS PubMed PubMed Central Google Scholar - Jones, C. D., Okhravi, N., Adamson, P., Tasker, S. & Lightman, S. Comparison of PCR detection methods for B1, P30, and 18S rDNA genes of T. gondii in aqueous humor. Invest. Ophthalmol. Vis. Sci. 41, 634–44 (2000).
CAS PubMed Google Scholar - Duarte, P. O. et al. Neonatal sepsis: evaluation of risk factors and histopathological examination of placentas after delivery. Biosci. J. 35, 629–39, https://doi.org/10.14393/BJ-v35n2a20198-41814 (2019).
Article Google Scholar - Lima Junior, M. S. D. C., Andreotti, R., Caetano, A. R., Paiva, F. & Matos, M. D. F. C. Cloning and expression of an antigenic domain of a major surface protein (Nc-p43) of Neospora caninum. Rev. Bras. Parasitol. Vet. 16, 61–66 (2007).
Google Scholar - Regitano, L. C. A., Coutinho, L. L. Biologia molecular aplicada à produção animal, Brasília: Embrapa Informação Tecnológica (2001).
- Bauer, H. M. et al. Genital human papillomavirus infection in female university students as determined by a PCR-based method. JAMA. 265, 472–77, https://doi.org/10.1001/jama.1991.03460040048027 (1991).
Article CAS PubMed Google Scholar - Buxton, D. et al. The pathogenesis of experimental neosporosis in pregnant sheep. J. Comp. Pathol. 118, 267–79, https://doi.org/10.1016/s0021-9975(07)80003-x (1998).
Article CAS PubMed Google Scholar - Bartley, P. M. et al. Detection of Neospora caninum DNA in cases of bovine and ovine abortion in the South-West of Scotland. Parasitol. 146, 979–82, https://doi.org/10.1017/S0031182019000301 (2019).
Article CAS Google Scholar - Hecker, Y. P. et al. Ovine abortion by Neospora caninum: first case reported in Argentina. Acta Parasitol. 1–6, https://doi.org/10.2478/s11686-019-00106-z (2019).
Article CAS Google Scholar - Burg, J. L., Grover, C. M., Pouletty, P. & Boothroyd, J. C. Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. J. Clin. Microbiol. 27, 1787–92 (1989).
Article CAS Google Scholar - Spalding, S. M., Amendoeira, M. R. R., Coelho, J. M. C. & Angel, S. O. Otimização da reação de polimerase em cadeia para detecção de Toxoplasma gondii em sangue venoso e placenta de gestantes. J. Bras. Med. Patol. Lab. 38, 105–10, https://doi.org/10.1590/S1676-24442002000200006 (2002).
Article CAS Google Scholar - Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 30, 2725–29, https://doi.org/10.1093/molbev/mst197 (2013).
Article CAS PubMed PubMed Central Google Scholar - Ronquist, F. & Huelsenbeck, J. P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 19, 1572–74, https://doi.org/10.1093/bioinformatics/btg180 (2003).
Article CAS PubMed Google Scholar - Tree Bio. FigTree [online]. London: Tree Bio; 2016 [cited 2019 November 10]. Available from, http://tree.bio.ed.ac.uk/software/figtree/.
- Csordas, B. G. et al. New insights from molecular characterization of the tick Rhipicephalus (Boophilus) microplus in Brazil. Rev. Bras. Parasitol. Vet. 25, 317–26, https://doi.org/10.1590/S1984-29612016053 (2016).
Article CAS PubMed Google Scholar
Acknowledgements
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) Finance Code 001. This study was financed in part by the Fundação Universidade Federal de Mato Grosso do Sul – UFMS/MEC – Brazil. The authors thank the Program for Technological Development in Tools for Health-PDTISFIOCRUZ for use of its facilities.
Author information
Authors and Affiliations
- Programa de Pós-Graduação em Doenças Infecciosas e Parasitárias- Universidade Federal de Mato Grosso do Sul (UFMS), Campo Grande, MS, Brasil
Pâmella Oliveira Duarte - Laboratório de Biologia Molecular do Carrapato, Departamento de Sanidade Animal, Embrapa Gado de Corte, Campo Grande, MS, Brasil
Leandra Marla Oshiro - Professor do Curso de Medicina Veterinária do Centro Universitário da Grande Dourados-UNIGRAN, Dourados, Brasil
Namor Pinheiro Zimmermann - Bolsista de Pós-Doutorado- FUNDAPAM, Laboratório de Biologia Molecular do Carrapato, Departamento de Sanidade Animal, Embrapa Gado de Corte, Campo Grande, MS, Brasil
Bárbara Guimarães Csordas - Laboratório de Toxonologia e Plantas Medicinais-Uniderp Agrárias, Campo Grande, Brasil
Doroty Mesquita Dourado - Empresa Brasileira de Pesquisa Agropecuária, Embrapa Gado de Corte, Campo Grande, MS, Brasil
Jacqueline Cavalcante Barros & Renato Andreotti
Authors
- Pâmella Oliveira Duarte
- Leandra Marla Oshiro
- Namor Pinheiro Zimmermann
- Bárbara Guimarães Csordas
- Doroty Mesquita Dourado
- Jacqueline Cavalcante Barros
- Renato Andreotti
Contributions
D.P.O. performed the experiments and wrote and edited the manuscript; O.L.M. and D.D.M. performed the experiments and conceptualized the study; Z.N.P. and C.B.G. performed the data analysis and statistical analyses; A.R. and C.J.B. acquired funding for the research, provided the study materials, and conceptualized the study. All authors read and approved the final manuscript.
Corresponding author
Correspondence toRenato Andreotti.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article
Cite this article
Duarte, P.O., Oshiro, L.M., Zimmermann, N.P. et al. Serological and molecular detection of Neospora caninum and Toxoplasma gondii in human umbilical cord blood and placental tissue samples.Sci Rep 10, 9043 (2020). https://doi.org/10.1038/s41598-020-65991-1
- Received: 03 December 2019
- Accepted: 13 May 2020
- Published: 03 June 2020
- Version of record: 03 June 2020
- DOI: https://doi.org/10.1038/s41598-020-65991-1