A phase I trial of cytotoxic T-lymphocyte precursor-oriented peptide vaccines for colorectal carcinoma patients (original) (raw)
Main
Recent advances in molecular biology and cellular immunology have resulted in identification of various antigens and epitopes recognised by human histocompatibility leukocyte antigen (HLA)-class-I-restricted cytotoxic T lymphocytes (CTLs) from various cancers (Bruggen et al, 1991; Brichard et al, 1993; Kawakami et al, 1994a, 1994b Fisk et al, 1995; Peoples et al, 1995; Robbins et al, 1996; Correale et al, 1997; Correale et al, 1998). Many clinical trials of peptide-based immunotherapy have shown an increased immune response to the vaccinated peptides (Rosenberg et al, 1998; Marchand et al, 1999; Jager et al, 2000; Gajewski et al, 2001; Gjertsen et al, 2001; Lau et al, 2001; Valmori et al, 2001). However, these initial clinical studies have produced few clinical responses in the various types of cancer treated including melanoma and colorectal cancer (Finn and Lotze, 2001; Miyagi et al, 2001; Sadanaga et al, 2001). This failure could be in part due to the insufficient and late stages of CTL induction by the current regimen, in which pre-vaccination screening of suitable peptides for each patient among a large number of candidate peptides is not conducted. As a result, vaccination in the present study began with immune priming. This standard regimen could be effective in preventing infectious disease. However, the main goal of a cancer vaccine is treatment of malignant disease. The time-lag necessary for priming of an antitumour response should be seriously considered, as the expected survival of most advanced cancer patients under these regimens is 3–9 months (Cole and Rodu, 2001; Miyagi et al, 2001). Therefore, the development of a new regimen of therapeutic vaccine is needed (Finn and Lotze, 2001). One regimen might include pre-vaccination measurement of peptide-specific CTL precursors in the circulation, followed by vaccination of only CTL-reactive peptides (CTL precursor-oriented vaccine). We have previously reported 14 vaccine candidate peptides that can induce HLA-A24-restriced and tumour-specific CTL in cancer patients (Gomi et al, 1999; Kikuchi et al, 1999; Yang et al, 1999; Harashima et al, 2000; Kawano et al, 2000; Nakao et al, 2000; Nishizaka et al, 2000). We have also shown that most cancer patients have peptide-specific CTL precursors for some of these peptides, and that peripheral blood mononuclear cells (PBMCs) stimulated with positive peptides show HLA-class-I-restricted and tumour-specific cytotoxicity (Hida et al, 2002; Maeda et al, 2002; Suzuki et al, 2002). In the present study, patients with advanced stages of colorectal cancer were immunised with up to four peptides identified in pre-vaccination measurement of peptide-specific CTL precursors to evaluate the toxicities and responses to CTL precursor-oriented peptide vaccination.
Materials and methods
Patients and eligibility criteria
The study protocol was approved by the Institutional Ethical Review Boards of Hokkaido University and Kurume University, respectively. Complete written informed consent was obtained from all of patients at the time of enrolment. According to the protocol, patients were required to be HLA-A24 positive, and to have a histologically confirmed lesion of colorectal carcinoma. Eligibility criteria included an age of 85 years or less, serum creatinine of less than 1.4 mg dl−1, bilirubin of less than 1.5 mg dl−1, a platelet count of 100 000 _μ_l−1 or more, haemoglobin of 8.0 g dl−1 or more, and total WBC of 3000 _μ_l−1 or more. Hepatitis B surface antigen, Hepatitis C antigen, and human immunodeficiency virus (HIV) were required to be negative. The patients, who had been untreated for at least 4 weeks before the study, had an Eastern Cooperative Oncology Group performance status of 0–2. The treatment was carried out at the First Department of Surgery, Hokkaido University School of Medicine (patients 1–9) and the Department of Surgery, Kurume University School of Medicine (patient 10) from November 2000 through April 2002. All immunological analyses were carried out at the Department of Immunology, Kurume University School of Medicine.
Screening of peptide-specific CTL precursors
A volume of 30 ml of peripheral blood was obtained pre- and post- (3rd, 6th, and 9th) vaccination, and PBMCs were isolated by means of Ficoll–Conray density gradient centrifugation, as reported previously (Miyagi et al, 2001). A previously reported method was used to detect peptide-specific CTL precursors in PBMCs (Hida et al, 2002; Suzuki et al, 2002). Briefly, PBMCs (1 × 105 cells well−1) were incubated with 10 μ M of a peptide in wells of u-bottom-type 96-well microculture plates (Nunc, Roskilde, Denmark) in 200 _μ_l of culture medium. The culture medium consisted of 45% RPMI-1640 medium, 45% AIM-V® medium (GIBCO BRL), 10% FCS, 100 U ml−1 of interleukin (IL)-2, and 0.1 μ M MEM nonessential amino-acid solution (GIBCO BRL). Every 3 days, half of the medium was removed and replaced with new medium containing a corresponding peptide (20 μ M). The assay was performed in quadruplicate. After a 12-day incubation, the cultured cells in one well were divided into four wells, two of which were used for corresponding peptide-pulsed C1R-A2402 cells, and the other two of which were used for HIV peptide (RYLRQQLLGI)-pulsed C1R-A2402 cells. The HLA-A24-binding HIV peptide was used as a negative control. After an 18-h incubation period, the supernatants were collected and the level of interferon (IFN)-γ was determined by enzyme-linked immunosorbent assay (ELISA) (limit of sensitivity: 10 pg ml−1). The background response to HIV peptide-pulsed C1R-A2402 cells was subtracted from the value. Assessment of peptide-specific CTL precursors was performed based on two parameters, the _P_-value by the Student's _t_-test and IFN-γ production, as described in legends of Tables 2 and 4. According to the results of this test, up to four positive peptides were selected for each patient, utilised in a skin test, and then, if negative, injected as a vaccination. The screening of peptide-specific CTL precursors was performed by the same method after the 6th vaccination to evaluate the effects of immunisation.
Table 2 Pre-vaccination screening of peptide-specific CTL precursors
Table 4 Summary of response to the peptide vaccination
Peptides and vaccination
The peptides utilised in the present study were prepared under conditions of Good Manufacturing Practice using a Multiple Peptide System (San Diego, CA, USA). Montanide ISA-51, an incomplete adjuvant, was manufactured by Seppic, Inc (Franklin Lakes, NJ, USA). The peptides were supplied in vials containing 3 mg ml−1 sterile solution for injection. A 3 mg portion of peptide with sterile saline was added in a 1 : 1 volume to Montanide ISA-51, then mixed in a Vortex mixer (Fisher, Inc, Alameda, CA, USA). The resulting emulsion was injected subcutaneously into the lateral thigh using a glass syringe. Patients were vaccinated every 14 days for a total of three injections to measure the toxicity. For the patients with no toxicity, the vaccinations were repeated biweekly up to 15 times with informed consent from each patient.
Delayed-type hypersensitivity (DTH) skin test
Skin tests were performed using 50 _μ_g of each peptide injected intradermally in a volume of 100 _μ_l using a tuberculin syringe and a 27-gauge needle. Saline was a negative control for assessment of DTH. At least 7 mm of induration or erythema read 48 h after injection was needed to score the skin test as positive (Nestle et al, 1998).
Assay of cytotoxicity
Cytotoxic activity was measured using a standard 6-h 51Cr-release assay (Gomi et al, 1999; Miyagi et al, 2001). Cryopreserved PBMCs were thawed and cultured in the medium consisting of 45% RPMI-1640 medium, 45% AIM-V® medium (GIBCO BRL), 10% FCS, 100 U ml−1 of IL-2, and 0.1 μ M MEM nonessential amino-acid solution (GIBCO BRL). On the 14th day of culture, the cells were harvested and served for the assay. To avoid the bias of bioassays, PBMCs harvested at different times from a single patient were thawed at the same time. SW620 (HLA-A24+, colon adenocarcinoma), COLO201 (HLA-A24−, colon adenocarcinoma), and PHA-blastoid T cells (HLA-A24+) were used as target cells (1 × 103 well−1), and 50-fold unlabelled K562 cells were added into wells to decrease nonspecific killing activity.
Purification of CD8+ or CD4+ T cells
CD8+ or CD4+ T cells were positively isolated from peptide-stimulated PBMCs using the CD8 Positive Isolation Kit (DYNAL, Oslo, Norway) or the CD4 Positive Isolation Kit (DYNAL) according to the manufacturer's instructions. In both cases, the percentage of purified CD8+ or CD4+ T cells was more than 90% (data not shown).
Kinetics of peptide-specific CTL precursors
To detect the kinetics of peptide-specific CTL precursor frequency in patient 1, PBMCs from before and after vaccination were incubated at 100 cells per well of a 96-well u-bottom microculture plate in the presence of feeder cells without the peptides. Cells from each well were harvested on the 14th day of culture and tested for their ability to produce IFN-γ by recognition of peptide-pulsed CIR-A2402 cells in duplicate assay. The well was considered positive if it contained effector cells producing much higher than 100 pg ml−1 and also statistically significant levels (P<0.05 by the Student's _t_-test) of IFN-γ in response to CIR-A2402 cells preloaded with a corresponding peptide as compared with those in response to the HIV peptide-pulsed CIR-A2402 cells.
Detection of serum immunoglobulin G (IgG) levels
An ELISA was used to detect the serum IgG levels specific to the peptides administered, as reported previously (Miyagi et al, 2001). Briefly, the peptide (20 _μ_g well−1)-immobilised plate was blocked with Block Ace (Yukijirushi, Tokyo, Japan), washed, and 100 _μ_l well−1 of serum samples diluted with 0.05% Tween 20-Block Ace were added to the plate. After a 2-h incubation, the plate was washed and further incubated for 2 h with an 1 : 1000-diluted rabbit anti-human IgG (DAKO, Glostrup, Denmark). The plate was washed, after which 100 _μ_l of 1 : 100-diluted goat anti-rabbit Ig-conjugated horseradish peroxidase-dextran polymer (En Vision, DAKO) was added to each well, and the plate was incubated for 40 min. After washing, 100 _μ_l well−1 of tetramethyl-benzidine substrate solution (KPL, Guildford, UK) was added, and the reaction was stopped by the addition of 1 M phosphoric acid. To estimate peptide-specific IgG levels, the optical density values of each sample were compared with those of serially diluted standard samples, and the values are shown as the optical density. The specificity of the ELISA was tested as follows. Serum samples (1 : 100 diluted) were incubated in wells precoated with corresponding or irreverent peptides to absorb the peptide-specific IgG. After a 2-h incubation at room temperature, samples were transferred to new wells precoated with the sample peptide used in the first absorption, and this protocol was repeated for a total of three times. The samples were then subjected to peptide-specific IgG ELISA.
Evaluation of treatment response
All known sites of disease were evaluated by CT-scan or X-ray examination pre- and post-vaccinations (the 3rd, 6th, 9th, and 12th). Patients were assigned a response category according to the response evaluation criteria in solid tumours, a revised version of the WHO criteria published in the WHO Handbook for reporting results of cancer treatment in June 1999.
Results
Demographics of the patients
In all, 10 patients with advanced colorectal carcinomas were enroled in this phase I study. Demographic details of the patients are shown in Table 1. The median patient age was 67 years (range 28–78). All patients had liver, lung, peritoneum, or lymph node metastases. All patients underwent surgical resection of the primary lesion; nine had failed previous chemotherapy. All 10 patients completed the first three vaccinations within the protocol under informed consent, and all of them received more vaccinations (6–15) under additional informed consent.
Table 1 Patient characteristics
Screening of peptide-specific CTL precursors
All tumour-related antigens were identified by screening of a cDNA library from tumour cells using tumour-reactive CTLs. The peptides used for this study have the potential to induce HLA-A24-restricted and tumour-specific CTL activity in PBMCs of HLA-A24+ cancer patients (Kikuchi et al, 1999; Yang et al, 1999; Gomi et al, 1999; Harashima et al, 2000; Kawano et al, 2000; Nakao et al, 2000; Nishizaka et al, 2000). Pre-vaccination PBMCs were provided for screening of the CTL precursors reactive to the 14 candidate peptides, followed by selection of peptides based on evaluation of the results with the criteria shown in Table 2. The assay was performed in quadruplicate. After a 12-day incubation, the cultured cells in one well were separated into four wells, two of which were used for corresponding peptide-pulsed C1R-A2402 cells, and the other two of which were used for HIV peptide-pulsed C1R-A2402 cells. The HLA-A24-binding HIV peptide was used as a negative control. The assessment of peptide-specific CTL precursors was performed based on two parameters, the _P_-value by the Student's _t_-test and IFN-γ production, as shown in the table legend. When these peptides were found to induce immediate-type hypersensitivity by a skin test, a fifth peptide was vaccinated if it proved negative in the skin test. SART2899, CyB91, ART1170, and ART413 were positive for immediate-type hypersensitivity in all patients tested and were not injected at all. As a result, five patients were injected with four peptides, three patients with three peptides, and two with two peptides. The vaccinated peptides for each patient are shown in Table 2. It is noteworthy that the profiles of the vaccinated peptides varied greatly among the 10 patients.
Toxicities
All 10 patients were evaluated for toxicity; the overall toxicities are shown in Table 3. The vaccinations were generally well-tolerated, but almost all patients (eight out of 10) had grade I or II local redness and swelling at the injection sites. Fever with mild flu-like symptoms was observed in four patients (grade I or grade II), although this symptom was transient and no medication was needed. Grade I fatigue or nausea was observed in two patients, and grade I anorexia, diarrhoea, or vomiting was observed in one. No vaccine-related grade III or IV toxicity was observed (data not shown). There was no clinical evidence of an autoimmune reaction as determined by symptoms, physical examination, or laboratory test.
Table 3 Toxicities associated with the peptide vaccination
Cellular immune responses
Post-vaccination (6th) PBMCs showed increased amounts of peptide-specific IFN-γ production compared to pre-vaccination PBMCs in five out of 10 patients (1, 2, 5, 6, and 10), as described in Table 4. Representative results of patients 1 and 2 are shown in Figure 1A. In patients 1 and 2, CTL response to the lck208 was apparently induced after the 6th vaccination. In five other patients, peptide-specific CTL response decreased. We further tested the reactivity of purified CD4+ or CD8+ T cells in response to the administered peptides. The pre- or post-6th vaccination PBMCs from patient 2 were in vitro stimulated, and purified CD4+ or CD8+ T cells were tested for their reactivity to the SART3109 peptide-pulsed C1R-A2402 cells. As shown in Figure 1B, purified CD8+ T cells from the post-vaccination PBMCs of patient 2 produced IFN-γ in an antigen-specific manner, although no definite IFN-γ production specific to the SART3109 peptide was observed when unseparated post-6th PBMCs from patient 2 were used (Figure 1A). Purified CD4+ T cells failed to produce IFN-γ in a peptide-specific manner. On the other hand, no peptide-specific IL-4 production was observed in the case with purified CD8+ or CD4+ T cells (data not shown).
Figure 1
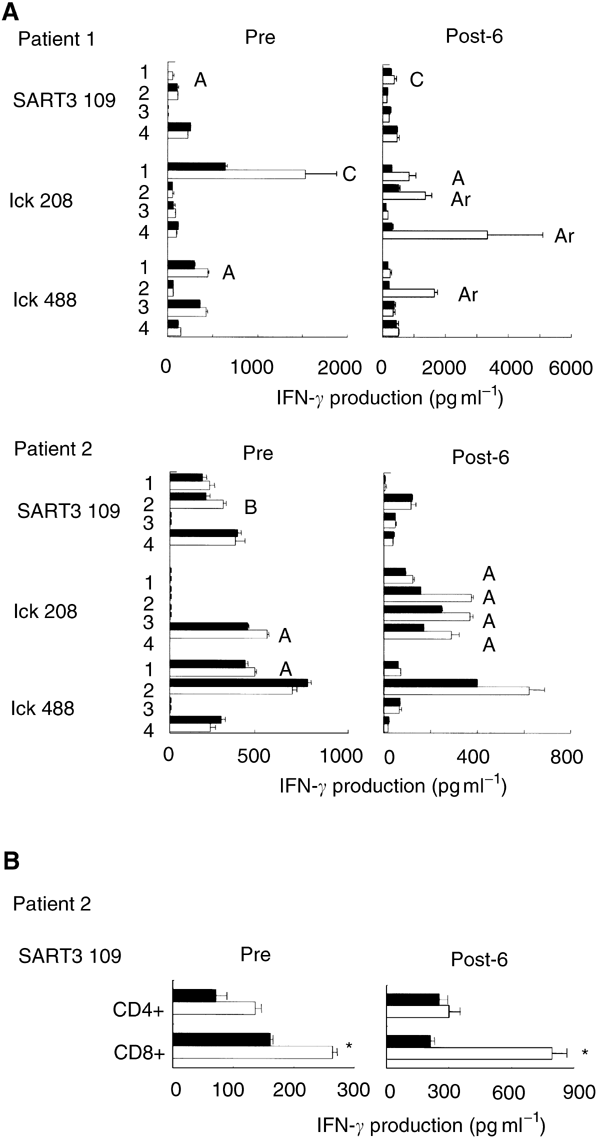
Assay of peptide-specific CTL precursors. (A) Pre- and post- (6th) vaccination PBMCs were provided for screening of reactivity to each of the 14 peptides listed in Table 2 in the quadruplicate assays. Representative results of patients 1 and 2 are shown in this figure. The peptide-stimulated PBMCs were cultured with C1R-A2402 cells that were preloaded with the corresponding peptide (open bar) or the HIV peptide (closed bar). The level of IFN-γ in the supernatant was determined by ELISA. The result was evaluated by the classification shown in the legend of Table 4. Each alphabet character represents the result of each well. (B) Pre- and post- (6th) vaccination PBMCs from patient 2 were stimulated in vitro with the SART3109 peptide. The peptide-stimulated PBMCs were harvested, and positively isolated CD4+ or CD8+ T cells were cultured in triplicate with C1R-A2402 cells that were preloaded with the SART3109 peptide (open bar) or the HIV peptide (closed bar). The level of IFN-γ in the supernatant was determined by ELISA. *Statistically significant at P<0.05.
We next examined cytotoxicity of pre- and post- (3rd, 6th, and 9th) vaccination PBMCs from eight patients against SW620 (HLA-A24+ colon tumour cells), COLO201 cells (HLA-A24− colon tumour cells), and PHA-activated T cells (HLA-A24+) (Figure 2). Tumour-related antigens from which all peptides used in this study were derived are nonmutated self-antigens overexpressed in tumour cells, including SW620 and COLO201 (Shichijo et al, 1998; Yang et al, 1999; Gomi et al, 1999; Harashima et al, 2000; Kawano et al, 2000; Nakao et al, 2000; Nishizaka et al, 2000). As shown in four cases of patients 2, 3, 7, and 9, cytotoxicity against HLA-A24+ SW620 increased after peptide vaccination compared to that against HLA-A24− COLO201. In the other four cases, no definite increase in cytotoxicity was observed after the peptide vaccination. No cytotoxicity against HLA-A24+ PHA-blastoid T cells was detected in any case. These results indicate that the peptide vaccination resulted in augmented CTL activity in four out of eight patients.
Figure 2
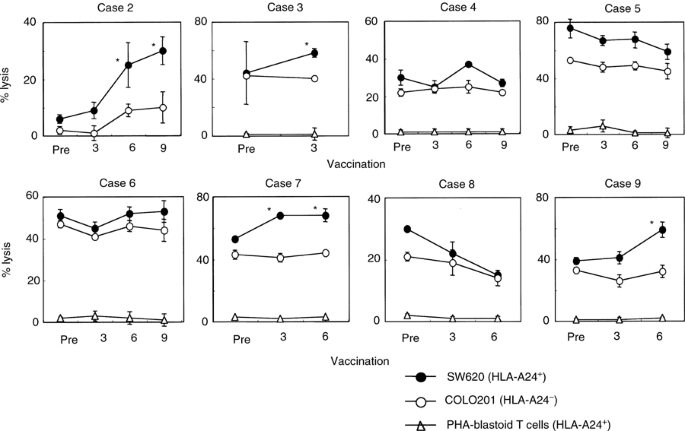
Cytotoxicity before and after the peptide vaccination. Pre- and post-vaccination PBMCs from eight patients were incubated for 14 days with IL-2 alone without any peptides in culture, followed by measurement of cytotoxicity against SW620 cells (HLA-A24+ colon cancer cell line), COLO201 cells (HLA-A24− colon cancer cell line), and PHA-activated T cells (HLA-A24+) by a 6-h 51Cr-release assay at an E/T ratio of 40/1. The assay was performed in triplicate, and the mean and s.d. are shown. *Statistically significant at P<0.05.
Serum IgG specific to peptides
No IgG reactive to any of the vaccinated peptides was detected in pre-vaccination sera from any of the 10 patients (Table 4). Significant levels of anti-peptide IgG reactive to SART3- or lck-derived peptides became detectable in the post-vaccination sera of seven patients. Detail results are shown in Figure 3. In patient 1, IgGs reactive to the lck208 and the lck488 peptides were induced after the 6th and 9th vaccinations, respectively (Figure 3A). A similar result was observed in six other patients (Figure 3B). It is noteworthy that three patients who showed strong peptide-specific CTL response (criteria Ar) after the peptide vaccination were also positive for antipeptide IgG to the corresponding peptides (patient 1 for lck208 and lck488, patient 6 for SART3109, and patient 10 for SART3315). In addition, the IgG response to the lck208 peptide in post-vaccination sera of patient 1 was neutralised by absorption with a corresponding peptide, but not with the lck408 peptide, whereas the opposite was observed in the case of the IgG response to the lck488 peptide (Figure 3C). This peptide-specific absorption demonstrates the validity of the ELISA system.
Figure 3
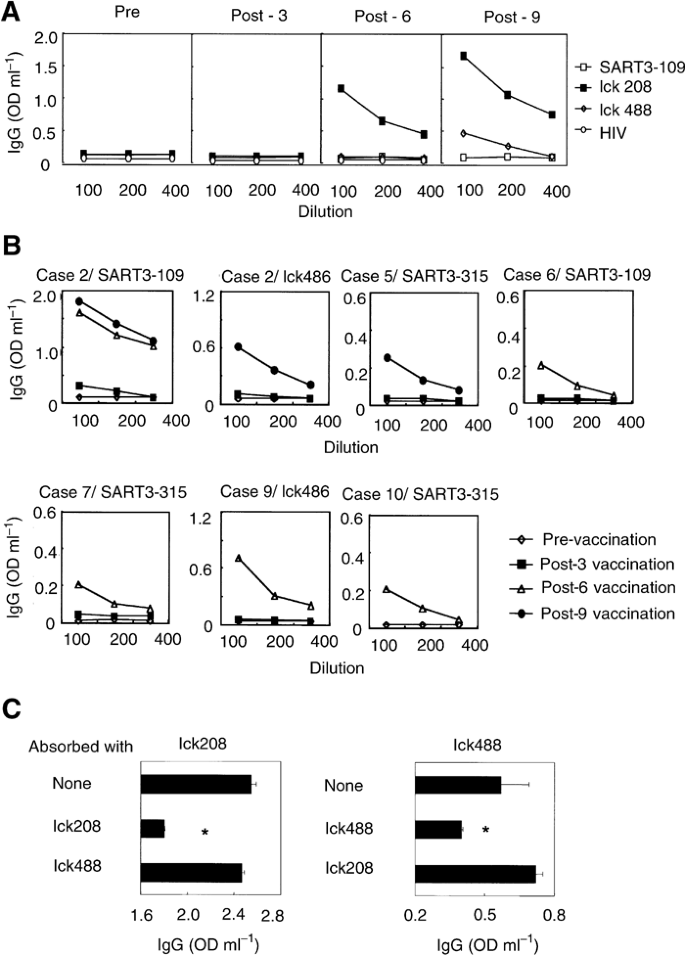
IgG reactive to the vaccinated peptides. (A) Pre- and post-vaccination sera from patient 1 were serially diluted and the levels of IgGs reactive to three administered peptides were determined by ELISA. (B) Pre- and post-vaccination sera from patients 2, 5, 6, 7, 9, and 10 were serially diluted and the levels of IgGs reactive to corresponding peptides were determined by ELISA. (C) Diluted sera of patient 1 after the 9th vaccination were cultured in the indicated peptide-coated wells and the levels of IgGs reactive to corresponding peptides were determined by ELISA. *Statistically significant at P<0.05.
DTH skin test
No DTH reaction against peptides was observed before vaccination in any patient, while peptide-specific DTH reactions were observed in three patients after the peptide vaccination (Table 4). In patient 1, DTH reactions to lck208 and lck488 were observed after the 6th vaccination, and DTH against SART3109 became detectable after the 9th vaccination. Patient 4 exhibited a DTH reaction against lck208, lck486, and lck488 after the 3rd vaccination. Patient 7 exhibited a DTH reaction against SART3109 after the 3rd vaccination.
Clinical results
The clinical responses of each patient are summarised in Table 4. CT scans of patient 1 pre- and post-vaccination are shown in Figure 4A. In all, 48% regression (27–14 mm) of liver metastasis was observed in patient 1. This patient had para-aortic lymph node metastasis at a diameter of less than 20 mm; it showed no change after the peptide vaccination (data not shown). Because his clinical response was considered a partial response (PR), this patient was analysed more in detail. The kinetic analysis of tumour cell lysis in patient 1 indicates that increased CTL activity to the SW620 cells became detectable in post-vaccination PBMCs (Figure 4B). The cytotoxicity against lck208 peptide-loaded CIR-A2402 cells became significant after the 3rd and 6th vaccinations (Figure 4C). While no peptide-specific IFN-γ production was detected in any of the 96 wells containing 100 cells well−1 of pre-vaccination PBMCs, SART3109-specific IFN-γ production was detected in two, two, and four wells among 96 wells containing post-vaccination (3rd, 6th, and 9th) PBMCs, respectively (Figure 4D). Production of lck208-specific IFN-γ production was detectable in three and four wells among 96 wells containing the 6th and 9th vaccination PBMCs, while lck488-specific IFN-γ production could be observed in one and two wells among 96 wells containing the 6th- and 9th-vaccination PBMCs, respectively. The patient has subsequently been treated only by vaccination (SART3109, lck208, and lck488) for 7 months as an outpatient, and is still doing well. Patient 2 had intrapelvic metastasis, and the disease has remained stable (s.d.) for 6 months. The eight other patients showed progressive disease (PD) 2–4 months after starting the vaccinations, although all have been treated as outpatients and their quality of life has been evaluated as quite high.
Figure 4
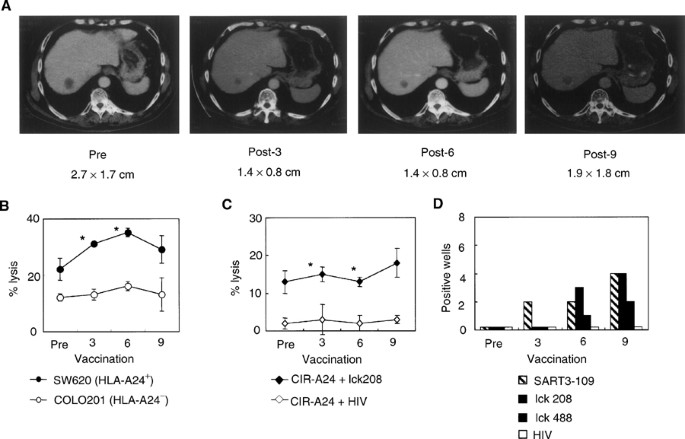
Clinical and immunological responses to the peptide vaccination. (A) CT scans show tumour regression of the liver metastasis after the peptide vaccination. The size of the liver metastasis (S8) is described. (B) CTL activity before and after vaccinations. Cytotoxicity to SW620 cells (HLA-A24+ colon cancer cell line), COLO201 cells (HLA-A24− colon cancer cell line), and PHA-activated T cells (HLA-A24+) was tested by a 6-h 51Cr-release assay at an E/T ratio of 40/1. Values are the means of triplicate assay. *Statistically significant at P<0.05. (C) Peptide-specific CTL activity before and after the peptide vaccinations. Cytotoxicity to CIR-A2402 cells preloaded with the lck208 or the control HIV peptide was tested by a 6-h 51Cr-release assay at an E/T ratio of 40/1. *Statistically significant at P<0.05. (D) Kinetics of peptide-specific CTL precursors. Pre- and post- (3rd, 6th, and 9th) vaccination PBMCs were incubated at 100 cells per well in a 96-well round-microculture plate in the presence of feeder cells. The number of wells producing a significant level of IFN-γ in a peptide-specific manner among 96 wells is shown.
Discussion
Patients undergoing this regimen received 3 mg of peptides biweekly for up to four peptides. All of the peptides used were derived from nonmutated self-antigens involved in cellular proliferation (Kikuchi et al, 1999; Gomi et al, 1999; Yang et al, 1999; Harashima et al, 2000; Kawano et al, 2000; Nakao et al, 2000; Nishizaka et al, 2000). However, there was no grade III or IV adverse effect, which is consistent with previous observations in studies of peptide-based vaccinations. Most of the patients received the vaccination as outpatients, and the performance status remained very good throughout the treatment periods. Therefore, in terms of safety, this regimen of CTL precursor-oriented peptide vaccine could be recommended as a cancer vaccine suitable for further clinical trials.
Another aim of our study was to assess the clinical response to the vaccination. In our limited number of case, we observed one PR and one s.d. continuing for more than 6 months. Both of these cases were treated with three kinds of SART3- and lck-derived peptides, suggesting that the combined use of these peptides might constitute a promising vaccine strategy for advanced colorectal carcinomas, thus encouraging us to plan a phase II trial utilising these peptides.
Vaccination-induced immunity was evaluated in this study by several different methods, including IFN-γ production in response to peptides, a standard 6-h 51Cr-release assay, measurement of antipeptide antibody, and DTH responses. An elevated immune response to lck208 and lck488 was detected in post-vaccination PBMCs by all of the methods used in the samples of patient 1, who showed PR. This patient's PBMCs also reacted to the SART3109 peptide, as measured by frequency analysis of cellular responses to peptides (Figure 4D) and also by DTH test (Table 4). These results indicate that the patient's PBMCs reacted to all three vaccinated peptides after the peptide vaccination. Post-vaccination PBMCs from patient 2, who had a long s.d., responded to lck208 peptide alone, and the post-vaccination sera became positive for both the SART3109 and lck486 peptides, although no DTH response was observed (Table 4). Besides patient 1, positive DTH response was observed in only two patients (4 and 7), with PD, but their post-vaccination PBMCs showed no increase in cellular responses to the administered peptides. On the other hand, besides patients 1 and 2, IgG reactive to the administered peptides became detectable in the post-vaccination sera of five other patients (5, 6, 7, 9, and 10) with PD. Although the post-vaccination PBMCs of patients 6 and 10 showed an increase in cellular responses to SART3109 and SART3315, respectively, no augmentation of peptide-specific cellular response was observed in other cases. Neither a cellular nor humoral immune response to administered peptides was detectable in the remaining two patients (3 and 8), who also had PD. These results suggest that vaccination-induced immunity varies considerably among patients. However, we recently reported that the in vivo induction of IgG reactive to administered peptides is positively correlated with clinical response or the survival of patients with prostate, lung, gastric, or gynaecological cancer (Mine et al, 2003; Noguchi et al, 2003; Sato et al, 2003; Tsuda et al, 2004). This may be the case with patients 1 and 2, who showed PR and s.d., respectively, because IgGs reactive to two different peptides were induced only in these two patients. We have no clear answer regarding the role of peptide-specific IgG in antitumour immune response, and are now grappling with this theme.
In addressing the mechanism for peptide-specific IgG induction after peptide vaccination, one possibility is that 9-mer or 10-mer peptide-recognizing CD4+ T cells were involved in this phenomenon. In general, in vivo generation of antigen-specific IgG requires a cytokine from helper T cells (Parker, 1993). Although peptides binding to MHC class II molecules have been suggested to be 12–25 amino acids in length, the core sites anchored to MHC class II molecules are sufficient even at a length of about nine amino acids (Rammensee et al, 1995). Indeed, we recently observed that peptide vaccination with a 9-mer peptide could induce peptide-specific and HLA-DR-restricted CD4+ T cells in vivo (Harada et al, 2004). CD4+ T-cell help is required during the generation and maintenance of effective anti-tumour CD8+ T cell-mediated immunity. The requirement of CD4+ T-cell help to initiate and sustain a CD8+ T-cell response has been well established and has led to the development of antitumour vaccines that attempt to induce both T-cell subsets (Knutson et al, 2001). The in vivo induction of IgG reactive to administered peptides may be indirect evidence of the involvement of CD4+ T lymphocytes.
We recently developed a culture system to evaluate CTL precursors against many peptides using a limited number of PBMCs from cancer patients (Hida et al., 2002); the same culture system was applied to this study. The main reason why we assessed peptide-specific CTL precursors based on two parameters, the _P_-value and IFN-γ production, was that the levels of IFN-γ produced by peptide-specific CTLs varied among quadruplicate wells. This finding might be due to the small number of cells (105 cells well−1) that were initially placed in each well. It is possible that one well may have contained peptide-specific CTL precursors, whereas another may have contained none. We concluded that each well should be individually estimated to screen for the presence of peptide-specific CTL precursors.
Recent reports revealed that a Th2 response is predominant in cancer patients (Pellegrini et al, 1996; Vita et al, 1999; Sheu et al, 2001). Therefore, we examined the level of IL-4 during peptide stimulation in vitro, but the level of IL-4 production was generally low, and no peptide-specific IL-4 production was observed. In contrast, the level of IFN-γ production was constantly substantial. Probably, the in vitro culture of PBMCs in the presence of IL-2 could preferentially activate natural killer cells, and natural killer cell-derived IFN-γ might provide an optimal condition for Th1 type cells.
In conclusion, vaccination of colorectal cancer patients with peptides by the CTL precursor-oriented method was a well-tolerated outpatient treatment and induced antigen-specific immunity as well as a clinical response. Even though only a small number of selected patients were treated, the encouraging clinical response demands further studies of CTL precursor-oriented vaccine in other human cancers.
Change history
16 November 2011
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
- Brichard V, Pel A, Wölfel T, Wölfel C, Plaen E, Lethé B, Coulie P, Boon T (1993) The tyrosinase gene codes for an antigen recognized by autologous cytotoxic T lymphocytes on HLA-A2 melanoma. J Exp Med 178: 489–495
Article CAS Google Scholar - Bruggen P, Traversari C, Chomez P, Lurquin C, Plaen E, Eynde B, Boon T (1991) A gene encoding an antigen recognized by cytotoxic T lymphocytes on a human melanoma. Science (Washington DC) 254: 1643–1647
Article Google Scholar - Cole P, Rodu B (2001) Descrictive epidemiology: cancer statistics. In Cancer Principles & Practice of Oncology, Vincent Jr TD, Samuel H, Rosenberg SA (eds) 6th Edition pp 229–230. Lippincott Williams & Wilkins
Google Scholar - Correale P, Walmsley K, Nieroda C, Zaremba S, Zhu M, Schlom J (1997) In vitro generation of human cytotoxic T lymphocytes specific for peptides derived from prostate-specific antigen. J Natl Cancer Inst 89: 293–300
Article CAS Google Scholar - Correale P, Walmsley K, Zaremb S, Zhu M, Schlom J, Tsang KY (1998) Generation of human cytotoxic T lymphocyte lines directed against prostate-specific antigen (PSA) employing a PSA oligoepitope peptide. J Immunol 161: 3186–3194
CAS Google Scholar - Finn OJ, Lotze MT (2001) A decade in the life of tumor immunology. Clin Cancer Res 7: 759–760
Google Scholar - Fisk B, Blevins TL, Wharton J, Ioannides C (1995) Identification of an immunodominant peptide of HER2/neu proto-oncogene recognized by ovarian tumor-specific cytotoxic T lymphocytes line. J Exp Med 181: 2109–2117
Article CAS Google Scholar - Gajewski TF, Fallarino F, Ashikari A, Sherman M (2001) Immunization of HLA-A2+ melanoma patients with MAGE-3 or MelanA peptide-pulsed autologous peripheral blood mononuclear cells plus recombinant human interleukin 12. Clin Cancer Res (Suppl) 7: 895s–901s
CAS Google Scholar - Gjertsen MK, Buanes T, Rosseland AR, Bakka A, Gladhaug I, Soreide O (2001) Interdermal ras peptide vaccination with granulocyte–macropharge colony-stimulating factor as adjuvant: clinical and immunological responses in patients with pancreatic adenocarcinoma. Int J Cancer 92: 441–450
Article CAS Google Scholar - Gomi S, Nakao M, Niiya F, Imamura Y, Kawano K, Nishizaka S, Itoh K (1999) A cyclophilin B gene encodes antigenic epitopes recognized by HLA-A24-restricted and tumor-specific cytotoxic T lymphocytes. J Immunol 163: 4994–5004
CAS Google Scholar - Harada M, Gohara R, Matsueda S, Muto A, Oda T, Iwamoto Y, Itoh K (2004) In vivo evidence that peptide vaccination can induce HLA-DR-restricted CD4+ T cells reactive to a class I tumor peptide. J Immunol 172: 2659–2667
Article CAS Google Scholar - Harashima N, Tanaka K, Sasatomi T, Shimizu K, Miyagi Y, Yamada A, Itoh K (2000) Recognition of the Lck tyrosine kinase as a tumor antigen by cytotoxic T lymphocytes of cancer patients with distant metastases. Eur J Immunol 31: 323–332
Article Google Scholar - Hida N, Maeda Y, Katagiri K, Takasu H, Harada M, Itoh K (2002) A simple culture protocol to detect peptide-specific cytotoxic T lymphocyte precursors in the circulation. Cancer Immunol Immunother 51: 219–228
Article CAS Google Scholar - Jager E, Gnjatic S, Nagata Y, Stockert E, Jager D, Karbach J, Neuman A, Rieckenberg J, Chen YT, Ritter G, Hoffman E, Arand M, Old LJ, Knuth A (2000) Induction of primary NY-ESO-1 immunity: CD8+ T lymphocyte and antibody responses in peptide-vaccinated patients with NY-ESO-1+ cancers. Proc Natl Acad Sci USA 97: 12198–12203
Article CAS Google Scholar - Kawakami Y, Eliyahu S, Delgado CH, Robbins PF, Sakaguchi K, Appella E, Rosenberg SA (1994b) Identification of a human melanoma antigen recognized by tumor-infiltrating lymphocytes associated with in vitro tumor rejection. Proc Natl Acad Sci USA 91: 6458–6462
Article CAS Google Scholar - Kawakami Y, Eliyahu S, Sakaguchi K, Robbins PF, Rivoltini L, Yannelli JR, Rosenberg SA (1994a) Identification of the immunodominant peptides of the MART-1 human melanoma antigen recognized by the majority of HLA-A2 restricted tumor infiltrating lymphocytes. J Exp Med 180: 347–352
Article CAS Google Scholar - Kawano K, Gomi S, Tanaka K, Tsuda N, Kamura T, Itoh K, Yamada A (2000) Identification of a new endoplasmic reticulum-resident protein recognized by HLA-A24-restricted tumor infiltrating lymphocytes of lung cancer. Cancer Res 60: 3550–3558
CAS Google Scholar - Kikuchi M, Nakao M, Inoue Y, Matsunaga K, Shichijo S, Yamna H, Itoh K (1999) Idnetification of a SART-1-derived peptides capable of inducing HLA-A24-restricted and tumor-specific cytotoxic T lymphocytes. Int J Cancer 81: 459–466
Article CAS Google Scholar - Knutson KL, Schiffman K, Disis ML (2001) Immunization with a HER2/neu helper peptide vaccine generates HER2/neu CD8 T-cell immunity in cancer patients. J Clin Invest 107: 477–484
Article CAS Google Scholar - Lau R, Wang F, Jeffery G, Marty V, Kuniyoshi J, Bade E (2001) Phase I trial of intravenous peptide-pulsed dendritic cells in patients with metastatic melanoma. J Immunol 24: 66–78
CAS Google Scholar - Maeda Y, Hida N, Niiya F, Katagiri K, Yamana H, Kamura T, Takahashi M, Sato Y, Todo S, Itoh K (2002) Detection of peptide-specific CTL-precursors in peripheral blood lymphocytes of cancer patients. Br J Cancer 87: 796–804
Article CAS Google Scholar - Marchand M, Baren VB, Weynants P, Brichard V, Dreno B, Tessier MH, Rankin E, Parmiani G, Arienti F, Humblet Y, Bourlond A, Vanwijck R, Lienard D, Beauduin M, Dietrich PY, Russo V, Kerger J, Masucci G, Jager E, De Greve J, Atzpodien J, Brasseur F, Coulie PG, Van Der Bruggen P, Boon T (1999) Tumor regressions observed in patients with metastatic melanoma treated with an antigen peptide encoded by MAGE-3 and presented by HLA-A1. Int J Cancer 80: 219–230
Article CAS Google Scholar - Mine T, Gouhara R, Hida N, Imai N, Azuma K, Rikimaru T, Katagiri K, Nishikori M, Sukehiro A, Nakagawa M, Yamada A, Aizawa H, Shirouzu K, Itoh K, Yamana H (2003) Immunological evaluation of CTL precursor-oriented vaccines for advanced lung cancer patients. Cancer Science 94: 548–556
Article CAS Google Scholar - Miyagi Y, Imai N, Sasatomi T, Yamada A, Mine T, Katagiri K, Nakagawa M, Muto A, Okouchi S, Isomoto H, Shirouzu K, Yamana H, Itoh K (2001) Induction of cellular immune responses to tumor cells and peptides in colorectal cancer patients by vaccination with SART3 peptides. Clin Cancer Res 7: 3950–3962
CAS Google Scholar - Nakao M, Shichijo S, Imaizumi T, Inoue Y, Matsunaga K, Yamada A, Itoh K (2000) Identification of a gene coding for a new squamous cell carcinoma antigen recognized by the CTLs. J Immunol 164: 2565–2574
Article CAS Google Scholar - Nestle FO, Alijagic S, Gilliet M, Sun Y, Grabbe S, Dummer R Burg G, Schadendorf D (1998) Vaccination of melanoma patients with peptide- or tumor lysate-pulsed dendritic cells. Nat Med 4: 328–332
Article CAS Google Scholar - Nishizaka S, Gomi S, Harada K, Oizumi K, Itoh K, Shichijo S (2000) A new tumor-rejection antigen recognized by cytotoxic T lymphocytes infiltrating into a lung adenocarcinoma. Cancer Res 60: 4830–4837
CAS Google Scholar - Noguchi M, Mine T, Suetsugu N, Tomiyasu K, Suekane S, Yamada A, Itoh K, Noda S (2003) Induction of cellular and humoral immune responses to tumor cells and peptides in HLA-A24 positive hormone-refractory prostate cancer patients by peptide vaccination. Prostate 57: 80–92
Article CAS Google Scholar - Parker DC (1993) T cell-dependent B-cell activation. Ann Rev Immunol 11: 331–360
Article CAS Google Scholar - Pellegrini P, Berghella AM, Del Beato T, Cicia S, Adorno D, Casciani CU (1996) Dysregulation in TH1 and TH2 subsets of CD4+ T cells in peripheral blood of colorectal cancer patients and involvement in cancer establishment and progression. Cancer Immunol Immunother 42: 1–8
Article CAS Google Scholar - Peoples GE, Geodegebuure PS, Smith R, Linehan DC, Yoshino I, Eberlein TJ (1995) Breast and ovarian cancer-specific cytotoxic T lymphocytes recognized the same HER2/neu-derived peptide. Proc Natl Acad Sci USA 92: 432–436
Article CAS Google Scholar - Rammensee HG, Frige T, Stevanovic S (1995) MHC ligands and peptide motifs. Immunogenetics 41: 178–228
Article CAS Google Scholar - Robbins PF, El-Gamil M, Li YF, Kawakami Y, Loftus D, Appella E, Rosenberg SA (1996) A mutated _β_-catenin gene encodes a melanoma-specific antigen recognized by tumor infiltrating lymphocytes. J Exp Med 183: 1185–1192
Article CAS Google Scholar - Rosenberg SA, Yang JC, Schwartzentruber DJ, Hwu P, Marincola FM, Topalian SL, Restifo NP, Dudley ME, Schwarz SL, Spiess PJ, Wunderlich JR, Parkhurst MR, Kawakami Y, Seipp CA, Einhorn JH, White DE (1998) Immunologic and therapeutic evaluation of a synthetic peptide vaccine for the treatment of patients with metastatic melanoma. Nat Med 4: 321–327
Article CAS Google Scholar - Sadanaga N, Nagashima H, Mashino K, Tahara K, Yamaguchi H, Ohta M, Fujie T, Tanaka F, Inoue H, Takesako K, Akiyoshi T, Mori M (2001) Dendritic cell vaccination with MAGE peptide is a novel therapeutic approach for gastrointestinal carcinomas. Clin Cancer Res 7: 2277–2284
CAS PubMed Google Scholar - Sato Y, Shomura H, Maeda Y, Mine T, Ueno Y, Akasaka Y, Kondo M, Takahashi S, Shinohara T, Katagiri K, Sato M, Okada S, Matsui K, Yamada A, Yamana H, Itoh K, Todo S (2003) Immunological evaluation of peptide vaccination for patients with gastric cancer based on pre-existing cellular response to peptide. Cancer Sci 94: 802–808
Article CAS Google Scholar - Sheu BC, Lin RH, Lien HC, Ho HN, Hsu SM, Huang SC (2001) Predominant Th2/Tc2 polarity of tumor-infiltrating lymphocytes in human cervical cancer. J Immunol 167: 2972–2978
Article CAS Google Scholar - Shichijo S, Nakao M, Imai Y, Takasu H, Kawamoto M, Niiya F, Itoh K (1998) A gene encoding antigenic peptides of human squamous cell carcinoma recognized by cytotoxic T lymphocytes. J Exp Med 187: 277–288
Article CAS Google Scholar - Suzuki N, Maeda Y, Tanaka S, Hida N, Mine T, Yamamoto K, Oka M, Itoh K (2002) Detection of peptide-specific cytotoxic T lymphocyte precursors used for specific immunotherapy of pancreatic cancer. Int J Cancer 98: 45–50
Article CAS Google Scholar - Tsuda N, Mochizuki K, Harada M, Sukehiro A, Kawano K, Yamada A, Ushijima K, Sugiyama T, Nishida T, Yamana H, Itoh K, Kamura T (2004) Vaccination with pre-designated or evidence-based peptides for patients with recurrent gynecologic cancers. J Immunother 27: 60–72
Article CAS Google Scholar - Valmori D, Dutoit V, Rubio-Goboy V, Chambaz C, Lienard D, Guillaume P (2001) Frequent cytotoxic T-cell responses to peptide MAGE-A10254–262 in melanoma. Cancer Res 61: 509–512
CAS PubMed Google Scholar - Vita FD, Orditura M, Galizia G, Romano C, Infusino S, Auriemma A, Lieto E, Catalano G (1999) Serum interleukin-10 levels in patients with advanced gastrointestinal malignancies. Cancer 86: 1936–1943
Article Google Scholar - Yang D, Nakao M, Shichijo S, Sasatomi T, Takasu H, Matsumoto H, Itoh K (1999) Identification of a gene coding for a protein possessing shared tumor epitopes capable of inducing HLA-A24-restricted cytotoxic T lymphocytes in cancer patients. Cancer Res 59: 4056–4063
CAS Google Scholar
Acknowledgements
We would like to express our gratitude to the patients who participated in this study and to the oncologists who referred their patients to us. This work was supported in part by Grants from the Ministry of Education, Science, Sport, Culture, and Technology of Japan (11178101 to KI), and the Ministry of Health, Labor, and Welfare of Japan (H2-genome-003, 11-16, and H12-cancer-004 to KI)
Author information
Authors and Affiliations
- First Department of Surgery, Hokkaido University School of Medicine, Sapporo, Japan
Y Sato, Y Maeda, H Shomura, M Takahashi, Y Une, M Kondo, T Shinohara, K Sato, M Sato & S Todo - Department of Immunology, Kurume University School of Medicine, Fukuoka, Japan
H Shomura, N Hida, K Katagiri, A Yamada, M Harada & K Itoh - Department of Surgery, Kurume University School of Medicine, Fukuoka, Japan
T Sasatomi & H Yamana
Authors
- Y Sato
You can also search for this author inPubMed Google Scholar - Y Maeda
You can also search for this author inPubMed Google Scholar - H Shomura
You can also search for this author inPubMed Google Scholar - T Sasatomi
You can also search for this author inPubMed Google Scholar - M Takahashi
You can also search for this author inPubMed Google Scholar - Y Une
You can also search for this author inPubMed Google Scholar - M Kondo
You can also search for this author inPubMed Google Scholar - T Shinohara
You can also search for this author inPubMed Google Scholar - N Hida
You can also search for this author inPubMed Google Scholar - K Katagiri
You can also search for this author inPubMed Google Scholar - K Sato
You can also search for this author inPubMed Google Scholar - M Sato
You can also search for this author inPubMed Google Scholar - A Yamada
You can also search for this author inPubMed Google Scholar - H Yamana
You can also search for this author inPubMed Google Scholar - M Harada
You can also search for this author inPubMed Google Scholar - K Itoh
You can also search for this author inPubMed Google Scholar - S Todo
You can also search for this author inPubMed Google Scholar
Corresponding author
Correspondence toY Sato.
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Sato, Y., Maeda, Y., Shomura, H. et al. A phase I trial of cytotoxic T-lymphocyte precursor-oriented peptide vaccines for colorectal carcinoma patients.Br J Cancer 90, 1334–1342 (2004). https://doi.org/10.1038/sj.bjc.6601711
- Revised: 28 November 2003
- Accepted: 16 January 2004
- Published: 02 March 2004
- Issue Date: 05 April 2004
- DOI: https://doi.org/10.1038/sj.bjc.6601711