Lactobacillus rhamnosus GG Lysate Increases Re-Epithelialization of Keratinocyte Scratch Assays by Promoting Migration (original) (raw)
Introduction
The potential beneficial effects of the oral consumption of probiotic bacteria on intestinal health have been extensively investigated. Probiotics are usually members of the genera Lactobacillus or Bifidobacterium and may exert a positive benefit on the gut using through a variety of mechanisms, including inhibition of the growth of pathogenic bacteria1, epithelial repair, enhancement of the gut barrier2 and modulation of the immune response3. Due to their potential for maintaining gut health and combating disease, various species of lactobacilli have been tested in tissues other than the gut, for example the vagina and the oral cavity4,5 and there is a growing body of evidence that probiotic bacteria may also be of benefit in these tissues. Our interest has been in the development of topically applied lactobacilli as treatments for skin in health and/or disease6,7,8. Our focus on lactobacilli is based on the fact that in contrast to skin microbiota, lactobacilli are relatively well characterised in terms of their safety and mechanisms of action. Thus, we believe that the development of lactobacilli as therapeutics for dermal applications may be more rapid than would be the case for numerically dominant members of the of skin microbiota.
The skin forms a barrier between the entire body and the outside environment without which life as a terrestrial organism would be impossible. The epidermis of skin prevents both water loss from within the body and ingress of potential pathogens9. The importance of the skin barrier is exemplified in conditions where it is severely breached. For example, morbidity and mortality in victims of severe burns are associated with the dehydration and infection that occurs due to the poor skin barrier in these conditions10. However, as well as these extreme situations, breaches in the skin barrier can occur in daily life due to wounding and in surgery11. In healthy individuals, wounds normally heal in a timely fashion12. However, due to underlying conditions such as diabetes, some wounds heal slowly or not at all. In these cases, infection is a potential complication that can then further inhibit wound healing13,14. Chronic wounds are therefore a significant pathology. It follows then that treatments that can promote the wound healing process will be of considerable benefit to patients.
In the gut, certain species of probiotic bacteria have been demonstrated to increase barrier repair in in vitro and vivo models15,16. For example, healing of acid-induced gastric ulcers in the rat is reportedly accelerated in the presence of L. rhamnosus GG and L. gasseri16. Because of their purported effectiveness at promoting the healing of gastric wounds, attention has begun to focus on whether probiotic bacteria can impact upon the healing of cutaneous wounds. A recent study in rabbits showed that patches containing L. fermentum accelerated wound healing17. L. plantarum has been shown to improve healing of burns in mice and humans in a limited number of studies and also reportedly promotes healing of leg ulcers in humans17,18,19,20. However, in general, the mechanisms underlying these effects have not been fully explored.
We hypothesised that bacterial lysates would represent a safer alternative to the use of live bacteria in a wound situation because although probiotic lactobacilli have a GRAS (generally regarded as safe) status for food, the potential risks of live probiotic entering the bloodstream through breached skin has not been assessed. Furthermore, the use of lysates may be of more utility to potential wound care manufacturers than live bacteria because the logistical requirements of maintaining viability of bacteria within a formulation or wound dressing can be bypassed. In this study therefore, we have compared the ability of lysates made from four different species of lactobacilli to accelerate an important aspect of wound healing, re-epithelialization. To this end the scratch assay a well-established wound model21, was used to assess the effect of Lactobacillus lysates on re-epithelialization of human primary keratinocytes, the main cell type present within the epidermal layer of skin.
Results
Bacterial lysates accelerate scratch closure in a species dependent manner
Re-epithelialization in keratinocyte monolayers treated with L. rhamnosus GG lysate was significantly accelerated compared to that of untreated monolayers. At 18 h, 95% (P = 0.03, n = 3) of the scratch area was re-epithelialized compared with 75.2% in the control monolayer (Fig. 1a,b). Similar results were found when scratched cells were subjected to treatment with L. reuteri lysate, where 90.4% (P = 0.05, n = 3) of the scratch area was closed at 18 h (Fig. 1a,b). In contrast, L. plantarum lysate did not stimulate monolayer re-epithelialization (70.5%, n = 3). L. fermentum lysates caused a significant reduction in monolayer re-epithelialization rate compared with the control (40%, P = 0.01, n = 3). In keeping with the literature, the positive control, Keratinocyte growth factor (KGF) stimulated re-epithelialization of scratches although it was no more efficacious than the L. rhamnosus GG lysate (Fig. 1).
Figure 1
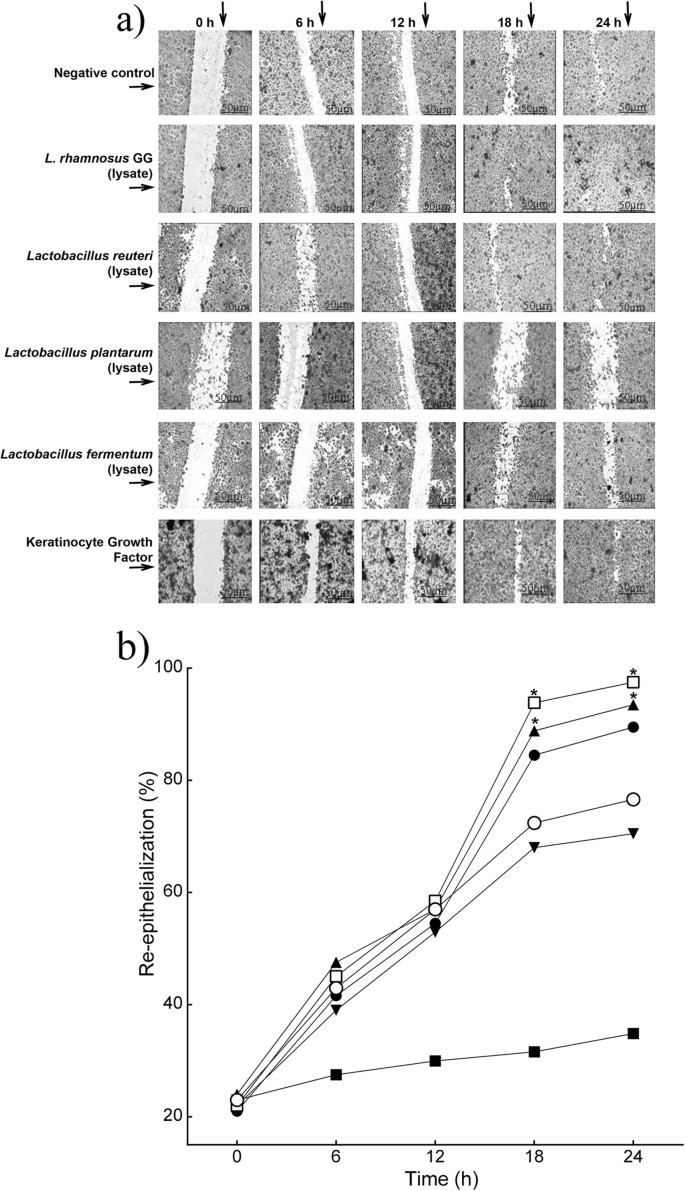
Specific probiotic lysates stimulate keratinocyte re-epithelialization in vitro.
(a) Representative images of monolayer re-epithelialization in the presence/absence of different treatments. (b) The graph shows the percentage of scratch re-epithelialization in cultures treated with/without probiotic lysates at different time points. Results are expressed as the mean ± SEM. *P < 0.05. (○), control, untreated; (□), L. rhamnosus GG lysate; (●), L. reuteri lysate; (▼), L. plantarum; (■), L. fermentum lysate; (▲), keratinocyte Growth Factor.
Bacterial lysates increase keratinocyte migration and proliferation
Since only L. rhamnosus GG and L. reuteri were efficacious in the scratch assay, these two lysates were analyzed further. Re-epithelialization could be due to keratinocyte migration and/or proliferation. Hence, the effect of L. rhamnosus GG and L. reuteri lysates on these keratinocyte activities was tested alongside that of the positive control KGF. In cultures treated with L. rhamnosus GG lysate and L. reuteri lysate there were about 48 × 105 and 24 × 105 migrated cells respectively after 4 h (P = 0.001, P = 0.004, respectively, n = 3), compared with 7 × 105 cells in control cultures (Fig. 2a). Importantly, there was a significant difference between two lysates (P = 0.002, n = 3), with L. rhamnosus GG lysate being the most efficacious (Fig. 2a) in stimulating cell migration. KGF stimulated 34 × 105 cells to migrate. However, there was no significant difference between the effects of KGF and L. rhamnosus GG lysate on keratinocyte migration. Incubation of keratinocytes with 100 μl of L. rhamnosus GG lysate or L. reuteri lysate resulted in significant proliferation as double the number of cells was found in these cultures relative to control cultures (P = 0.02, P = 0.03, respectively, n = 3) at 12 h post-inoculation (Fig. 2b). KGF caused a two-fold increase in the number of cells compared with the untreated control (Fig. 2b). Interestingly, both lysates and KGF afforded equal stimulation of proliferation.
Figure 2
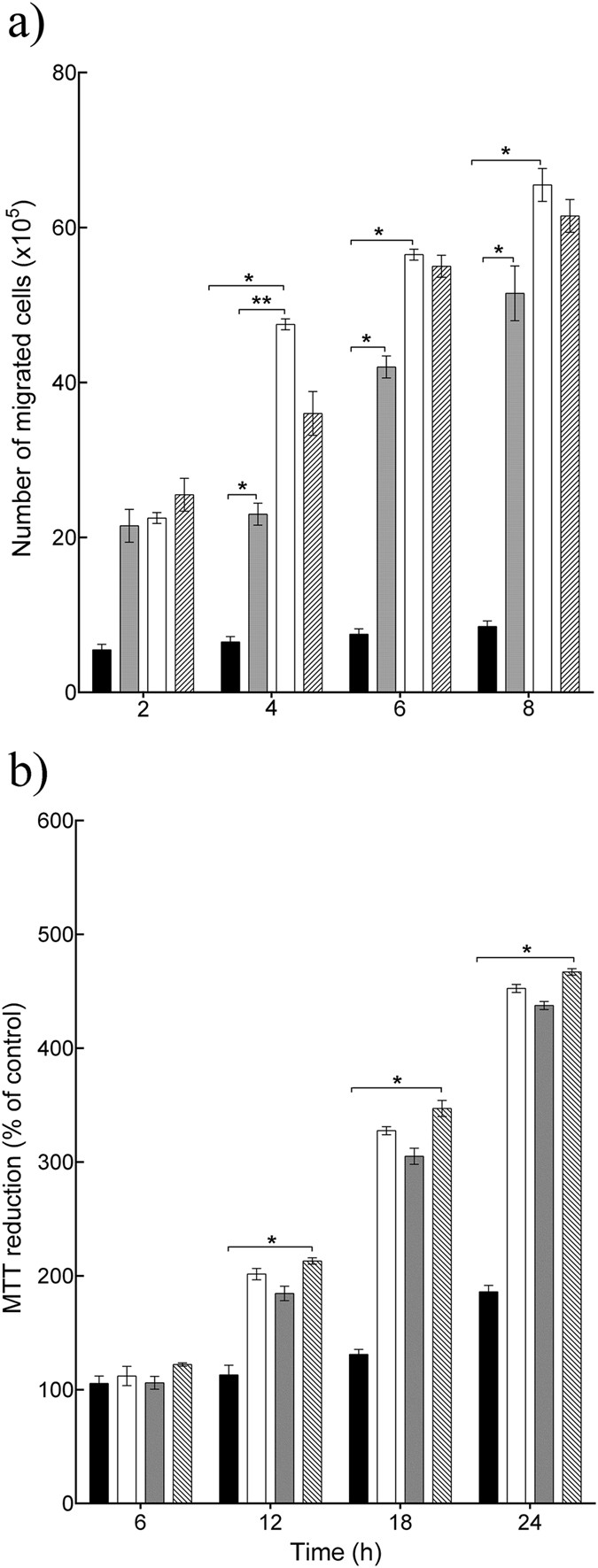
L. rhamnosus GG lysate and L. reuteri lysate increased keratinocyte migration and proliferation.
(a) L. rhamnosus GG lysate and L. reuteri lysate significantly increased the number of migrated cells (P = 0.001, P = 0.004, respectively n = 3) after 4 h incubation. However, L. rhamnosus GG lysate was more efficacious than L. reuteri lysate (**P = 0.002, n = 3) and had similar effect to Keratinocyte Growth Factor. (b) L. rhamnosus GG lysate and L. reuteri lysate significantly increased cell proliferation after12h incubation. Results are expressed as the mean ± SEM, *P < 0.05. (■), control; (□) L. rhamnosus GG lysate; ( ), L. reuteri lysate; (
), L. reuteri lysate; ( ), keratinocyte Growth Factor.
), keratinocyte Growth Factor.
L. rhamnosus GG induces cell migration as the dominant mechanism
Since the L. reuteri and L. rhamnosus GG lysates both stimulated migration and proliferation of keratinocytes, we next asked which of these mechanisms was most dominant in the acceleration of re-epithelialization induced by the two lysates. The rate of re-epithelialization of scratched keratinocyte monolayers was measured in the presence of the well-characterised inhibitor of cell proliferation, Mitomycin C. The data in Fig. 3a demonstrate that in monolayers treated with L. rhamnosus GG lysate and Mitomycin C, 94.5% of the scratch area re-epithelialized after 18h incubation (P = 0.03, n = 3) which was not significantly different to the re-epithelialization of monolayers treated with L. rhamnosus GG alone. However, in the presence of Mitomycin C the L. reuteri lysate- treated monolayer was only 52% re-epithelialized compared to 90% in cells treated with L. reuteri lysate alone (P = 0.01, n = 3, Fig. 3b).
Figure 3
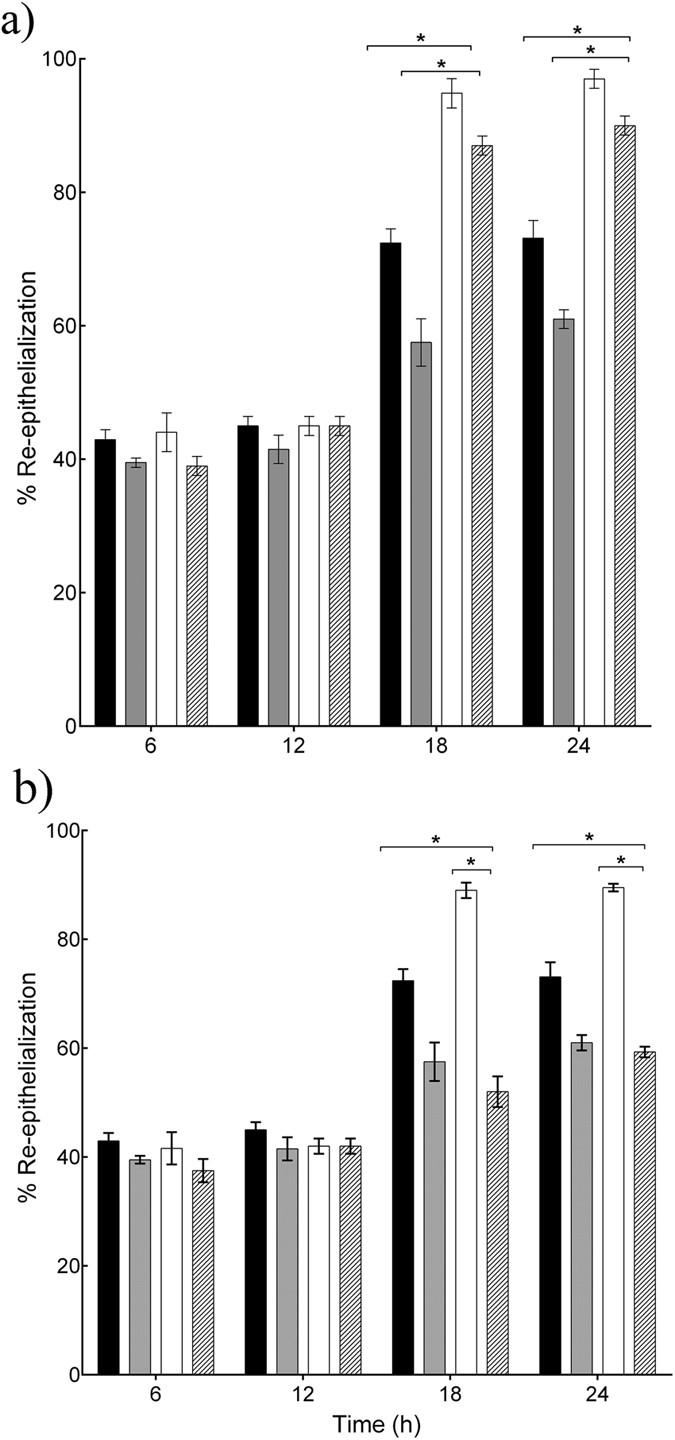
L. rhamnosus GG and L. reuteri enhance re-epithelialisation by stimulating migration and proliferation of keratinocytes.
(a) L. rhamnosus GG lysate accelerated re-epithelialisation of scratches even in the presence of Mitomycin C (*P = 0.03, n = 3) after 18h incubation. (b) However, in cells treated with L. reuteri lysate and Mitomycin C, the percentage of re-epithelialisation was reduced significantly to 52% ± 2.32 at 18 h, compared to 90% ± 1.13 in cells treated with L. reuteri lysate alone (P = 0.01, n = 3) at 18 h. Results are expressed as the mean ± SEM, *P < 0.05. (■), control; ( ), control and Mitomycin C; (□), L. reuteri lysate; (
), control and Mitomycin C; (□), L. reuteri lysate; ( ), L. reuteri lysate and Mitomycin C.
), L. reuteri lysate and Mitomycin C.
Genome-wide Affymetrix microarray study of the effects of the L. rhamnosus GG lysate on keratinocyte gene expression
Our data suggest that the L. rhamnosus GG lysate increases re-epithelialization by enhancing keratinocyte migration. Therefore, the pathways underlying this effect were explored using microarray analysis of scratched keratinocytes treated vs. untreated with L. rhamnosus GG lysate. The genes identified in the microarray were grouped using IPA according to cellular function. Table 1 considers functions known to be relevant to the wound-healing process10,22,23. The groups of genes involved in ‘cell movement’, ‘migration’ and ‘proliferation’ had the highest P value suggesting modulation of these genes by the bacterial lysate. The group of genes that was most significantly altered in expression was ‘cell movement’. However, all the genes contained within this group were also common to the group called ‘cell migration’. More than 1760 genes involved in migration, were altered in response to the L. rhamnosus GG lysate. The top 10 genes from the common group between ‘migration’ and ‘cell movement’ are presented in Table 2.
Table 1 Bioinformatics analysis of the effects of L. rhamnosus GG lysate on keratinocyte activities.
Table 2 L. rhamnosus GG modulates the expression of genes involved in cell migration.
Table 2 shows that the expression of two genes, CXCL2 and its receptor CXCR2 were highly up-regulated by the L. rhamnosus GG lysate. The microarray was validated by performing quantitative PCR analysis of CXCL2 and CXCR2 mRNA levels in scratched keratinocytes treated with L. rhamnosus GG lysate vs untreated controls (data not shown). Since the L. rhamnosus GG lysate up-regulated the gene expression of CXCL2 and its corresponding receptor, we next investigated whether the proteins corresponding to these genes were also increased by the lysate. The secretion of CXCL2 by scratched keratinocytes in response to L. rhamnosus GG lysate after 12 h and 18 h incubation was measured using ELISA. These showed that increased CXCL2 secretion was stimulated by L. rhamnosus GG lysate-treated keratinocytes, with 6.30 ± 0.4 μg/ml, compared with 3.10 ± 0.7 μg/ml in untreated cells after 12h incubation (P = 0.002, n = 3, Fig. 4a). Immunoblot analysis was performed to quantify the changes in the levels of CXCR2 protein receptors in cells 12 h post-treatment with L. rhamnosus GG lysate. Analysis of extracts demonstrated that L. rhamnosus GG lysate caused an up-regulation of CXCR2 protein levels amounting to four times greater than in untreated cells (P = 0.0009, n = 3, Fig. 4b,c).
Figure 4
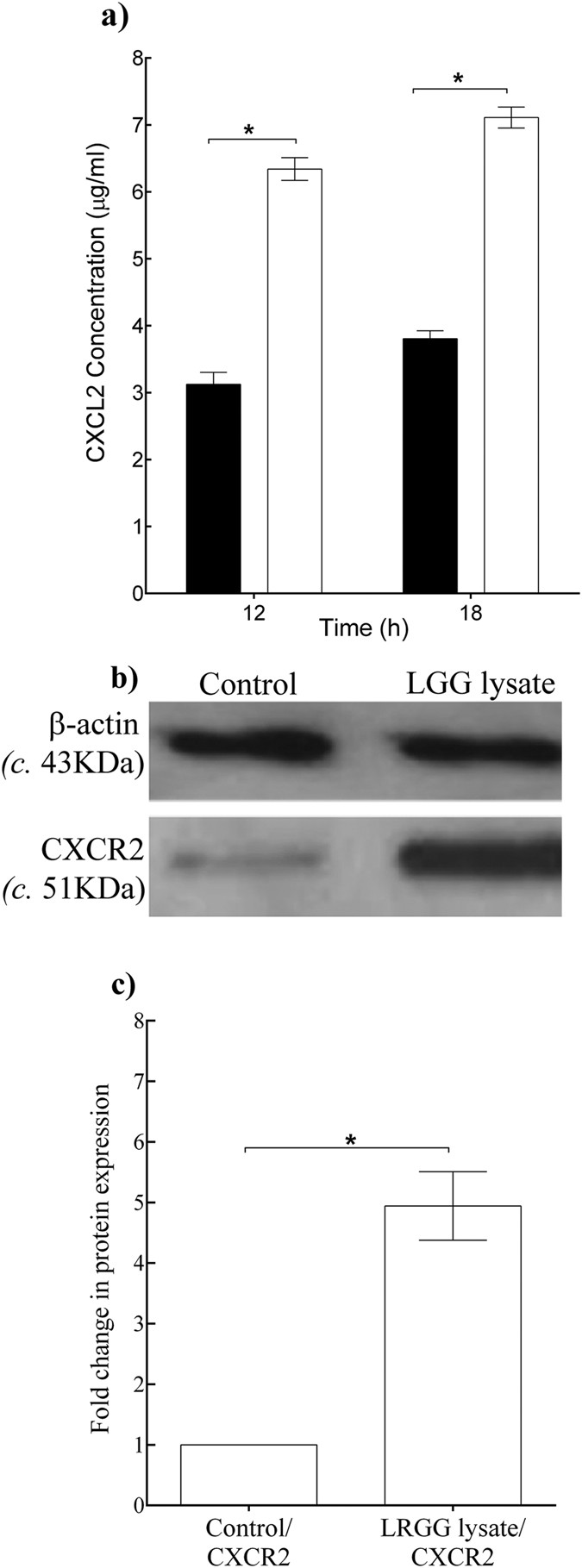
L. rhamnosus GG lysate increased the protein levels of CXCL2/CXCR2 in scratched keratinocyte cultures.
(a) Untreated control cultures produced 3.10 μg/ml ± 2.32 of CXCL2 whereas treated cultures produced significantly higher concentrations of CXCL2, compared with untreated controls (P = 0.002, n = 3) after 12 h incubation. (b) Representative images of immuno-blots for CXCR2. (c) Densitometry analysis of the immunoblots was performed to quantify the change in protein levels for treated cell cultures vs. untreated controls (control). Changes were normalized against β-actin. L. rhamnosus GG lysate increased CXCR2 protein level compared to untreated control (P = 0.0009, n = 3). Data are representative of three individual experiments and are expressed as the mean ± SEM, *P < 0.05. For (a) (■), control; (□) L. rhamnosus GG lysate.
Discussion
Non-healing wounds such as diabetic foot ulcers are a significant cause of morbidity and can lead to mortality24. Over-use of antibiotics has given rise to many antibiotic-resistant infections that are frequently associated with poor outcomes in wound healing, especially following surgery25. Currently, there is an unmet, clinical need for new therapies in the treatment of wounds. Since probiotics have been demonstrated to promote wound healing in the gut15, the aim of this study was to examine the effects of enteric probiotic lysates on keratinocyte migration and proliferation that are important aspects of the wound healing response.
Lysates of L. rhamnous GG and L. reuteri promoted the re-epithelialization of keratinocyte monolayers. In contrast, lysates from the other lactobacilli tested either had no effect on re-epithelialization, (L. plantarum) or inhibited re-epithelialization (L. fermentum). This latter result is probably related to our previous observation that L. fermentum reduces keratinocyte viability6,8. A previous study examined the effects of cell-free supernatants of L. plantarum on the wound healing process especially, the proliferation and migration of human keratinocytes in vitro26. The study demonstrated that the L. plantarum cell free supernatant promoted re-epithelialization in keratinocyte cultures. It was hypothesized that L. plantarum produces a specific substance, Plantaricin A that stimulates keratinocyte migration and proliferation in a scratch assay26. The authors investigated the mechanism underlying improved migration and proliferation and showed increased expression of transforming growth factor (TGF-β1), vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF-7). In the present study, the effect of spent culture fluid of L. plantarum on the stimulation of keratinocyte re-epithelialization was not measured. Therefore, it is not possible to exclude the likelihood that the spent culture fluid from L. plantarum may have been efficacious in our assay. What is clear is that lysates of both L. reuteri and L. rhamnosus GG are more efficacious than the L. plantarum lysate in promoting keratinocyte proliferation/migration.
The enhanced re-epithelialization induced by L. rhamnosus GG and L. reuteri was probably due to increased keratinocyte migration and proliferation although the two lysate were not equally effective in promoting cell migration. L. rhamnosus GG was much more effective at stimulating migration than L. reuteri. Indeed, migration is probably the dominant mechanism promoted by L. rhamnosus GG because in the presence of Mitomycin C, scratches were still able to re-epithelialize at a significantly faster rate than untreated scratches. Conversely, the dominant mechanism for the L. reuteri lysate was probably stimulation of proliferation, because Mitomycin C completely negated the stimulatory effects of the L. reuteri lysate. This suggests that lysates from different lactobacilli have species-specific effects on keratinocyte functions and is in agreement with previous observations6,7,8.
The mechanism underlying the increase in re-epithelialization induced by L. rhamnosus GG potentially involves increased expression of the chemokine, CXCL2 and its receptor, CXCR2. The expression of these genes was greatly enhanced at both the mRNA and protein levels as demonstrated by microarray, q-PCR, ELISA and immunoblotting data. CXCL2 is known to have several roles in the wound-healing process such as the stimulation of keratinocyte migration, proliferation and adhesion27,28. Indeed, it has been suggested that CXCL2 may be a chemoattractant for human keratinocytes27,28. Moreover, a number of vitro studies have shown that CXCL2 stimulates the production of additional chemokines, such as CXCL12 and CXCL10 also act as chemoattractants for keratinocytes in the wound-healing process26,29,30. The CXCL2 receptor, CXCR2 is also known to be important to the wound healing process. Mice lacking CXCR2 have delayed wound healing and this was related to a delay in cell recruitment to the wound site and a delay in re-epithelialization26,29,30. Based on these findings and the present data, we hypothesize that L. rhamnosus GG lysate increases re-epithelialization potentially via the interaction of CXCR2 and CXCL2 that accelerates keratinocyte migration, although other factors could also be involved.
A study conducted in 2011 by Polk31 reported that material derived from L. rhamnosus GG regulated intestinal epithelial cell survival and growth in injury via stimulating epidermal growth factor (EGF) and its receptor31,32. The microarray data in the present study did not highlight a role for EGF nor its receptor alpha chain EGFR-α since the gene levels of neither of these were significantly altered in cells treated with L. rhamnosus GG lysate. However, some of the main genes in the EGF signaling cascade, such as EGFR-β, MEK1, STAT3 and ADAM17, were significantly up regulated in response to L. rhamnosus GG lysate. CXCL2 and CXCR2 were both significantly upregulated and higher than the expression of genes associated with the EGF pathway. Polk’s study32 also identified specific proteins from L. rhamnosus GG spent medium fluid that promoted cell growth in cultured human and mouse colon epithelial cells. In the current study, the spent culture fluid was tested but found not to be as efficacious as the lysate (Figure S1). It is possible that this is due to a concentration effect i.e. the effective concentration is not sufficiently high in the spent culture fluid for an effect on re-epithelialization to be observed. Conversely, the L. rhamnosus GG lysate may be working via different receptors and ligands in keratinocytes than has been reported for enterocytes29,30,31.
In conclusion, L. rhamnosus GG lysate is a potential new wound healing treatment. We have demonstrated previously6 that this lysate also has protective effects against S. aureus toxicity towards keratinocytes. Thus, it is possible that L. rhamnosus GG lysate may have utility in the treatment of infected, non-healing wounds.
Materials and Methods
Bacterial cell culture
Lactobacillus rhamnosus GG (ATCC 53103) Lactobacillus reuteri (ATCC55730), Lactobacillus plantarum (UCC118) (ATCC10241) and Lactobacillus fermentum (ATCC14932) were purchased LGC Ltd, Ireland, UK. These were grown routinely in Wilkins-Chalgren Broth or Agar (Oxoid, Basingstoke, UK) at 37 °C incubated in an anaerobic cabinet (atmosphere, 10:10:80, H2-CO2-N2). Lysates of all these species were prepared as previously described6.
Primary keratinocyte culture and scratch assay
Primary human epidermal keratinocytes were cultured as described in7. For scratch assays, keratinocytes were plated on 24 well tissue culture plates and allowed to reach confluence. At this point a scratch was introduced into the monolayer using a sterile p200 pipette tip. The cells were then washed in phosphate buffered saline (0.01M PBS, pH = 7.4) to remove the debris and the medium was replaced with either fresh medium (control samples) or medium containing 100 μl bacterial lysate (0.93 mg/μl). In some experiments, keratinocyte growth factor (KGF, 0.5 mg/ml) was included as a positive control.
In other experiments, 0.5 mg/ml Mitomycin C from Streptomyces caespitosus was added before the addition of the lysate to inhibit cell proliferation. Scratches were then monitored and documented at 0,6,12,18 and 24 h post scratch by staining cells with crystal violet solution. Images were captured under the Keyence microscope (x5 magnification, Keyence, Osaka, Japan) and analysed using IMAGE J-64 software (http://imagej.nih.gov). This enabled determination of the percentage of scratch area (gap) with respect to the percentage of starting scratch area at time zero. For each image, the area between one side of the scratch and the other was measured. Quantification of scratch closure was performed by comparing the area of the scratch at set time points according to the equation:

where t = t is a specific time point post scratching. The experiment was performed three times with triplicate samples within each individual experiment by using different keratinocytes donors.
Keratinocyte migration assays
A migration assay was performed by plating c. 2.5 × 105 cells per well in the upper wells of a 24-TranswellTM chamber (Invitrogen, Life Technologies Ltd, Paisley) and then the 100 μl bacterial lysate under test, or the positive control, keratinocyte growth factor (KGF, 0.5 mg/ml) was placed in the lower chamber. The chambers were separated by an 8 μm pore-size permeable membrane that allowed the keratinocytes to move down toward chemo-attracting lysates. At 2, 4, 6 and 8h post inoculation, the membrane was stained with crystal violet to visualise the cells. Non-migrating cells in the upper surface were removed, whereas cells adhering to the lower surface of the membrane were considered as having migrated. The number of migrated cells was determined by counting the cells in three high powered fields (images were captured at 40 × magnification; bars = 100 μm) and this number was subtracted from the initial number of cells seeded i.e. 2.5 × 105. The experiment was performed three times with triplicate samples within each individual experiment.
Keratinocyte proliferation assay
The proliferation assay (CellTiter 96® AQueous) was performed according to the manufacturer’s instructions (Promega, Madison, USA). Cells were seeded at an initial density of 2.5 × 105 cells per well in 96-well plates. After 24 h incubation, cells were exposed to 100 μl of probiotic lysates or to the positive control, KGF at 0.5 mg/ml. Plates were then incubated at 37 °C, for 6, 12, 18 and 24 h. Once each time point was reached, the medium was aspirated and replaced with 100 μl per well of the CellTiter 96® Aqueous reagent. After 3 h of incubation, the absorbance of the solutions was measured at 490 nm in a Titertek micro-plate reader (Flow Laboratories Ltd. Leyland, UK). The absorbance of cells in the lysate-treated wells was compared with that in their untreated counterparts. The absorbance of 2.5 × 105 cells (original seeding number) was taken as the control absorbance and that of treated cells was normalized to this control. The experiment was conducted on three separate occasions with triplicate samples within each individual experiment.
Micro-array analysis of scratch assays
Confluent keratinocytes were scratched and exposed to bacterial lysate for 12 h at 37 °C in 5% CO2. Cells were washed twice in phosphate buffered saline (0.01M PBS, pH = 7.4) and the total RNA was extracted from treated and untreated control cells using 1.0 ml Trizol (Invitrogen, Life Technologies Ltd, Paisley, UK) per 10 cm2 of the culture dish. RNA was purified using an Ambion PureLink® RNA Mini Kit according to the manufacturer’s instructions (Invitrogen, Life Technologies Ltd, Paisley). The samples gathered from three individual extractions were analysed on an Agilent 2100 Bioanalyzer Instruments (Agilent Technologies, CA, United States) using the Human Genome U133 plus 2.0 Affymetrix GeneChips according to the manufacturer’s instructions. Bioinformatics analysis was performed with dChip (V2005) (www.dchip.org). The model-based analysis of oligonucleotide arrays and gene expression analysis were carried out using Bioconductor as described in33. Filtering for probe sets with a P ≤ 0.05 created a list of differentially expressed genes. P value refers to the alteration in gene expression in treated vs untreated cells. Data was analysed using the Ingenuity Knowledge Base program (IPA, http://www.ingenuity.com/products/ipa) with the aim of identifying the genes related to the wound-healing process. The microarray data was submitted to the array express repository, ID=E-MTAB-3485.
Analysis of chemokine production
A scratch assay was performed and the cells treated with or without lysate as described above. At 12 h or 18 h after scratching, the spent cell culture fluid yielded from the plates was removed. The CXCL2 level was determined by ELISA (Invitrogen, Life Technologies Ltd, Paisley, UK), in adherence to the manufacturer’s instructions. The optical density of wells was determined using a Labtek LT400 microplate reader (Labtech International Ltd, UK) set to read absorbance at 450 nm. The experiment was carried out on three separate occasions with triplicate samples within each individual experiment.
SDS-PAGE and Immunoblotting
This was performed as described previously8 using primary antibodies CXCR2/IL-8 RB (2.5 μg in1 ml of PBS) overnight at 4 °C. Membranes were washed for 3 × 5 minutes with TBS/Tween containing 5% skimmed milk solution and then incubated with secondary antibody (HRP-conjugated goat anti-mouse IgG) at 1:5000 dilutions for 1h at room temperature. Then the immunoblotting assay was performed as described in8.
Statistical analyses
All experiments were performed three times with triplicate samples within each individual experiment. For experiments comparing two or more treatments, a two-way ANOVA with post hoc Tukey test were utilised to analyse the main effects of and interactions between, multiple factors. Results were considered significant if P ≤ 0.05 and all analysis were performed using the SPSS (IBM SPSS Statistics version 16.0) program.
Additional Information
How to cite this article: Mohammedsaeed, W. et al. Lactobacillus rhamnosus GG Lysate Increases Re-Epithelialization of Keratinocyte Scratch Assays by Promoting Migration. Sci. Rep. 5, 16147; doi: 10.1038/srep16147 (2015).
Change history
25 January 2024
A Correction to this paper has been published: https://doi.org/10.1038/s41598-024-52533-2
References
- Coconnier, M. H. et al. Inhibition of adhesion of enteroinvasive pathogens to human intestinal Caco-2 cells by Lactobacillus acidophilus strain LB decreases bacterial invasion. FEMS Microbiol letters. 110, 299–306 (1993).
Article CAS Google Scholar - Anderson, R. et al. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 10, 305–316 (2010).
Article Google Scholar - Servin, A. L. & Coconnier, M. H. Adhesion of probiotic strains to the intestinal mucosa and interaction with pathogens. Best Pract Res Clinic Gastroenterol. 17, 741–54 (2003).
Article CAS Google Scholar - Reid, G. 2001. Probiotic agents to protect the urogenital tract against infection. Am J Clinic Nutr. 73, 437–443 (2001).
Article Google Scholar - Haukioja, A., Loimaranta, V. & Tenovuo, J. Probiotic bacteria affect the composition of salivary pellicle and streptococcal adhesion in vitro. Oral Microbiol and Immuno. 23, 336–343 (2008).
Article CAS Google Scholar - Mohammedsaeed, W., McBain, A. J., Cruickshank, S. & O’Neill, C. A. Lactobacillus rhamnosus GG inhibits the toxic Effects of Staphylococcus aureus on epidermal keratinocytes. Appl Environ Microbiol. 80, 1–14 (2014).
Article Google Scholar - Prince, T., McBain, A. J. & O’Neill, C. A. Lactobacillus reuteri protects epidermal keratinocytes from _Staphylococcus aureus_-induced cell death by competitive exclusion. Appl Environ Microbiol. 78, 5119–26 (2012).
Article ADS CAS PubMed PubMed Central Google Scholar - Sultana, R., McBain, A. J. & O’Neill, C. A. Strain-dependent augmentation of tight-junction barrier function in human primary epidermal keratinocytes by Lactobacillus and Bifidobacterium lysates. Appl Environ Microbiol. 79, 4887–94 (2013).
Article ADS CAS PubMed PubMed Central Google Scholar - Matsui, T. & Amagai, M. Dissecting the formation, structure and barrier function of the stratum corneum. Inter Immunol. 27(6), 269–280 (2015).
Article CAS Google Scholar - Alibardi, L. Structural and Immunocytochemical characterisation of keratinisation in vertebrate epidermis and epidermal derivatives. Inter Review of Cytol. 253, 177–259 (2006).
Article CAS Google Scholar - Santoro, M. M. & Gaudino, G. Cellular and molecular facets of keratinocyte re-epithelization during wound healing. Exp Cell Res. 304, 274–86 (2005).
Article CAS PubMed Google Scholar - Bjarnsholt, T., Kirketerp-Møller, K. & Jensen, P. Ø. Why chronic wounds will not heal: a novel hypothesis. Wound Repair Regen. 16, 2–10 (2008).
Article PubMed Google Scholar - Yates, C. C., Hebda, P. & Wells, A. Skin wound healing and scarring: fetal wounds and regenerative restitution. Birth Defects Res C Embryo Today. 96, 325–33 (2012).
Article CAS PubMed PubMed Central Google Scholar - Bowler, P. G. & Duerden, B. I. Wound microbiology and associated approaches to wound management. Clinic Microbiol Review. 14, 244–269 (2001).
Article CAS Google Scholar - Halper, J., Leshin, L. S., Lewis, S. J. & Li, W. I. Wound healing and angiogenic properties of supernatants from Lactobacillus cultures. Exp Biol Med (Maywood). 228, 1329–37 (2003).
Article CAS PubMed Google Scholar - Lam, E. K. Y., Yu, L. & Wong, H. P. S. Probiotic Lactobacillus rhamnosus GG enhances gastric ulcer healing in rats. Eur J Pharm. 565, 171–9 (2007).
Article CAS Google Scholar - Jones, M., Ganopolsky, J. G. & Labbé, A. Novel nitric oxide producing probiotic wound healing patch: preparation and in vivo analysis in a New Zealand white rabbit model of ischaemic and infected wounds. Inter Wound J. 9(3), 330–43 (2012).
Article Google Scholar - Peral, M. C., Rachid, M. M., Gobbato, N. M., Martinez, M. A. H. & Valdez, J. C. Interleukin-8 production by polymorphonuclear leukocytes from patients with chronic infected leg ulcers treated with Lactobacillus plantarum. Clinic Microbiol and Infect. 16, 281–286 (2009).
Article Google Scholar - Valdéz, J. C., Peral, M. C., Rachid, M., Santana, M. & Perdigón, G. Interference of Lactobacillus plantarum with Pseudomonas aeruginosa in vitro and in infected burns: the potential use of probiotics in wound treatment. Clinic Microbiol and Infect. 11, 472–9 (2005).
Article Google Scholar - Peral, M. C., Martinez, M. A. H. & Valdez, J. C. Bacteriotherapy with Lactobacillus plantarum in burns. Inter Wound J. 6, 73–81 (2009).
Article Google Scholar - Bolstad, B. M. 1., Irizarry, R. A. & Astrand M. A comparison of normalization methods for high de nsity oligonucleotide array data based on variance and bias. Bioinformatics. 2, 185–93 (2003).
Article Google Scholar - Werner. S. & Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol Review. 3, 835–870 (2003).
Article Google Scholar - Lampugnani, M. G. Cell migration into a wounded area in vitro. Meth Molecular Biol. 96, 177–82 (1999).
CAS Google Scholar - Reyzelman, A. M. Human acellular dermal wound matrix for treatment of DFU: literature review and analysis. J Wound Care. 24, 128–34 (2015).
Article CAS PubMed Google Scholar - Trivedi, U. et al. Prevalence of multiple antibiotic resistant infections in diabetic versus nondiabetic wounds. J Pathol. 17, 30–53 (2014).
Google Scholar - Daniela, P., Marzani, B. & Minervini, F. Plantaricin A synthesized by Lactobacillus plantarum induces in vitro proliferation and migration of human keratinocytes and increases the expression of TGF-β1, FGF7, VEGF-A and IL-8 genes. Peptides. 32, 1815–24 (2011).
Article Google Scholar - Zlotnik, A., Yoshie, O. & Nomiyama, H. The chemokine and chemokine receptor superfamilies and their molecular evolution. Genome Biol. 20, 7–243 (2006).
Google Scholar - Devalaraja, R. M., Nanney, L. B., Yu, Y. & Richmond, A. Delayed Wound Healing in CXCR2 Knockout Mice. J Invest Dermatol. 115, 234–244 (2000).
Article CAS PubMed PubMed Central Google Scholar - Werner, S. & Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol Review. 3, 835–870 (2003).
Article Google Scholar - Rennekampff, H. O. et al. Bioactive interleukin-8 is expressed in wounds and enhances wound healing. J Surg Res. 93, 41–54 (2000).
Article CAS PubMed Google Scholar - Yan, F. et al. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterol. 132, 562–75 (2007).
Article ADS CAS Google Scholar - Yarrow, J. C., Perlman, Z. E., Westwood, N. J. & Mitchison, T. J. A high-throughput cell migration assay using scratch wound healing, a comparison of image-based readout methods. BMC Biotechnol. 4, 21 (2004).
Article PubMed PubMed Central Google Scholar - Yan, F. & Polk, D. B. Characterization of a probiotic-derived soluble protein which reveals a mechanism of preventive and treatment effects of probiotics on intestinal inflammatory diseases. Gut Microbes. 3, 25–28 (2011).
Article Google Scholar
Acknowledgements
Walaa Mohammedsaeed was sponsored by Taibah university and was supported by a Scholarship from Ministry of Higher Education of Saudi Arabia.
Author information
Authors and Affiliations
- Institute of Inflammation and Repair, The University of Manchester, Manchester, UK
Walaa Mohammedsaeed & Catherine A. O’Neill - Faculty of Life Sciences, The University of Manchester, Manchester, UK
Sheena Cruickshank - Manchester Pharmacy School, The University of Manchester, Manchester, UK
Andrew J. McBain
Authors
- Walaa Mohammedsaeed
You can also search for this author inPubMed Google Scholar - Sheena Cruickshank
You can also search for this author inPubMed Google Scholar - Andrew J. McBain
You can also search for this author inPubMed Google Scholar - Catherine A. O’Neill
You can also search for this author inPubMed Google Scholar
Contributions
W.M. performed the studies and wrote the first draft of the paper. S.C. advised on the scratch assay. A.J.M. advised on the microbiology. C.A.O. supervised the studies. All authors edited the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article
Cite this article
Mohammedsaeed, W., Cruickshank, S., McBain, A. et al. Lactobacillus rhamnosus GG Lysate Increases Re-Epithelialization of Keratinocyte Scratch Assays by Promoting Migration.Sci Rep 5, 16147 (2015). https://doi.org/10.1038/srep16147
- Received: 03 July 2015
- Accepted: 08 October 2015
- Published: 05 November 2015
- DOI: https://doi.org/10.1038/srep16147