■ (original) (raw)
2024 年8月6日投稿
DNA methylation-edited mouse
前回紹介したCpG-free cassetteをpromoter近傍のCGIsに挿入する技術を使って、target genesのCGIsのmethylationを誘導したESCsを作った。図1に方法を示した。左から、target geneのCGIsで、CpGはmethyl化されていない(〇)。矢印にしたがって、CpG-free cassetteが挿入された状態になった。すると、methyl化されていなかったsitesがmethyl化された(●)状態に変化する。さらに矢印で示したのは、CpG-free cassetteを外しても、methyl化の状態は変化しない。
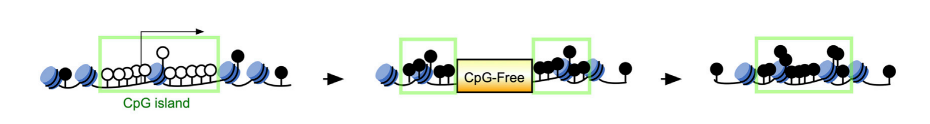
図1.文献17より引用
Target genesとして、Ankyrin repeat domain 26 (Ankrd26)およびlow-density lipoprotein receptor (Ldlr)、それぞれのpromotersにassociatedしているCGIsを選んだ。Ankrd26_のknockout mouseはobesityを発症するが、生存率や生殖に影響を与えない。また、_Ldlr_のknockoutはhypercholesterolemiaを発症することが、知られている。これらのtarget CGIsはactive status markersのH3K4me3とH3K27acの修飾を有する。Targeting vectorsは図2(Ankrd26のケース)のような構成で、CpG-free DNAの両側にPiggyBac inverted repeats (ITRs)を付加した(cassetteが挿入されると、境界のTTCTがTTAAになる)。mESCsに、targeting vectorsをCas9 nucleaseを発現させるplasmidとa single guide RNAを一緒にtransfectした。G418選択によって、homologous recombination (HR) clonesを作製した。_Ankrd26 CGI targetingの場合、取得した8 HR clonesの内、4 clonesでde novo DNA methylationが起こっていた。特に、2 HR clonesでは、CpGの55%がmethyl化されていた。Ldlr CGI targetingの場合、de novo DNA methylationを示す2 HR clonesを取得した。それらのHR clonesから、excision-only PiggyBac transposaseによって、CpG-free cassetteを切り出し除いた(HRex)。さらに、whole-genome bisulfite sequencing (WGBS)によって、targetした領域にのみ、methylationが誘起されていることを確認した。そして、Ankrd26 HRex cloneの一つでは、Ankrd26 mRNAの発現がWTの10%にまで減少していた。CPIのmethylationおよび_Ankrd26_ mRNA合成の抑制は、HRexを25 passages培養しても変化しなかった。
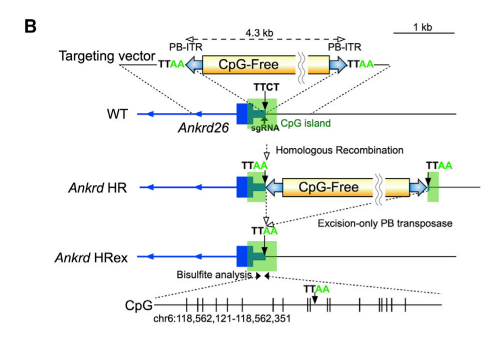
図2.文献17より引用。cassetteが組み込まれる様子
Targeted DNA methylation-edited mice の作製
様々なDNA methylation-edited ESCsをwhite coat colorのマウスの8-cell stage embryosにmicroinjectした(キメラを確認するために白マウスを使った)。Embryoをsurrogate mother miceに移した(図3)。Ankrd HR1exをinjectして作った♂マウス(Ankrd c3)は身体全体がagouti coat colorを示し、高度なchimeraが形成されたことを示した。さらに、chimeric miceのsomatic cellsの_Andrd26_ CGIのCpGsの90%がmethyl化されていた。この結果は、mESCsの中で、Ankrd26 CGIのmethyl化は、発生の過程で保持されたことを意味する。また、Ankrd c3マウスのsomatic cellsで_Ankrd26_ mRNA発現が低下していた。
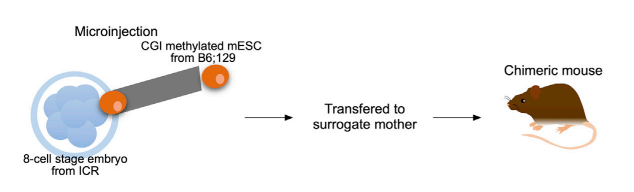
図3:文献17より引用
Ankrd26 knockout mouseがobestyなどの症状を示すことが報告されている。作製した_Ankrd26_ chimerias(離乳後10か月)はWT miceより体重が重く、knockout mouseと同様の表現形質を示した。一方、平行して作製した、low methylated clone Ankrd26 HR7exから作製した、Ankrd26 CGIのmethylationの程度の低いchimeric mouseでは、obestyのような症状は認められなかった。
同じ手法で、Ldlr HR1exをmaicroinjectionして、chimeric miceを作製した。
Ankrd26 CGI のDNA methylationのtransgenerational inheritance
CGIに誘起されたmethylationが次世代に伝わるかを調べた。♂Ankrd c3を♀ICR mouseと交配した。Ankrd c3親と同様に、TTAAalleleのF1 miceでは_Ankrd26_ CGIsはほぼ完全にmethyl化されていた(95%)。実験結果は図4に示してあるが、説明は細かくなるので省略。ただ、♂が□で、♀が〇。TTAA allele (homologous recombinationしたallele)では、CGIがmethyl化されていること、を見て取れる Low methylated chimera Ankrd HR7c1親にして作ったF1 miceでは、Ankrd26 CGIの約25%がmethyl化されていた。これらの結果は、親miceの_Ankrd26_ CGIsのmethylationはそのままF1 miceに伝達されたこと示している。
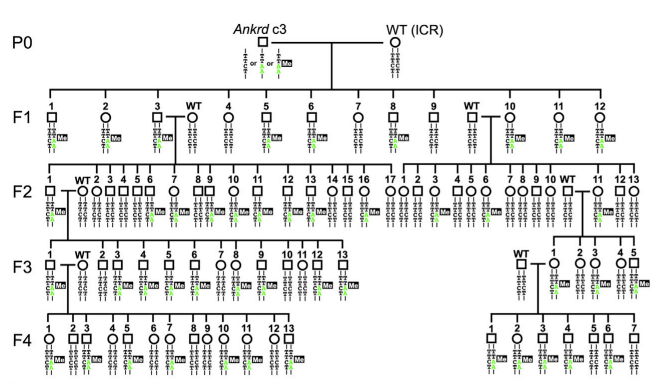
図4.文献17より引用
さらに、♂F1 mouseと♀ICR mouseを交配して作製したF2 miceでもまったく同じようにCGIsはmethyl化されていた。少なくとも、F4 generationまでは、methylationの状態は維持されていた。これまでは、♂のAnkrd c3を使った結果であるが、♀Ankrd c3を♂ICRと交配しても、同じような結果が得られた。すなわち、Ankrd26 CGIに誘起したmethylationは、♂および♀の生殖細胞を介して、子孫に伝達されることが示された。また、メチル化のパターンが配偶子から、体細胞に正確に伝達されることも明らかとなった。
Ankrd26 CGI methylation 次世代への伝達は表現形質の伝達をともなう
Heterozygous methylated F1 male (F1-1)とheterozygous methylated F1 female (F1-2)を交配した(図5)。次に、homozygous F2の♀ (F2-6)とhomozygous F2の♂ (F2-8)を交配し、F3を作った。Ankrd26 CGIのmethylationを調べたところ、methylation stateは安定に子孫に伝達された(図6)。Chromatin statusがどうなっているかを調べた。H3K4me3はactive promoterの特徴であり、H3K9me3はgene silencingの特徴である。ChiP-qPCRによって解析した結果が、図7である。Ankrd26_の発現が抑制されているF2およびF3ではH3K9me3のChipに多くの_Ankrd26 DNAが含まれており、逆にH3K4me3のChipでは少なくなっている。_Ankrd26_の発現が多いWTでは、逆の傾向がみられた。さらに、F2 and F3 homozygous methylated miceでは、obese phenotypesが認められた。Serum leptin levelsも高かった。
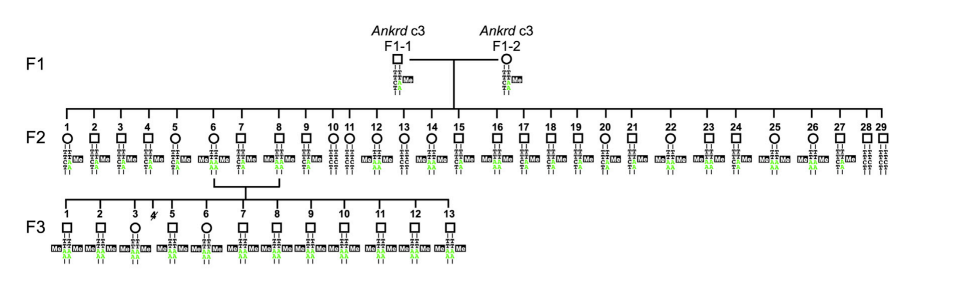
図5.文献17より引用。
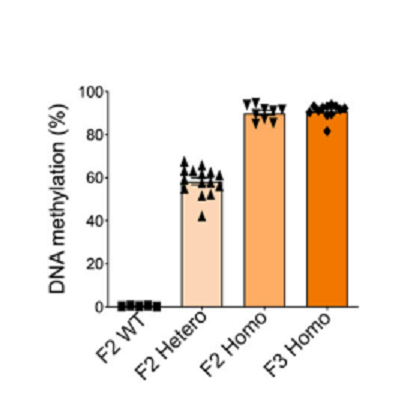
図6.文献17より引用
同様な実験を_Ldlr_ CGI methylationについても行っているが、データの紹介は省略する。ただ、F2およびF3 ATAA homoであっても、60%程度のCGI methylation-positiveであるところが、Ankrd26 CGIのケース(図6)と違っている(図7)。
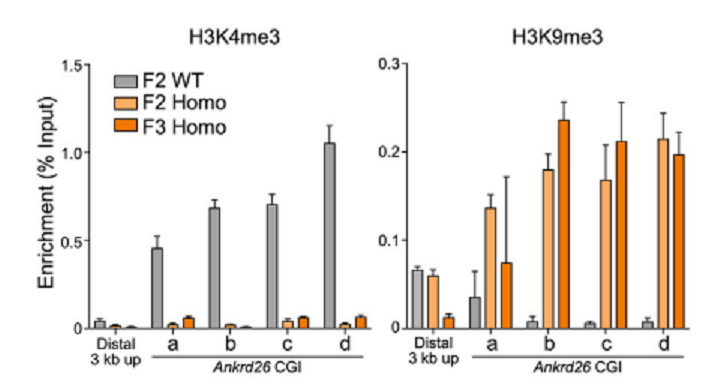
図7.文献17より引用
DNA methylation of CGI は生殖細胞系列でreprogramingを受け、embriosで再構築される
図8に示したように、胚が発生して体細胞の分化が進むと、DNA methylationは不可逆的に進行する(太い青線)。生殖細胞系列では、E13.5の時期にDNA methylationは消去され、受精時期を経て、着床後、発生にともないDNA methylationがプログラム通りに再構築される。では、CGIsに誘導されたDNA methylationはどういう経緯をたどって子孫に伝達されるのか、が問題である。まず、Pou5f1(Oct4)-driven GFP ♀mice(PGCsがGFPを発現する)とhomogeneous Ldlr CGI-methylated ♂mice ( Ldlr c1 line)を交配し、E13.5にFACSでGFP陽性のPGCsを分離した。PGCsとsomatic cellsのgenomesをbisulfite sequencing解析を行った。E13.5 embryoのsomatic cellsには、TTAA alleleにおける_Ldlr_ CGI methylationをはっきりと確認できたが、単離したPGCsには_Ldlr_ GCIのmethylationはかすかにしか認められなかった。この結果は、親マウスの_Ldlr_ GCI methylationがPGCsでは受精後E13.5の間に消去されたことを示している。また、F1 miceから単離したspermでは、Ldlr CGI methylationがまったく検出できなかった。同様に、Ldlr CGI-methylated ♀miceからoocytesを調べても、Ldlr CGI methylationは認められなかった。これらのF1 miceから生まれたF2 miceでは、Ldlr CGI methylationが再構築されていた(図8の下側の赤線)。結果は、Ldlr CGI methylationはtransgenerationalにinheritしたことを確認した。
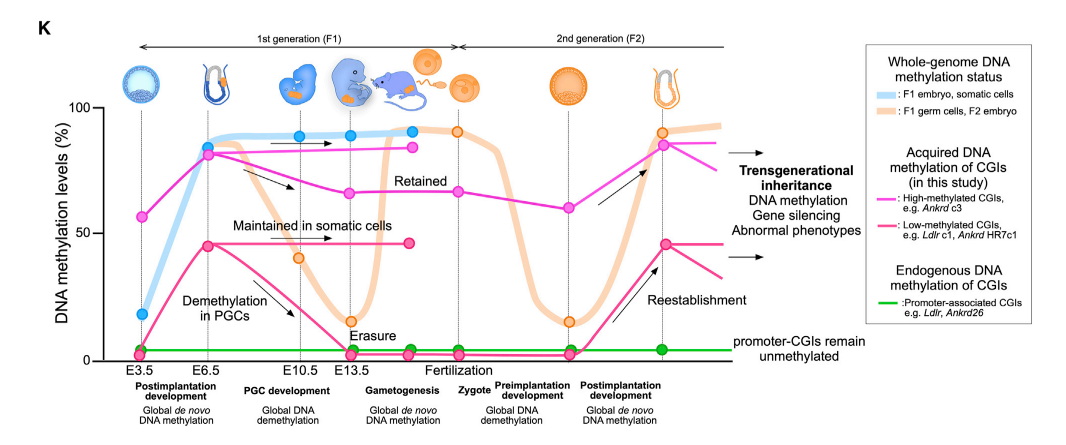
図8。文献17より引用
図8に示したあるように、Ankrd c3 miceのE13.5 PGCsでは、部分的なdemethylationしか起こっていなかった。Ankrd miceでも、somatic cellsの_Ankrd26_ CGI methylationがTTAA alleleで33%の系統では、Ldlr miceと同じように、spermでmethylationが認められなかった。
まとめ
本実験では、Ankrd26 CGIのhypermethylation、および_Andrd26 CGIと_Ldlr CGIのhypomethylationのケースを解析した。前の論文で示したように、CpG-free cassetteをmESCの_Ankrd26_ CGIおよび_Ldlr_ CGIをtargetとして、導入し、それぞれのCGIをmethyl化したHR clonesを得た。仮親に導入して作製したDNA methylation-edited miceから、安定してF1などの子孫にCGI methylationの状態が伝わった。CGI methylationの程度によって、target promoterが支配するgenes (Ankrd26 and Ldlr)の発現が抑制された。Target CGIs methylationはPGCsで減少し、gametogenesisの過程、受精、着床前までdemethylationの状態が続いた。しかし、着床後、以前と同じmethylationの状態に戻った。CGI hypermethylationのケースでは、PGCsにおけるdemethylationは不完全にしか起こらなかった。一方、hypomethylationの場合、元のCGI methylationの状態に戻るためには、methylationについてのmemoryがなければならない。この問題については、実験データはなく、植物でH3K9とnon-coding RNAがde novo DNA methylationを支配している結果などが引用され、議論されているだけである。