GET AWAY TRIKE ! (original) (raw)

素でブログ開設11周年を忘れる程度には忙しい日々が続いているが、そうはいっても隙を見てやるべきことは色々とある(メックウォリア5Cとか)。お盆はわりあいにのんびりできた格好で、出稼ぎ先の人々からそそのかされたりなんだりでバンダイスピリッツは1/32のイマジナリースケルトンにようやく(発売から2年を経て)手を出したのであった。ここ最近で再販がかかっているわけではないようだが、とはいえ店頭在庫はときおり見かけるし、最近では博物館のショップに並んでいることもある。
今さら書くまでもなく筆者の「いちばんすきなきょうりゅう」はトリケラトプスである。従って、トリケラトプスを標榜するなにがしかの出来のよいアイテム(できれば「化石」に近しいものがいい)を手元に置いておきたいという欲はだいぶあるのだが、一方で化石をあれこれして骨格図をこさえて日銭を稼いでいる身分としては、生体復元模型――「復元のビジュアル化」の極致であり、ある種もっとも化石からかけ離れたものである(トリケラトプスの皮膚痕は恐竜の中でも相当な面積が知られているとはいえ)――のできあいのものに「満足」することは遠ざかって久しく、自分でどうこうする気も遠くなって久しい。
しかしながら(出来のよい)骨格復元模型であれば、自分の脳内で都度都度そこへ軟組織のレイヤーを重ねることができる。組み立てに多少の問題があっても、骨の形状さえ正確であれば、切り刻んで「ぼくのかんがえたさいきょうのふくげんこっかく」として組み立て直すことは造作もない。というか、「ぼくのかんがえたさいきょうのふくげんこっかく」を(脳内にせよその外にせよ)こねくり回すのは、筆者のようなタイプにとっては恐竜の模型のもっとも楽しい遊び方のひとつである。
そんなことはずいぶん前からわかっていたわけだが、様々な点において、トリケラトプスのマスプロダクティブな骨格模型は不遇と言ってよい(ティラノサウルスの骨格模型が初手でマスターピースに肉迫しているにもかかわらず)。ハイエンドかつマスプロダクトな骨格模型としてはフェバリットのものがあるが、現行(と言ってもずいぶん昔の発売で、生産は終了しているようだ)のバージョンも厳しさにあふれており、材質上改造も困難である。近年では博物館が公開している復元骨格の3Dモデルが出力・販売されていることもあるが、そもそも既存のトリケラトプスの復元骨格は大なり小なり不適切なアーティファクトを抱えており(あるいは変形がひどく)、その縮小3Dプリント品を単に組み直しただけでは「ぼくのかんがえたさいきょうのふくげんこっかく」には程遠いのである。
既存の骨格模型、あるいはその再マウントだけでは欲しいものが手に入らないことがいい加減わかった以上、手に入れるべきものは改造しやすい(単なる切り貼りだけでなく、盛る・削るといった作業にも供しやすい)ものである。この点イマジナリースケルトンのトリケラトプスは(このところの市場を鑑みれば)手頃な価格で入手もしやすく、なによりプラモデルであるため改造しやすい。結果的にセミスクラッチといえる状態になったとしても、キットというとっかかりがある時点でゼロベースからフルスクラッチするよりははるかに楽(既存のものの気に入らない部分をいじればいいだけなのだ)で、かつ一定の完成度も担保できるだろう。
「イマジナリースケルトン」のシリーズ名に筆者は泡を吹いた(結果、発売から2年にわたってキットを手に取ることはなかった)わけだが、キットの箱には「”化石”という太古の記憶から“生”を想像する」という景気のいい文句が書かれている。つまり組み上がったキットから生きているトリケラトプスの姿を想像してね♡ということらしいのだが、しかしまず「我々」がすべきことは化石から骨格そのものの復元を行うことであり、そしてこのキットはトリケラトプスの化石の1/32スケールモデルとしてみるには不可解な点があまりに多い。本キットから「ぼくのかんがえたさいきょうのふくげんこっかく」――それは本来、実際の復元骨格に課された制約から解き放たれているプラモデルが目指す場所でもあるはずだ――を完成させるためにはそれなりに大規模な改造が必要になるようだが、とはいえ先述の通りキットベースで作業できるという気楽さはある。
若き日の筆者とて無からカラウルやゼロスをこしらえたわけではない。イントロが1000字をぶっちぎったが、そういうわけで筆者の製作記録について書いておく。
―――――――――――――――――――――――――――――――――――――――
1. 概観
一度キットを組み上げたら全体をよく観察し、要所要所で計測しておく。キットの(長さ方向に関する)プロポーションは実のところトリケラトプスの大型成体(として筆者が制作した骨格図)にかなり近く、
・頭蓋がでかい
・胴椎が1個多い
・肩帯が細長い
・第IV指、第V指が小さい
・腸骨がでかい
・大腿骨が短い
・中足骨が長い
点を除けば特に問題ないようだ。
本キット最大の問題は、胴体の骨格が生物として成立しない状態でマウントされている(トリケラトプスの胴体がほぼ可動しないだろうという点はさておいても、肋骨の背側に恥骨が乗っており、スタンド攻撃を受けたかのような状態である)ところにある。また、肋骨の形状そのものがそもそもおかしい(これは事実上あらゆるマスプロダクトのトリケラトプスの骨格縮小模型にいえる)うえ、胴椎の横突起と肋骨の骨頭がまったく関節していない(謎の溝が生じている)のもこの状況を悪化させているといえよう。頭骨の造形のまずさ(説明書では_T._ ホリドゥスであることを示唆しているが、T. プロルススめいた何かにしか見えない)はこの際おまけでしかないし、四肢の組み立てのおかしさはもはや微笑ましい。

↑長骨はそれほど手を加えなくていいことがわかり一安心する。頭蓋と肋骨さえいじれば見違えるだろう。
逆に言えば、このあたりの問題さえ解消できれば本キットはトリケラトプスの縮小骨格模型としては他に類を見ないレベルの代物に化けるはずである。改造方針が定まったところで作業に移る。必要な道具は、
・瞬間接着剤各種(中粘度、ゼリー状、パテタイプのもの)、スプレー式硬化促進剤
・エポキシパテ(切削性に優れたタイプのもの)
・ニッパー、デザインナイフ(どちらもなるべくよく切れるもの)、スパチュラ
・紙ヤスリ(筆者はだいたい320番を使用)
・棒ヤスリ(先端のカーブしたダイヤモンドヤスリを使用)
そして、様々な図版をコラージュしてキット(というよりこれから作ろうとする模型)の実寸サイズでプリントした図面である。トリケラトプス・ホリドゥスの(化石としては事実上ほぼ知られていないクラスの)大型個体を作るとして、参照とする標本は主に
・YPM 1820(ホロタイプ):顔面
・ゼンケンベルク自然史博物館の頭蓋(標本番号不詳):フリル
・USNM 4842(“ハッチャー”):骨格の主要部位
・NSM PV 20379(“レイモンド”):“ハッチャー”の欠損部
・HMNS PV.1506(“レイン”):同上
・BSP 1964 I 458(トリケラトプス・プロルスス):同上
をあたることとした(他に下顎や肩帯は別の標本を参考としている)。“ハッチャー”や“レイモンド”、BSP 1964 I 458はそれぞれ記載があり、非常によい参考となる。一方でこうした標本では肋骨の詳細な記載がないため、スティラコサウルスの記載も参考とする。
2. 形状加工
2-1. 仙椎・腰帯
腸骨と恥骨は肋骨をマウントする上で重要な基準点となるため、何よりもまず手を付けるべきは骨盤まわりである(ただし、パテを盛るのは肋骨ができあがってからでよい)。仙椎(第10仙椎は尻尾のパーツB20に割り振られている)と腰帯は素晴らしい立体感だが、腸骨が異様に大きく肉厚で、第10仙椎も長すぎる。第10仙椎はB20から切り離したうえで前方の関節面を削って短くし、第9仙椎に接着する。腸骨は天面のパーツ(C19、C20)は使用せず、ブレード全体を削り込んでパテを盛り、形を整える。恥骨と座骨の形状はそのままでよいが、大腿骨の骨頭をねじこめるよう、寛骨臼は(本来ある骨の薄い裏打ちは無視して)削り込んで開口しておく。
骨盤と仙椎の関節位置はキットでは妙なことになっているため、左右それぞれの骨盤の工作が終わったら、骨盤がより前方・腹側で関節するよう仙椎に接着する。左右の座骨の嵌合部は削り取り、遠位端を瞬間接着剤で強引に接着してしまうとよい。第1~第5仙椎の棘突起もパテなどで一体化させておくこと。
2-2. 仙前椎と肋骨
「躍動的に」大きく曲がった胴体はおそらくこのキットのセールスポイントであり、きちんとした形のトリケラトプスの骨格模型が欲しいのであればこれを完全に殺す必要がある。胴体と胴肋骨のブロック(実際にはいくつかの頸椎と頚肋骨も含まれている)は一度完全に組み上げ、きっちり接着しておく。胴椎はなぜか1個多いので、最終胴椎をまるごとカットする(肋骨は細く加工したうえで第1仙胴肋骨として仙椎と腸骨に接着する)。
接着した胴体ブロックは椎骨ごとに切り分け(ひとつひとつ切り分けるのは大変なので、要所で切断するのがよい)、胴体ブロックが真っすぐになるよう接着する。続いて肋骨をすべて切り離し(それぞれの肋骨に番号を書きこんでおくこと)、胴椎の横突起ときちんと関節するよう注意しながら接着する。
本キットの肋骨はケラトプス科の特徴的な形態とはまるで一致せず、個別の肋骨の形態をきちんと表現するにはすべて自作する必要がある(それ以前に胴椎の横突起のつくりからして正確ではない)。そもそもトリケラトプスの肋骨がきちんと記載・図示されたことはなく、参考にできる復元骨格も“レイン”程度である(“レイン”にしても肋骨は少なからず変形しており、またマウントにも多少なりとも問題がある)という問題がある。
あくまでキットベースの改造に留めるならば、肋骨の近位部の形状が不正確にしかならない、肋骨籠前方の長さが足りなくなるといった問題はあるが、キットの肋骨の近位部をカットし、スティラコサウルスの記載を参考に手で肋骨を曲げていく(案外折れないし脆化もしない)というのが現実的な落としどころだろう。個々の肋骨の形態ではなく、肋骨籠全体の(その中でも側面の)アウトラインを追い込んでいくことを目標に作業を進めていく。
腸骨・恥骨を頼りに第12胴肋骨を接着し、そこを起点により前方の肋骨を接着していく。肋骨の「肩」の描くラインが腸骨ときれいにつながるよう意識するとスムーズである。頸椎ブロックはそれほどいじる必要もないが、後方の頚肋骨は胴体ブロックとラインがきれいにつながるよう加工する。

↑肋骨と椎骨の関節まわりもきちんと作りこみたいところだが、見ての通り本キットの胴椎はそもそも横突起の形状がだいぶ不正確であり、手を入れるならば本来そこから作り直すべきである。そこまで手を入れ始めると確実に完成しなくなる(それはそれで豊かな経験ではあるはずだが、そこまでいくとキットベースの改造という意義が失われていく)ため、そのあたりは潔くスルーし、肋骨の骨頭は(どうせ瞬間接着剤まみれになるため)切りっぱなしのまま済ませてしまうことにした。頚肋骨の改造は、肩帯のフィッティングが終わってから、そこに合わせて行うとよいだろう。

↑胴椎と左側の胴肋骨を再マウントしたところ。かはくの“レイモンド”(下の写真)でも、こうした肋骨と恥骨の位置関係を見て取ることができる。

2-3. 尾
血道弓が意味不明な場所に関節させられていたり、神経弓の前方/後方関節突起が省略されたうえ妙な処理をされているほかは全体的によくできている。血道弓の関節位置の調整は、よほどのことがなければ避けた方が無難だろう(筆者もしれっと無視することとした)。関節突起が省略された結果生じた隙間は瞬間接着剤で埋めるとよい。筆者は第1尾椎をやや切り詰めたほか、血道弓の個数や形状、横突起の形状、遠位尾椎の数を修正した(付属の解説書には尾椎の数について言及があるが、厳密な個数がはっきりしている角竜はわずかであり、ケラトプス科内でもそれなりに変異があるようだ)。

↑ここまでの産物をキットの素組と比較する。“化石”という太古の記憶から“生”を想像する」のであれば、こうした作業は本来避けては通れない。なにより、こうした作業は「古生物学者ごっこ」のおいしい部分である。
2-4. 肩帯
肩甲烏口骨・胸骨は大がかりな改造が必要である。サポート用のパーツC5は使用せず、肋軟骨(付録の解説書にはなぜか腹肋骨と書かれている)は初めに切除しておく。
肩甲烏口骨は肩甲骨・烏口骨の関節で切断し、肩甲骨はそこから長さを切り詰める。肩甲骨ブレードを走る稜は完全に削り落とし、肩甲骨の高さを増したうえで新造する。烏口骨は一回り小さくなるようカットし、加工した肩甲骨に接着する。そのうえで肩関節窩を新造する(ポールらによる写真と図がよい参考になる)。胸骨は一回り小さくカットし、左右を揃えて接着しておく(AMNH 971やセントロサウルスの記載がよい参考になるだろう)。
肩甲骨烏口骨の加工が完了したら、完成した肋骨籠に接着する。肋骨籠の上方から胴体全体を吊り下げるイメージで、かつ胸骨とうまく関節する位置を探る(“レイン”の復元骨格をはじめ、肋骨籠の下方から胴体を支えるように組み立てられた肩帯はしばしばみられるが、これは恐らく完全な誤りである)。肩甲烏口骨を接着したら、さらに胸骨も接着してしまう。

↑肩帯のフィッティングに際しては四肢と同時並行で行うべきでもあろうが、そうなると実作業としてはあまりにも混沌とするので、まずは肩帯の位置を仮決めする。胴体前半部の肋骨のつくりが(改造後も)あまりよろしくないことを考えれば、実際には(肩関節の位置はこのままで)肩甲骨ブレードの後半部はもっとおさまりがよくなるだろう。(骨化しない)間鎖骨や鎖骨(少なくともプロトケラトプス科段階までは骨化する)の存在を考慮すると、左右の烏口骨にはある程度の間隔があるはずだ。
2-5. 四肢
手と足を除けば四肢はよくできており、加工は最低限で済む。各骨を関節で切り分け、嵌合にかかわる部分はすべて切り落としておく。
上腕骨は内側上顆がおかしな位置にあるので削り落とし、より内側の位置に新造する。この時、三角筋稜や外側上顆もパテでボリュームアップさせておく。前腕と関節させる際に重要なため、遠位部の関節形状は注意して造形すること。
橈骨・尺骨はともにボリューム不足だが、形状そのものはよくできている(ただし橈骨は左右逆のようだ)。橈骨は左右を入れ替え、尺骨にきちんと関節させてやる。橈骨・尺骨はパテでボリュームアップさせること。
手は中足骨に沿って切り分け、前腕部に接着する。第IV指・第V指は新造する。筆者は省略したが、腕に覚えのある人は手根骨も作るとよい。第I指は指骨がひとつ多いので関節部を削ってそれっぽくごまかす。末節骨のサイズがおかしなことになっているので削って調整する。
大腿骨は骨頭を作り直し、内側顆をボリュームアップさせる。中央部で切断し、延長する。
脛骨は遠位端の内側部をボリュームアップさせる。距骨は前方側が嵌合の都合で大きくえぐれているのできちんと復元しておくこと。
足は中足骨が異様に長く、また趾もわずかに長いようである。趾の長さには目をつぶることとし(末節骨は削っておく)、中足骨の短縮で足のプロポーションを改善させることとした。中足骨に沿って切り分け、セントロサウルスなどの足や足跡化石の図を参考に、指が放射状に広がるようにして再接着する。

↑関節で切り分けて骨頭にパテを持ったりしているとはいえ、四肢の改造は背骨まわりと比べればはるかに楽である。キットの腰帯や四肢は明らかに“ハッチャー”の記載に基づき造型されているが、にもかかわらず妙な部分で不正確でもある。
2-6. 頭骨
頭蓋は下顎に対してやや大きく、全体として非常にきびしい造形である。後頭顆周辺は位置取りがおかしなことになっているが、嵌合の都合で本来の後頭顆-環椎関節とボールジョイントの凹凸が逆になった結果、頭骨を椎骨に繋いで吻を下げている分にはむしろ問題ない。説明書を無視してA9、A13、A1、A4を接着し、フリル(A13)は鱗状骨の近位部を残してすべて切除する。また、A9は下顎を固定するピンを削り込んでおくと、下顎が可動するようになる。前上顎骨~頬骨(A1・A4)はほぼ消失するまで削り込んでおき、前上顎骨の先端側はA13ごと切り詰めておく。
上眼窩角はキットパーツ(A2・A3)を使用する場合、左右逆にするとアウトラインをそのまま活かすことができる。老齢個体ではどうも相対的に角があまり長くない(大型亜成体の頃が相対的に最も長い角を持っている)ようなので、このあたりは自分が作るものと相談である。

↑パテを盛る都合上、頭蓋はこのくらいまで削り込んでおく必要がある。筆者はトリケラトプス・ホリドゥスとして製作したが、トリケラトプス・プロルススを製作する場合でも同様である。
こうして加工した頭蓋にエポキシパテを盛りつけ、整形していく。筆者はYPM 1820とゼンケンベルグの頭骨(プロルスス風の吻を据えられているが、鱗状骨の形態は明らかにホリドゥス型である)をコンポジットにしたイメージで改造し、頭蓋にあわせて前歯骨もパテで延長した。キットのスケール表記通りの大型個体として制作するならば、縫合線は特段彫りこむ必要はない。筆者は前上顎骨のもろもろの構造は潔く省略したが、腕に覚えがあるなら再現しておくべきところでもある。キットの構造上、下側頭窓の開口はリスキーだろう。
下顎の相対的なサイズ・プロポーションはやけに正確で、形状もほぼ問題ない。上角骨の外側面に突出する部分を少し削り、背側・腹側から見た際の歯骨のくびれを軽く削って強調してやれば十分である。

↑細部を作り込むほどの腕はないので、とにかくできる範囲でシルエット・プロポーションを追い込んでいく。キットの表記通り1/32スケールで換算するとYPM 1828もびっくりの大型個体ということになるので、若者らしくは見えないように作ることとした。
3. マウント
すでに胴体のポージングはキットとは別物である。キットでは左肘が胸郭にめりこみかけているので、そのようなことがないよう注意して四肢を接着していく。接着しては剥がし、を繰り返すつもりで進めていくとよいだろう。肩や肘・股や膝関節は(軟骨を介するとはいえ)骨頭の位置関係に注意しなくてはならない。

↑はじめに体軸骨格に皮膚に見立てたマスキングテープを貼り、内臓の入っている領域を可視化しておく。そのうえで、骨格だけでなくそこに付着するはずの筋肉がマスキングテープを圧迫しない姿勢で、かつ足跡化石とよく一致するポージングを探っていく。肩関節の姿勢はこの写真の撮影後にもう一度修正した。
こうしてすり合わせていくと、突拍子のない姿勢を取らずとも、体化石・生痕化石双方と矛盾することはないようだ。先述の通り肋骨は(肋骨籠全体のアウトラインはともかく)実際の形態とはやや異なっているが、そこを勘案しても特に影響はないようである。


↑なにしろエポキシパテによる顔面再建手術のあとなので絶妙に自立せず、ひっつき虫で適当に立たせることとした。生々しい改造の跡(ある種アーティファクトである)を残しておくのも全然よいのだが、それはそれとして最初からこの形だったら(少なくとも、各要素がこの形状をしていれば)どんなに嬉しかったかというところの気持ちを込めて塗装することとした。
4. 塗装
塗装は全体の作業からすればほんのおまけだが、ここまでに挙げた強引な作業の形跡を隠し、さもスマートな作業の連続だったふうに見せかける役割がある。どんな色で塗装してもよい(あえて化石ではなく「骨」風に塗ってしまうのも一興だろう)が、YPM 1820や“ハッチャー”、“レイン”といったランス層産の標本を改造のインスパイア元としたこともあり、それっぽい色で塗ってやりたいところでもある。ランス層産の化石といえば黄土色や白っ茶けた色の印象が強いが、標本によってはヘル・クリーク層産のものと同様のきれいなチョコレート色を呈するものもある。YPM 1820に至っては(恐らく)風化の進んでいなかった部位がチョコレート色、風化して崩れかけていた部分が褐色から黄土色、というように変化に富んでいる。このあたりの折衷として、褐色と黄土色の入り混じったような色をランス層産の化石風ということにして塗装することとした。
改造で生じたキズや段差を塗膜が埋めてくれることを期待して、ダークアースを筆で厚めに塗り、乾燥後にサンディブラウン→セールカラーの順に雑に(下塗りのダークアースがだいぶ隠れる程度まで)ドライブラシする。仕上げにシェイドブラウンでウォッシングし、つや消しクリアでコートしてやれば完成である。




↑頭骨の細かい部分を容赦なく省略したり、肋骨の完全再現をあっさり諦めた結果肋骨基部のカーブがおかしかったり胴肋骨の長さが全体的に足りていなかったりもするが、とはいえプロポーションは基本的に骨格図と同じである。同じプロポーション、形状の骨を並べたものであっても、実際の骨格を真横から(なるべくパースを排除して)眺めるのとポール式骨格図では少なからず見た目に違いが表れる。ポール式骨格図に肉付けしても生体復元画にはなり得ないのだ。
―――――――――――――――――――――――――――――――――――――――
かくして丸4日でトリケラトプスの製作は終わった。1/32というキットのスケール表記通りにいけば、全長(キットからほぼ変化なし)およそ8.8m、頭蓋長2.5mにして大腿骨長も1.3m強と、既知の最大級の標本と同等かそれ以上のサイズの個体という計算になる。スケール換算はさておくとしても、自分の知っていること・考えたこと・できることのせめぎ合いから生まれただけあって満足感は相当にある(どこをどういじればより完成度が高くなるかを知っている、というところも含め)。決して楽ではないが、とはいえ手頃なサイズで出来のよいトリケラトプスの縮小復元骨格模型――「化石のスケールモデル」を手にする道は確かにあるのだ。
筆者が原書の第一報を目にしたのは、振り返ってみれば3年ほど前のことであった。どういうわけか発売日はその時点で確定しており(現にその通りだったように思う)、ちょっと眉をひそめた覚えもある。
そんなわけで原書を購入した筆者であったが、実のところ一通り斜め読みしたきり、ほとんど単なるインテリアとして本棚の最下段に置くにまかせていた。より正確に言えば、前半の「翼竜概説」は(どの程度骨格図が出ているかを確認した程度で)斜め読みさえしていなかったのである。原書の発売前に取り留めもない話をしたためたりもしていた(これは筆者の本の制作が佳境に入っていた時期で、自らを鼓舞しようとした側面も大きい)のだが、つまり――少なくとも斜め読みする限りでは――原書は筆者の期待を裏切ることも上回ることもなかった。その後に発売された海竜フィールドガイドの食味にすべてを上塗りされたふうでさえあったのである。
恐竜フィールドガイドの第二版はかくして邦訳され、日本語で読めるもっとも網羅的な恐竜図鑑としての地位を保っている。であれば、翼竜フィールドガイド――海外でさえ網羅的な書籍は数えるほどしかなく、ウィットンによる教科書がこれを除くと現代的なものとしては唯一であろう――が邦訳されるのは既定路線だったのかもしれない。なによりもまず、現代的な翼竜の、それも非常に網羅的な図鑑が邦訳されたことを喜ぶべきである(ヴェルンホーファーの訳書である平凡社『動物大百科 別巻 翼竜』が色あせることはないが、なにしろ原書は1991年の刊で、現代的というよりは近代的という位置付けになる)。そして翻訳について何ら心配をする必要がないのは大変に頼もしい限りである。原書に一人で立ち向かうより格段に“安全”であるのだ。
そうは言いつつ、筆者は一匹狼気取りである。一人でどうとでも相手はできるとタカをくくり、あげく先述の通り斜め読みすら満足にしていなかった。そうこうしているうちに出版社(&訳者)のご厚意で、前回と同様に邦訳版をいただいてしまったわけである。かくして、筆者はやっとこさ腹を括って『グレゴリー・ポール 翼竜事典』ときちんと向き合うこととした。実際問題として、福井県立大学恐竜学研究所と福井県立恐竜博物館の援護射撃があればこそ、(『グレゴリー・ポール 恐竜事典』と比べればだいぶボリュームは小さいが)本書を相手取ることができるようになった格好である。これは本書を手に取るであろう日本語話者の方には例外なく当てはまるはずだ。
『グレゴリー・ポール 恐竜事典』の出版によって日本国内でポールの復権が起きたと思いたい筆者だが、果たしてそのあたりはどうなのだろう。年若い読者にとっては、(自覚的に)初めて触れるグレゴリー・ポールの復元図がおそらく『グレゴリー・ポール 恐竜事典』であることには違いない(『恐竜図鑑展』でポールの骨格図が事実上まったく扱われていなかったことを思えばなおさらである)。筆者くらいの歳であれば00年代初頭までにポールの描く恐竜の骨格図には確実に触れているはずだが、そうした世代であっても、ポールの描く翼竜の骨格図はほとんど未知の存在である。なにしろ、本書に載っている翼竜の相当な数――ポールの言葉を借りれば“有効な種の約1/3が今世紀に入ってから命名されている”のだ。ウィットンの教科書(繰り返しになるが、邦訳はない)で翼竜の分類の現代的な知見をまざまざと見せつけられた筆者だが、あらためてそれを「ポール式」の骨格図で見るのはさすがに壮観である。翼竜の現代的かつ網羅的な日本語の図鑑が本書だけであることを鑑みれば、本書が日本における恐竜マニア(あえてそう言い切ってしまおう)の翼竜に対する興味を相当に深化させることは間違いない。であれば、『グレゴリー・ポール 翼竜事典』の出版――The Princeton Field Guide to Pterosaursの邦訳は諸手を上げて歓迎すべきである。相当にやんちゃな著者の本だとしても、だ。
当然のごとく、原書で半分以上を占めていた前半パートの「翼竜概説」は、日本語版たる本書でも同様のウェイトを占めている(本書は単なる翼竜骨格図集ではないのだ)。幸いというべきか、前作『グレゴリー・ポール 恐竜事典』のように訳者による脚注の嵐が吹き荒れることはなく、先行研究のレビューという意味では割合におとなしくまとまっているようだ。翼竜の地上での姿勢の復元について熱弁を奮うあたりは往年の輝きを感じさせるものでもあるが、一方でそれを(「ポール式」の)骨格図で扱うことについてはいささかの自己矛盾をはらんでいるようにも聞こえる。
前半で抑え気味だったポール節は後半の骨格図パートで爆発する――が、前作と比べてやはり控えめのようにも思われるのは、筆者が翼竜に疎いせいか、翼竜の分類が(特に成長に伴う形態の変化と関連して)いまだ混沌としているせいなのか、それともデイビッド・ピーターズ――ウィットンと並んで翼竜の骨格図ではよく知られてしまっている――の影が脳裏にちらつくからなのだろうか。翼竜の骨格図は同じくポールの手による恐竜の骨格よりも相当に模式的に描かれているものが多いが、これはすなわち参照できる資料の質、つまり化石の保存状態やそれらを詳細に記載した文献の有無に影響されているようだ。骨格図から滲み出すものも含め、本書の抱えている膨大な情報をどこまでモノにできるかは前作と同様やはり読者に委ねられている。「ケツァルコアトルスの未命名種」のように原書の出版直後に記載されたものもあり、本書の先には広大な一次資料の海が広がっているのだ。本書を繰り返し手に取るたび、読者は自らの成長を実感していくはずである。このあたりの付き合い方は、前作について書いた時と何ら変わることはない。
繰り返しになるが、本書『グレゴリー・ポール 翼竜事典』は翼竜の図鑑としてはもっとも網羅的な書籍の邦訳版であり、翼竜に関する現代的な本としてもやはり日本語で読めるものとしては唯一無二である。やんちゃで知られた著者の本とはいえ、本棚に加える理由としてはそれで十二分にすぎるだろう。前作『グレゴリー・ポール 恐竜事典』に福井県立大学恐竜学研究所・福井県立恐竜博物館の訳者たちと共に立ち向かった読者であれば、臆することはないはずだ。訳者の援護射撃の下、本書をモノにできるかは読者次第。今一度、存分に腕を振るう時である。
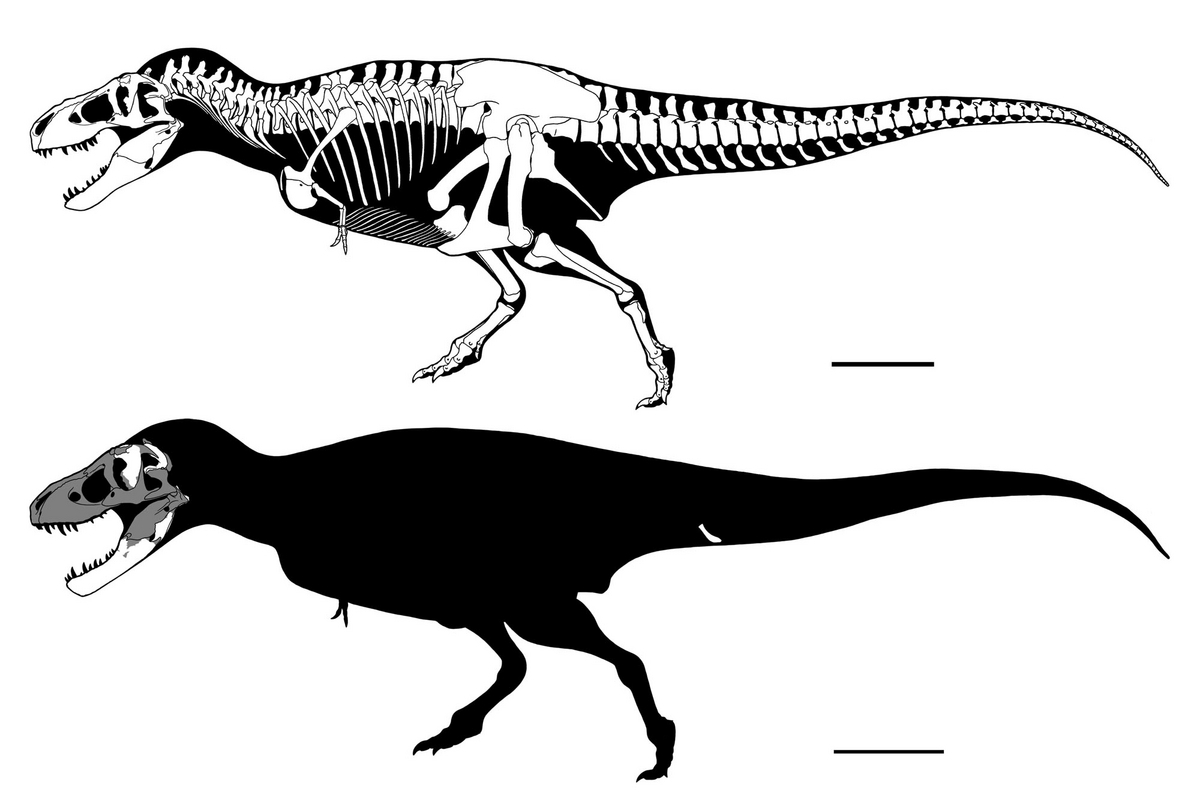
↑Skeletal reconstruction of Tyrannosaurus rex (top: largely based on USNM PAL 555000)
and Tyrannosaurus mcraeensis (bottom: holotype NMMNH P-3698). Scale bars are 1m.
いい加減で3月である。年明け早々にでも書こうと思っていた記事は、あけおめメールの到来によってここまで後ろ倒しになった格好である。
さて、かれこれ10年近く前にこんな記事を書いたりしていたわけだが、年明け早々に“エレファント・ビュートのティラノサウルス類”ことNMMNH P-3698(古い文献だとNMMNH P-1013-1となっている)がティラノサウルス属の新種として記載されたわけである。長らく「ニューメキシコ産ティラノサウルス・レックス」として知られていた本標本は、紆余曲折の末にひとまずティラノサウルス・レックスとは別物と見なされるようになったのだ。
ニューメキシコの上部白亜系といえば北西部はサン・フアン堆積盆のもの(フルーツランドFruitland層やその上位のカートランドKirtland層、オホ・アラモOjo Alamo層)が有名だが、中西部のマクレーMcRae層群(最近までマクレー層扱いだった)でも20世紀の初頭から恐竜化石が産出することが知られていた。1980年代に入ると調査が活発化したが、採集された化石は単離した要素ばかりで、おまけにひどく風化したものばかりであった。
とはいえ、それなりに同定のできそうなものはないわけではなかった。トロサウルスかペンタケラトプスと思しき大型角竜(頭頂骨に窓を持つのは確かだった)に大型の竜脚類――この時代のアメリカ南西部の竜脚類といえばアラモサウルスを置いて他にはない――、それに鎧竜の皮骨のスパイクが採集されたが、大本命と言えるのがエレファント・ビュート貯水池の東岸――ホール・レイクHall Lake層(当時は部層扱いだった)で1983年に採集された巨大な獣脚類の歯骨だった。この歯骨NMMNH P-3698(この時点ではNMMNH P-1013-1のナンバリングだった)はほぼ完全かつ保存もなかなかのもので、しかも前関節骨や血道弓といった他の要素も一緒に採集されたのである。歯骨は紛れもないティラノサウルス科のそれで、しかももろもろの特徴は何よりもティラノサウルス・レックスとよく一致するものだった。かくして、ジレットらはこれを嬉々としてニューメキシコ初となるティラノサウルス・レックスとして記載したのである。
1993年に同じホール・レイク層の下部でトロサウルスのかなり大きな部分骨格が採集されたこともあり、マクレー層群の恐竜相は典型的なアラモサウルス相――“ランス期”(≒マーストリヒチアン後期)のアメリカ南西部に存在した、アラモサウルスやトロサウルス、ティラノサウルスを特徴とした恐竜相であるとみなされるようになった。が、2017年になってホセ・クリークJose Creek層(マクレー層群の最下部層である)の上部やホール・レイク層の最下部付近の火山灰層からカンパニアン後期を示す絶対年代が報告されたことで話は妙な方向へと動き始めた。「掘り残し」を得てみれば、ホール・レイク層下部から産出した「トロサウルスの部分骨格」はトロサウルスなどではなかったのである。
さて、当初ティラノサウルス・レックスとして記載されたNMMNH P-3698だったが、当時イケイケだったレーマンとカーペンターは1990年に「ニューメキシコ産アウブリソドンの部分骨格」(今日ビスタヒエヴェルソルとみられている)を記載した際にこれが新属新種にであることを(カーペンターの印刷中だった論文を引用する形で)示唆した(が、出版された論文ではテキサスはハヴェリナJavelina層産の上顎骨TMM 41436-1しか言及されていなかった;後述)。当然これといった理由は述べられていなかったわけで、カーとウィリアムソンは2000年のニューメキシコ産ティラノサウルス類のレビューでこれを一蹴したわけである。当時知られていた目ぼしいティラノサウルス・レックスの(大人サイズの)標本を精査したうえで、カーらは改めてNMMNH P-3698をティラノサウルス・レックスとみなしたのだった。
この見方はその後も踏襲され、カーによる近年のティラノサウルス・レックスの成長過程に関する大著ではNMMNH P-3698はやや若い成体とみなされた。カーはNMMNH P-3698が(2000年とは比べ物にならないほど比較できる標本が増えた中にあって)ティラノサウルス・レックスであることを改めて示した格好でもあったのだが、一方でマクレー層群の最近の時代論については特に注意を払うことはなかったのである。
1986年にNMMNH P-3698を記載するにあたり、ジレットらは「掘り残し」がまだ現地にあるらしいこと、エレファント・ビュート貯水池の水位が下がって再び現地を訪れられるようになるのは今後2年では厳しいだろうという見通しについても述べていた。果たして、「掘り残し」はまだあったのである。
「トロサウルスの部分骨格」改めシエラケラトプスNMMNH P-76870の記載に勢いづき、追加要素も含めたNMMNH P-3698の再記載が進められた。マクレー層群の絶対年代を報告した論文中ではNMMNH P-76870にせよNMMNH P-3698にせよ(ホール・レイク層における)産出層準が不詳であることが述べられてはいたのだが、改めて行われた地質調査で両者の産出層準がホール・レイク層の最下部付近――7320万±70万年前(カンパニアン後期の後半)――からそれほど上位にあるわけでもないことが判明した。絶対年代による「挟み撃ち」はできなかったが、とはいえ推定される堆積速度からしてNMMNH P-76870にせよNMMNH P-3698にせよ、カンパニアン末からせいぜいマーストリヒチアン前期の前半のものとみてよさそうだったのである。
かくしてティラノサウルス属の新種――ティラノサウルス・マクレーエンシスのホロタイプとなったNMMNH P-3698だが、とはいえティラノサウルス・レックスとの形態的差異として挙げられている特徴のほとんどはかなり微妙であり、歯骨の特徴(腹側縁が角骨との関節部に至るまで背側にカーブする)を除けば成長過程や個体変異、化石の破損・変形によるものとも言えてしまうようなものばかりである(このあたり筆頭著者の悪名高さについてはいちいち書かない;カーが同サイズ個体についても成長段階による差異らしきものを拾い出していることはよく顧みるべきである)。とはいえ歯骨の特徴はティラノサウルス科全体を見渡しても他に見られないものであり、であればひとまず別種と見ておいた方がよいのだろう。

↑北米産のティラノサウルス族の頭骨。上段と下段でそれぞれ、左から右へ向かって成長段階が進んでいく。カーによってTMM 41436-1は亜成体、それ以外は成体とされている。“スタン”とUMNH VP 11000では眼窩周辺の皮骨を透過して描いており、それ以外の標本では省略している点に注意。スケールバーは50cm。
さて、筆頭著者のダールマンといえば“アラモティラヌス”で悪名高いわけである。かつて本ブログでも取り上げた通り、どうもオホ・アラモ層(これまでの時代論は決定打に欠けるが、恐竜化石の産出するナアショイビトNaashoibito部層はおそらくC31rすなわちマーストリヒチアン前期~後期初頭(ざっと7140万~6930万年前)に収まるようだ)産のティラノサウルス類――ほとんどが単離した首から後ろの骨格であり、部分骨格に至ってはカーらのレビューで触れられこそすれど図示されない程度の代物だった――を“Alamotyrannus brinkmani”として記載するつもりだったようなのだが、「印刷中」(一般論として査読が終わっていることを意味する)として引用された論文はとうとう出版されず、今日ではアメリカ南西部のティラノサウルス族の俗称のようなものとして用いられる始末となっている。
(ダールマンが“アラモティラヌス”として触れた唯一の標本がACM 7975である。これは保存のぱっとしない部分的な右歯骨であるようだが、今日まで特に記載されておらず、その実態は定かではない。ダールマンがこれをホロタイプにしようとしていたのかも謎である。)
NMMNH P-3698と並んでアメリカ南西部のティラノサウルス・レックスとしてよく知られていたのがテキサスはハヴェリナ層最上部(ケツァルコアトルス・ノースロッピのホロタイプの産出層準と実質的に同じである;もろもろを勘案するとマーストリヒチアン後期の中ごろのようだ)から産出した部分的な上顎骨TMM 41436-1である。これはローソンの修士論文の中で“ティラノサウルス・ヴァヌスTyrannosaurus vanus”として記載されたが、その後1976年に改めてティラノサウルス・レックスとして記載されたものであった。カーペンターは先述の論文の中でこの標本が他の(ティラノサウルス・レックスとされている)上顎骨に見られる個体変異から外れていることを指摘し、未知の属であることを示唆した(が、追加標本の産出を待つと記したのみそれ以上は特に何もしなかった)。実のところこの上顎骨はティラノサウルス・レックスの成体と比べるとかなり小さく、もっぱらティラノサウルス・レックスの亜成体である(≒なにかしらの別種ないし別属にできるような根拠が特にない)とみなされている。
もうひとつ、ユタ州はノース・ホーンNorth Horn層(マーストリヒチアン後期からダニアンにかかっているが、恐竜化石が産出する層準はマーストリヒチアン後期初頭あたりに相当するようだ)ではティラノサウルス・レックスとされる部分骨格UMNH 11000が産出している。これはかなり風化の進んだ骨格だが、後眼窩骨(およびそこに付着する皮骨)と鱗状骨が関節して保存されており、アメリカ南西部産のものとしては非常に貴重である。
オホ・アラモ層産の標本はさておくとしても、ハヴェリナ層やノース・ホーン層産の標本の分類についてはどういうわけかティラノサウルス・マクレーエンシスの記載論文中では触れられておらず、UMNH 11000がティラノサウルス・レックスの一例として比較用に(SIの中で)図示されている程度である。一方で、実のところUMNH 11000とNMMNH P-3698は後眼窩骨の特徴を共有しており(UMNH 11000の後眼窩骨は皮骨の付着した状態で図示されているが、皮骨を外した状態だとNMMNH P-3698と同様の形態を示す)、もしこの特徴が本当に(筆者は先述の通り歯骨以外の独自性については懐疑的であるが)ティラノサウルス・マクレーエンシス特有のものであるならば、UMNH 11000もティラノサウルス・マクレーエンシスと言えそうである。TMM 41436-1の正体は判然としない(なにしろ同様の形態の上顎骨が他に知られていない)が、あるいはこれもティラノサウルス・レックスというよりはティラノサウルス・マクレーエンシスの亜成体なのかもしれない。
NMMNH P-3698がカンパニアン末~マーストリヒチアン前期の前半のものであることはほぼ確実であり、一方で「トリケラトプス相」――コロラド以北から産出したティラノサウルス・レックス(と断言できるもの)で最古のものはMOR 1125や“スー”といったヘル・クリーク層の最下部付近から産出したもの――どうやってもマーストリヒチアン後期の前半には遡らない――である。両者の間には少なくとも380万年に渡るギャップが存在する格好だが、先述の通りUMNH 11000やTMM 41436-1はマーストリヒチアン後期の前半のものであるらしく、であればこのギャップを埋める存在ということになる。マーストリヒチアン前期から後期の前半にかけてのアメリカ西部の恐竜相はカナダと比べても謎だらけだが、鍵はとうに手の中にある。

↑マイプの合成骨格図(上)と不定のメガラプトル類の合成骨格図(下)。
上は恐竜博2023用に制作したもの、下は2014年6月に制作したもの。基本的に同じ標本を組み合わせて描いており、実質的にプロポーションの違いがない点に注意。
暇をもてあました末に、それなりの下心をもって本ブログを立ち上げて10年が過ぎた。暇だったのは最初の1年だけで(学部2年の後期から3年の前期という、必修科目も少なければ卒業研究もまともに始まっていない稀有な時期であった)、ここ数年は毎年新機動戦記ガンダムWもびっくりな急展開が続いているところである。10年ともなればさすがにあっという間に過ぎたというわけもなく、振り返ればそれなりに長く、ほとんど消えかかった獣道が藪の中に伸びている。
さて、筆者が骨格図を制作する仕事を請け負うようになって7年ほどになるが、その中でしばしば描いてきたのが合成骨格図――同一種の複数標本はおろか、複数種の部分的な骨格さえ寄せ集めたコンポジットである。筆者が初めて本格的に描いたコンポジットの骨格図は2014年に描いたメガラプトル類のもので、以来同じテーマでたびたび個人的に描くことがあった。どっこい、ひょんなことから因縁の相手(筆者とて茨城うまれなので適当に因縁を付けがちである)――最大最新のメガラプトル類であるマイプの骨格図を描くという機会を得、ひとまず(この手の話は常に“ひとまず”止まりだ)の決着をつけるに至った格好である。10年目にして決着の機会を与えてくれたマイプについて、今さらながらここに書き記しておきたい。
パタゴニアといえば恐竜化石の世界的な大産地としてよく知られている。広大なパタゴニアに大小さまざまな堆積盆が広がっている格好だが、白亜紀後期の獣脚類の良好な記録はもっぱら最北部のネウケンNeuquen堆積盆(ネウケン州とその周辺)とその南に位置するカナドン-アスファルトCañadón Asfalto堆積盆(チュブ州とリオ・ネグロ州)に限られてきた。パタゴニア南部――南米大陸の南端に広がるマガヤネスMagallanes/オーストラルAustral 堆積盆ではオルコラプトル_Orkoraptor_やアウストロケイルス_Austrocheirus_を除いて命名されたものはなく、オルコラプトルにせよアウストロケイルスにせよ分類さえおぼつかないほどに断片的な標本に基づいていたのである。
(オルコラプトル、アウストロケイルスとも原記載では「下部マーストリヒチアンのパリ・アイケPari Aike層」産とされている。このあたりの層序と年代論についてはつい最近まで著しい混乱があり、オルコラプトルについてはその後「セノマニアン~サントニアンのマタ・アマリヤMata Amarilla層」に、アウストロケイルスについては「上部チューロニアン~下部コニアシアンのパリ・アイケ層」に修正された。最終的にオルコラプトル、アウストロケイルスとも「中部カンパニアン~下部マーストリヒチアンのセロ・フォルタレザCerro Fortaleza層」産とされるようになり、今日に至っている。これはプエルタサウルスやドレッドノートゥス、タレンカウエンと同じ地層から産出したことを意味している。これらの恐竜の生息時代についてより踏み込んだことは言えないのが現状だが、ざっくりカンパニアン後期としておくのが妥当なところだろう。アウストロケイルスはケラトサウルス類とみて間違いないようで、わずかに残された化石を見るにエラフロサウルスやデルタドロメウスが連想される。)
一連のセロ・フォルタレザ層産恐竜が発掘されたのは2000年代に入ってからだが、マガヤネス/アウストラル堆積盆で恐竜化石が発見されたのはそれが初めてのことではなかった。チョリーヨChorrillo層(上部カンパニアン~下部マーストリヒチアン)では1940年代までにはすでに恐竜の化石が発見されていたのである。1980年には竜脚類の部分骨格が発見され、駆け付けたボナパルテによって部分的な頸椎(だけ)が採集された。骨格の残りの部分はそのまま現地に残されたが、ボナパルテは同じ産地から獣脚類の尺骨や歯冠をも採集していた。
竜脚類の頸椎はボナパルテによってcf. アンタークトサウルス sp.とされ、その後アエオロサウルス類とされたり不定のティタノサウルス類とされたりもしたが、依然として骨格の残りは現地に残されたままだった。この標本に再びスポットが当たったのは2019年の1月――ノヴァス率いる“ベルナルディーノ・リヴァダヴィア”アルゼンチン国立自然科学博物館のチームがこの地を訪れてからのことである。
発掘隊はボナパルテが頸椎を採集した産地を再特定し、首尾よく骨格の残りを採集した。この骨格(ボナパルテが採集した頸椎MACN-PV 18644および掘り残しMPM 21542)は新属新種の大型ティタノサウルス類(おそらくコロッソサウルス類)ヌロティタン・グラキアリス_Nullotitan glaciaris_のホロタイプとなったが、発掘隊はヌロティタンの追加標本に加え、他にも新属新種のエラスマリア類イサシクルソル・サンタクルセンシス_Isasicursor santacrucensis_といった様々な恐竜化石を採集していった。その中にはチョリーヨ層中部の下部から産出したMPM 21545――ひどく断片的だが単一個体に由来すると思しき大型獣脚類の骨格があったが、これはメガラプトル類、それもアエロステオンと同等のサイズのものだったのである。
実のところ、チョリーヨ層からメガラプトル類の化石が発見されたのはこれが最初ではなかった。1981年にボナパルテが採集していた獣脚類の歯MACN-PV 19066(ヌロティタンのホロタイプと同じ産地で発見されたが、厳密に共産したかについては記録がない。チョリーヨ層下部の産出であることは確かである)もメガラプトル類のもので間違いないことが判明したのである。さらに推定全長2mほどの個体の胴椎(MPM 21546;MPM 21545より10mほど下の層準からの産出)まで発見され、どうもチョリーヨ層の主だった大型獣脚類がメガラプトル類らしいことが明らかになった。まごうことなきマーストリヒチアンのメガラプトル類が、パタゴニアの南部に存在したのだ。2019年のうちにチョリーヨ層産の化石はまとめて記載され、マガヤネス/アウストラル堆積盆――南極そしてオーストラリアへと続くパタゴニア南部のマーストリヒチアンの生物相の一端が初めて示されたのであった。
(上述したオルコラプトルはさておくとして、チョリーヨ層産のもののほかにマーストリヒチアンの可能性があるメガラプトル類が記載されていないわけでもない。マガヤネス/アウストラル堆積盆の北東に位置するゴルフォ・サン・ホルヘGolfo San Jorge堆積盆のラゴ・コルウエ・ウアピLago Colhué Huapí層(セケルノサウルスの産出が知られる)ではほぼ完全なメガラプトル類の“シックル・クロー”や中足骨が産出しているのだ。ラゴ・コルウエ・ウアピ層はカンパニアンからマーストリヒチアンにまたがっており、セケルノサウルスの産出層準はマーストリヒチアンに相当するとされている。メガラプトル類の化石はいずれもセケルノサウルスのホロタイプよりも下の層準から産出しているが、これがカンパニアンなのかマーストリヒチアンなのかははっきりしていないようである。いずれにせよカンパニアン以降と非常に新しい時代のものであることは確実で、にもかかわらず“シックル・クロー”の形態はメガラプトルのそれと酷似している。)
2020年の3月になり、“ベルナルディーノ・リヴァダヴィア”の発掘隊がチョリーヨ層の一連の産地を抱えたラ・アニタ・ファームへ帰ってきた。今度は国立科学博物館も加えた国際チームである。チームはチョリーヨ層からさらなる化石を採集したが、その中には前年に掘り残していたMPM 21545の続きも含まれていた。
母岩は硬く、車で直接乗り入れることのできない現場ということもあった発掘は難航したが、それでも2020年の発掘で採集されたMPM 21545のパーツの量は前年をはるかに上回った。

↑マイプ・マクロトラックス_Maip macrothorax_のホロタイプMPM 21545の骨格図。
恐竜博2023用に描き起こした(2022年8月)もの(上)と原記載の出版直後(2022年4月)に描いたもの(下)。
腰帯の相対的な位置関係の違いに注意。下のものの腹肋骨はごく模式的に描いている。
マーストリヒチアンのメガラプトル類のまとまった骨格に現場は色めき立ったが、コロナ禍という情勢の急転を受け、ラ・アニタ・ファームでの調査は中断される格好となってしまった。アルゼンチン軍のヘリによるジャケットの搬出予定はなくなり、現地に置き去りにせざるを得なくなった標本さえあったのである。MPM 21545にしても、(関節が完全にばらけた状態であったにもかかわらず)5m×3mほどの範囲を掘ったところで発掘は打ち止めとせざるを得なくなった。標本の行く先であった“ベルナルディーノ・リヴァダヴィア”も閉鎖される中で、MPM 21545はプレパレーターの自宅にてクリーニングが進められることになったのだった。
不幸中の幸いというべきかMPM 21545の保存状態は非常によく、断片的な骨格とはいえ仙前椎の要所や相当数の腹肋骨、そしてほぼ完全な烏口骨が採集されていた。重複する椎骨はアエロステオンのホロタイプMCNA-PV-3137よりも一回り大きな動物のものであり、そして烏口骨は(椎骨がMCNA-PV-3137のそれよりも大きいことを差し引いても)妙に巨大でがっしりしたつくりであった。依然として断片的ではあるが、それでも最新かつ最大級のメガラプトル類の確かな骨格がそこにあったのである。
MPM 21545の椎骨の細部には既知のメガラプトル類には見られない特徴がいくつか確認され、固有の形質らしいものもいくらか見出された。烏口骨の形態はメガラプトル類にしてもかなり独特で、MPM 21545が未記載種に属していることは明らかだった。
かくしてMPM 21545をホロタイプとして2022年に正式に記載された(論文の草稿は2021年の年末には公開されていた)マイプだが、「掘り残し」がいまだラ・アニタ・ファームの片隅に埋もれているとみられている。これまでに採集された部位のほとんどは椎骨や肋骨といった体軸骨格や、それに隣接する肩帯や腰帯に限られており、しかもばらけているとはいえそれなりに狭い範囲にまとまって産出した。とはいえ中足骨の破片らしきものも採集されており、であれば四肢の要素も周辺に散乱しているかもしれない。メガラプトル類の標本としては珍しく第2頸椎も産出しており、なにがしかの頭骨要素が残されている可能性もある。

↑マイプの合成骨格図とその参照元。歯骨と上腕骨、大腿骨はアウストラロヴェナトルのホロタイプも参照している。趾骨はバホ・バレアル層産のメガラプトル類に基づく。また、前上顎骨-上顎骨-鼻骨はメガラプトルの幼体MUCPv 595を大まかに参照している。
マイプのホロタイプから既知のメガラプトル類とは異なる様々な特徴が見出されたことは先述した通りだが、とはいえこれまでに採集されたMPM 21545の概形は他のメガラプトル類とよく似ている。従って、既知のメガラプトル類の標本を注意深く組み合わせることで、マイプの骨格をそれらしく復元することができるはずである。メガラプトル類のトレードマークである前肢は今のところマイプではひとかけらも知られていないが、そこへつながる烏口骨はメガラプトル類としては極めつけにごつく、相対的なサイズもアエロステオンやメガラプトルと比べて大きい。マイプと最も近い時代のメガラプトル類であるラゴ・コルウエ・ウアピ層のものが(相対的なサイズは不明だが)メガラプトルと酷似した“シックル・クロー”を保持していたことも考えれば、前肢についてもメガラプトルと同等以上に発達していたとみるべきだろう。
恐竜博2023でマイプの復元骨格が展示されることはなかった(計画されなかったわけではないものの、いかんせん現状では産出部位も少なく、制作したところでほとんどメガラプトルの拡大バージョンになってしまうだろうということもあって見送られたようである)が、かくして筆者はマイプの合成骨格図を仕事として描く機会を得た。南米産の派生的なメガラプトル類の標本を結集してなお足りない部位も少なくないが、とはいえマイプの骨格は上の図からかけ離れたものでもないはずだ。
白亜紀末のゴンドワナ――南半球の陸上生物相のうち、多少なりとも実態が知られているのはパタゴニア北部やインド、マダガスカルに限られており、陸成層の残るパタゴニア南部も先述の通り近年ようやくスポットライトが当たった格好である。南極やオーストラリアではこの時代の陸成層が知られておらず、特に後者の陸上生物相はまったくの謎に包まれている。とはいえ南極やオーストラリアはパタゴニア南部と古地理上密接なつながりがあり、そうした意味でもチョリーヨ層や隣接する(側方変化とも言える)チリのドロテアDrotea層(ステゴウロスやゴンコケンの記載でにわかに知名度を得た)の研究は非常に大きな意味を持っている。
かくしてゴンドワナにおける最後の頂点捕食者として鮮烈なデビューを果たしたマイプだが、ここまで散々書いた通り骨格はごく断片的にしか知られていない。チョリーヨ層では先述の通り2タイプのメガラプトル類の歯化石が知られているが、これらとマイプの関係さえ現状では定かではないのである。それでも、マイプのホロタイプはこの時代のパタゴニア南部(や南極、オーストラリア)の大型獣脚類としては最良の標本である。おそらく存在する「掘り残し」やチョリーヨ層やドロテア層のポテンシャルからすれば、こうした問題はいずれ解決されるものとみてよいだろう。2023年の2月にはラ・アニタ・ファームでの発掘が再開され、次なる成果に期待が高まっている。
この10年でメガラプトル類に関する知見は大きく広がったが、それでもなお残された謎は多い。次の10年でマイプは、メガラプトル類の骨格復元はどれほどの修正を要することになるだろうか。今日のものは10年後に「妥当な程度に正確」だったと評せるものになっているだろうか。先のことなどわからないが、筆者は気の向くままに描き続けるだけである。
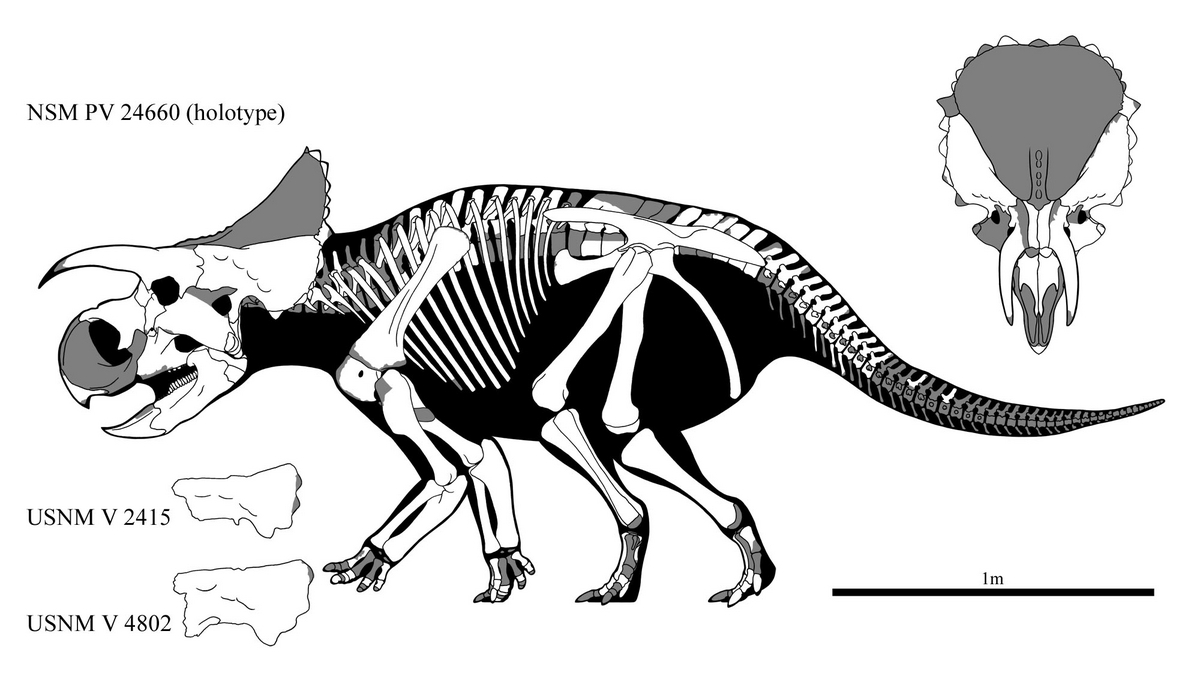
↑Skeletal reconstruction of Furcatoceratops elucidans holotype NSM PV 24660
("AVA", formerly RMDRC 12-020) and some other closely related specimens.
恐竜博2023にてお披露目された国立科学博物館所蔵の「ケラトプス科の未記載種」はナストケラトプス族の新属新種であることが仄めかされており、そもそも本ブログの古参読者のみなさまにとってはずいぶん昔からおなじみの顔でもあった。“AVA”の愛称を与えられたそれはかつて「アヴァケラトプスの新標本」として喧伝されたものだったが、発見から11年を経てついにフルカトケラトプス・エルキダンス_Furcatoceratops elucidans_のホロタイプとして記載されたのである。
本ブログでも散々取り上げてきたジュディス・リバーJudith River層はアメリカ・モンタナ州の北西部に露出しており、デイノドン、トラコドン、トロオドン、パラエオスキンクスと、北米で初めて記載された恐竜たちが産出したことで知られている。ミズーリ川の支流であるジュディス川にちなんで命名されたその地層は19世紀後半から精力的に化石採集が行われてきたが、比較的最近になるまで恐竜のまとまった骨格は知られていなかった。1888年にハッチャーが「最初の角竜」たる“ケラトプス”を発見した(もっと言えば、1876年にコープのチームがモノクロニウスを発見してもいた)地層であるにもかかわらず、1981年に“アヴァケラトプス”のホロタイプANSP 15800が個人コレクターによって発見されるまで、「まとも」な角竜の化石が発見されることはなかったのである。
なにしろ国境を挟んですぐのカナダ側にはダイナソー・パークDinosaur Park層が露出しており、どう考えてもそちらを探したほうが採れ高がよかった。モンタナ州のカンパニアンを見渡しても、西部に露出するトゥー・メディスンTwo Medicine層では1970年代以降マイアサウラをはじめとする恐竜化石の産出が知られるようになり、研究者の目はそちらに向きがちであった。
ケアレス・クリーク・クオリー(様々な動物の入り混じった大規模なボーンベッドであり、近年でもハドロサウルス類の記載がなされるなどしている)における“アヴァケラトプス”(もっぱら疑問名として扱われている)の発見とそれに続く記載は、ジュディス・リバー層を巡る停滞した状況に風穴を開けたようだった。1990年代に入るとモンタナ州のあちこちで民間の発掘業者が活発に活動するようになり、ジュディス・リバー層はマクレラン・フェリーMcClelland Ferry部層の下部で大規模な角竜のボーンベッド(マンスフィールドのボーンベッド)が発見されたのである。
1999年に“アヴァケラトプス”の再記載が出版されたが、このときUSNM 2415とUSNM 4802――ハッチャーによる1888年夏の調査の際、“ケラトプス”のホロタイプUSNM 2411に続いて採集された鱗状骨も“アヴァケラトプス”と同定された。どちらの標本もかつてハッチャーによって採集され、“ケラトプス”名義で図示されたこともある(マーシュはこれを“ケラトプス”の背中に並べようとしていた。“ケラトプス”が剣竜類とされていた時代の話である)、歴史的な標本である。
USNM 2415とUSNM 4802、そして“アヴァケラトプス”のホロタイプを結び付けていたのは、鱗状骨の外側面に走る、顕著な隆起群の存在であった。これほど顕著な構造はケラトプス科を見渡しても“アヴァケラトプス”にしか見られない特徴のように思われたのである。
(暫定的に“アヴァケラトプス”の成体とされた標本MOR 692の鱗状骨にも、同様の顕著な隆起が確認された。今日、こうした構造はナストケラトプス族全般やその他の基盤的なセントロサウルス類でよく発達することが知られている。セントロサウルスのようなより派生的なセントロサウルス類やカスモサウルス類でも同様の構造がみられるが、骨格上ではそれほどはっきりしたものではない。こうした隆起は大きな飾り鱗の起点となっていたとみられており、骨格上で見るよりはるかに目立つ構造をなしていたと思われる。)
その後もジュディス・リバー層における角竜化石の発見は続き、2010年代に入るとメデューサケラトプス(マンスフィールドのボーンベッド産の標本に基づく)、ジュディケラトプス、メルクリケラトプスとジュディス・リバー層産角竜の命名が相次いだ。もっとも、メデューサケラトプスは実態のはっきりしないボーンベッドの産であり(最終的にボーンベッドに含まれている角竜化石は事実上すべてメデューサケラトプスのものらしいことが判明し、頭骨全体を復元可能な状況へと変化した)、ジュディケラトプスはひどく部分的な頭骨の寄せ集め(セントロサウルス類が参照標本の中に紛れ込んでいたことが後に判明した)、メルクリケラトプスは部分的な鱗状骨2点のみと、興味深いながらも扱いに困る代物だらけだったのである。”JUDITH”の愛称で呼ばれていた頭骨の大部分を含む部分骨格はジュディス・リバー層産の角竜としては“アヴァケラトプス”のホロタイプ以来となるまとまった1個体分の骨格であったのだが、商業標本であったことが祟ってかなかなか研究が進まず、ROMによって購入されてからようやくスピクリペウスとして命名される始末だった。
こうした中でトリーボールド社によってジュディス・リバー層はコール・リッジCoal Ridge部層の上部で2012年に発見された標本RMDRC 12-020は、“アヴァケラトプス”と同様、鱗状骨にわりあい目立つ隆起群が走っていた。また、手の「ひづめ状」の末節骨の中に顕著に小さいもの(リンク先では第IVとされているが、言うまでもなく第IIIである)が存在する点も“アヴァケラトプス”のホロタイプと似ているように思われた。かくしてこの標本は“アヴァケラトプス”の新標本として諸手を挙げて歓迎され、“AVA”(そのまんますぎるネーミングである)の愛称で呼ばれることになったのだった。
“AVA”の発掘は翌年のシーズンまで続き、頭骨の相当な部分を含んだ骨格が姿を現した。骨格は完全にばらけていたものの、頭骨の要素は比較的まとまった状態であり、首から後ろについてもほぼ完全な前後肢と肩帯、腰帯、相当数の肋骨が含まれていた。2005年に発見された”JUDITH”をはるかにしのぐ、ジュディス・リバー層の研究が始まって以来となる完全度の角竜の骨格がそこにあったのである。化石化の過程で潰れている骨も少なくなかった一方で、概して骨の表面のテクスチャーはよく保存されており、総じてダイナソー・パーク層でもめったに見つからないような代物であった。
発掘が進むにつれて“AVA”にはMOR 692と同様の長い上眼窩角が確認され、MOR 692が“アヴァケラトプス”の眷属であることにも疑いの余地がなくなった。しかも"AVA"の鼻骨には鼻角が存在せず(ちょっとした隆起があるのみである)、ナストケラトプスのそれとよく似た代物であった。ナストケラトプスは“アヴァケラトプス”とごく近縁である可能性が指摘されていたが、"AVA"はそれを補強するものだったのである。"AVA"の椎骨はまるっきり癒合が進んでおらず、見るからに未成熟個体ではあったが、とはいえ分類上重要な特徴はすでに立ち現れているようでもあった。
“AVA”の復元頭骨は2014年のツーソンショーでお披露目され、復元骨格についても2015年の秋にSVPにあわせて公開された(ついでにハートマンの手による骨格図も発表され、なぜか本ブログの画像がネットの海を漂った末に転載された)。この頃になると“AVA”はもはや“アヴァケラトプス”とは別物とみられるようになっており、トリーボールド社の販売サイトには「ジュディス期の角竜の新種」の文字が躍るようになった。一方で、“AVA”のオリジナルに関する情報はそれきり途絶えてしまった。“アヴァケラトプス”あるいは「ジュディス・リバー層産の新種」として“AVA”の復元頭骨に基づいた生体復元が競うように描かれる一方で、オリジナルの行く末は判然としなかったのである。
(“AVA”の復元頭骨をきちんと見てやればわかることだが、額の部分は左右から圧縮されたような状態でマウントされており、結果的に左右の上眼窩角が正中面でくっつかんばかりとなっている。左右でくっつかんばかりの上眼窩角はしばしば(いまだに)“AVA”の重要な特徴として描かれがちであるが、つまるところこれは化石の保存状態による問題(をマウントの際に矯正しきれなかっただけ)である。後眼窩骨を見れば、“AVA”の上眼窩角は左右ともやや内側(そして腹側)へ向かってカーブしつつ、正中線と平行に伸びていることがわかる。フルカトケラトプスすなわち「フォーク状の角の顔」は、こうした「二又のフォーク状」あるいは音叉状、さすまた状をなしている上眼窩角にちなんだものである。)
さて、2017年になると“AVA”を取り巻く状況に転機が訪れた。国境を挟んですぐのカナダ・アルバータ州はオールドマンOldman層の最上部近くで発見された角竜の断片的な頭骨CMN 8804が属種不定のナストケラトプス族(この時初めて設立された)として記載されたのである。この標本は1937年にチャールズ・モートラム・スターンバーグによって採集されて以来、フィールドでざっくり“ブラキケラトプス”として同定されたきり収蔵庫で眠りについていたものであった。
(CMN 8804に限らず、「オールドマン層上部」産の標本の時代論については少し注意しておく必要がある。オールドマン層とその上のダイナソー・パーク層の境界は南へ行くほど若くなっており、国境付近でみられるオールドマン層の上部は、州立恐竜公園におけるダイナソー・パーク層の下部と同時代ということになる。CMN 8804はつまるところ、州立恐竜公園でいうところのダイナソー・パーク層下部と同じ時代のものである。ちょうどセントロサウルス・アペルトゥスのレンジに重なるようだ。ジュディス・リバー層とベリー・リバーBelly River層群(オールドマン層+ダイナソー・パーク層)の対比については長年議論があったが、最近になって詳細な絶対年代データに基づく見直しが図られており、フォアモストForemost層とオールドマン層の境界がマクレラン・フェリー部層の最下部に、州立恐竜公園におけるオールドマン層-ダイナソー・パーク層の境界がマクレラン・フェリー部層とコール・リッジ部層の境界に対比されている。また、コール・リッジ部層は州立恐竜公園におけるダイナソー・パーク層の下半全体に対比される。もろもろを勘案すると、CMN 8804はMOR 692とおおよそ同時代のものであり、"AVA"よりもやや古いくらいのものであるようだ。)
CMN 8804も未成熟個体ではあり("AVA"よりもやや小さな頭骨であるようだ)、なにしろ断片的ということで命名は差し控えられたが、既知のナストケラトプス族のいずれとも異なる分類群のようでもあった。断片からして上眼窩角はナストケラトプスほど長くはなく、それでいてMOR 692(こちらは相当に成熟したものであるらしい;"AVA"より一回りは大きい)とは異なり、やや内側に向かってカーブしたものであるように思われたのである。
そして、MOR 692を“アヴァケラトプス”とする積極的な根拠がもはやどこにもないことも明白となった。ナストケラトプス族であるということ以上に両者を結び付けるような特徴は見当たらなかったうえ、両者の産出層準は派手に離れていたのである。オールドマン層やダイナソー・パーク層では、ケラトプス科角竜の種がせいぜい数十万年で入れ替わっていくことが知られている。“アヴァケラトプス”が例外的に長命な分類群であった可能性はもちろんあるが、そう考えるべき積極的な理由もまったくない。かくして“アヴァケラトプス”はホロタイプANSP 15800に限定され、MOR 692はANSP 15800とは別物として扱われるようになった。
(このとき“アヴァケラトプス”が疑問名送りを宣言されることは(どういうわけか)なかったが、幼体であるうえに頭蓋天井をごっそり欠いたANSP 15800から独自性らしいものを見出せないという話は繰り返し述べられている。かくして、フルカトケラトプスの記載では“アヴァケラトプス”は容赦なく疑問名として扱われている。)
こうした背景にあって、未成熟個体であるとはいえ頭骨の相当な部分が良好な状態で保存されている"AVA"――頭頂骨をまるっきり欠いているとはいえ、見るからにナストケラトプス族だった――は、このあたりの問題になにがしかのヒントを与えてくれる存在でもありそうだった。なにしろ、ジュディス・リバー層やベリー・リバー層群では文句なしに最良のナストケラトプス族の標本なのである。“AVA”の記載が待ち望まれたが、音沙汰はこの7年間途絶えていた。
とはいえ、こうした状況は(表向きには)急転直下で動くものである。恐竜博2023の展示の概要が明らかになったのは開幕までひと月となった時だったが、そこには紛れもない"AVA"の写真があった。会場には”AVA”――NSM PV 24660が(置けるだけ)並べられており、傍らにはかはくのスペシャル仕様(産出部位とアーティファクトを塗り分けてある)で仕上げられたマウントが佇んでいる。図録にはクローズアップの写真がいくらでも載っており、「ケラトプス科の未記載種」名義の展示だったとはいえ、記載論文の出版が近いらしいことがうかがえた。
(実のところ記載論文は2023年の2月上旬には投稿済みであり、7月20日付で校正前の受理原稿が(印刷中という扱いで)公表された。8月11日付で受理原稿は正式に出版され、晴れて命名と相成った(日本語のプレスリリースが公開されたのは盆明けであった)のである。だいぶ以前から“AVA”の骨格図を制作する機会にあずかっていた筆者だったりもするのだが、最初に描いたものはとうとう一度も表に出ることがなかった。このあたりの話は、そのうちどこかで書くこともあるだろう。)
かくして新属新種として世に送り出されたフルカトケラトプス・エルキダンスのホロタイプNSM PV 24660は(かねてからの指摘通り)未成熟もいいところであった。椎骨の癒合はさっぱり進んでおらず、頭骨も縁後頭骨はおろかほとんどの部分で癒合がみられなかったのである。長骨の薄片に成長停止線はたった2本しか観察されず、(ナストケラトプス族としてはそれなりの体格であるにもかかわらず)生後ものの数年で死んだことは確かなようだ。
未成熟個体ということで分類に細心の注意が必要であることは今さら書くまでもないが、NSM PV 24660では鼻骨と前上顎骨の(腹側の)関節面をはじめ、成長過程で変化しないとみて間違いない独自性がいくつも見いだされた。NSM PV 24660の産出層準はMOR 692と比較的近いとみられているが、両者は明らかな別物であることが確認されたのである。やはり産出層準の近いCMN 8804の上眼窩角は(MOR 692とは異なり)NSM PV 24660と似ているようだったが、あまりにも断片的であるためにそれ以上の比較は困難であった。
(“アヴァケラトプス”ANSP 15800はそっくり鼻骨を欠いており、フルカトケラトプス(上述の通り、鼻骨と前上顎骨の関節が特殊化している)との比較がまともに行えない。とはいえANSP 15800の前上顎骨は(鼻骨との関節面は途中で欠損しているものの)いたって普通のつくりであるらしく、であればフルカトケラトプス的な鼻骨の持ち主ということもなさそうだ。ANSP 15800(カンパニアン中期の中ごろ;ざっと7800万年前ごろ)とNSM PV 24660(カンパニアン後期の前半;7560万年前ごろか)の産出層準のギャップはANSP 15800とMOR 692のそれより大きく、カンパニアンにおけるケラトプス科角竜の生存期間が概して数十万年程度ということからしても、同じ種とは考えにくい。)
NSM PV 24660の頭骨は半ばバラバラになっていた一方で、相当な数の縁後頭骨(頭頂骨の正中線上にあるもの(P0)を除き、事実上すべて発見された計算になる)が採集された。うち、縁鱗状骨(esq)と縁頭頂鱗状骨(eps)については自信をもって元の位置に関節させることのできるものだったが、かくして復元されたフルカトケラトプスの鱗状骨とホーンレットはUSNM 2415や4802――“アヴァケラトプス”が疑問名となって宙に浮いていたオールドコレクションとよく似たものであった。どちらの標本も今となっては産地・産出層準ともはっきりしないものであり、結局のところ“アヴァケラトプス”に属する可能性は十分あるが、ひょっとするとフルカトケラトプスは最初期に発見された角竜のひとつだったということさえあり得るのである。
(MOR 692の鱗状骨は不完全だが、老齢個体であるらしいとはいえ保存されているホーンレットはごく低く、USNM 2415や4802にみられるよく癒合し、かつ突出したものとはだいぶ趣が異なる。USNM 2415とUSNM 4802にはepsの関節面と思しきものがみられ、この点もNSM PV 24660とよく似ている。ANSP 15800はサイズからして相当な若年個体であると思われ、このあたりについては何とも言えないところである。)
フルカトケラトプスの原記載論文は頭骨の記載に重きを置いており(下顎についてはあらゆるケラトプス科角竜の中で最高と言える標本である)、首から後ろの要素についてはわりあい簡潔な記載に留められている。とはいえ端々で詳細な再記載――将来的なモノグラフの出版を匂わせており、今後の研究に非常に期待が持てるところである。なにしろフルカトケラトプスのホロタイプはジュディス・リバー層産の角竜としては空前の保存状態・完全度を誇るものであり、これまで発見されたナストケラトプス族としても最も完全な骨格であり、そしてなにより公的な研究機関――博物館の所蔵する標本なのである。
フルカトケラトプスのホロタイプの産地はちょっとしたボーンベッドとなっており、ハドロサウルス類の四肢骨をはじめとする共産化石もまとめて国立科学博物館の標本となるようだ。コール・リッジ部層の上部――州立恐竜公園におけるダイナソー・パーク層下部と対比される層準――にぽっかりと開いた窓から差し込む光もまた、我々の手の中にある。
角竜の研究の始まった場所、ジュディス・リバー層における悪夢のような状況を打破しうるものとして当初より期待が寄せられていた“AVA”は、かくしてフルカトケラトプス・エルキダンスのホロタイプNSM PV 24660として国立科学博物館に安住の地を得た。「解き明かす」という意味の種小名を背負ったNSM PV 24660は、相当部分を非常によく保存した数少ないケラトプス科角竜として、ジュディス・リバー層産の角竜としてはもっとも完全な骨格として、そして謎の多いナストケラトプス族の中でも最良の標本として、これからも歩みを止めることはない。

↑Skeletal reconstruction of cf. Denversaurus schlessmani
“Tank” FPDM-V9673 (formerly HMNS 11). Scale bar is 1m.
Note: recovered postpectoral osteoderms are not illustrated.
「デンヴァーサウルス」という名前を覚えたのはいつのことだったろう。豊橋市立自然史博物館へ贈られたホロタイプのキャストや、かつての林原が購入した“タンク”の愛称のついた部分骨格の存在によって日本とも縁深い恐竜であり、読者の方はどこかしらで一度は目にしたことがあるだろう。デンヴァーサウルス・シュレスマニ、あるいはエドモントニアの一種として扱われるこの恐竜はその実、相当に実態の不明瞭なものでもある。
パラエオスキンクス_Palaeoscincus_――なにがしかのノドサウルス類の歯の発見で始まった北米産鎧竜類の研究だが、しばらくのうちはまとまった骨格要素が見つかることはなかった。歯や単離した皮骨の発見が精々であり、それらが同じグループの恐竜に由来するとさえ思われていなかったのである。
ララミー層あるいは“ケラトプス層”では、ハッチャーの時代からそうした化石の産出が知られていたが、それらの化石が妥当な同定を受けるまでには程遠い状態であった。一帯ではなにかしらの剣竜の類縁――角竜の化石がよく知られており、直接的な証拠は何もなかったものの、この一帯で見つかる単離した皮骨(様々な鎧竜のものに加え、パキケファロサウルス類の頭骨に見られるスパイクのクラスターも含まれていた)が角竜のものであることは間違いないように思われていたのである。こうした状況にあって、ハッチャーがランス層から持ち帰ったスパイクUSNM 5793が(採集者の当人によって)角竜、おそらくはトリケラトプスの尾の基部に2列で並ぶものと考えられたのは無理もないことであった。
とはいえ、1910年代になるとこうした見方は大きく変わるようになった。カナダやアメリカ――カンザス周辺から様々な「装甲恐竜」が発見されるようになりはじめ、「角竜の鎧」がそうした恐竜の皮骨と区別できないらしいことが明らかになったのである。1914年にスミソニアンの「装甲恐竜」についてのモノグラフをまとめたギルモアは、USNM 5793がホプリトサウルス_Hoplitosaurus_――もともとステゴサウルス類とみなされていたが、ギルモアによる再記載以降はもっぱらポラカントゥス類とされている――のスパイクとよく似ていることを見て取った。ギルモアはこのほかにもハッチャー採集の「トリケラトプスの鎧」が鎧竜のものであるらしいことを見抜いたのだった。
このあたりの時期から加速度的に北米産の鎧竜に関する理解が進んだ。全体像がはっきりしないのは相変わらずであったが、それでも関節のつながった部分骨格――しばしば鎧を(おおむね)生前の位置に留めていた――がカナダで複数発見されるようになり、「鎧」の実態が明らかになりつつあったのである。
とりわけ大きな発見がレッド・ディアー川――ベリー・リバー層(現ダイナソー・パーク層)であった。第二次化石戦争もほぼ終わっていた1915年、すでにカナダ地質調査所の調査隊から離れていたリーヴァイ・スターンバーグによって発見されたその骨格――AMNHが入手した――は、ほぼ完全な状態で保存された鎧竜の前半身であった。この標本AMNH 5665――その壮絶な装甲っぷりにマシューは「動物界の超ド級戦艦」と呼んだ――はラングらAMNHのキュレーターの根性で三次元的にクリーニングされ、今日まで至るAMNHの看板展示のひとつに収まったのである。
頭骨の保存はあまりよくはなかったのだが、とはいえ歯はいくらか保存されており、マシューはAMNH 5665をパラエオスキンクスと同定した。マシューとブラウンはこれをパラエオスキンクスの新種として記載するつもりであったのだが、どういうわけかとうとう記載せずじまいに終わった。マシューによる(属止まりの)AMNH 5665の同定はギルモアら後続に追認され、しばらくの間(1919年にパノプロサウルスが、1928年にはエドモントニアが命名されてはいたのだが)アルバータやモンタナの上部白亜系から見つかるノドサウルス類はざっくりとパラエオスキンクス属にまとめられるようになったのである。
(1930年にギルモアはモンタナのトゥー・メディスン層産の標本USNM 11868を記載するにあたり、マシューによる1922年のAMNH 5665の「紹介」に大きな影響を受けた。ギルモアはパラエオスキンクス・コスタトゥス(模式種)のホロタイプ(ジュディス・リバー層産)とUSNM 11868の歯の形態が異なることからこれをパラエオスキンクス・ルゴシデンスと命名し、1928年に命名されたばかりであったエドモントニア・ロンギケプスもパラエオスキンクス属に過ぎない可能性をも指摘したのである。ギルモアはこの時AMNH 5665がパラエオスキンクス・コスタトゥスであるという前提の下で自らの議論を展開していたのだが、実のところマシューはAMNH 5665については上述の通り(歯の形態に基づいて)パラエオスキンクス属の新種と考えていた。やがてパラエオスキンクス・ルゴシデンスはエドモントニア属に移され(後述)、AMNH 5665も結局エドモントニア・ルゴシデンスとされて今日に至っている。)
こうした状況が整理されたのは1940年になってからで、ラッセル(ロリスの方)によってこれらのノドサウルス類は2属3種――パノプロサウルス・ミルス_Panoplosaurus mirus_、エドモントニア・ロンギケプス_Edmontonia longiceps_そしてエドモントニア・ルゴシデンス_E. rugosidens_(上述の通り、原記載ではパラエオスキンクス属だった)へとまとめられた。このコンセプトは(紆余曲折ありつつも)究極的には現代まで踏襲されるもので、パラエオスキンクスは疑問名として扱われるようになったのである。
その後表立った動きのない状況が長く続いた(1970年代後半に鎧竜の分類の大改訂に取り組んだクームズがついでにエドモントニア属をパノプロサウルス属のシノニムとしたくらいで、種レベルの話は特に何もなかった)のだが、1986年になり、カーペンターは北米西部における“ランス期”のノドサウルス類(カーペンター自身が一時パラエオスキンクスと呼んでいた一連の標本)をまとめて記載した。恐竜の絶滅に関する議論が盛んになりつつある中で当然ノドサウルス類の絶滅した時期も問題になったのだが、そのあたりをまともに取り扱った研究が何もなかったのである。
先述のスパイクUSNM 5793はここでやっとエドモントニア属のスパイクであるとされたが、この時カーペンターはサウスダコタのヘル・クリーク層から産出したほぼ完全な頭蓋の記載も行った。この標本DMNH 468は上下方向からひどく押しつぶされていた(鎧竜の頭骨は程度の差はあれだいたい上下方向に潰れているものでもある)が、もろもろの特徴からカーペンターはこれをパノプロサウルス属ではなくエドモントニア属とした――が、種については特に言及することはなかった。カーペンターはランス期のノドサウルス類の化石記録が白亜紀の最末期にまでは到達していない(ヘル・クリーク層やランス層の上部では産出していない)ことを指摘し、恐竜の「緩やかな絶滅」の証拠のひとつとしたのだった。
かくして種不定のまま記載されたDMNH 468に目を付けたのはバッカーであった。1988年にバッカーはノドサウルス「上科」に関する総説を出版し、その中でパノプロサウルス属とエドモントニア属をやたら細分化したのである。エドモントニア・ルゴシデンスを新亜属チャズスターンバーギア_Chassternbergia_とし、そしてバッカーはDMNH 468を新属新種のエドモントニア「科」、デンヴァーサウルス・シュレスマニ_Denversaurus schlessmani_としたのであった。
バッカーのこの細分化の受けは極めて悪かった(例によってオルシェフスキーは別だったが)。事実上、標本1つにつき(命名の有無はさておき)1つの種を設けたような状況だったのである。他のエドモントニア類よりもずっと新しいDMNH 468に学名を与えたという意義(だけ)はあったが、その程度であった(そしてノドサウルス上科がアンキロサウルス科よりもステゴサウルス類に近縁であるとするバッカーの意見はその後特に顧みられなかった)。カーペンターは1990年の論文でこのあたりをばっさり切り捨て(同時期にクームズも同様の意見に達していた)、改めてロリス・ラッセルによる分類コンセプトを提示するとともに、DMNH 468が(より時代の近いエドモントニア・ロンギケプスではなく)エドモントニア・ルゴシデンスと最もよく似ていることを指摘した。バッカー言うところのデンヴァーサウルスの特徴のうちのいくつかはアーティファクトめいていた(バッカーはDMNH 468の頭骨復元図に基づいて論じている部分が少なからずあり、一方でバッカーの復元は相当に客観性を欠いていた)のである。一方で変形のために形態のはっきりしない部分もあることから、カーペンターはやはりDMNH 468をエドモントニアの一種としたのだった。
(カーペンターによる1990年の論文は、AMNH 5665の骨学的記載としてはいまだにほぼ唯一のものとなっている。小柄なカーペンターをしてマウントの下に潜り込んで椎骨の観察を行うというだいぶ無茶な作業によるものであった。)
そんなわけでランス期のエドモントニア類の分類については判然としないままだったのだが、その間に大きな動きがあった。1988年、ワイオミングのランス層で大きな鎧竜の部分骨格――“タンク”の愛称で呼ばれるそれが発見されたのである。“タンク”を発見したのは業者であり、化石は一通りのプレパレーション(復元骨格まで制作された)を経て日本の企業――林原へと渡った。
(このあたりの経緯はつまびらかではないのだが、“タンク”の復元骨格は2タイプある。林原(→福井)の所蔵であったキャストは最初に作られたタイプであるのだが、これと同型のもの(に一体成型の「甲羅」を加えたもの)がトリーボールドの私設博物館で展示されている。福井県博で常設展示されている実物のマウントも、アーティファクト部分に関してはこれと全く同型である。BHIは林原からキャストの制作・販売権を取得し、アーティファクトをリメイクした(後述)バージョンを販売しているが、初期のタイプを制作していた業者は(トリーボールドの可能性はあるが)はっきりしない。)
かくして“タンク”の形態やら分類やらは当初(カーペンターやラーソンらによる)「目撃談」頼りではあったのだが、頭蓋と尾を除く骨格の相当な部位を保存しており、かつ前方へ向けられた「二重スパイク」――パノプロサウルスではみられない――も残されているらしいとの話であった。つまるところこれは(少なくともカーペンター言うところの)エドモントニア属であるらしく、(直接比較はできないものの)産地からしてヘル・クリーク産の「エドモントニアの一種」と同じ種であるように思われたのである。
はるばる岡山へと渡った“タンク”はそこでそれなりの研究材料としての扱いを受けた。頭蓋はごく断片的だったものの、明瞭な分岐のある「二重スパイク」の存在からエドモントニア・ルゴシデンスと同定され(言うまでもなくエドモントニア・ロンギケプスも二重スパイクを持っており、このあたりの判断は相当に恣意的ではある)、予察的な機能形態学的、古病理学的な解析をも受けたのである。林原自然科学博物館が設置したパナソニックセンターで“タンク”のキャストが展示されるようになったが、一方で配列のはっきりしなかった方から後ろの皮骨については(相当な数が採集されていたとはいえ)マウントされないままであった。
さて、林原から“タンク”のキャストの販売権を取得したBHIであったが、それとは別に手元にヘル・クリーク層産のノドサウルス類の頭骨が2つ――頭蓋天井といくらかの皮骨を含むBHI 6225と左右からひどく圧縮されているものの完全な頭骨BHI 6332があった。どちらもデンヴァーサウルス・シュレスマニのように思われ、産出層からすれば“タンク”と同種のようにも思われたのである(“タンク”の頭蓋はごく断片的であり、いずれの頭骨とも直接比較できない点に注意)。これらの頭骨と“タンク”に据えられていた出来のよくないアーティファクトとの違いは歴然としていた。BHI 6332は完全な頭蓋ではあったが(ノドサウルス類の頭骨としては珍しく)左右方向からぺしゃんこになっていたため、BHI 6225のキャストに欠損部を継ぎ足したものが新たに量産された“タンク”に据えられた。仕上げに手足のアーティファクトもブラッシュアップされた(原型はほぼそのままだったが)“タンク”は、かくして“Denversaurus (Edmontonia) cf. _schlessmani_”の「商品名」で、デンヴァーサウルスの代表格のごとく世界へ出荷されていったのである。一方で、林原から特別展へと貸し出される“タンク”のキャストはあくまでも「エドモントサウルスの一種」あるいは「エドモントニア・ルゴシデンス」止まりのままであった。
{kind=link}
{kind=link}
{kind=link}
(実のところBHI 6225は“タンク”よりもやや大きな個体の頭蓋天井であり、従ってBHIの“タンク”は本来よりも大きな頭蓋を持つものとしてマウントされている(適切なアーティファクトとは言い難いが、サイズ感については当然オリジナルの“タンク”の方が正確である)。BHIは“タンク”のハーフリングをオリジナルのマウントとは違ったパターンで組み立てている(オリジナルの二重スパイクを第2ハーフリングへ移し、その後ろにあったスパイクをそのまま第3ハーフリング(二重スパイクの位置)へスライドさせている)が、これは明らかな誤りで、林原-FPDMのマウントの方が正確である。)
デンヴァーサウルスの名を冠した「商品」が各地へ広がる一方(なにしろエドモントニア類もといパノプロサウルス類の復元骨格として流通しているのは“タンク”が唯一無二であった)、DMNH 468の再検討はなかなか進まなかった。研究に相当な支障をきたすレベルで変形しているのは間違いなかったし、比較されるべきパノプロサウルスやエドモントニアのレビューさえ1990年が最後だったのである。
このあたりの問題――パノプロサウルス亜科の再評価に取り組んだのはバーンズであり、博士課程の研究においてパノプロサウルスやエドモントニア各種、そしてBHIの「デンヴァーサウルス」まで広範な再検討が行われた。DMNH 468やBHI 6225にはパノプロサウルスやエドモントニア2種の特徴がモザイク的にみられた一方で固有の特徴らしいものは見出されなかったが、それら3種と全体の形態が一致することもなかった。そして系統解析の結果この2標本は姉妹群をなした一方、エドモントニアやパノプロサウルスの標本群とは独立した枝をなした(パノプロサウルスとそれらが姉妹群をなし、エドモントニア属はその基盤の側系統をなした)のである。バーンズはここに(いまだ博論止まりではあるのだが)デンヴァーサウルス・シュレスマニを復活させたのである(左右方向の変形がひどかったために頭蓋天井の比較ができなかったBHI 6332はひとまずパノプロサウルス亜科の不定種とされ、そもそも頭蓋がほぼ残っていない“タンク”は研究材料とはされなかった)。
バーンズによるパノプロサウルス亜科のレビューはいまだ出版されてはいないのだが、とはいえ(時代の違いが明確であるものを積極的に同種にまとめることもなかろうという昨今の流れもあって;必ずしも適切な判断とも限らない)デンヴァーサウルス・シュレスマニは研究者のコミュニティでも独立種として扱われるようになった。BHI 6332、ひいては“タンク”も(現状直接比較が困難ではあるが)デンヴァーサウルス・シュレスマニと見て問題はないだろう。
林原自然科学博物館のうち主だったコレクションは福井県立恐竜博物館へ渡り、“タンク”の標本番号もHMNS 11からFPDM-V9673へと変わった。アーティファクトはそのままに、(頭骨以外は)初めて実物でマウントされた“タンク”は、林原時代からおなじみのキャストとは異なり、肩から後ろまで完全に装甲された姿を現すこととなった。
今度こそ終の棲家を得た“タンク”だが、研究材料――博物館の収蔵物としての正念場はここからである。林原時代に行われた研究は予察的なところで止まっており、肩から後ろの皮骨の配置もはっきりしないままだが、しかしこれはパノプロサウルス亜科としてはAMNH 5665さえ遥かにしのぐ完全度の骨格なのである。眩しい点光源を浴びながら、“タンク”は新たな研究者を待ち続けている。
(カーペンターらによる1986年の研究の後、ランス期のノドサウルス類に関する生層序学的な議論は特に行われておらず、カーペンターらの言うようにデンヴァーサウルスがマーストリヒチアンの終わりを前に絶滅したのかどうかははっきりしていない。そもそもの産出数が相当に少ないのも間違いない話で、少なくともトリケラトプスのような話は現状望むべくもない。)
いい加減で春であり、筆者の花粉症のピークは先々週くらいだったらしい。そんなわけで春の特別展はすでに始まっており、今度こそは(2019は招待されつつ当時の本職の都合で行けなかったので)内覧会に顔を出しつつ初日にもう一度観に行けた筆者である。そういうわけで、この半年ちょっとの間筆者を苦しめていた恐竜博2023についてつらつらと紹介していきたい。

のっけから事前情報の一切なかったスクテロサウルスである。最近の再記載やらを踏まえたマウント(昨年のツーソンショーでお披露目されたばかり)で、出来もなかなかといったところ。

吻やら肘から先がアーティファクトだったりはするのだが、それをさておいてため息の出る産状である。開催のわりあいに直前になって情報が解禁されたスケリドサウルスだが、これ目当てに2200円をつぎこめるだけの代物である。

しばらく前からいろいろと描き溜めていた超大型ティタノサウルス類を結集して描いたものである。実物の6割弱のサイズであることは見ての通りである。

しれっと置かれているが、胴椎(というか神経弓)は特に既知の恐竜の中では最も左右幅があり(アルゼンチノサウルスやパタゴティタンは物の数ではない)、アクリル板越しに置かれるでもなく観察ができる。この手の巨大な骨化石のキャストとしては非常に抜きがよく、原記載の写真が悲惨だったこともあって大変にありがたいところ。

先日報告されたピナコサウルスの咽頭骨(の3Dプリント)やFPDMのデンヴァーサウルスやらオヴィラプトル類の胚のキャスト(どうもそのうち実物に展示更新されそうな気配がある)やらサイカニアのホロタイプの頭骨キャストやらいぶし銀の標本(とピンチヒッターのFPDM出張装盾類セット)が続くが、そこを抜ければもうズールのお出ましである。
頭骨を全周囲から観察できる粋なレイアウトであり、この空間だけ照明が暗いのだが、とはいえありがちな点光源照明下とは比べ物にならないほど見やすい。見せ所をよくわかっている展示のありがたみを痛感させてくれる。

腹側はもうこれ以上クリーニングができない(というよりジャケットをもう外せない)ため、ズールの胴体ブロックの腹側はこのキャストである意味見納めである。意外なほど関節がゆるんでおり、背側の産状とあわせて興味深い。壁に垂直に貼り付けてあるので、人波が小さくなった隙を突くことで自在に写真が撮れるという思いやりにあふれている。

ズールの胴体ブロックはどう展示してくるかと思いきや、床置きなのはわかるとして尾のブロックとそれから頭部、首(頸椎と第1ハーフリングは実物)を並べた、全身骨格と言える状態でのレイアウトで度肝を抜かれた筆者である。第3頸椎や第2ハーフリング(あまりにも保存が悪かったためかキャストである)は未記載であり、胴体ブロックに至っては直近の研究で図示された状態からさらにクリーニングが進んだ(というかこれで打ち止めだろう)最終形態といえるものである。写真やイラストで示される情報量とは比べ物になるわけがなく、なんなら1回や2回行ったごときでは一瞬で脳がオーバーフローを起こすことうけあいである。ズールのホロタイプとはさりげなく付き合いの長い本ブログであり、このあたり謎の感慨も(勝手に)ある。

恐ろしいことにズールの「全身骨格」は通路の上からも(ガラス越しではあるが)観察可能なレイアウトで、ひたすらにズールをしゃぶりつくさせようという展示設計者の慈悲がうかがえる。全体的に明るくかつ点光源が悪さをしない照明、ゆとりのある(=人波が引くのを待てる)空間、標本の見せたいポイントと見たいポイントをきっちり押さえたレイアウトと、恐竜博2023の展示設計は神がかり的な噛み合いのよさを見せており、筆者の記憶にある限りでは数ある恐竜絡みの展示の中でも断トツで優勝である。

キービジュアルになってはいるのだが、展示品のそうそうたる並びからすると箸休めといったところである。とはいえゴルゴサウルスROM 1247(のキャスト)もズール(用の改造パーツを組み込まれたエウオプロケファルス・トゥトゥスないしスコロサウルス・スロヌスのホロタイプROM 1930のキャスト)のマウントもキャストの精度は良好で、特に後者は色々と観察のし甲斐がある。

内覧会でご一緒した相場大祐博士(ポケモン化石博物館の前に異常巻きアンモナイトやさん同士なんかしら引き合うものがあるらしい)にROM 1247のマウントの頭骨の写真の撮り方を披露してドヤっていた筆者だったのだが、しれっとROM 1247の頭骨の実物が来ていたというオチである。(話には多少聞いてはいたが)おどろくほど保存がよく、キャスト用の型を制作した後で徹底的な再クリーニングが行われたことがうかがえる。恐ろしいことに図録にはバラした各要素がでかでかと(頭骨のマウントには組み込まれていない角骨・上角骨も含めて)図示がある。

しばらく前にトリーボールド社が掘っていた標本("Bert"という愛称も付いている)なのだが、かはくが(恐竜博2019の時のイクチオルニスや、次に紹介する”AVA”と同様に)購入していたわけである。依然としてテスケロサウルスの保存のよい頭骨は珍しく、(すでに歯のマイクロウェアに関する研究には供されているとのことだが)いじりがいのある標本だろう。

どこかで見たことがあると思っていた古参読者の方は大正解、本ブログでずいぶん昔にネタにしていた“AVA”はいつの間にかかはくに購入されていたのである。なんならすでに記載論文は投稿済みという話であり、何もかもうまくいけば年内には学名が付くようだ。

ジュディス・リバー層がどうのこうのは置いといて保存は非常によい。下顎のデンタルバッテリーなどはちょっと異様なほどである。
例のごとく(おかげさまで例のごとく、になりつつあるようだ)筆者が骨格図を描いていたりするのだが、(単にレイアウト上の問題で)図録に載っているのはバストアップだけである。なんなら同人誌に載せた頭骨図はダミーの古い図だった(し、その時点での最新バージョンだった図は結局没にした)という話は秘密である。

大人の事情の薫り高い展示であり、正直なところ他に書くことは特にない。“スコッティ”(なりなんなり、素性のはっきりしたマウント)が隣にいて初めて成立する展示でもある。

カルノタウルスのマウントはいつもの茨城県博の遠征要員(初来日した時のマウントであり、どうしていいのかわからなかった手も含めて元祖ボナパルテ復元を今に伝えている)なのだが、足元にはフルクリーニングされた右前肢のキャスト(本展にあわせてフルクリーニングしたとのことで、いずれは論文のネタになることだろう)が置かれている。つまるところカルノタウルスの手は完全な状態であったらしく、今後の波乱を予感させる。

福井からのピンチヒッターであるフクイラプトルとメガラプトルを従えつつ、どうにかして来日したマイプのホロタイプの実物(と胸郭の複製)が展示されている。このご時世もあってプレパレーターが在宅でクリーニングしたという代物らしいのだが、保存状態は非常によく、クリーニングも大変丁寧である。そこかしこの破断面からハニカム状の含気構造が顔を覗かせている。

マイプの第6胴椎(胴体の左右幅が最も広くなる位置である)まわりはだいぶ断片的なのだが、とはいえアーティファクトの出来は(あらゆる意味で)非常にしっかりしており、マイプの種小名の意味、ひいてはメガラプトル科の胸郭のなんたるかを物語っている。マイプのホロタイプはメガラプトルのマウントのベースになっている標本よりも(全長で)35%大きい(筆者調べ)ので、そのサイズは頭上のメガラプトルのマウント(前方胴椎が一つ多かったり、尻尾がだいぶごつかったりはするのだが)から推して知るべしである。
さんざん書いたのだが、これでも展示の紹介はずいぶんかいつまんでいる。海外からの標本を用意しにくい(コロナ禍ということもあるし、少し前の円相場は相当悪さをしたようだ)という中にあって来日した標本はズールを筆頭に凄まじいものばかりで、ズールに関して言えばおそらくもう二度と来日することはないだろう。そうした標本の合間を国内の博物館からの出張組がうまくつないでおり、かはくの特別展会場の規模からしてみればボリューム不足ということは全くない。
上にも書いたことであるが、標本の点数と空間のバランスというところも含め、照明や展示レイアウト等々、展示設計としても本展は出色のものであった。一昨年の恐竜科学博もそうだったわけだが、(昨今の情勢による怪我の功名的な側面も多分にありそうだが)標本一つ一つをしっかりと見せる/見られる特別展示が確立されてきたということでもあるのだろう。
そんなこんなで筆者の骨格図(を原図としたもの)がそびえたっていたり、ほかにもマイプや“AVA”のフル骨格図を描くなりなんなり(マイプについては3Dペーパーパズルのたたき台にもなっている)、筆者なりに今まで勝手に因縁をつけてきた相手とひとまずの決着をつけるいいおしごとをこの半年間いただいていた格好でもある。これだけの特別展の賑やかしになれば、この手のおしごととしては何よりなのだ。