The orthography-specific functions of the left fusiform gyrus: Evidence of modality and category specificity (original) (raw)
. Author manuscript; available in PMC: 2011 Feb 1.
Abstract
We report on an investigation of the cognitive functions of an individual with a resection of the left fusiform gyrus. This individual and a group of control participants underwent testing to examine the question of whether or not there are neural substrates within the left fusiform gyrus that are dedicated to orthographic processing. We evaluated the modality specificity (written vs. spoken language) and the category specificity (written language vs. other visual categories) of this individual’s impairments. The results clearly reveal deficits affecting lexical processes in both reading and spelling. Specifically, we find disruption of normal, rapid access to meaning from print in reading and of accurate retrieval of the spellings of words from their meaning in writing. These deficits stand in striking contrast with intact processing of spoken language and categories of visual stimuli such as line drawings of objects and faces. The modality and category specificity of the deficits provide clear evidence of neural substrates within the left mid-fusiform gyrus that are specialized and necessary for normal orthographic processing.
Keywords: fusiform gyrus, VWFA, reading, spelling, orthographic processing
Introduction
The question of whether or not there are specific regions of cortex dedicated to the representation of category-specific knowledge or processes is easily one of the oldest and most fiercely debated questions in the history of neuroscience. Variations on the functional localization versus holism (mass action, equipotentiality) debates of early neuroscientists such as Broca, Ferrier, Golgi, Goltz, Hughlings Jackson, Lashley, and others (for a review see Finger, 1994), have been rekindled over more than a century as different approaches and techniques have gained prominence, from phrenology to fMRI. Furthermore, similar discussions have played out over a wide range of cognitive domains. For example, the past decade has seen a great deal of research directed at determining whether face recognition or specific semantic category knowledge (e.g., foods, animals, tools) have dedicated neural substrates or if, instead, the apparent specificity of their neural instantiation is a simply a mirage emerging from the distributed interactions of networks carrying out broader functions (see Baker, et al., 2007a; Baker et al., 2007b; Downing et al., 2006; Tarr and Gauthier, 2000; for reviews). A domain that has quite recently entered this debate is that of written language processing, and the brain area that has received considerable attention is the left mid fusiform gyrus, referred to by Cohen and Dehaene and colleagues (Cohen et al., 2000; Cohen et al., 2002; Dehaene et al., 2002; Cohen et al., 2004; Dehaene et al., 2004) as the “visual word form area”. Certain research findings have been put forward in support of the conclusion that there are neural substrates largely dedicated to or indispensable for written language processing, while other findings have been put forward as evidence that written language is, instead, processed by neural substrates that have broader polymodal language functions or more general visual functions. That is, the debate concerns the modality specificity and the category specificity of the neural substrates that have been identified with orthographic processing.
The data relevant to the debate have come primarily from functional neuroimaging studies, but also include cognitive neuropsychological studies of individuals with acquired deficits in written language processing (Cohen, Henry et al., 2004; Cohen, Lehericy et al., 2004; Cohen et al., 2003; Gailard et al., 2006; Raymer et al., 1997). Each approach has well-known strengths and weaknesses (Gailard et al., 2006; Kleinschmidt and Cohen, 2006; Price et al., 2003). While functional neuroimaging can reveal areas that are active during processing, it does not reveal which of these areas are causally necessary for performing the task. Neuropsychological studies relating deficits to lesions can reveal necessary areas and establish a causal relationship, but may be limited by uncontrollable factors such as the extent and nature of the damage to the brain. Ideally, evidence from lesion and functional neuroimaging studies should converge to identify the brain areas that make causal and necessary contributions to specific cognitive functions. In this paper we report on the case of an individual who had a resection of the mid and anterior portion of his left fusiform gyrus. Because the lesion was restricted and because we were able to carry out detailed testing of written and spoken language, as well as object and face processing, the results provide strong constraints on the current debate. On the basis of our findings, we will conclude that there are substrates within the left fusiform that are specialized and necessary for normal written language comprehension (reading) and production (spelling).
Orthography-specific neural substrates
To many neuroscientists, the possibility that specific neural tissue would be largely dedicated to orthographic processing seems particularly unlikely given the recency (3,000 years) with which written language has entered the human repertoire (Farah and Wallace, 1991). Nonetheless, the notion of orthography-specific substrates dates back to the beginnings of cognitive neuroscience. Critical evidence was Dejerine’s (1892) cognitive neuropsychological and post-mortem anatomical observations of an individual who, as a result of cerebral infarcts, acquired written, but not spoken language or object recognition, impairments. In the Dejerine case, the neuroanatomical data pointed to the critical role of the left angular gyrus in orthographic processing. More recently, a great deal of attention has been focused on the role of left mid-fusiform gyrus, and both functional neuroimaging and cognitive neuropsychological evidence primarily from reading, but also from spelling, has been brought to bear on the question.
Reading
Cohen et al. (2000, 2002) specifically proposed a visual word form area (VWFA) centered approximately on Talairach coordinates (x= −43, y = −54, z = −12) and extending some two centimeters in the rostral-caudal dimension along the fusiform gyrus. They argued that this region “plays a particular and largely indispensable role in the recognition of visual words, while it may not be strictly necessary to the efficient perception of other visual categories such as faces, objects or scenes” (Gaillard et al., 2006). While a number of functional neuroimaging studies of reading had previously reported activation in the inferior, posterior temporal areas (among others: Beauregard et al., 1997; Nobre et al., 1994; Puce et al., 1996; Pugh et al., 1996, 2001; Salmelin et al., 1996; Wagner et al., 1998), Cohen et al. (2000) drew specific attention to the mid-fusiform region. They carried out a number of studies that indicated that this area is very reliably activated (at the group and individual subject level) by written words and letter stings relative to low-level visual stimuli such as checkerboards and also relative to other categories of visual objects such as faces and houses (Cohen et al., 2000, 2002; Cohen et al., 2003; Cohen et al., 2004; Cohen and Dehaene, 2004; Dehaene et al., 2004; Dehaene et al., 2005; Gaillard et al., 2006; Vinckier et al., 2007). These results supported the category specificity of the visual processing within this mid-fusiform region. With regard to the modality specificity, evidence was also provided that this area is not activated by auditory stimuli. For example, Dehaene et al. (2002) specifically evaluated the response of functionally identified VWFA voxels to the presentation of auditorily presented words. In both group and single-subject analyses they failed to find any significant response to spoken stimuli. Similarly, Cohen et al. (2004) found that voxels in a functionally identified VWFA were responsive to the repetition of written words but showed no response to repeated spoken words. Furthermore, Binder et al. (2000) also failed to find left inferior temporal activations for passive listening of speech.
Subsequent work has been dedicated to elucidating the specific orthographic functions of this region. For example, Dehaene, et al. (2005) and Cohen et al. (2003) proposed that letter stings are hierachically coded in the left fusiform gyrus such that as processing proceeds in a posterior to anterior direction, it is carried out by neuronal detectors that are increasingly complex, abstract and location-invariant. Subsequent findings provide further support for this gradient of complexity in the organization of the fusiform (Binder et al., 2006; Vinckier et al., 2007). Dehaene et al. (2005) specifically proposed that there is a progression from processing in visual areas V1–V4 (TC y= −90 to −70) that are sensitive to physical characteristics such as word length, visual contrast, rate and duration, through processing by letter detectors located more anteriorly (y= −64), on to bigram detectors (y=−56) and then, finally, morpheme detectors (y=−48). Consistent with this general notion of increasing caudal-to-rostral abstraction, the most anterior region of the fusiform has been associated with multimodal word processing or access to semantics by a number of researchers. That is, even researchers such as Cohen and colleagues who have proposed orthography-specific processing in the mid to posterior areas of the fusiform have proposed that the anterior region may be polymodal. This anterior region has been given different names such as Lateral Inferior Multimodal Area (LIMA; centered on x= −48, y= −60, z= −16 ) by Cohen et al., (2004), or Basal Temporal Language Area (BTLA; centered on x= −50, y= −44, z= −10) by Luders et al. (1991; and see Jobard et al., 2007) and this characterization is generally consistent with Damasio’s (1989) claim that the left lateral temporal cortex constitutes a convergence zone supporting the linkage of orthographic, phonemic, and semantic information (Damasio, 1989; see also, Krauss et al.,1996; Demonet et al., 1992, 1994; Binder et al., 1996; Booth et al. 2002a, 2002b; Buchel et al., 1998; Cappa et al., 1998; Fiez et al., 1999; Hagoort et al., 1999; Thompson-Schill et al., 1999; Buckner et al., 2000; Crinion, et al., 2003; Giraud and Price, 2001; Hillis et al., 2005; Jobard et al., 2003; Kreiman et al., 2000; Lambon Ralph, et al., 2003; Cohen et al., 2004; Sharp et al., 2004; Jobard et al., 2007).
With regard to the lesion evidence, there have been various cases of a type of acquired alexia that is referred to as “pure alexia” or “letter by letter reading”, in which there is a deficit of parallel processing in word reading accompanied, at least in some cases, by sparing of spelling abilities and auditory word comprehension (Damasio and Damasio, 1983; Leff et al., 2001; Binder and Mohr, 1992). Cohen et al. (2000, 2002) reviewed a number of these cases underscoring the lesion site in the occipitotemporal cortex. Cohen et al. (2003) presented a series of six cases with varying lesion sites and behavioral patterns that supported the claim that the lesion site for pure alexia is within the vicinity of the VWFA. As concerns the category specificity of the VWFA, there have been several reports of cases of selective alexia without prosopagnosia (Feinberg et al., 1994) and vice versa (Farah et al., 1998), and of selective visual object agnosia without alexia or prosopagnosia (Humphreys and Rumiati, 1998; Rumiati et al., 1994) and vice versa (Buxbaum et al., 1999; De Renzi et al., 1987). Such cases would seem to constitute compelling evidence for category-specific orthographic substrates.
Nonetheless, as Price and Devlin (2003) have noted, in most of these cases the lesions are usually large and also compromise occipital cortex and, as a result, pure alexia cases almost invariably have some additional visual processing deficits in color naming and/or object processing. However, quite recently a very compelling case has been reported by Gaillard et al. (2006). The case concerns an individual who underwent surgical resection of a circumscribed area of the left fusiform for treatment of epilepsy. Prior to surgery he exhibited normal reading, spelling to dictation, written lexical decision and oral language comprehension, production and repetition. Also, before surgery he underwent fMRI scanning to functionally identify his VWFA which was found to be centered on prototypical coordinates (x= −42, y= −57, z= −6). The surgical lesion was relatively small, extending from y=−60 to −80. Given that the lesion was located posterior to his VWFA, the authors suggested that the lesion effectively deafferented the VWFA from visual input. Consistent with this characterization, they report that, subsequent to surgery, spelling, face and object recognition and spoken language remained intact in the face of a selective and severe reading impairment. In reading, the patient made large numbers of errors with briefly presented stimuli, showed overall increased response latencies, and exhibited a marked length effect. Arguably, the combined elements of this case make it one of the strongest and clearest pieces of evidence to date for a causal role of the left-fusiform in reading.
Spelling
Although there has been a dearth of functional neuroimaging work in written language production (spelling), evidence for an orthographic role for the mid-fusiform region in spelling has been reported. In an fMRI study, Beeson et al. (2003) reported significant activation at the prototypical VWFA coordinates (x= −44, y= −54, z= −12) when generative writing of words was compared to alphabet writing. Similarly, in an fMRI study of spelling, Rapp and Hsieh (Rapp and Hsieh, 2002 and Hsieh and Rapp, 2004) also reported significant spelling-related activation in this region. Moreover, when Rapp et al. (2006) examined both reading and spelling activations in the same individuals using the tasks employed by Cohen and colleagues to localize the VWFA, they found significant spelling-related activation within the functionally identified VWFA (x= −42, y= −43, z= −8). With regard to the cognitive neuropsychological evidence, Rapcsak and Beeson (2004) reviewed a number of cases of acquired dysgraphia with dyslexia. In these cases, the dysgraphia typically affected the lexical system (exception word spelling), leaving relatively intact sublexical processes (nonword spelling) and the lesions affected the left fusiform and posterior inferior temporal gyri.
In sum, there is a considerable body of functional neuroimaging and lesion evidence from both reading and spelling in support of the claim of orthography-specific functions of a region of the left mid-fusiform gyrus.
Challenges to the claims of modality- and category- independent orthographic functions of the left fusiform
Both the modality- and the category-specificity of the functions of mid-fusiform have been vigorously challenged (Hillis, et al., 2005; Price and Devlin, 2003, 2004; Price et al., 2003; Mechelli et al., 2005; Price and Mechelli, 2005; Devlin et al. 2006). These opposing views tend to posit either that this region instantiates functions that are not limited to the visual modality or, if they assume that the functions are visual, they claim they are not specifically orthographic. Empirically these positions are based on functional neuroimaging and neuropsychological findings indicating associations among modalities and/or visual categories rather than on dissociations between them.
With regard to the question of category independence, Starrfelt and Gerlach (2007), for example, argued that the mid-fusiform region may be responsible for certain types of complex visual analysis that applies across categories (see also Moore and Price, 1999; Martin and Chao, 2001). Furthermore, as indicated above, Price and Devlin (2003) argued that the lesion data largely indicate that individuals with acquired reading deficits typically suffer from other visual processing deficit (Farah and Wallace, 1991; see also, Damasio and Damasio, 1983; De Renzi et al., 1987; Geshwind, 1965; Behrmann et al., 1998).
With regard to the issue of modality specificity, Price and Devlin (2004) describe this region as “a polymodal area that is clearly driven by visual input but also responds to tactile and auditory stimuli even during tasks that do not entail top-down activation of visual processes” (pg 478). These conclusions are based on a series of functional neuroimaging experiments and reviews of neuropsychological and functional neuroimaging data that report fusiform activation across a very wide range of tasks in which auditory words are presented, including tasks such as listening for meaning, rhyme detection, repetition of heard words, semantic decisions to heard definitions (Thompson-Schill et al., 1999; Booth et al., 2002; Price et al., 2002; see also Vigneau et al., 2005; Jobard et al., 2003; Price and Devlin, 2003, 2004). Furthermore, with regard to lesion data, it is worth noting that there is evidence that lesions to this area may result in spoken word production deficits. For example, Raymer, et al. (1997) described an individual with acute damage to the left posterior inferior temporal lobe who suffered from anomia with impaired picture naming and naming to definition as well as reading difficulties(see also Hillis et al., 2002). Similarly, many of the cases reviewed by Rapcsak and Beeson (2004) with lesions in this area and primary reading and spelling deficits also suffered at least mild anomia. In addition, Hillis et al. (2005) reported that lesions and/or hypoperfusion of the left fusiform were significantly associated with reading, spelling and spoken naming deficits.
Interestingly, just as Cohen and colleagues proposed subdivisions within the fusiform, Mechelli et al. (2005) did so also, but within a framework that assumes that fusiform functions are not modality or domain specific. They proposed distinctions between the anterior fusiform (y= −20 to −50) that is involved in semantic and supramodal tasks, middle fusiform (y= −50 to −60) that is critical for lexical retrieval in tasks such as reading and picture naming (McCandliss et al., 2003; Price and Devlin, 2003; Price and Friston, 1999), and posterior fusiform (y= −60 to −70) that is involved in sublexical processes such as in pseudoword reading (Mechelli et al., 2003). Mechelli et al. (2005) present evidence that these different fusiform subregions show differential patterns of functional connectivity with regions of the left frontal lobe that are thought to be involved in semantic and phonological processing.
In sum, opponents of the notion of orthography-specific neural areas argue that, although the left posterior inferior temporal region is clearly involved in visual word processing, the activations observed in this area likely reflect either more general visual processes or that the function of the area “can only be defined by specifying the set of interacting regions” (Price and Devlin, 2003).
The debate
The conflicting conclusions regarding the orthographic functions of the left fusiform have arisen essentially because of the different weight given to findings of associations and dissociations both in the performance of individuals with acquired deficits affecting this area and in the patterns of activation produced by different tasks. With regard to the functional neuroimaging data, differences across studies in terms of patterns of association and dissociation of activation can be attributed to a variety of factors, including differences in tasks, baseline conditions, thresholding, power, etc. Furthermore, because functional neuroimaging data can provide information regarding which areas are active but not which are necessary for a given cognitive operation, it has been argued that some of the findings of activation in the fusiform in non-orthographic tasks are the result of (automatic) unnecessary processing that subjects engage in. For example, Dehaene et al. (2002) suggested that fusiform activation in auditory tasks may come from subjects (unnecessarily) picturing the orthographic forms of the words. Others (Vigneau, 2005) claim that auditory and visual words activate similar areas because in processing the meanings of the heard or seen words subjects bring to mind the visual attributes of the objects that the words correspond to. As concerns the lesion evidence, associations among deficits are readily attributed to the fact that lesion size and location are not controlled and multiple geographically proximal functions may be coincidentally affected by a single large lesion. Generally speaking, dissociations in performance resulting from lesions represent the most compelling evidence of functional causality, linking a brain area and a cognitive function. In these cases, the major criticism is typically that the other cognitive functions that were seemingly intact were actually impaired, but insufficiently evaluated.
In the present study we address the questions of modality and category specificity of the functions of the left fusiform through the detailed investigation of both written and spoken language processing as well as object and face recognition in an individual with a lesion affects the left fusiform and some of the adjacent inferior temporal gyrus. The lesion is anterior to the one reported by Gaillard et al. (2006) and, as a result, the case provides a unique opportunity to make a contribution to the ongoing debate. We first report on testing directed at characterizing the written language deficits and then we go on to examine the questions of modality and category specificity.
Participants
DPT: Case history
DPT is a right-handed male (DOB: 9/1969) who currently works as a tax attorney. He reported that prior to his surgery he read extensively for his work and that, several years earlier, he scored in the top 1% on standardized reading comprehension tests used for law school admissions in the United States. He also indicated that his spelling had been comparable to that of other law school graduates.
In 2001, DPT experienced a single 3–4 minute episode of aphasia that led him to seek medical attention. He reported that during this episode of aphasia he could understand spoken words but lost the ability to speak, producing random words and “gibberish”. He was diagnosed with an oligodendroglioma in the left fusiform gyrus and underwent surgical resection of the tumor. He reported that immediately after surgery he had difficulties in spoken naming, reading comprehension, spelling and short-term memory. He returned to work one month after the surgery and has worked successfully since then, although he has continued to experience mild difficulties in reading, moderate difficulties in spelling and occasional difficulties in medium-term memory. With regard to reading, he specifically noted that since the surgery he has been able to read aloud easily but cannot always immediately understand the meaning of the words.
In spring 2006 (4 ½ years post-surgery) signs of regrowth of the tumor were detected, although there were no behavioral symptoms. As a result he underwent chemotherapy from 6/06 to 12/07 which successfully stopped tumor growth. The data from the experimental tasks reported on in this paper were collected between July 2005 and August 2007.
Structural MRI was carried out (9/05) and the images were registered to Talairach coordinates. The scans indicated that DPT’s resection lesion extended along the medial-lateral axis from −29 to −63, along the anterior-posterior axis from −15 to −66, and along the superior-inferior axis from −30 to −6. This places DPT’s lesion largely anterior to the lesion of the patient reported by Gaillard et al (2007) which extended rostro-caudally from y= −60 to −80. In terms of a gyral characterization, the lesion comprised a large part of the fusiform gyrus (including the whole mid-fusiform area claimed to be dedicated to orthographic processing) as well as some parts of the inferior temporal gyrus primarily mainly along the anterior and the lateral edges of the lesion (see figure 1).
Figure 1.
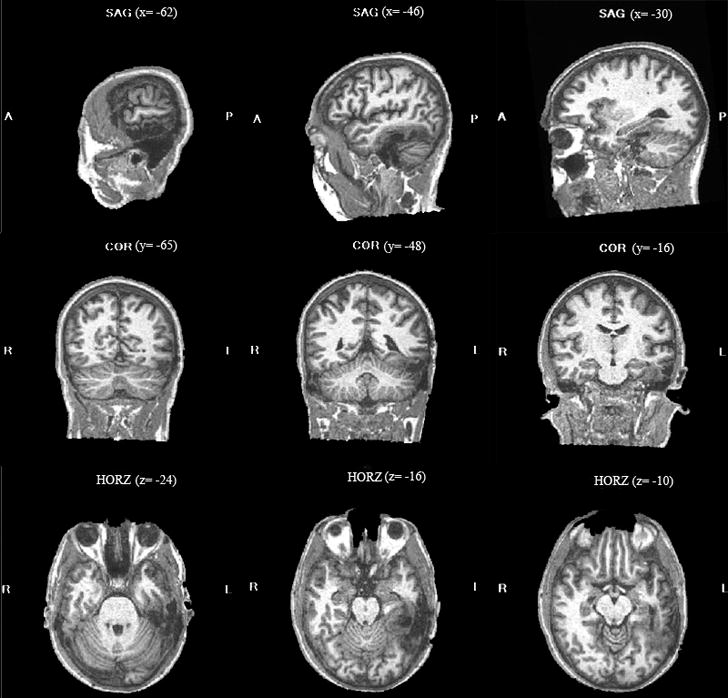
Sagittal, coronal and horizontal views of DPT’s lesion
A clinical neuropsychological evaluation was carried out 21 months after surgery (6/2003). It reported an MMSE score of 28/30 and normal or superior performance in virtually all cognitive areas that were evaluated. Specifically, as indicated in Table 1, DPT’s performance was normal to above normal for verbal working memory (Wechsler digits forward and backward), visual perception and memory (Rey Complex Figure-copy and delayed), fine motor speed and precision (Grooved pegboard; Trail Making), verbal learning (Rey Auditory Verbal Learning Test), spoken word naming and fluency (Boston naming, Word Association Test) and oral reading (American National Adult Reading Test), single word auditory comprehension (PPVT) and in recognition memory for words and faces (Warrington Recognition Memory Test. Performance was below normal only on the Stroop color-word task (15%ile) where he showed some slowness in naming the color of the ink in which color words were written.
Table 1.
DPT’s performance on a set of standardized tests: the Wechsler memory scale (Wechsler, 1987), the Rey Complex Figure Test (Osterrieth, 1993; Rey, 1993), the Grooved Pegboard Test (Bornstein, 1986), the Trail Making Test (Army Individual Test Battery, 1944), the Rey Auditory Verbal Learning Test (Rey, 1964), the Boston Naming Test (Kaplan, Goodglass, & Weintraub, 1983), the Warrington Recognition Memory Test for Words and Faces (Warrington, 1984), the Peabody Picture-Vocabulary Test (Dunn, 1959), the Controlled Oral Word Association Test (Benton et al., 1994), the American National Adult Reading Test (Blair & Spreen, 1989), the Stroop Test (Stroop, 1935).
| TASK | SUBTASK | DPT’S SCORE | CONTROL MEAN (SD) OR PERCENTILE |
|---|---|---|---|
| Wechsler memory scale (verbal working memory) | Information and Orientation | 13/14 | 13.7 (0.5) |
| Digits Forward | 16/16 | 8.7 (1.8) | |
| Digits Backwards | 11/14 | 6.8(2.1) | |
| Rey Complex Figure (visual perception & memory) | Copy | 36/36 | 31.75(3.21) |
| Delayed | 25.5/36 | 17.20(7.08) | |
| Grooved pegboard (fine motor speed & precision)Trail making Rey auditory verbal learning (verbal learning) | Dominant Hand | Time= 60, Error Rate=0 | percentile 53 |
| Nondominant Hand | Time=66, Error Rate=0 | percentile 50 | |
| Form A | 29sec | 29.9 (15.6) | |
| Form B | 62sec | 58.9 (22.1) | |
| Total | 54/75 | 53.6 (8.3) | |
| Interference Trial | 10/15 | 6.6 (2.1) | |
| Recall after Interference | 11/15 | 11.4 (2.4) | |
| Delayed Recall | 12/15 | 11.2 (2.8) | |
| Written Recognition | 13 | 13.6 (1.9) | |
| Boston Naming Test (spoken word naming) | Spontaneous | 60 correct | 55.9 (2.8) |
| Warrington’s Recognition Memory Test for Words and Faces | Words | 47/50 | percentile: 50 |
| Faces | 43/50 | percentile: 50 | |
| PPVT (Peabody Picture-Vocabulary Test) | Auditory word comprehension | 171/175 | percentile: 92 |
| Controlled Oral Word Asscociation Test (fluency) | F-A-S (Total # Words) | 50 | 40.5 (10.7) |
| American National Adult Reading Test (oral reading) | IQ Equivalent = 122.34 (Superior) | ||
| Stroop Test | Color | 112 | 111.94 (0.23) |
| Color-word | 93 | 104.90 (10.22) 15th percentile |
Control participants
Eleven control participants (6 men and 5 women) were recruited from the Johns Hopkins University community and were comparable to DPT with respect to age (age range 31–41) and years of education. (2 participants had BA degrees, 1 had a Ph.D. and the remaining 8 participants had MA degrees). It was not always possible to test all control participants on all tasks but there was a core group of seven to eight participants who served as controls for most tasks. Two participants had corrected-to-normal vision and none had any history of reading or spelling disorders. Normal spelling ability was verified for all participants by means of a spelling screener.
Methods
General testing procedures
All computer-based tasks were administered on the same Compaq Presario 2100 laptop, using E-prime 1.2.1 software (Psychological Software Tools, Pittsburgh, PA) for stimulus presentation and data collection. For all timed tasks, DPT and control participants were instructed to respond as quickly and accurately as possible. Unless noted otherwise, word frequency counts are from Francis and Kucera (1982).
Data Analysis
In order to limit response time variability, all reaction time analyses comparing DPT to control participants are based on median reaction times. DPT’s median RTs were statistically evaluated relative to those of control participants using the modified t-test and revised tests for dissociations proposed by Crawford and Garthwaite (2002, 2005). The “Crawford-t” is a quite stringent evaluation of differences between a single subject and a group. Because of this, in order to decide if DPT’s performance was abnormal, in the small number of cases where the p value of a Crawford-t was between .05 and .1 we compared DPT’s median RT to the control range of median RTs and we also considered DPT’s Z-score. Therefore, in addition to cases where the Crawford-t p value was below .05, we also considered his results to be significantly different from those of the control group if: the Crawford-t p value was between n .05 and .1, DPT’s median RT fell outside the normal range and his Z-score was greater than 1.5.
When individual subject effect sizes were statistically evaluated (e.g., the magnitude of frequency, regularity or priming effects), the analyses were based on mean (rather then median) RTs for DPT as well as individual control participants. This was, of course, necessary in order to obtain standard deviations for individual subjects.
DPT’s accuracy levels and error types on most tasks were comparable to those of control subjects and, therefore, although error rates are reported, error types are not described in any detail except where they differed from control participants. In addition, in tasks in which spoken response times were recorded, all participants made some voice-key errors, tripping the voice key prematurely due to coughs, false starts or other vocal sounds. The rate of voice key errors for controls subjects typically ranged from 1–13% and DPT’s range of 1–5% fell clearly within normal range and will not be discussed for each individual task. RT analyses do not include voice-key or actual errors.
Results
Section 1: Characterizing the orthographic impairments
In this section we present the results of a number of tasks used to evaluate DPT’s orthographic processing of words and nonwords in spelling, oral reading, and lexical decision and in two tests of comprehension of written words.
Briefly, the orthographic processing evaluation assumes fairly standard theories of reading and spelling (Coltheart, 1982; Ellis and Young, 1988) which assume a distinction between lexical and sublexical processes. We make no assumptions about whether the same or different lexical and sublexical processes are used in both reading and spelling. We assume that the spellings of familiar words are stored in long-term memory in what we refer to as an orthographic lexicon and that these long-term memory representations need to be accessed for the correct spelling of irregular words, as well as for their comprehension in reading. Sublexical processes that relate letters to sounds in reading and writing (grapheme-phoneme or phoneme-grapheme conversion processes, respectively) are necessary for the spelling and reading of unfamiliar (non)words. These sublexical processes are recruited when access to the orthographic lexicon fails and, when this occurs, plausible readings and spellings (e.g., ‘once” spelled as WUNS or read as “oans”) are produced..
Written Spelling
Task 1.1: Spelling words to dictation
DPT and 6 control participants were administered a list of 68 monomorphemic words for writing to dictation. There were no time restrictions and the first response was scored.
Control participants performed extremely well on this task (error range=0–3/68). In contrast, DPT made four times as many errors as the most errorful control subject (13/68). All of DPT’s errors were phonologically plausible. For example, “speak” was spelled as SPEEK, “type” --> TIPE, “toss” -> TAUSS.
DPT’s excellent repetition of the dictated stimuli, his ability to explain the meanings of words he did not spell correctly, and the fact that errors were phonologically plausible indicate an intact sublexical phoneme-grapheme conversion system, with errors originating from a deficit either within the orthographic lexicon or in gaining access to it (surface dysgraphia). To confirm this impairment locus, additional testing described below was carried out with DPT only.
Task 1.2: Spelling nonwords
DPT was administered 34 pseudowords (4 to 8 letters in length) from the JHU Dysgraphia Battery (Goodman, Caramazza, 1985) for spelling to dictation. DPT spelled 97% (33/34) of them correctly (see Table 2). This excellent performance is clearly consistent with an intact phoneme-grapheme conversion system which would be the source of the phonologically plausible spellings of words reported for List 1.
Table 2.
Spelling accuracy for DPT and age-matched control participants.
| Spelling accuracy | DPT’s accuracy | Control accuracy (range) |
|---|---|---|
| Words List 1 | 81% (55/68) | 96%–100% (0–3/68) |
| Nonwords List 2 | 97% (33/34) | |
| Words List 3 | ||
| Long words | 88% (15/17) | |
| Short words | 94% (16/17) | |
| Words List 4 | ||
| HF words | 98% (97/99) | |
| LF words | 80% (105/132) |
Tasks 1. 3 and 1.4: Spelling words: Evaluating effects of length and frequency
DPT was administered 17 long (7 and 8 letters) and 17 short (4 and 5 letters) frequency-matched words from the JHU Dysgraphia Battery Length List. The results indicated that there was no effect of length on his performance, with 88% accuracy (15/17) for short words and 94% (16/17) for the long words (x2 (1) = 0, p<0.1).
The effect of frequency was evaluated from data obtained by combining a number of word lists. A significant effect of frequency was found with high frequency words spelled significantly more accurately (98%, 97/99) than low frequency words (80% 105/132) (x2(1) = 15.9, p<.001). All errors but two (27/29) were phonologically plausible. It is interesting to note that, not infrequently, DPT produced multiple phonologically plausible spellings (“riot”-> RIAT, RIUT, RIOT; “elbow” -> ELBO, ELBOUGH; “gross” -> GROSE, GROAS).
In summary, the effect of frequency, the absence of a length effect, excellent pseudoword spelling and the production of phonological plausible spellings are the classical symptoms of an impairment to the orthographic lexicon (or in access to it) in the context of intact: sublexical processing, grapheme buffering and letter shape selection and production (see Tainturier and Rapp, 2001 for a review).
Reading
Task 1.5: Nonword reading
DPT and eleven control participants were asked to orally read 64 pronounceable nonword stimuli from Andrews and Scarratt (1998-Experiment 2). Each trial consisted of a fixation cross presented in the center of the computer monitor for 500 msec, followed by a single lowercase nonword that remained on the screen until the participant responded orally; response times were recorded by a voice key and the specific responses were recorded by the experimenter.
With regard to accuracy, DPT was highly accurate with 0 errors, a rate well within the control range of 0–3 errors. In terms of response times (as indicated in Table 3), DPT’s median RT of 759 msec. was not significantly different from the control group’s medians (Crawford’s t= 1.448, p = .178).
Table 3.
DPT and control participants’ performance on reading tasks. DPT’s performance is compared to that of the controls using the Crawford t-test (Crawford & Garthwaite, 2002)
| Task | DPT: median RT | Controls: mean (range; SD of medians) | Statistical comparison of DPT to controls |
|---|---|---|---|
| Nonword reading | 759 | 582 (465–804) (117) | ns |
| Word Reading | |||
| High frequency | 583 | 470 (388–596) (62) | ns |
| Low frequency | 615 | 477 (398–617) (65) | p<.1 |
| Regular | 588 | 475 (391–608) (64) | ns |
| Exception | 632 | 477 (393–614) (63) | p<.05 |
| Word Reading: Brief presentation | |||
| LVF | 668 | 429 (296–585) (105) | ns |
| RVF | 642 | 357 (301–426) (62) | p<.05 |
| High frequency | 602 | 351 (288–409) (53) | p<.05 |
| Low frequency | 738 | 405 (295–550) (105) | p<.05 |
| Short | 571 | 338 (263–411) (69) | p<.05 |
| Long | 743 | 417 (298–557) (99) | p<.05 |
| Visual Lexical Decision | |||
| High frequency | 675 | 572 (465–766) (91) | ns |
| Low frequency | 807 | 640 (537–809) (84) | p<.1 |
| Reg/consistent | 718 | 602 (490–767) (89) | ns |
| Reg/inconsistent | 712 | 574(535–801) (82) | ns |
| Strange | 727 | 597 (484–767) (86) | ns |
| Visual Synonym Judgment | 976 | 762 (564–965) (105) | p<.1 |
| Visual Semantic Priming | |||
| Overall lexical decision | 698 | 633 (486–880) (128) | ns |
| Priming effect: (unrelated-related) | 12 | 43 (29–73) (18) | p<.05 |
Task 1.6: Word reading: Evaluating effects of frequency and regularity
DPT and 11 control participants were asked to orally read single words from Jared (2002-Experiment 2). The list consisted of 160 monosyllabic words, half of high and half of low frequency (mean frequency of HF words=321; mean frequency of LF words=6) that were closely matched for neighborhood characteristics. Half of the words in each frequency group were exception words and half were regular-consistent words. A fixation cross appeared at the center of the monitor for 1000 msec, was followed by a blank screen for 500 msec, and then the stimulus word appeared until the participant responded orally triggering the voice key.
DPT made no reading errors on this task, clearly falling within the normal range of 0–4/160 errors. DPT’s median RTs for high frequency and regular words were no different from those of control participants (t=1.745, p=0.112 and t=1.69, p=0.122 respectively). However, his median RT for exception words was significantly slower than those of the control participants (t=2.280, p=.046). Furthermore, although his median RT of for low frequency words was not significantly different from those of controls (t=2.018, p=.07), we do consider it to be abnormal as the p value was less than .1, the Z-score was 1.7 and his median RT fell outside the control range of medians.
With regard to the magnitude of frequency and regularity effects, an evaluation of DPT’s mean RTs indicated a significant frequency effect that was comparable in magnitude to that exhibited by the control participants (Crawford’s t(11)=1.489, p=.167), who also showed significant frequency effects both as a group (t(10)=5.2, p=.000) and as individuals (range of p-values for controls= .022 to .000).
With regard to regularity, the control participants did not exhibit significant differences in their mean response times to exception versus regular words either as a group (t(10)=0.372, p= .718) or as individuals (p values ranged from .23 to .89). This contrasts with the significant regularity effects reported by Jared (2002) with these stimuli. On the other hand, DPT did exhibit a significant difference between mean response times for regular versus irregular words (t(146)=3.45, p=.0007). Furthermore, this difference was significantly larger than that of controls (Crawford’s RSDT: t(11)=2.872, p=.017)
In sum, despite the fact that DPT’s oral reading accuracy was comparable to that of the control participants, his reading times were significantly longer for low frequency and exception words. He exhibited significant effects of frequency and regularity, even though the control participants did not exhibit the latter. The significant regularity effect is likely to have resulted from his abnormally slow reading times for exception words. Thus, in clear contrast to DPT’s intact oral reading of nonwords, in his oral reading of words there was a significant slowing of responses, particularly for low frequency and exception words.
Task 1.7: Word reading with brief presentation: Evaluating the effect of visual fields, frequency and length
In this task DPT and 5 control participants were asked to read word stimuli presented randomly in the right or the left visual field. The beginning of each word in the right visual field (RVF) and the end of each word in the left visual field (LVF) were at 2.3 degrees of visual angle from the fixation point at the center of the monitor. Short words (4–5 letters) occupied 2.28 degrees of visual angle and long words (7–8 letters) 3.4 degrees. Each word was presented for 200 msec and the participant had to pronounce it as quickly and as accurately as possible. There were 240 stimuli, half of high and half of low frequency (mean frequency for HF words = 97, mean frequency for LF words = 2.2). In each frequency group, half of the words were short and half long. Long and short words were matched for frequency and high- and low-frequency words were matched for length.
DPT produced 17 errors, well within the control range of 11–45 errors. DPT produced more errors in the left visual field (LVF) than in the right (RVF) and more errors overall for low than high-frequency words (LVF: HF errors=3, LF errors= 8; RVF: HF errors=2, LF errors = 4).
With regard to frequency, DPT was significantly slower than control participants both for high (Crawford’s t=3.9, p=.012) and low-frequency words (t=2.895, p=.044). Nonetheless, the difference between high- and low-frequency words for DPT was not significantly different than that of controls (Crawford’s RSDT: t (4) = 0.71, p=.95).
With regard to effects of visual fields, for the stimuli presented in the LVF, DPT’s median RT of 668 msec, was not significantly different than controls (Crawford’s t=2.078, p=.106). However, for stimuli presented in the RVF, DPT’s median RT of 642 msec was significantly slower than controls(Crawford’s t=4.196, p=.014). Consistent with what is commonly reported in the literature (e.g., Brysbaert et al., 1996; Ellis, 2004; Whitney and Lavidor, 2004), control participants exhibited a RVF advantage. This RVF advantage was significantly smaller for DPT in comparison to controls (Crawford’s RSDT: t (4)= 2.682, p=.055). Presumably this is due to the fact that DPT’s response times were more abnormal with RVF versus LVF stimuli.
Finally, with regard to word length we found that DPT’s median RTs were significantly slower than those of controls for both short and long words (Crawford’s t=3.083, p=.037 and t=3.006, p=.04). However, the effect of word length was not significantly different from that of controls (Crawford’s RSDT: t(4)= 0.122, p=.91). The length effect was significant for DPT and all individual control participants (DPT: t(212)= 6, p=.0000; p-value range for controls: .01 to .0000).
In sum, in this oral reading task with brief stimulus presentation we see high accuracy but abnormally slow responses in all categories, although especially for stimuli presented in the RVF. The magnitude of DPT’s frequency and length effects both fall within the normal range; however, his RVF advantage is significantly smaller than that of control participants. The fact that DPT does not exhibit an abnormal length effect is important as it indicates that, unlike a number of other cases of individuals with damage to the fusiform and/or nearby tissue, DPT is not a letter-by-letter reader, we return to this point in the General Discussion.
Task 1.8: Visual Lexical Decision: Evaluating effects of frequency and regularity
DPT and 10 control participants performed a visual lexical decision task with stimuli from Seidenberg, Waters, Barnes, and Tanenhaus (1984-Experiment 4). Following a central fixation cross presented for 300 msec, each stimulus was presented in the center of the computer monitor until the participant made a key response to indicate whether or not the stimulus was a real word of English. There were a total of 90 words, half of high and half of low frequency (mean frequency for HF words =319; for LF words: 11). In each frequency category, there were three levels of regularity with 15 regular-consistent words, 15 regular-inconsistent words and 15 strange words. There were 90 pronounceable nonwords.
DPT’s overall error rate of 5/180 fell within the control range of 3–6/180. As indicated in Table 3, DPT’s median RTs for words were not significantly different than those of the control participants on any of the sublists. However, it is worth mentioning that his median RT of 807 to low frequency words, although within control range, yielded a p value between .05 and .1 (Crawford’s t=1.709, p=0.89) and a somewhat elevated Z-score of 1.61.
With regard to frequency effects, DPT and all individual control participants showed significant effects of frequency (DPT: t(83)=2.78, p=.0085, controls: p values .001–045). In addition, the magnitude of DPT’s frequency effect was significantly greater than that of the controls (Crawford’s t=2.42, p=.019). With regard to regularity, although Seidenberg et al. (1984) reported a significant difference between response times for strange versus regular words, we did not find a significant effect for either DPT or for the control participants (except for one). Furthermore, DPT’s difference score for strange versus regular words was not significantly different from that of controls (Crawford’s t=1.27 p=.235). With regard to nonwords, DPT’s median RT of 868 for nonwords was significantly different from control participants (Crawford’s t=2.346, p=.044).
In brief, DPT’s performance in the lexical decision task was generally comparable to that of controls’ both with respect to error rates and RTs, and with regard to the presence/absence of frequency and regularity effects. There was, however, some indication of slowed responses to nonwords and to low frequency words, the latter most likely served to accentuate the effect of frequency, producing a significantly larger frequency effect for DPT as compared to controls.
Arguably, the oral reading and lexical decision tasks reported on above do not necessarily require access to semantics from orthography. In order to examine this process specifically we carried out tasks requiring explicit (Task 1.9) or implicit (Task 1.10) semantic access.
Task 1.9: Written word comprehension: Synonym judgment task
DPT and 10 control participants performed the synonym judgment task from the Johns Hopkins University Dyslexia Battery which consists of 54 pairs of high frequency words (mean frequency =154), half of which are synonyms and half are not. Each pair of words was presented simultaneously on the computer monitor and participants were instructed to decide as quickly and as accurately as possible whether the words were related or not by pressing one of two responses keys,. The stimuli were visible until the participant responded.
This was the only reading task on which DPT performed outside the control range with respect to accuracy. His error rate of 5/54 fell just outside the control range of 1–4/54 errors. In addition, his response times were abnormal with his median RT of 979 ms. falling outside the normal range (564–965 ms), corresponding to a p value of .08 (t(10)=1.9) and a Z score of 1.64
His abnormal error rate and slowed reaction times on this task indicate specific difficulties in accessing meaning from print; this possibility was examined further in the following task.
Task 1.10: Semantic priming with orthographic stimuli
Semantic priming tasks that involve presenting two semantically related or unrelated stimuli in close temporal succession allow for an evaluation of the time course of access to meaning from print, as the prime word must access meaning with sufficient speed so as to influence the processing of the target stimulus that follows it closely in time.
DPT and 7 control participants made lexical decision judgments on the second stimulus of a pair of written stimuli. There were equal numbers (120) of word/nonword and word/word stimulus pairs. The first word of each pair (appeared in lower case) and remained on the screen for 200msec and was immediately followed by the second stimulus and on which the lexical decision was made. The lexical decision stimulus appeared in upper case and remained on the screen until the participant pressed either of the ‘yes’ or ‘no’ designated response keys. There were 40 pairs of semantically related words and 40 pairs of unrelated words that were matched for frequency, length, and regularity. The experiment included another 40 word/word filler pairs. The word primes were of the same frequency and length for both word/nonword and word/word pairs. Within pairs, word primes and targets were matched for frequency (mean frequency of primes=65; targets=67).
DPT’s overall lexical decision accuracy (5/240 errors) fell within the control range (2–14/240 errors). In addition, his median lexical decision RT of 698 msec (both words and nonwords included) did not differ from those of controls (Crawford’s t=.47, p=.649). This excellent lexical decision performance was not surprising since DPT was also no different from controls in the lexical decision task reported above, especially for words in the high frequency range.
Crucially, however, with regard to priming, we found that DPT did not show significant facilitation when mean lexical decision times for the unrelated and the related pairs were compared (t(74)=.528, p=.6). In contrast, all individual control participants exhibited significant semantic priming effects in this task (controls’ range of p values: 0004-.032).
In DPT’s case, the absence of semantic priming in the face of normal lexical decision times to target words and nonwords is precisely what would be predicted if there were slowed access to semantics. That is, if access to semantics is abnormally slow for the prime word, then we would not expect to see facilitation of processing (faster lexical decision times) for the target.
Section Summary: Orthographic processing deficits
The evaluation of DPT’s orthographic skills reveals the following: (1) With regard to spelling, DPT shows clear impairment in the lexical route in spelling, presumably arising at the level of the orthographic lexicon or in access to it, with sparing of the sublexical process. This conclusion is supported by his intact auditory comprehension of single words, his accurate spelling of nonwords, a significant effect of frequency in word spelling and the production of phonologically plausible errors in his spelling of irregular words; (2) In reading, DPT appears also to suffer a deficit to the lexical system, leaving intact his sublexical and prelexical processing. His oral reading of nonwords is apparently entirely normal indicating that letter processing, orthographic analysis, grapheme-to-phoneme conversion and general phonological output processes are intact; (3) Unlike spelling, the reading deficit does not generate abnormal error rates or regularization errors, instead reading difficulties are manifested in slowed reading times, especially for low frequency and irregular words. This pattern is consistent with damage somewhere along the lexical route for reading: in accessing the orthographic lexicon and/or in semantic access from the orthographic lexicon. The abnormal synonym judgment times and especially the absence of a semantic priming effect for high frequency words, is strongly indicative of slowed access to semantics from orthographic input. We take up the issue regarding the relationship between the reading and spelling deficits in the General Discussion.
These findings while clearly indicating disruption to orthographic processing do not address the question of whether or not the lexical deficits in reading and spelling -in going from semantics to orthography (spelling) and in going from orthography to semantics (reading)- are modality and category specific. We take up these questions in the next two sections.
Section 2: Modality specificity?
Task 2.1: Spoken picture naming
Is the difficulty observed in producing written words in spelling limited to the written modality or is it a more general lexical retrieval deficit (see Hillis et al., 2005)? This question was evaluated by examining DPT’s accuracy and response times in a task of spoken picture naming.
DPT and 6 control participants were administered line drawings for spoken naming from Rossion and Pourtois (2004)’s color and texture adaptation of the Snodgrass and Vanderwart (1980) line drawings. Drawings were presented individually on the computer monitor until the participant responded.
Responses were scored as correct or incorrect based on published norms (Snodgrass and Vanderwart, 1980; Bates et al., 2003; Rossion and Pourtois, 2004). On this basis, DPT made 5/260 naming errors, well within the control range of 2–19/260. Similarly, DPT’s median RT of 977 was not significantly different from those of controls (Crawford’s t=1.844, p=.13).
In order to evaluate his naming performance in a more detailed manner we considered a subset of 50 high- and 50 low-frequency words (mean frequency for HF=102, for LF=2.2) whose images were matched for visual complexity according to the visual complexity ratings provided by Snodgrass and Vanderwart (1980) and updated by Rossion and Pourtois (2004) (mean visual complexity = 2.9 for HF and 2.7 for LF). Furthermore, we did not include any low-frequency words whose names were compound words. Again (as indicated in Table 5), DPT’s median RTs were not significantly different from those of controls for high or low frequency items. Furthermore, the magnitude of DPT’s frequency effect was comparable to the magnitude of the frequency effects produced by controls (Crawford’s t= 1.2. p=.29).
Table 5.
DPT and control participants’ performance on spoken language tasks. DPT’s performance is compared to that of the controls using the Crawford t-test (Crawford & Garthwaite, 2002)
| TASK | DPT’s median RT | Controls mean (range; SD of medians) | Statistical comparison of DPT to controls |
|---|---|---|---|
| Spoken Picture Naming | |||
| High frequency | 822 | 687 (579–881; 116) | ns |
| Low frequency | 1035 | 788 (683–932; 128) | ns |
| High complexity | 1098 | 806 (681–915; 122) | ns |
| Low complexity | 1140 | 817 (654–1059; 164) | ns |
| Auditory synonym judgment | 1382 | 1036 (719–1456; 248) | ns |
| Auditory semantic priming | |||
| Overall Lexical Decision | 1183 | 1101 (964–1363; 150) | ns |
| Priming effect: (unrelated-related) | 169 | 200 (152–283; 46) | ns |
The fact that DPT’s spoken word naming accuracy and response times do not differ from those of control participants, not only indicates that DPT’s written language deficits may be modality specific (affecting written vs. spoken language responses) but also that they may be category specific (affecting written but not picture stimuli).
Although we take up the question of category specificity more directly in the next section, in the data analysis for this task we did examine whether there was any effect of the visual complexity of the picture stimuli on DPT’s naming times. We identified two sets of items of 40 items that differed in visual complexity as established by Snodgrass and Vanderwart (1980) and updated by Rossion and Pourtois (2004). The low complexity set had an average complexity rating of 1.7 (on a scale from 1 to 5) and the high complexity set’s average complexity rating was 3.7. The two sets were matched for name frequency (High Complexity (HC), name frequency = 26, Low Complexity (LC), name frequency = 23). As indicated in Table 5, DPT’s naming latencies did not differ from those of normal controls for either set (HC: Crawford’s t= 1.64, p=.162; LC: Crawford’s t=1.438, p=.21). Furthermore, DPT’s naming time difference of 42 msec for high versus low complexity stimuli did not differ from those of controls (Crawford’s t=.27, p=.71). When we compared HC versus LC items within each individual participant, DPT showed no visual complexity effect (t(80)=.44, p=.66); this results was comparable to controls (except for one) who also showed no visual complexity effects (range of p values for controls: 0.044 to 0.86).
From these results we can conclude: (1) the significant problems in orthographic lexical retrieval (manifested in word spelling) did not extend to lexical retrieval in spoken word production and (2) the significant slowing in oral reading was unlikely to have been caused by a primary difficulty in accessing the phonological forms of words.
Task 2.2: Auditory synonym judgment task
In order to further evaluate the question of modality specificity we examined DPT’s auditory comprehension with tasks similar to the reading comprehension tasks described earlier (Tasks 1.9 and 1.10).
DPT and the same 10 control participants who participated in Task 1.9, were administered the synonym judgment task (Task #49) from the PALPA (Kay et al., 1992). This consisted of 60 pairs of high frequency words (mean=48), half of which were synonyms and half were not. Stimulus presentation was as follows: a fixation point for 500msec, a pause for 300msec, followed immediately by a pair of related or unrelated words with 100msec silence before and after each stimulus. The participants were instructed to decide as quickly and as accurately as possible whether the pair of words were semantically related or not by pressing one of two response keys. Response times were recorded from the onset of the second stimulus in the pair.
DPT’s error rate of 1/60 was within the control range of 0–3/60 errors. Moreover, and in striking contrast to his performance with written stimuli in Task #9, his median RT of 1382 was not significantly different from those of the control participants (Crawford’s t=1.33, p=.216).
Task 2.3: Semantic priming in the auditory modality
Five of the six control participants that previously participated in the visual-visual semantic priming Task 1.10 as well as one additional control subject participated in this experiment. Participants made lexical decision judgments on the second stimulus of either word/nonword or word/word stimulus pairs that were presented sequentially in the auditory modality. The items were comparable to those used in the visual-visual experiment (i.e., they were matched for prime and target frequency, length (number of letters and syllables) and imageability as shown in Table 4. There were 40 pairs of semantically-related words and 40 pairs of unrelated words; the two sets were matched for frequency, length, and regularity. The experiment also included another 40 word/word filler pairs and 120 word/nonword pairs where the word prime was of the same frequency and length as the word prime of the word/word pairs. The mean frequencies of the primes and targets were comparable (mean frequency of primes= 68; targets=76). Stimulus presentation was as follows: a fixation cross for 500msec, the prime word, 200msec of silence immediately followed by the target word or nonword, then another 100msec of silence. The participant had to make a lexical decision by pressing the designated ‘yes’ or ‘no’ keys.
Table 4.
Characteristics for each stimulus category for the visual and auditory semantic priming experiments (Studies 1.10 and 2.3). Each cell reports the mean value and (standard deviation). Word frequencies and imageability ratings are from MRC database (Coltheart, 1981) in which the frequencies are based on Kucera and Francis (1967). and imageability ratings on Pavio et al. (1968)
| MODALITY | STIMULUS CHARACTERISTICS | RELATED PRIME | RELATED TARGET | UNRELATED PRIME | |
|---|---|---|---|---|---|
| Visual | Word frequency | 72 (82) | 68 (77) | 70 (79) | 71 (63) |
| Imageability | 515 (82) | 531 (80) | 516 (61) | 497 (39) | |
| # of letters | 4.57 (1.1) | 4.55 (0.9) | 4.23 (0.9) | 4.55 (1) | |
| # of syllables | 1.35 (0.5) | 1.15 (0.4) | 1.13 (0.3) | 1.275 (0.5) | |
| Auditory | Word Frequency | 73 (77) | 74 (85) | 71 (63) | 75 (56) |
| Imageability | 539 (72) | 522 (98) | 521 (40) | 507 (44) | |
| # of letters | 4.52 (1) | 4.12 (1) | 4.7 (1) | 4.7 (0.9) | |
| # of syllables | 1.27 (0.5) | 1.1 (0.3) | 1.375 (0.5) | 1.3 (0.5) |
DPT’s lexical decision accuracy and median RT did not differ from those of controls. In fact, DPT was more accurate than controls with 0/240 errors (control range = 1–13/240) and his median lexical decision RT of 1183 msec (both words and nonwords included) did not differ from those of controls (Crawford’s t=.65, p=.544). When we examined the magnitude of priming effect for the unrelated compared to the related pairs, DPT showed a significant priming effect (t(75)= −4.5, p=.0008) as did all individual control participants (range of p=.00001-.0003). Furthermore, with regard to the priming effect size, DPT’s priming effect was well within the control range of effect sizes 152–283 and did not differ significantly from them (Crawford’s t=−0.6, p=.572).
Section Summary-Modality Specificity?
The results are quite clear with regard to the question of modality specificity. In contrast to his performance in the written modality, DPT’s performance in spoken naming and in tasks requiring rapid access to the meaning of auditorily presented words was no different from that of normal control participants. Furthermore, the finding of normal semantic priming in the auditory modality rules out the possibility that a semantic deficit contributed to DPT’s difficulties in reading comprehension.
Section 3: Category specificity?
In this section we report a series of experiments that examine the issue of whether or not DPT’s difficulties in reading were limited to the visual category of orthographic stimuli or if they more broadly affected visual object processing. We did so by examining DPT’s ability to extract meaning from faces and objects.
Task 3.1: Faces: Fame judgment
DPT and 7 control participants were shown a face on a computer monitor and were asked to respond as quickly and accurately as possible if the face corresponded to a famous person or not by pressing one of two response keys. The face was visible until the participant responded. The task consisted of a total of 210 faces, half of which were famous and half of which were not1. The famous individuals had professions in the following categories: sports, politics, business and entertainment.
DPT’s performance was no different from that of control participants with respect to accuracy and median RT (see Table 6). His error rate of 17/210 was well within the control range of 8–31/210 and his median RT of 1183 was no different from that of controls (Crawford’s t=.98, p=.4).
Table 6.
DPT and control participants’ performance on face and object processing tasks.. DPT’s performance is compared to that of the controls using the Crawford t-test (Crawford & Garthwaite, 2002)
| TASK | DPT: median RT | Control mean (range; SD of medians) | Statistical comparison of DPT to controls |
|---|---|---|---|
| Faces: fame judgment | 1183 | 938 (772–1434; 235) | ns |
| Faces: forced choice categorization | 838 | 823 (523–1123; 163) | ns |
| Pyramid and Palm Trees (timed) | 2269 | 1765 (1147–2565; 504) | ns |
Task 3.2: Faces: Forced choice categorization of profession
DPT and 8 control participants were tested with the same photos used in the previous task. A face appeared on the computer monitor and the participant was instructed to choose from two categories the profession that corresponded to the target face (on different blocks of trials the choices were: sports-politics, politics-entertainment, sports-entertainment, business-sports, business-politics). Stimuli remained on the monitor until the participant responded. There were a total of 266 forced choice trials.
DPT’s error rate of 3/266 was lower than that of the controls, whose error rates ranged from 5–25/266. His median RT of 838 msec was not different from those of the control participants (Crawford’s t=−0.38, p=.79).
Task 3.3: Objects - Pyramids and Palm Trees (timed)
To examine DPT’s ability to access semantic information for pictured objects we used a timed, computerized adaptation of the Pyramids and Palm Trees task (Howard and Patterson, 1992).
DPT and 8 control participants were presented with 55 stimuli; each consisting of a triad of line drawings presented on a computer monitor—one displayed above the other two. Participants were asked to decide as quickly and as accurately as possible which of the two pictures presented at the lower part of the monitor was related in meaning to the picture shown in the upper part. Stimuli remained on the monitor until participants responded with a button press (see Table 6).
DPT’s error rate of 6/55 fell well within the control range of 3–15/55 and his median RT of 2269 was no different from that of control participants (Crawford’s t=.94 p=.38).
Section Summary: Category Specificity?
DPT showed no signs of difficulty in processing non-orthographic categories of visual stimuli such as line-drawings of objects and photographs of faces. These findings are also consistent with the normal effects of visual complexity observed for the picture naming task reported in Section 2. These results clearly indicate that the orthographic difficulties that we have documented are not part of a more general deficit in processing visual stimuli. These findings reveal that DPT can access semantics normally from visual input as long as the stimuli are not orthographic.
General Discussion
In the present study we investigated the cognitive functions subserved by the left fusiform gyrus through the detailed investigation of the performance of an individual (DPT) who had undergone a resection of the mid and anterior portions of his left fusiform gyrus (as well as adjacent regions of the inferior temporal gyrus). We were interested in evaluating the hypothesis that there are neural substrates in the left fusiform that are specifically necessary for the processing of orthographic information, and not language nor visual categories more generally. That is, we were specifically interested in the claim regarding the modality and category specificity of substrates within the left fusiform. Our investigation revealed the following: (1) DPT suffered deficits in orthographic processing that affected both the reading and spelling of words (leaving intact the processing of nonwords) and, more specifically, the deficit/s affected the retrieval of meaning from orthographic forms (in reading) and the retrieval of orthographic forms from meaning (in spelling). (2) The modality specificity of the deficit was supported by the finding that, in contrast to his difficulties in generating written word forms in spelling, DPT was able to generate spoken word forms with normal speed and accuracy in picture naming; and, furthermore, that, in contrast to his difficulties in extracting meanings from written forms in reading, DPT was able to extract meaning from spoken forms with normal speed and accuracy in auditory word comprehension tasks. (3) The category specificity of the deficit was supported by the finding that DPT showed normal speed and accuracy in his processing of photographs of faces and line drawings of objects, and was not affected abnormally by the visual complexity of the stimuli. This constellation of results clearly supports the conclusion that at least some portion of the mid-anterior left fusiform is specifically necessary for the normal processing of lexical orthographic information in reading and spelling.
Functional localization of the deficits in reading and spelling
In spelling, the characterization of the deficit locus is quite straightforward. DPT’s entirely normal nonword spelling, with difficulties restricted to the spelling of words with unpredictable spellings (in the face of excellent auditory word comprehension) and the production of phonologically plausible responses (e.g., ‘riot” -> RIAT) clearly point to a specific difficulty in accessing the long-term memory representations of the spellings of words from intact representations of word meanings.
With regard to reading, the fact that DPTs was able to read nonwords with normal accuracy and response times, provides clear evidence that early visual and letter recognition processes were intact and, therefore, that the reading deficit affected some aspect of lexical processing. The fact that auditory semantic priming was normal indicates that semantic representation and processing were intact, narrowing the possible deficit locus to some aspect of processing the orthographic word forms themselves or in accessing their meaning. The visual semantic priming task (Task 1.10) is the most revealing with regard to characterizing the specific nature of DPT’s reading deficit. His normal lexical decision times and accuracy for the target words in this task stand in contrast to the absence of significant semantic priming effects in the same task. The normal lexical decision times indicate intact processing through the lexical orthographic level (at least for high frequency words which make up the stimuli in this task) and given this, the absence of semantic priming indicates a failure to activate word meanings in a normal time frame. It is this within-task dissociation and the contrasting results in the orthographic and spoken modalities that provide the clearest evidence of a deficit specifically affecting access to semantics from orthography. This conclusion finds additional support in DPT’s elevated error rate and response times in the written synonym judgment task (Task 1.9). The conclusion that there is a deficit affecting access to meaning from print is strongly supported by the evidence, however, it is important to note that we cannot rule out some additional disruption to lexical orthographic processing itself, given the signs of slowed lexical decision times for low frequency words (Task 1.8).
As we have noted, the spelling and reading deficits have considerable functional “symmetry”. They differ, however, in terms of their impact on errors. That is, while the lexical deficit in spelling resulted in frank phonologically plausible errors, the reading deficit manifested itself largely in slowed reaction times, rather than elevated error rates and regularization errors. One may wonder how oral reading and reading comprehension are so accurate. This prompts the question: What exactly is the nature of the functional deficit such that it only slows access from orthography to meaning but does not eliminate it nor does it result in regularization errors? There is considerable evidence that normal reading (and spelling) involves the interaction between and integration of information from both lexical and sublexical processes (for reading: Hillis and Caramazza, 1991; Miceli et al, 1994; Plaut et al., 1996; for spelling: Folk and Rapp, 2004; Rapp, et al., 2002). Some of these investigations reveal that errors in word reading can be reduced when lexical-sublexical interaction is available. Thus, one possibility is that we are observing the consequences of a disrupted lexical system functioning in the context of the support provided an intact sublexical system and even perhaps an intact non-semantic lexical process (Schwartz et al., 1980)2. Without additional, extensive testing it is not possible to more precisely characterize the nature of the deficit that affects speed and efficiency of access to meaning in reading while leaving accuracy intact.
The relationship between reading and spelling
The fact that DPT’s performance in reading and written spelling tasks is strikingly similar and “symmetrical” quite naturally raises questions regarding the functional and neural relationship between reading and spelling. In both reading and spelling, DPT exhibited intact processing of nonwords, the absence of any length effect, and lexical deficits were indicated by frequency effects (in reading: Tasks 1.6 and 1.8; spelling: Tasks 1.3 and 1.4), as well as by effects of regularity. The latter were manifested in reading in his increased response latencies to irregular words (Task 1.6) and, in spelling, by the production of phonologically plausible spellings (Tasks 1.1, 1.3 and 1.4). The question arises: Do these similarities indicate that the comprehension and production of orthography share one or more processing components that suffered damage in DPT’s case?
According to most theories of word processing, the reading and spelling of words make use of a lexical semantic system that is also shared with spoken word processing. Semantic mediation is typically considered to be critical at least for the correct spelling and reading of words with non-predictable spellings and pronunciations. As a result, neural damage affecting the lexical semantic system can be expected to affect both reading and spelling of low-frequency, exception words; furthermore, in the presence of intact sublexical processing, phonologically plausible pronunciations and spellings should be produced. This pattern of impaired word comprehension accompanied by surface dyslexia and dysgraphia has been reported either subsequent to stroke (Hillis and Caramazza, 1991), or to trauma (Tainturier, 1996) and in context of semantic dementia (Graham et al., 2000). However, since DPT has intact lexical semantics (as evidenced by normal auditory semantic priming, synonym judgment, etc.) this cannot be the basis of the association between his reading spelling deficits. This, therefore, raises the possibility that reading and spelling may share orthography-specific processing structures.
The notion that reading and spelling share orthography-specific processing components/neural substrates is consistent with DPT’s performance pattern as well as with other cases exhibiting both acquired dysgraphia and dyslexia subsequent to lesions affecting the left inferior temporal lobe (Rapcsak and Beeson, 2004). As striking as the similarities are across reading and spelling, Hillis and Rapp (2004) and also Tainturier and Rapp (2001) discuss the complexities involved in interpreting patterns of association and dissociation in reading and spelling performance. They caution that it is difficult to conclude that a striking association of deficits reflects damage to a shared process, because the alternative conclusion that similar processes are neurally continguous cannot be ruled out. While this is undeniable, it is also important to consider that recent functional neuroimaging studies have also indicated the involvement of common substrates for reading and spelling within this left-mid fusiform region (Rapp et al., 2006; see also Beeson et al., 2003). Nonetheless, it is possible that these functional neuroimaging findings do not provide sufficient resolution to distinguish independent but contiguous processes. In sum, although not entirely unambiguous, there is a convergence of the functional neuroimaging and neuropsychological findings on the conclusion that at least some shared orthography-specific processes/processing resources for reading and spelling may be subserved by the left-fusiform. However, the specific computational function of these substrates has not been identified. In the case at hand, it is interesting to consider whether or not a single functional deficit that reduced the speed or efficiency of processing between lexical semantic and orthographic representations could produce the observed asymmetrical consequences in reading and spelling. Without a better understanding of these systems and the timing of lexical and sublexical processes, it is not clear why a reduction in lexical processing speed would allow for correct retrieval of pronunciations in reading, but yield phonologically plausible responses in spelling.
The neural substrates of orthographic processing
Constraints on the interpretation of DPT’s lesion-deficit pattern are provided both by cases of semantic dementia–with lesions typically anterior to DPT’s- as well as by cases such as the one by Gaillard et al. (2006)–whose lesion was posterior to DPT’s.
A number of studies have indicated that the most commonly affected region in semantic dementia is the left anterior temporal lobe (with atrophy also reported in lateral and anterior areas of the middle and inferior temporal cortex (Chan et al., 2001; Davies et al., 2005; Mummery et al., 2000; Mummery et al., 1999). The fact that DPT did not have a semantic deficit and that his lesion was situated posterior to these anterior temporal regions associated with semantic processing, provides support for the hypothesis that orthography-specific functions required for the normal mediation between lexical semantics and orthography are situated in the region of the mid fusiform gyrus that was lesioned in DPT’s case.3
As indicated earlier, DPT’s lesion is largely anterior to that of the individual described by Gaillard et al., extending (in Talaraich space) from −15 to −66 while Gaillard et al.’s patient’s lesion extended from −60 to −80. Both the similarities and differences in their cognitive abilities and deficits provide important information regarding the nature of orthographic functions and their neural substrates.
With regard to similarities, it is important to note that both individuals exhibited normal processing of stimuli from visual categories such as objects and faces. Furthermore, they both exhibited normal performance in spoken language production and comprehension. The differences between them lie in the nature of their orthographic deficits. The first and most clearly documented difference concerns the effect of word length in reading. Gaillard et al. presented clear evidence that their patient’s overall reading times significantly increased from pre to post surgery, and that, after surgery, he exhibited a significant effect of length on his reading times that was absent prior to the surgery; furthermore, when stimuli were flashed briefly, his reading accuracy decreased significantly from pre to post surgery (for other relevant cases see Cohen et al., 2003; Damasio and Damasio, 1983; Henry et al., 2005; Leff et al., 2001). In contrast, although DPT’s overall reading time were slower than those of controls, he showed no abnormality in the magnitude of his length effect. That is, the difference in response times for long and short words was no greater for DPT than it was for control subjects. Also consistent with an absence of length effects (or letter-by-letter reading behavior) is the fact that DPT’s reading times for pseudowords were no different from those of control subjects4.
Abnormal length effects in reading are typically interpreted as indicating difficulties in the parallel processing of the orthographic stimulus itself. Consistent with this interpretation, Gaillard et al. conclude that their patient suffered a deafferentation of the orthographic processing system from visual input. DPT’s deficit clearly affects a later reading process, one involved in the processing of lexical orthographic forms after their constituent letters have been identified in a normal manner. As indicated above, DPT’s reading deficit is best characterized as one affecting his access to word meaning after relatively intact processing of orthographic word forms. Importantly, the distinction between the deficits exhibited by these two individuals and the differences in the location of their lesions, is generally consistent with the notion that the left fusiform, in its posterior-anterior extent, is hierarchically organized to carry out increasingly more “abstract” orthographic functions (Cohen, et al., 2003; Vinckier et al., 2007; Binder et al., 2006). Under this interpretation, the Gaillard et al. subject reveals the role of more posterior areas of the left fusiform in processing orthographic stimuli, while DPT’s performance sheds light on the role of mid-fusiform areas specifically in word processing and in access to meaning from print.
Another difference between the two individuals concerns their spelling performance. While DPT clearly suffered from a deficit affecting his ability to retrieve the stored orthographic forms of words, Gaillard et al.’s patient’s spelling was flawless for those items he was asked to spell. However, a limitation in interpreting this difference is that Gaillard et al.’s patient was not extensively tested in spelling to dictation (36 high frequency words, 3–9 letters in length). Without data on irregular and low frequency words it is difficult to draw firm conclusions. Nonetheless, the difference between the two patients with regard to their spelling abilities is certainly consistent with the conclusion that processes/representations involved in accessing abstract orthographic word forms from long-term memory (a function referred to as the orthographic lexicon by many researchers) are located more anteriorly along the left fusiform.
Modality specific processing in the left fusiform?
As we discussed in the Introduction, most researchers have posited amodal/polymodal processing at some location within the left fusiform. While some have argued that there are no orthography-specific processes whatsoever within the fusiform, and that, instead, all non-visual processing is polymodal (Booth et al., 2002; Buchel et al., 1998; Price et al., 2003; Mechelli et al., 2005; Mummery et al., 1998; Noppeney and Price, 2002), others have argued that polymodal processing is limited to more anterior regions of the fusiform (Cohen et al., 2004; Jobard et al., 2003, 2007). DPT’s lesion extended from −15 to −66 in the anterior-posterior dimension, occupying regions that have been associated by virtually all investigators with polymodal/spoken language processing. Interestingly, however, DPT performed normally in terms of response times, accuracy and magnitude of semantic priming in the auditory modality (Tasks 2.2 and 2.3) and the semantic priming task is a particularly rigorous measure of the integrity of auditory word processing. In addition, with regard to spoken production, although DPT did report spoken naming difficulties post-surgery, his picture naming response times and accuracy were no different from those of normal subjects.
How can we account for the fact that DPT’s performance would seem to be at odds with rather widespread claims of (anterior) fusiform involvement in auditory/spoken word processing? One possibility is that, the critical polymodal fusiform areas are more anterior to the anterior edge of his lesion. That is, fusiform gyrus extends to approximately y=0 in Talairach coordinates, whereas the anterior edge of DPT’s lesion was at approximately y=−15. Although this very anterior involvement in polymodal/spoken language processing is a possibility, this would seem to be clearly more anterior than has been reported in a number of studies which typically report activation centered around y=−40 (Jobard et al., 2007; Mechelli et al., 2003, 2005; Mummery et al., 1998; Price et al., 2003). A second possibility is that although, in functional neuroimaging studies, these anterior areas (centered on y-40) have been reported as active in polymodal/spoken language contexts, they are not strictly necessary for performing these functions. Possibly other regions (such as superior temporal, or posterior superior temporal areas) play a more critical role in access to (and from) semantics for spoken language. Consistent with this, in the relevant functional neuroimaging studies, activation associated with polymodal/spoken language processing is typically reported in multiple areas outside the fusiform. However, this still leaves to be accounted for the reports of lesion-deficit correlation or perfusion studies reporting an association of spoken naming deficits with fusiform lesions (Hillis, et al., 2005; Price et al., 2003; Raymer et al., 1999). However, if we look at these cases we find that although the lesions/hypoperfusion affected the left (and sometimes anterior) fusiform gyrus, they also typically affected other inferior temporal or even middle to superior temporal areas as well. Therefore, it is difficult to attribute the spoken naming deficits specifically to fusiform or anterior fusiform areas.
It is apparent that, in order to resolve these issues, functional neuroimaging and lesion studies that can specifically isolate anterior portions of the left fusiform will be required.
Category-specific orthographic processing in the left fusiform?
DPT’s performance is consistent with a number of neuropsychological reports indicating dissociations in performance across the visual categories of written words, face and objects (see Farah, 1994; Kanwisher and Yovel, 2006; for discussions). Along similar lines, in the functional neuroimaging literature, a number of studies have reported “activation dissociations” between written words/letter strings and/or visual objects and faces (Gauthier et al., 2000; Hasson et al., 2002; Puce et al., 1996; Bar et al., 2001; Malach et al., 2002). However, it is also true that a number of functional neuroimaging studies have found no such activation dissociations (Joseph et al 2003, 2006; Price and Devlin, 2003; Wright et al., 2008; Bookheimer et al., 2000; Indefrey et al., 1997; Moore and Price, 1999).
How can we account for this range of functional neuroimaging and neuropsychological findings? One possibility is that different visual categories may indeed recruit and require different subregions of the fusiform, but that these regional differences may not always be detected by the relatively large grain of functional neuroimaging techniques. However, given that the lesions suffered in cases of selective alexia, agnosia or prosopagnosia are not always especially small, this does not seem to be the most likely account of the observed neuropsychological dissociations. More likely is the possibility that while a range of different types of visual stimuli may engage the left fusiform, it is only orthographic processing that actually requires this region. Under this hypothesis, object processing requires networks localized in other cortical areas and/or in the right-hemisphere (Hasson, et al., 2002; Hemond et al., 2007). In fact, in the case of face processing the evidence seems to more clearly point to the critical role of the right-hemisphere mid-fusiform (for reviews and discussions see Grill-Spector et al., 2004; Kanwisher and Yovel, 2006; Kleinschmidt and Cohen, 2006; Yovel and Kanwisher, 2007).
In sum, with regard to the functional localization of face and object processes, DPT and the Gaillard et al. case add to the already considerable evidence that the left fusiform, is not specifically necessary face or object processing, although it may typically be activated by these stimuli in functional neuroimaging studies.
The question of functional reorganization
It is important to discuss the possibility that cognitive functions that were supported by mid-fusiform substrates may, in DPT’s case, have been taken up by other brain regions. One hypothesis is that, although prior to the resection multiple tasks may have required the left mid-fusiform area, due to post-surgical reorganization of function performance on these tasks was normal at the time DPT was evaluated. Specifically, the concern would be that the mid/anterior fusiform region is normally neither category nor modality specific but only appears that way after functional reorganization. Despite its surface plausibility, if we consider this possibility carefully, we see that we are still led to the conclusion that the left mid-fusiform supports modality and category specific orthographic functions. We lay out this reasoning in the following paragraphs.
As Price and Devlin (2003; see also Hillis et al., 2005) discussed there are at least three ways to think about the assignment of function/s to the left fusiform (1) there are several smaller and functionally specialized areas within the same fusiform area, (2) there is one underlying “higher” function subserved by this region and (3) the very same area subserves different functions depending on the “extra-fusiform” area/s it interacts with for each particular task. We consider each of these in turn in the context of possible functional reorganization. The first possibility is consistent with our conclusion of category and modality-specific orthographic functions within the left fusiform, as it allows for specifically orthographic functions in the fusiform to co-exist with other visual and spoken language functions. Under this hypothesis, the reason we see selective orthographic impairment in DPT’s case is that either (a) the visual and spoken language functions were spared by his lesion (they were, respectively, more posterior and anterior to the lesion) or (b) if the multiple independent orthographic, visual and spoken language functions were all affected by the lesion, functional reorganization or pre-existing redundant circuitry was available for visual and spoken language functions but not for orthographic ones.
The second possibility assumes that the left mid-fusiform supports a single function that is recruited by a wide range of tasks–reading, spelling, spoken naming, object recognition, etc. The first problem is that, without specifying what such a function might be, it is difficult to evaluate the hypothesis. Nonetheless, in the context of the question of functional reorganization if we assume that the reason that spoken language and visual processing are intact in DPT’s case (and also for Gaillard et al.’s patient) is because this single “general” function has been relocalized to another brain area, then the challenge is to explain why orthographic processing did not also benefit from this reorganization. The fact that orthographic processing remains impaired reveals that there was something uniquely necessary for orthographic processing that is subserved by this brain region-our conclusion precisely.
Finally, there is the possibility that the very same neurons within the mid-fusiform region take on different functions depending on which extra-fusiform they are interacting with in the task at hand. In that case, one could posit that after resection a different brain area comes to interact with those critical extra-fusiform regions to allow for intact spoken language and visual processing. The question would be—why didn’t this reorganization benefit written language processes? The critical point is: the fact that written language alone was unable to achieve this reorganization indicates that, in some manner, normal written language processing uniquely relies upon mid-fusiform substrates.
That is not to say that we can rule out that there has been any functional reorganization of orthographic processing functions. Furthermore, in considering the possibility of functional reorganization, it may be relevant to consider the nature of the neurological deficit. Specifically, a relatively slow-growing tumor may provide opportunities for reorganization or outcomes of reorganization that are either not possible with stroke or require a similarly long post-stroke time course. For example, one could speculate that the difference between DPT and the Gaillard et al. patient as well as many of the other letter-by-letter readers with lesions to the inferior temporal lobe (reviewed in the Introduction) with regard to presence versus absence of letter-by-letter reading is not due to differences in the location of the lesions along anterior-posterior axis of the fusiform (as we suggested above), but rather to the fact that DPT suffered from a slow-growing tumor that allowed for the gradual reassignment of the parallel processing of orthographic stimuli to other brain regions either within the left fusiform or to other brain regions. Although certainly a possibility, the fact remains that at least DPT and the Gaillard et al. patient differed in the location of their lesions. Similarly, the relative “mildness” of DPT’s reading impairment relative to what is often reported for individuals with inferior temporal lobe damage could, in some way, be the result of gradual reorganization.
The questions of reorganization of function and how it is affected by etiology of neural damage is an important one and functional neuroimaging of DPT’s reading and spelling performance might provide a useful means of revealing differences in activation patterns between DPT and normal readers and spellers, which, in turn, may provide insights into possible reorganization. Nonetheless, as we have argued above, these possibilities do not weaken the argument that DPT’s case provides clear evidence of orthography-specific functions of the fusiform.
Conclusions
There has been an extensive and ongoing debate regarding the possibility that neural substrates within the left fusiform gyrus may be dedicated to the modality and category specific processing of orthographic stimuli. A large set of neuroimaging and neuropsychological findings have been marshaled in support of both positions in this controversy. Although all sources of information play a valuable role, dissociations of functions associated with relatively restricted lesions are a particularly powerful form of evidence. As Kleinschmidt and Cohen (2006) noted ‘functional specialization in the sense of a critical role, i.e. loss of a function in case of damage, reflects whether a given region is the exclusive cortical locus for that particular function, and not whether it is exclusively engaged by that function’ (pg. 389). The case we have described in this paper provides strong evidence for the critical role of neural substrates within the left fusiform in orthographic processing and representation.
Acknowledgments
This research was made possible through the support of NIH grant DC006740 to the second author. We are deeply grateful to DPT for his patience and good humor through many hours of testing and for his generosity in donating his valuable time to this project.
Footnotes
1
Many photos of famous people were borrowed from Michele Miozzo’s laboratory database (used by permission) whereas non-famous peoples’ faces were taken from several internet resources.
2
Consistent with this characterization of the underlying deficit, many letter-by-letter readers, some of whom suffer from damage to the inferior temporal lobe, produce regularization errors and have slowed reading responses
3
Since DPT’s lesion was not entirely restricted to the mid-fusiform gyrus, but also included portions of the adjacent inferior temporal gyrus, we cannot conclusively attribute such functions to the mid-fusiform gyrus.. Nonetheless, our data clearly support orthography-specific substrates somewhere within the substrates corresponding to DPT’s lesion. Given the considerable evidence implicating the fusiform in orthographic processing and the current debate on how to specifically characterize the properties of this area, it is appropriate to assume that the fusiform is likely to be the relevant area.
4
Although Gaillard et al. do not report on nonword reading performance, presumably this would also have been carried out in a letter-by-letter fashion.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Army Individual Test Battery. Manual of directions and scoring. Washington, DC: War Department, Adjutant General’s Office; 1944. [Google Scholar]
- Baker CI, Hutchison TL, Kanwisher N. Does the fusiform face area contain subregions highly selective for nonfaces? Nature neuroscience. 2007a;10(1):3–4. doi: 10.1038/nn0107-3. [DOI] [PubMed] [Google Scholar]
- Baker CI, Liu J, Wald LL, Kwong KK, Benner T, Kanwisher N. Visual word processing and experiential origins of functional selectivity in human extrastriate cortex. Proceedings of the National Academy of Sciences of the United States of America. 2007b;104(21):9087–9092. doi: 10.1073/pnas.0703300104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bar M, Tootell RB, Schacter DL, Greve DN, Fischl B, Mendola JD, et al. Cortical mechanisms specific to explicit visual object recognition. Neuron. 2001;29(2):529–535. doi: 10.1016/s0896-6273(01)00224-0. [DOI] [PubMed] [Google Scholar]
- Bates E, D’Amico S, Jacobsen T, Szekely A, Andonova E, Devescovi A, et al. Timed picture naming in seven languages. Psychonomic bulletin & review. 2003;10(2):344–380. doi: 10.3758/bf03196494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beauregard M, Chertkow H, Gold D, Karama S, Benhamou J, Babins L, et al. Word priming with brief multiple presentation technique: Preservation in amnesia. Neuropsychologia. 1997;35(5):611–621. doi: 10.1016/s0028-3932(96)00108-x. [DOI] [PubMed] [Google Scholar]
- Beeson PM, Rapcsak SZ, Plante E, Chargualaf J, Chung A, Johnson SC, Trouard TP. The neural substrates of writing: A functional magnetic resonance imaging study. Aphasiology. 2003;17(67):647–665. [Google Scholar]
- Behrmann M, Nelson J, Sekuler EB. Visual complexity in letter-by-letter reading: “pure” alexia is not pure. Neuropsychologia. 1998;36(11):1115–1132. doi: 10.1016/s0028-3932(98)00005-0. [DOI] [PubMed] [Google Scholar]
- Benton AL, Hamsher KS, Sivan AB. Multilingual Aphasia Examination. Iowa City: AJA Associates; 1994. [Google Scholar]
- Binder JR, Frost JA, Hammeke TA, Bellgowan PS, Springer JA, Kaufman JN, et al. Human temporal lobe activation by speech and nonspeech sounds. Cerebral cortex (New York, N.Y.: 1991) 2000;10(5):512–528. doi: 10.1093/cercor/10.5.512. [DOI] [PubMed] [Google Scholar]
- Binder JR, Medler DA, Westbury CF, Liebenthal E, Buchanan L. Tuning of the human left fusiform gyrus to sublexical orthographic structure. NeuroImage. 2006;33(2):739–748. doi: 10.1016/j.neuroimage.2006.06.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binder JR, Mohr JP. The topography of callosal reading pathways. A case-control analysis. Brain : a journal of neurology. 1992;115(Pt 6):1807–1826. doi: 10.1093/brain/115.6.1807. [DOI] [PubMed] [Google Scholar]
- Blair JR, Spreen O. Predicting premorbid IQ: a revision of the National Adult Reading Test. The Clinical Neuropsychologist. 1989;3:129–136. [Google Scholar]
- Bookheimer SY, Zeffiro TA, Blaxton TA, Gaillard PW, Theodore WH. Activation of language cortex with automatic speech tasks. Neurology. 2000;55(8):1151–1157. doi: 10.1212/wnl.55.8.1151. [DOI] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TB, Mesulam MM. Functional anatomy of intra- and cross-modal lexical tasks. NeuroImage. 2002;16(1):7–22. doi: 10.1006/nimg.2002.1081. [DOI] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TB, Mesulam MM. Modality independence of word comprehension. Human brain mapping. 2002;16(4):251–261. doi: 10.1002/hbm.10054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth MC, Rolls ET. View-invariant representations of familiar objects by neurons in the inferior temporal visual cortex. Cerebral cortex (New York, N.Y.: 1991) 1998;8(6):510–523. doi: 10.1093/cercor/8.6.510. [DOI] [PubMed] [Google Scholar]
- Bornstein RA. Normative data on intermanual differences on three tests of motor performance. Journal of Clinical and Experimental Neuropsychology. 1986;8:12–20. doi: 10.1080/01688638608401293. [DOI] [PubMed] [Google Scholar]
- Buchel C, Price C, Frackowiak RS, Friston K. Different activation patterns in the visual cortex of late and congenitally blind subjects. Brain : a journal of neurology. 1998;121(Pt 3):409–419. doi: 10.1093/brain/121.3.409. [DOI] [PubMed] [Google Scholar]
- Buckner RL, Koutstaal W, Schacter DL, Rosen BR. Functional MRI evidence for a role of frontal and inferior temporal cortex in amodal components of priming. Brain : a journal of neurology. 2000;123(Pt 3):620–640. doi: 10.1093/brain/123.3.620. [DOI] [PubMed] [Google Scholar]
- Buxbaum LJ, Glosser G, Coslett HB. Impaired face and word recognition without object agnosia. Neuropsychologia. 1999;37(1):41–50. doi: 10.1016/s0028-3932(98)00048-7. [DOI] [PubMed] [Google Scholar]
- Cappa SF, Perani D, Schnur T, Tettamanti M, Fazio F. The effects of semantic category and knowledge type on lexical-semantic access: A PET study. NeuroImage. 1998;8(4):350–359. doi: 10.1006/nimg.1998.0368. [DOI] [PubMed] [Google Scholar]
- Carreiras M, Mechelli A, Estevez A, Price CJ. Brain activation for lexical decision and reading aloud: Two sides of the same coin? Journal of cognitive neuroscience. 2007;19(3):433–444. doi: 10.1162/jocn.2007.19.3.433. [DOI] [PubMed] [Google Scholar]
- Chan D, Fox NC, Scahill RI, Crum WR, Whitwell JL, Leschziner G, et al. Patterns of temporal lobe atrophy in semantic dementia and alzheimer’s disease. Annals of Neurology. 2001;49(4):433–442. [PubMed] [Google Scholar]
- Cohen L, Dehaene S. Specialization within the ventral stream: The case for the visual word form area. NeuroImage. 2004;22(1):466–476. doi: 10.1016/j.neuroimage.2003.12.049. [DOI] [PubMed] [Google Scholar]
- Cohen L, Dehaene S, Naccache L, Lehericy S, Dehaene-Lambertz G, Henaff MA, et al. The visual word form area: Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain : a journal of neurology. 2000;123(Pt 2):291–307. doi: 10.1093/brain/123.2.291. [DOI] [PubMed] [Google Scholar]
- Cohen L, Henry C, Dehaene S, Martinaud O, Lehericy S, Lemer C, et al. The pathophysiology of letter-by-letter reading. Neuropsychologia. 2004;42(13):1768–1780. doi: 10.1016/j.neuropsychologia.2004.04.018. [DOI] [PubMed] [Google Scholar]
- Cohen L, Lehericy S, Chochon F, Lemer C, Rivaud S, Dehaene S. Language-specific tuning of visual cortex? functional properties of the visual word form area. Brain : a journal of neurology. 2002;125(Pt 5):1054–1069. doi: 10.1093/brain/awf094. [DOI] [PubMed] [Google Scholar]
- Cohen L, Lehericy S, Henry C, Bourgeois M, Larroque C, Sainte-Rose C, et al. Learning to read without a left occipital lobe: Right-hemispheric shift of visual word form area. Annals of Neurology. 2004;56(6):890–894. doi: 10.1002/ana.20326. [DOI] [PubMed] [Google Scholar]
- Cohen L, Martinaud O, Lemer C, Lehericy S, Samson Y, Obadia M, et al. Visual word recognition in the left and right hemispheres: Anatomical and functional correlates of peripheral alexias. Cerebral cortex. 2003;13(12):1313–1333. doi: 10.1093/cercor/bhg079. [DOI] [PubMed] [Google Scholar]
- Coltheart M. The MRC Psycholinguistic Database. Quarterly Journal of Experimental Psychology. 1981;33A:497–505. [Google Scholar]
- Coltheart M. The psycholinguistic analysis of acquired dyslexias: Some illustrations. Philosophical transactions of the Royal Society of London. Series B, Biological sciences. 1982;298(1089):151–164. doi: 10.1098/rstb.1982.0078. [DOI] [PubMed] [Google Scholar]
- Crawford JR, Garthwaite PH. Investigation of the single case in neuropsychology: Confidence limits on the abnormality of test scores and test score differences. Neuropsychologia. 2002;40(8):1196–1208. doi: 10.1016/s0028-3932(01)00224-x. [DOI] [PubMed] [Google Scholar]
- Crawford JR, Garthwaite PH. Testing for suspected impairments and dissociations in single-case studies in neuropsychology: Evaluation of alternatives using monte carlo simulations and revised tests for dissociations. Neuropsychology. 2005;19(3):318–331. doi: 10.1037/0894-4105.19.3.318. [DOI] [PubMed] [Google Scholar]
- Crinion JT, Lambon-Ralph MA, Warburton EA, Howard D, Wise RJ. Temporal lobe regions engaged during normal speech comprehension. Brain : a journal of neurology. 2003;126(Pt 5):1193–1201. doi: 10.1093/brain/awg104. [DOI] [PubMed] [Google Scholar]
- Damasio AR. Time-locked multiregional retroactivation: A systems-level proposal for the neural substrates of recall and recognition. Cognition. 1989;33(1–2):25–62. doi: 10.1016/0010-0277(89)90005-x. [DOI] [PubMed] [Google Scholar]
- Damasio AR, Damasio H. The anatomic basis of pure alexia. Neurology. 1983;33(12):1573–1583. doi: 10.1212/wnl.33.12.1573. [DOI] [PubMed] [Google Scholar]
- De Renzi E, di Pellegrino G. Prosopagnosia and alexia without object agnosia. Cortex; a journal devoted to the study of the nervous system and behavior. 1998;34(3):403–415. doi: 10.1016/s0010-9452(08)70763-9. [DOI] [PubMed] [Google Scholar]
- De Renzi E, Zambolin A, Crisi G. The pattern of neuropsychological impairment associated with left posterior cerebral artery infarcts. Brain : a journal of neurology. 1987;110(Pt 5):1099–1116. doi: 10.1093/brain/110.5.1099. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Cohen L. Response to carreiras et al. The role of visual similarity, feedforward, feedback and lateral pathways in reading. Trends in cognitive sciences. 2007;11(11):456–457. [Google Scholar]
- Dehaene S, Cohen L, Sigman M, Vinckier F. The neural code for written words: A proposal. Trends in cognitive sciences. 2005;9(7):335–341. doi: 10.1016/j.tics.2005.05.004. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Jobert A, Naccache L, Ciuciu P, Poline JB, Le Bihan D, et al. Letter binding and invariant recognition of masked words: Behavioral and neuroimaging evidence. Psychological science : a journal of the American Psychological Society/APS. 2004;15(5):307–313. doi: 10.1111/j.0956-7976.2004.00674.x. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Le Clec HG, Poline JB, Le Bihan D, Cohen L. The visual word form area: A prelexical representation of visual words in the fusiform gyrus. Neuroreport. 2002;13(3):321–325. doi: 10.1097/00001756-200203040-00015. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Naccache L, Cohen L, Bihan DL, Mangin JF, Poline JB, et al. Cerebral mechanisms of word masking and unconscious repetition priming. Nature neuroscience. 2001;4(7):752–758. doi: 10.1038/89551. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Tzourio N, Frak V, Raynaud L, Cohen L, Mehler J, et al. Cerebral activations during number multiplication and comparison: A PET study. Neuropsychologia. 1996;34(11):1097–1106. doi: 10.1016/0028-3932(96)00027-9. [DOI] [PubMed] [Google Scholar]
- Dejerine J. Contribution a l’etude anatomoclinique et clinique des differentes varietes de cecite verbale. CR Hebdomadaire des Seances et Memoires de la Societe de Biologie. 1892;4:61–90. [Google Scholar]
- Demonet JF, Chollet F, Ramsay S, Cardebat D, Nespoulous JL, Wise R, et al. The anatomy of phonological and semantic processing in normal subjects. Brain : a journal of neurology. 1992;115(Pt 6):1753–1768. doi: 10.1093/brain/115.6.1753. [DOI] [PubMed] [Google Scholar]
- Demonet JF, Price C, Wise R, Frackowiak RS. Differential activation of right and left posterior sylvian regions by semantic and phonological tasks: A positron-emission tomography study in normal human subjects. Neuroscience letters. 1994;182(1):25–28. doi: 10.1016/0304-3940(94)90196-1. [DOI] [PubMed] [Google Scholar]
- Devlin JT, Jamison HL, Gonnerman LM, Matthews PM. The role of the posterior fusiform gyrus in reading. Journal of cognitive neuroscience. 2006;18(6):911–922. doi: 10.1162/jocn.2006.18.6.911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Virgilio G, Clarke S. Direct interhemispheric visual input to human speech areas. Human brain mapping. 1997;5:347–354. doi: 10.1002/(SICI)1097-0193(1997)5:5<347::AID-HBM3>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- Downing PE, Chan AW, Peelen MV, Dodds CM, Kanwisher N. Domain specificity in visual cortex. Cerebral cortex (New York, N.Y.: 1991) 2006;16(10):1453–1461. doi: 10.1093/cercor/bhj086. [DOI] [PubMed] [Google Scholar]
- Dunn L. Peabody Picture Vocabulary Test. Circle Pine. v., MN: American Guidance Service; 1959. [Google Scholar]
- Ellis AW, Young AW. Human cognitive neuropsychology. Hillsdale, NJ: Lawrence Erlbaum Associates; 1988. [Google Scholar]
- Farah MJ. Specialization within visual object recognition: clues from prosopagnosia and alexia. In: Farah MJ, Ratcliff G, editors. The Neuropsychology of High-Level Vision. Erlbaum; Hillsdale, NJ: 1994. pp. 133–146. [Google Scholar]
- Farah MJ, Wallace MA. Pure alexia as a visual impairment: A reconsideration. Cognitive Neuropsychology. 1991;8:313–334. [Google Scholar]
- Farah MJ, Wilson KD, Drain M, Tanaka JN. What is “special” about face perception? Psychological review. 1998;105(3):482–498. doi: 10.1037/0033-295x.105.3.482. [DOI] [PubMed] [Google Scholar]
- Feinberg TE, Schindler RJ, Ochoa E, Kwan PC, Farah MJ. Associative visual agnosia and alexia without prosopagnosia. Cortex; a journal devoted to the study of the nervous system and behavior. 1994;30(3):395–411. doi: 10.1016/s0010-9452(13)80337-1. [DOI] [PubMed] [Google Scholar]
- Fiez JA, Balota DA, Raichle ME, Petersen SE. Effects of lexicality, frequency, and spelling-to-sound consistency on the functional anatomy of reading. Neuron. 1999;24(1):205–218. doi: 10.1016/s0896-6273(00)80833-8. [DOI] [PubMed] [Google Scholar]
- Finger S. Origins of Neuroscience: A history of explorations into brain function. New York, NY: Oxford University Press; 1994. [Google Scholar]
- Folk J, Rapp B. Interaction of lexical and sublexical information in spelling : Evidence from nonword priming. Applied Psycholinguistics. 2004;25:565–585. [Google Scholar]
- Foundas AL, Daniels SK, Vasterling JJ. Anomia: Case studies with lesion localisation. Neurocase. 1998;4:35–43. [Google Scholar]
- Francis WN, Kucera H. Frequency analysis of english usage. Boston: Houghton-Mifflin Company; 1982. [Google Scholar]
- Gaillard R, Naccache L, Pinel P, Clemenceau S, Volle E, Hasboun D, et al. Direct intracranial, FMRI, and lesion evidence for the causal role of left inferotemporal cortex in reading. Neuron. 2006;50(2):191–204. doi: 10.1016/j.neuron.2006.03.031. [DOI] [PubMed] [Google Scholar]
- Gauthier I, Tarr MJ, Moylan J, Skudlarski P, Gore JC, Anderson AW. The fusiform “face area” is part of a network that processes faces at the individual level. Journal of cognitive neuroscience. 2000;12(3):495–504. doi: 10.1162/089892900562165. [DOI] [PubMed] [Google Scholar]
- Geschwind N. Disconnexion syndromes in animals and man. I. [review] Brain : a journal of neurology. 1965;88:237–294. doi: 10.1093/brain/88.2.237. [DOI] [PubMed] [Google Scholar]
- Giraud AL, Price CJ. The constraints functional neuroimaging places on classical models of auditory word processing. Journal of cognitive neuroscience. 2001;13(6):754–765. doi: 10.1162/08989290152541421. [DOI] [PubMed] [Google Scholar]
- Goodman RA, Caramazza A. The Johns Hopkins University dysgraphia battery. Baltimore, MD: The Johns Hopkins University; 1985. [Google Scholar]
- Graham NL, Patterson K, Hodges JR. The impact of semantic memory impairment on spelling: Evidence from semantic dementia. Neuropsychologia. 2000;38(2):143–163. doi: 10.1016/s0028-3932(99)00060-3. [DOI] [PubMed] [Google Scholar]
- Grill-Spector K, Kanwisher N. Visual recognition: As soon as you know it is there, you know what it is. Psychological science : a journal of the American Psychological Society/APS. 2005;16(2):152–160. doi: 10.1111/j.0956-7976.2005.00796.x. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Indefrey P, Brown C, Herzog H, Steinmetz H, Seitz RJ. The neural circuitry involved in the reading of German words and pseudowords: A PET study. Journal of cognitive neuroscience. 1999;11(4):383–398. doi: 10.1162/089892999563490. [DOI] [PubMed] [Google Scholar]
- Hasson U, Levy I, Behrmann M, Hendler T, Malach R. Eccentricity bias as an organizing principle for human high-order object areas. Neuron. 2002;34(3):479–490. doi: 10.1016/s0896-6273(02)00662-1. [DOI] [PubMed] [Google Scholar]
- Hemond CC, Kanwisher NG, Op de Beeck HP. A preference for contralateral stimuli in human object- and face-selective cortex. PLoS ONE. 2007;2(6):e574. doi: 10.1371/journal.pone.0000574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillis AE, Caramazza A. Mechanisms for accessing lexical representations for output: Evidence from a category-specific semantic deficit. Brain and language. 1991;40(1):106–144. doi: 10.1016/0093-934x(91)90119-l. [DOI] [PubMed] [Google Scholar]
- Henry C, Gaillard R, Volle E, Chiras J, Ferrieux S, Dehaene S, et al. Brain activations during letter-by-letter reading: A follow-up study. Neuropsychologia. 2005;43(14):1983–1989. doi: 10.1016/j.neuropsychologia.2005.04.007. [DOI] [PubMed] [Google Scholar]
- Hillis AE, Caramazza A. Mechanisms for accessing lexical representations for output: Evidence from a category-specific semantic deficit. Brain and language. 1991;40(1):106–144. doi: 10.1016/0093-934x(91)90119-l. [DOI] [PubMed] [Google Scholar]
- Hillis AE, Newhart M, Heidler J, Barker P, Herskovits E, Degaonkar M. The roles of the “visual word form area” in reading. NeuroImage. 2005;24(2):548–559. doi: 10.1016/j.neuroimage.2004.08.026. [DOI] [PubMed] [Google Scholar]
- Hillis AE, Rapp B. Cognitive and neural substrates of written language comprehension and production. In: Gazzaniga MS, editor. The new cognitive neurosciences. 3. Cambridge, MA: MIT Press; 2004. pp. 755–88. [Google Scholar]
- Humphreys GW, Rumiati RI. Agnosia without prosopagnosia or alexia: Evidence for stored visual memories specific to objects. Cognitive Neuropsychology. 1998;15(3):243–277. doi: 10.1080/026432998381177. [DOI] [PubMed] [Google Scholar]
- Howard D, Patterson K. The pyramid and palm trees test: A test of semantic access from words and pictures. Bury St. Edmunds: Thames Valley Test Company; 1992. [Google Scholar]
- Hsieh L, Rapp B. Functional magnetic resonance imaging of the cognitive components of the spelling process. Brain and language. 2004;91(1):40–41. [Google Scholar]
- Indefrey P, Kleinschmidt A, Merboldt KD, Kruger G, Brown C, Hagoort P, et al. Equivalent responses to lexical and nonlexical visual stimuli in occipital cortex: A functional magnetic resonance imaging study. NeuroImage. 1997;5(1):78–81. doi: 10.1006/nimg.1996.0232. [DOI] [PubMed] [Google Scholar]
- Jobard G, Crivello F, Tzourio-Mazoyer N. Evaluation of the dual route theory of reading: A metanalysis of 35 neuroimaging studies. NeuroImage. 2003;20(2):693–712. doi: 10.1016/S1053-8119(03)00343-4. [DOI] [PubMed] [Google Scholar]
- Jobard G, Vigneau M, Mazoyer B, Tzourio-Mazoyer N. Impact of modality and linguistic complexity during reading and listening tasks. NeuroImage. 2007;34(2):784–800. doi: 10.1016/j.neuroimage.2006.06.067. [DOI] [PubMed] [Google Scholar]
- Joseph JE, Gathers AD, Piper GA. Shared and dissociated cortical regions for object and letter processing. Brain research. Cognitive brain research. 2003;17(1):56–67. doi: 10.1016/s0926-6410(03)00080-6. [DOI] [PubMed] [Google Scholar]
- Kanwisher N, Yovel G. The fusiform face area: A cortical region specialized for the perception of faces. Philosophical transactions of the Royal Society of London. Series B, Biological sciences. 2006;361(1476):2109–2128. doi: 10.1098/rstb.2006.1934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan E, Goodglass H, Weintraub S. The Boston Naming Test. Philadelphia: Lea and Febiger; 1983. [Google Scholar]
- Krauss GL, Fisher R, Plate C, Hart J, Uematsu S, Gordon B, et al. Cognitive effects of resecting basal temporal language areas. Epilepsia. 1996;37(5):476–483. doi: 10.1111/j.1528-1157.1996.tb00594.x. [DOI] [PubMed] [Google Scholar]
- Kreiman G, Koch C, Fried I. Category-specific visual responses of single neurons in the human medial temporal lobe. Nature neuroscience. 2000;3(9):946–953. doi: 10.1038/78868. [DOI] [PubMed] [Google Scholar]
- Kronbichler M, Bergmann J, Hutzler F, Staffen W, Mair A, Ladurner G, et al. Taxi vs. taksi: On orthographic word recognition in the left ventral occipitotemporal cortex. Journal of cognitive neuroscience. 2007;19(10):1584–1594. doi: 10.1162/jocn.2007.19.10.1584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kronbichler M, Hutzler F, Wimmer H, Mair A, Staffen W, Ladurner G. The visual word form area and the frequency with which words are encountered: Evidence from a parametric fMRI study. NeuroImage. 2007;21(3):946–953. doi: 10.1016/j.neuroimage.2003.10.021. [DOI] [PubMed] [Google Scholar]
- Kucera M, Francis WN. Computational Analysis of Present-Day American English. Providence: Brown University Press; 1967. [Google Scholar]
- Lambon-Ralph MA, Patterson K, Garrard P, Hodges JR. Semantic dementia with category specificity: A comparative case-series study. Cognitive Neuropsychology. 2003;20(3456):307–326. doi: 10.1080/02643290244000301. [DOI] [PubMed] [Google Scholar]
- Leff AP, Crewes H, Plant GT, Scott SK, Kennard C, Wise RJ. The functional anatomy of single-word reading in patients with hemianopic and pure alexia. Brain : a journal of neurology. 2001;124(Pt 3):510–521. doi: 10.1093/brain/124.3.510. [DOI] [PubMed] [Google Scholar]
- Luders H, Lesser RP, Hahn J, Dinner DS, Morris HH, Wyllie E, Godoy J. Basal temporal language area. Brain : a journal of neurology. 1991;114(2):743–754. doi: 10.1093/brain/114.2.743. [DOI] [PubMed] [Google Scholar]
- Malach R, Levy I, Hasson U. The topography of high-order human object areas. Trends in cognitive sciences. 2002;6(4):176–184. doi: 10.1016/s1364-6613(02)01870-3. [DOI] [PubMed] [Google Scholar]
- Martin A, Chao LL. Semantic memory and the brain: Structure and processes. Current opinion in neurobiology. 2001;11(2):194–201. doi: 10.1016/s0959-4388(00)00196-3. [DOI] [PubMed] [Google Scholar]
- McCandliss BD, Cohen L, Dehaene S. The visual word form area: Expertise for reading in the fusiform gyrus. Trends in cognitive sciences. 2003;7(7):293–299. doi: 10.1016/s1364-6613(03)00134-7. [DOI] [PubMed] [Google Scholar]
- Mechelli A, Crinion JT, Long S, Friston KJ, Lambon-Ralph MA, Patterson K, et al. Dissociating reading processes on the basis of neuronal interactions. Journal of cognitive neuroscience. 2005;17(11):1753–1765. doi: 10.1162/089892905774589190. [DOI] [PubMed] [Google Scholar]
- Mechelli A, Gorno-Tempini ML, Price CJ. Neuroimaging studies of word and pseudoword reading: Consistencies, inconsistencies, and limitations. Journal of cognitive neuroscience. 2003;15(2):260–271. doi: 10.1162/089892903321208196. [DOI] [PubMed] [Google Scholar]
- Miceli G, Capasso R, Caramazza A. The interaction of lexical and sublexical processes in reading, writing and repetition. Neuropsychologia. 1994;32(3):317–333. doi: 10.1016/0028-3932(94)90134-1. [DOI] [PubMed] [Google Scholar]
- Moore CJ, Price CJ. Three distinct ventral occipitotemporal regions for reading and object naming. NeuroImage. 1999;10(2):181–192. doi: 10.1006/nimg.1999.0450. [DOI] [PubMed] [Google Scholar]
- Mummery CJ, Patterson K, Hodges JR, Price CJ. Functional neuroanatomy of the semantic system: Divisible by what? Journal of cognitive neuroscience. 1998;10(6):766–777. doi: 10.1162/089892998563059. [DOI] [PubMed] [Google Scholar]
- Mummery CJ, Patterson K, Price CJ, Ashburner J, Frackowiak RS, Hodges JR. A voxel-based morphometry study of semantic dementia: Relationship between temporal lobe atrophy and semantic memory. Annals of Neurology. 2000;47(1):36–45. [PubMed] [Google Scholar]
- Mummery CJ, Patterson K, Wise RJ, Vandenberghe R, Price CJ, Hodges JR. Disrupted temporal lobe connections in semantic dementia. Brain : a journal of neurology. 1999;122(Pt 1):61–73. doi: 10.1093/brain/122.1.61. [DOI] [PubMed] [Google Scholar]
- Nobre AC, Allison T, McCarthy G. Word recognition in the human inferior temporal lobe. Nature. 1994;372(6503):260–263. doi: 10.1038/372260a0. [DOI] [PubMed] [Google Scholar]
- Noppeney U, Price CJ. A PET study of stimulus- and task-induced semantic processing. NeuroImage. 2002;15(4):927–935. doi: 10.1006/nimg.2001.1015. [DOI] [PubMed] [Google Scholar]
- Noppeney U, Price CJ. Retrieval of visual, auditory, and abstract semantics. NeuroImage. 2002;15(4):917–926. doi: 10.1006/nimg.2001.1016. [DOI] [PubMed] [Google Scholar]
- Osterrieth PA. The complex figure copy test. In: Corwin J, Bylsma FW, translators. The Clinical Neuropsychologist. Vol. 7. 1993. pp. 9–15.Reprinted from Osterrieth PA. Le test de copie d’une figure complexe. Archives de Psychologie. 1944;30:286–356.
- Pavio A, Yuille JC, Madigan SA. Concrete- ness, imagery and meaningfulness values for 925 words. Journal of Experimental Psychology Monograph Supplement. 1968;76(3 part 2) doi: 10.1037/h0025327. [DOI] [PubMed] [Google Scholar]
- Plaut DC, McClelland JL, Seidenberg MS, Patterson K. Understanding normal and impaired word reading: Computational principles in quasi-regular domains. Psychological review. 1996;103(1):56–115. doi: 10.1037/0033-295x.103.1.56. [DOI] [PubMed] [Google Scholar]
- Price CJ, Devlin JT. The myth of the visual word form area. NeuroImage. 2003;19(3):473–481. doi: 10.1016/s1053-8119(03)00084-3. [DOI] [PubMed] [Google Scholar]
- Price CJ, Devlin JT. The pro and cons of labelling a left occipitotemporal region: “the visual word form area”. NeuroImage. 2004;22(1):477–479. doi: 10.1016/j.neuroimage.2004.01.018. [DOI] [PubMed] [Google Scholar]
- Price CJ, Friston KJ. Scanning patients with tasks they can perform. Human brain mapping. 1999;8(2–3):102–108. doi: 10.1002/(SICI)1097-0193(1999)8:2/3<102::AID-HBM6>3.0.CO;2-J. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price CJ, Gorno-Tempini ML, Graham KS, Biggio N, Mechelli A, Patterson K, et al. Normal and pathological reading: Converging data from lesion and imaging studies. NeuroImage. 2003;20 (Suppl 1):S30–41. doi: 10.1016/j.neuroimage.2003.09.012. [DOI] [PubMed] [Google Scholar]
- Price CJ, Mechelli A. Reading and reading disturbance. Current opinion in neurobiology. 2005;15(2):231–238. doi: 10.1016/j.conb.2005.03.003. [DOI] [PubMed] [Google Scholar]
- Puce A, Allison T, Asgari M, Gore JC, McCarthy G. Differential sensitivity of human visual cortex to faces, letterstrings, and textures: A functional magnetic resonance imaging study. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1996;16(16):5205–5215. doi: 10.1523/JNEUROSCI.16-16-05205.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pugh KR, Mencl WE, Jenner AR, Katz L, Frost SJ, Lee JR, et al. Neurobiological studies of reading and reading disability. Journal of communication disorders. 2001;34(6):479–492. doi: 10.1016/s0021-9924(01)00060-0. [DOI] [PubMed] [Google Scholar]
- Pugh KR, Shaywitz BA, Shaywitz SE, Constable RT, Skudlarski P, Fulbright RK, et al. Cerebral organization of component processes in reading. Brain : a journal of neurology. 1996;119(Pt 4):1221–1238. doi: 10.1093/brain/119.4.1221. [DOI] [PubMed] [Google Scholar]
- Rapcsak SZ, Beeson PM. The role of left posterior inferior temporal cortex in spelling. Neurology. 2004;62(12):2221–2229. doi: 10.1212/01.wnl.0000130169.60752.c5. [DOI] [PubMed] [Google Scholar]
- Rapp B, Caramazza A. From graphemes to abstract letter shapes: Levels of representation in written spelling. Journal of experimental psychology. Human perception and performance. 1997;23(4):1130–1152. doi: 10.1037//0096-1523.23.4.1130. [DOI] [PubMed] [Google Scholar]
- Rapp B, Caramazza A. The modality-specific organization of grammatical categories: Evidence from impaired spoken and written sentence production. Brain and language. 1997;56(2):248–286. doi: 10.1006/brln.1997.1735. [DOI] [PubMed] [Google Scholar]
- Rapp B, Epstein C, Tainturier MJ. The integration of information across lexical and sublexical processes in spelling. Cognitive Neuropsychology. 2002;19:1–29. doi: 10.1080/0264329014300060. [DOI] [PubMed] [Google Scholar]
- Rapp B, Glucroft B, Tsapkini K. Investigating the shared cognitive and neural substrates of reading and spelling. Paper presented at: Conference on the Mental Lexicon; Montreal, Canada. 2006. [Google Scholar]
- Rapp B, Hsieh L. Functional magnetic resonance imaging of the cognitive components of the spelling process. Paper presented at: Cognitive Neuroscience Society Meeting; San Francisco, CA. 2002. [Google Scholar]
- Raymer AM, Foundas AL, Maher LM, Greenwald ML, Morris M, Rothi LJ, et al. Cognitive neuropsychological analysis and neuroanatomic correlates in a case of acute anomia. Brain and language. 1997;58(1):137–156. doi: 10.1006/brln.1997.1786. [DOI] [PubMed] [Google Scholar]
- Rey A. Psychological examination of traumatic encephalopathy. In: Corwin J, Bylsma FW, translators. The Clinical Neuropsychologist. Vol. 7. 1993. pp. 4–9.Reprinted from Rey A. L’examen psychologique dans les cas d’encephalopathie traumatique. Archives de Psychologie. 1941;28:286–340.
- Rey A. L’examen clinique en psychologie. Paris: Presses Universitaires de France; 1964. [Google Scholar]
- Rossion B, Pourtois G. Revisiting snodgrass and vanderwart’s object pictorial set: The role of surface detail in basic-level object recognition. Perception. 2004;33(2):217–236. doi: 10.1068/p5117. [DOI] [PubMed] [Google Scholar]
- Rumiati RI, Humphreys GW. Recognition by action: Dissociating visual and semantic routes to action in normal observers. Journal of experimental psychology. Human perception and performance. 1998;24(2):631–647. doi: 10.1037//0096-1523.24.2.631. [DOI] [PubMed] [Google Scholar]
- Salmelin R, Service E, Kiesila P, Uutela K, Salonen O. Impaired visual word processing in dyslexia revealed with magnetoencephalography. Annals of Neurology. 1996;40(2):157–162. doi: 10.1002/ana.410400206. [DOI] [PubMed] [Google Scholar]
- Schwartz MF, Saffran EM, Marin OSM. Fractionating the reading process in dementia: Evidence for word-specific print-to-sound associations. In: Coltheart M, Patterson KE, Marshall JC, editors. Deep Dyslexia. London: Routledge & Kegan Paul; 1980. pp. 259–269. [Google Scholar]
- Seidenberg MS, Waters GS, Barnes MA. When does irregular spelling or pronunciation influence word recognition? Journal of Verbal Learning and Verbal Behavior. 1984;23:383–404. [Google Scholar]
- Sharp DJ, Scott SK, Wise RJ. Retrieving meaning after temporal lobe infarction: The role of the basal language area. Annals of Neurology. 2004;56(6):836–846. doi: 10.1002/ana.20294. [DOI] [PubMed] [Google Scholar]
- Snodgrass JG, Vanderwart M. A standardized set of 260 pictures: Norms for name agreement, image agreement, familiarity, and visual complexity. Journal of experimental psychology. Human learning and memory. 1980;6(2):174–215. doi: 10.1037//0278-7393.6.2.174. [DOI] [PubMed] [Google Scholar]
- Starrfelt R, Gerlach C. The visual what for area: Words and pictures in the left fusiform gyrus. NeuroImage. 2007;35(1):334–342. doi: 10.1016/j.neuroimage.2006.12.003. [DOI] [PubMed] [Google Scholar]
- Stroop JR. Studies of interference in serial verbal reactions. Journal of Experimental Psychology. 1935;18:643–662. [Google Scholar]
- Tainturier M-J. Phonologically based spelling errors and their implications in the specifcation of phonology to orthography conversion processes. Brain and Cognition. 1996;32(2):148–51. [Google Scholar]
- Tainturier M-J, Rapp B. The spelling process. In: Rapp B, editor. The handbook of cognitive neuropsychology: What deficits reveal about the human mind. Philadelphia, PA: Psychology Press (Taylor and Francis Group); 2001. pp. 263–289. [Google Scholar]
- Tainturier MJ, Rapp B. Complex graphemes as functional spelling units: Evidence from acquired dysgraphia. Neurocase : case studies in neuropsychology, neuropsychiatry, and behavioural neurology. 2004;10(2):122–131. doi: 10.1080/13554790409609943. [DOI] [PubMed] [Google Scholar]
- Tarr M-J, Gauthier I. FFA: A flexible fusiform area for subordinate-level visual processing automatized by expertise. Nature neuroscience. 2000;3(8):764–769. doi: 10.1038/77666. [DOI] [PubMed] [Google Scholar]
- Thompson-Schill SL, Aguirre GK, D’Esposito M, Farah MJ. A neural basis for category and modality specificity of semantic knowledge. Neuropsychologia. 1999;37(6):671–676. doi: 10.1016/s0028-3932(98)00126-2. [DOI] [PubMed] [Google Scholar]
- Vigneau M, Jobard G, Mazoyer B, Tzourio-Mazoyer N. Word and non-word reading: What role for the visual word form area? NeuroImage. 2005;27(3):694–705. doi: 10.1016/j.neuroimage.2005.04.038. [DOI] [PubMed] [Google Scholar]
- Vinckier F, Dehaene S, Jobert A, Dubus JP, Sigman M, Cohen L. Hierarchical coding of letter strings in the ventral stream: Dissecting the inner organization of the visual word-form system. Neuron. 2007;55(1):143–156. doi: 10.1016/j.neuron.2007.05.031. [DOI] [PubMed] [Google Scholar]
- Wagner JC, Rogers JE, Baud RH, Scherrer JR. Natural language generation of surgical procedures. MEDINFO. 1998;9 (Pt 1):591–595. [PubMed] [Google Scholar]
- Warrington EK. The Recognition Memory Test. Windsor: NFER-Nelson; 1984. [Google Scholar]
- Wechsler D. Wechsler Memory Scale—Revised manual. San Antonio, TX: Psychological Corporation; [Google Scholar]
- Wright ND, Mechelli A, Noppeney U, Veltman DJ, Rombouts SA, Glensman J, et al. Selective activation around the left occipito-temporal sulcus for words relative to pictures: Individual variability or false positives? Human brain mapping. 2008;29(8):986–1000. doi: 10.1002/hbm.20443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yovel G, Kanwisher N. Face perception: Domain specific, not process specific. Neuron. 2004;44(5):889–898. doi: 10.1016/j.neuron.2004.11.018. [DOI] [PubMed] [Google Scholar]