Ecological and Morphological Profile of Floating Spherical Cladophora socialis Aggregations in Central Thailand (original) (raw)
Abstract
The unique beauty of spherical aggregation forming algae has attracted much attention from both the scientific and lay communities. Several aegagropilous seaweeds have been identified to date, including the plants of genus Cladophora and Chaetomorpha. However, this phenomenon remains poorly understood. In July 2013, a mass occurrence of spherical Cladophora aggregations was observed in a salt field reservoir in Central Thailand. The aims of the present study were to describe the habitat of the spherical aggregations and confirm the species. We performed a field survey, internal and external morphological observations, pyrenoid ultrastructure observations, and molecular sequence analysis. Floating spherical Cladophora aggregations (1–8 cm in diameter) were observed in an area ~560 m2, on the downwind side of the reservoir where there was water movement. Individual filaments in the aggregations were entangled in each other; consequently, branches growing in different directions were observed within a clump. We suggest that water movement and morphological characteristics promote the formation of spherical aggregations in this species. The molecular sequencing results revealed that the study species was highly homologous to both C. socialis and C. coelothrix. However, the diameter of the apical cells in the study species was less than that of C. coelothrix. The pyrenoid ultrastructure was more consistent with that of C. socialis. We conclude that the study species is C. socialis. This first record of spherical aggregations in this species advances our understanding of these formations. However, further detailed physical measurements are required to fully elucidate the mechanism behind these spherical formations.
Introduction
The unique morphology of spherical growth seaweeds has gained them considerable popularity [1], both socially and scientifically. Mass media have sensationally referred to sporadic occurrences of aegagropilous seaweeds as mystery balls, UFOs, and green alien balls [2, 3]. An extremely rare freshwater alga, Aegagropila linnaei, is known for its beautiful spherical filamentous aggregations [4]. It has become popular in the souvenir and aquarium trades, and is depicted on postage stamps from Japan and Iceland [1, 5]. This alga is an endangered or protected species in Japan, Iceland, United Kingdom, Germany, Sweden, and Russia, because of declining populations, and is included in national red lists and other conservation strategies [5, 6]. Scientifically, however, the mechanism for forming spherical aggregations has been researched and discussed. It is generally assumed that the ball shape is a result of both mechanical factors, such as wave-induced rolling motion, and morphological features leading to entanglement of the filament [1, 5].
Besides A. linnaei, several aegagropilous seaweeds are known [7–9], e.g., at least 18 green, 11 brown, and 25 red algae form aggregations [2]. Of the Cladophorales family species, Chaetomorpha picquotiana [2], Cladophora coelothrix [10], C. prolifera [9, 11, 12] have ball-like forms including loose-lying balls and spherical forms. However, information on the habitat and ecology of spherical aggregation forms of this taxon remains rare.
Cladophora is one of the largest and most common green algal genera, having a worldwide distribution [13]. Despite having simple external morphologies, there are 242 recognized species in this genus [14]. However, it is difficult to identify the species in this genus by external morphology, because of its few morphological characteristics and extensive morphological plasticity related to environmental conditions, age of the alga, and season [15–18]. Additionally, Cladophora is not monophyletic, and even the same morphospecies are distributed in a number of different clades in a phylogenetic tree, as a result species identification using only nucleic acid sequences is very difficult [13, 19, 20]. Thus, various methods are required to identify the Cladophora species [21, 22], including internal morphology and ultrastructure [18, 22–25], external morphology, and molecular sequencing analysis.
The aims of this study were: 1) to describe the habitat of the spherical Cladophora aggregations that occurred in July 2013 in Central Thailand and 2) to use a combination of external and internal morphological characteristics, ultrastructure, growth pattern, and molecular sequence analysis to identify the aggregations to the species level.
Materials and Methods
Ethics Statement
All research activities in this study, including field surveys, observations, and laboratory experiments, were permitted by the National Research Council of Thailand (NRCT) (Project ID 2011/005). The field survey site was a privately owned pond and the authors obtained oral approval for sampling from the pond owner. The field study did not involve endangered or protected species.
Field survey
A field survey was conducted in a salt field reservoir in Samut Sakhon Province (13.504629N and 100.217659E) to identify the habitat of spherical Cladophora aggregations in July 2013 during a large-scale occurrence (Fig 1). Salinity and water temperature were measured on site with a conductivity meter (Condo 3210, WTW Inc., Weilheim, Germany). The pond area was measured with a laser distance meter (GLM 250 VF, Robert Bosch Inc., Stuttgart, Germany). A plastic ruler was used to measure water depth. Total coverage of floating spherical aggregations was directly observed by eye and estimated. The water movement was analyzed from video images of seaweed movement captured by mobile phone (GT-i9500, Samsung Electronics Co., Ltd., Suwon, South Korea). A small number of spherical aggregations of this alga were collected by hand at the survey site and immediately placed in plastic bags with water, after which they were transported to the Shrimp Co-culture Research Laboratory (SCORL), Faculty of Agricultural Technology, King Mongkut’s Institute of Technology Ladkrabang (KMITL), Thailand for observation and laboratory experiments. Some of the samples were dried, pressed, and preserved in the herbarium as specimens at SCORL.
Fig 1. Map showing field survey location in Central Thailand.

Morphological observations
Samples brought back to SCORL were rinsed in artificial seawater (Rhoto Marine II, Rei-Sea Co. Ltd., Tokyo, Japan) with a salinity of 20 PSU. Part of the spherical aggregations was cut off and observed under an optical microscope (BX 41, Olympus Co. Ltd., Tokyo, Japan) to study the external morphology. For internal morphology, such as nuclei, the samples were fixed with 3:1 ethanol: glacial acetic acid and stained with aceto-iron-hematoxylin chloral hydrate [26].
Ultrastructure observations
The samples were fixed with 2% paraformaldehyde (PHA) and 2% glutaraldehyde (GA) in 0.05 M cacodylate buffer pH 7.4 at 4°C overnight. After fixation, the samples were washed three times with 0.05 M cacodylate buffer for 30 min, and postfixed with 2% osmium tetroxide (OsO4) in 0.05 M cacodylate buffer at 4°C for 3 h. The samples were dehydrated in graded ethanol solutions; 50% for 30 min at 4°C, 70% for 30 min at 4°C, 90% for 30 min at room temperature, 100% ethanol was changed four times at room temperature. After the dehydration process, the samples were continuously dehydrated in 100% ethanol at room temperature overnight. The samples were infiltrated with propylene oxide (PO) twice for 30 min and put into a 7:3 mixture of PO and resin (Quetol-651, Nisshin EM Co., LTD., Tokyo, Japan) for 1 h. The tube cap was left open overnight and PO was volatilized. The samples were transferred to fresh 100% resin and polymerized at 60°C for 48 h. The polymerized resins were ultra-thin sectioned at 70 nm with a diamond knife using an ultra-microtome (Ultracut UCT, Leica, Vienna, Austria), and the sections were mounted on copper grids. These samples were stained with 2% uranyl acetate at room temperature for 15 min, and washed with distilled water followed by secondary staining with lead stain solution (Sigma Aldrich Co. Ltd., Tokyo, Japan) at room temperature for 3 min. The samples on grids were observed on a transmission electron microscope (JEM-1400Plus, JEOL ltd., Tokyo, Japan) at an acceleration voltage of 80 kV. Digital images were taken with a CCD camera (VELETA, Olympus Soft Imaging Solution GMBH, Münster, Germany).
Growth pattern experiment
Prior to the experiments, powdered, artificial seawater (Rhoto Marine II, Rei-Sea Co. Ltd., Tokyo, Japan) was diluted with distilled water and adjusted to 20 PSU with a conductivity meter (Condo 3210, WTW Inc.). This experimental medium was sterilized at 121°C for 20 min in an autoclave (HVE-50, Hirayama Manufacturing Corp., Saitama, Japan), after which the pH was adjusted to 8.0 with NaOH using a pH meter (SevenGo, Mettler Toledo, Schwerzenbach, Switzerland). As a nutrient source, a ratio of 3 mL of stock solution of Provasoli’s enriched seawater (PES) in 1 L artificial seawater (approximately 1/6 concentration of PES) was used [27], germanium dioxide (3 mg L−1) was added to prevent diatom growth [28]. A small clump of seaweed was kept in a 50 mL conical tube with distilled artificial seawater and oscillated to separate it into small particles with a mixer (VTX-3000L, LMS Co, Ltd, Tokyo, Japan) for 1 min. Healthy filaments without epiphytes or damage, measuring 1.5 mm in length were selected with a stereomicroscope (SZX12, Olympus Co. Ltd.) for the growth pattern experiment. Filaments were individually placed in a 6-well cell culture plate (approximately 4 cm in diameter per well) with 10 mL of medium. The plate was kept at room temperature (approximately 29–30°C) with a 12:12 h light: dark cycle and a photon flux density of 340 μmol m−2 s−1. Algal development was observed every morning under the stereomicroscope for 7 days.
Molecular sequencing
Samples were frozen in liquid nitrogen until use and DNA was extracted with a DNeasy Plant Mini Kit (Qiagen Sciences, Germantown, MD, USA) according to the manufacturer’s instructions. The isolated DNA was used for PCR amplification of ribosomal RNA genes using the 18S rDNA specific primers SR-1f (5′-TACCTGGTTGATCCTGCCAG-3′) and 18S-C2r (5′-TCCGCAGGTTCACCTACGGAG-3′) [29] and the 28S rDNA specific primers C1FL (5′-ACCCGCTGAACTTAAGCATATC-3′) and D2FL (5′-GGTCCGTGTTTCAAGACGG-3′) [20]. The PCR products were purified with a NucleoSpin Gel and PCR Clean-up Kit (Macherey-Nagel Inc., Düren, Germany) and subcloned into a pCR4-TOPO vector using a TOPO TA Cloning Kit for sequencing (Thermo Fisher Scientific Inc., Weltham, MA, USA). Sequencing was carried out by Eurofins Genomics K.K., Tokyo, Japan. The 18S rDNA and 28S rDNA sequences of this alga were submitted to the DNA Data Bank of Japan (DDBJ) and were searched for homology using the program BLAST at the DDBJ. This nucleotide sequence and the published sequences of other filamentous algae were aligned by Clustal W [29]. The Maximum likelihood (ML) and maximum parsimony (MP) algorithms were used to construct a phylogenetic tree. Bootstrap values were obtained from 500 resamplings. Cladophora herpestica (Z35419) (syn. Cladophoropsis zollingerii) and Cladophora sericea (Z35320) were used as out-groups for the 18S rDNA phylogenetic tree, and C. herpestica (AM503460) and C. sericea (AM503474) were used as out-groups for the 28S rDNA phylogenetic tree, based on Leliaert et al. [20].
Results
Chronology
A mass occurrence of spherical Cladophora aggregations was observed in a salt field reservoir in Samut Sakhon Province in July 2013 (Fig 1). Thereafter, only a few residual spherical aggregations were observed and all had disappeared by August. Spherical aggregation blooms do not occur every year, and have only been observed twice (2010 and 2013) in at least 50 years in this area, according to the pond owner (pers. comm. Mr. and Mrs. Yasathon).
Habitat
The reservoir where the mass of spherical aggregations occurred was approximately 70 m long, 50 m wide, and 40–50 cm deep. Water temperature, salinity, and pH at the reservoir were 32.8°C, 35 PSU, and 8.97, respectively. Extensive populations of spherical aggregations floated, accumulated, and colonized on the water surface close to the shore downwind in an area of approximately 560 m2 with a density of 706 aggregations m−2 (Fig 2). Aggregations were slightly rolling and trembling horizontally 132–142 times per min by wind-generated ripples (Table 1 and Fig 3 and S1 Movie). Individual aggregations were soft, solid, with irregularly entangled filaments, and dark green in color (Fig 4A and 4B). They ranged in diameter from 1.2–8.2 cm (mean 3.6 cm) along the major axis and 1.0–7.7 cm (mean 2.9 cm) along the minor axis (Fig 5). Mat-like forms with semi-spherical aggregations (not yet completely aggregated) were observed attached to or lying on the clay bottom offshore on the downwind side of the reservoir (S1 Movie). However, indeterminate turfed forms were observed loosely attached to or lying on the clay bottom offshore at the windward side of the reservoir, where there was almost no water movement (Fig 4C).
Fig 2. Mass floating spherical aggregations of Cladophora socialis in a salt field reservoir in Central Thailand.

Table 1. Outline of habitat and habit of several Cladophora socialis forms at the study site.
| Southwest side | Northeast side | |
|---|---|---|
| Aggregation form | Indeterminate turfed form | Floating spherical aggregation, mat-like forms with semi-spherical aggregation |
| Water surface | Calm and smooth | Ripple with wind |
| Water movement (times m−1) | ~0 | 132–142 |
| Position related to wind direction | Windward | Downwind |
| Water depth (cm) | 40–50 | 40–50 |
Fig 3. Transverse schematic diagram showing the location of several Cladophora socialis forms at the study site.
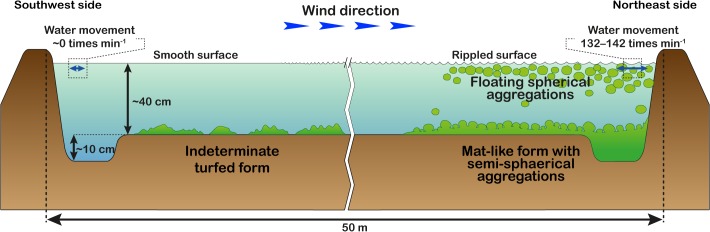
Fig 4. Floating spherical aggregations of Cladophora socialis in a salt field reservoir, a natural habitat.
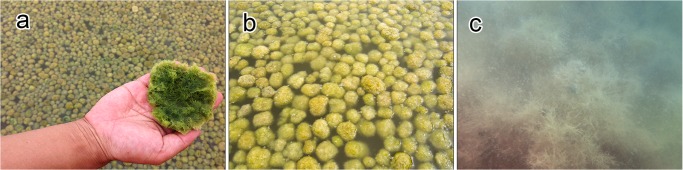
(a) A lower hemisphere of spherical aggregations of a solid, dark green color: (b) On-land view of floating spherical aggregations: (c) Underwater view of Indeterminate turf forms.
Fig 5. Scattergraph showing the size of spherical Cladophora socialis (major and minor axis) aggregations.

External and internal morphology
Individual filaments were green to light green and measured 2–3 cm in total length when fully developed (Fig 6A). Rhizoids were formed apically or intercalary and attached to another filament (Fig 6B); consequently, branches within a clump grew in different directions. The apical cells were cylindrical, round ended, 27.7–42.6 μm (mean = 36.0, SD ± 4.84) in diameter, 7.2–16.5 (9.44 ± 1.92) in length/diameter (L/D) ratio, and 0.9–2.6 μm (1.5 ± 0.41) in cell wall thickness (Fig 6C and 6D). The main axis cells were straight or slightly curved, cylindrical, 60.2–99.6 μm (72 ± 6.54) in diameter, 3.3–7.1 (4.8 ± 0.96) in length/diameter (L/D) ratio, and 1.7–4.6 μm (2.7 ± 0.62) in cell wall thickness (Fig 6E). Branching angles were normally 23.8–70.6° (47.7° ± 11.82), and were occasionally close to 180°, with single and sometimes double branching systems (Fig 6F and 6G). Newly formed laterals inclined to without cross-walls at their base (Fig 6H). The cells were multi-nucleate, with 12–38 (23.3 ± 6.65) nuclei in apical cells and 10–66 (23.5 ± 11.08) in main axis cells (Fig 6I). The nuclei major axes were 3.2–5.7 μm (4.2 ± 0.59) and 2.1–7.2 μm (3.6 ± 1.02) in apical and main axis cells, respectively. The chloroplasts were parietal along the cell wall (Fig 6J).
Fig 6. Morphological characteristics of Cladophora socialis.

(a) A fresh individual thallus: (b) Filaments attaching to one another by rhizoids formed apically (filled-arrow) and intercalary (open-arrow): (c) Apical cell in natural individual: (d) Apical cell cultured in static water showing longer length: (e) Main axis cells: (f) Branching system: (g) Filaments showing different growing directions because of a bent obtuse angle: (h) Newly formed lateral (filled-arrow) without cross-wall at its base: (i) Multi-nuclei in a cell stained by the Wittmann method: (j) Parietal chloroplasts along the cell wall: (k) Transmission electron microscopic image of bilenticular type pyrenoid: (l) Transmission electron microscopic image of zonal type pyrenoid.
Pyrenoid ultrastructure
The pyrenoids were spherical to ellipsoidal: 1.2–3.1 μm (2.3 ± 0.45) along the major axis, 0.8–3.1 μm (1.9 ± 0.48) along the minor, and 1.0–2.4 (1.3 ± 0.24) in L/D ratio. Two different types of pyrenoids were observed; approximately 80% were bilenticular, which was composed of a matrix traversed by a single thylakoid band surrounded by two cup-shaped starch sheaths (Fig 6K), and approximately 20% were zonal, in which the matrix was divided by two or more parallel thylakoid bands and surrounded by three or more starch sheaths (Fig 6L).
Growth pattern
Filamentary particles, 1.5 mm long, without branching (six cells) were cultured (Fig 7A). Lateral formations were observed at the apical pole of each of the four intermediate cells on the 2nd day (Fig 7C). The cell walls were formed at the basal portion of laterals, and the apical cell divided in two on the 3rd day (Fig 7D). The lateral cells extended, divided, and became branchlets on the 4th and 5th days (Fig 7E and 7F). Lateral formations were observed from branchlets on the 6th day (Fig 7G). The cell walls were formed at the basal portion of new laterals on the 7th day (Fig 7H). Thus, growth of this seaweed was by division of apical and lateral cells, which were produced at intermediate cells.
Fig 7. Growth of Cladophora socialis in laboratory experiments.

(a) Day 0: (b) Day 1: (c) Day 2: (d) Day 3: (e) Day 4: (f) Day 5: (g) Day 6: (h) Day 7.
Molecular sequencing analysis
The almost complete 18S rDNA sequence of this alga consisted of 1769 nucleotides. It was registered on DDBJ under the accession number AB971263. Homologies of 18S rDNA between this seaweed and other related seaweed sequences in the DDBJ were 99.5% in C. coelothrix from Philippines and 99.1% in C. socialis from Australia and Panama (Table 2 and Fig 8). The partial 28S rDNA sequence of this alga consisted of 608 nucleotides. It was registered on DDBJ under the accession number AB971264. Homologies of 28S rDNA between this seaweed and other related seaweed sequences in the DDBJ were 98.8% in C. socialis from South Africa, Jamaica, Panama, and Australia (Table 3 and Fig 8).
Table 2. Homology of Cladophora socialis 18S rDNA sequence to related species.
| Species | Voucher | Accession No. | Identities (bp) | Similarity (%) | Locality | Reference |
|---|---|---|---|---|---|---|
| C. socialis | SCORL 000358 | AB 971263 | 1769 | Samut Sakhon, Thailand | Present study | |
| C. coelothrix | F135 | AM498749 | 1679/1687 | 99.5 | Mactan Is., Philippines | [20] |
| C. socialis | F225 | AM498751 | 1673/1688 | 99.1 | Rottnest Is., Australia | [20] |
| C. socialis | F220 | AM498752 | 1671/1687 | 99.1 | Balboa, Panama | [20] |
| C. socialis | F224 | AM498753 | 1657/1688 | 98.2 | Virgin Is., USA | [20] |
| C. liebetruthii | Z35318 | 1728/1770 | 97.6 | Republic of Cabo Verde | [17] |
Fig 8. Maximum likelihood phylogenetic trees for Cladophora socialis and relatives.

(a) A tree based on nearly complete 18S rDNA sequences: (b) A tree based on partial 28S rDNA sequences. Other data on Cladophorales were downloaded from GenBank for comparison. Numbers close to the nodes are ML/MP bootstrap values.
Table 3. Homology of Cladophora socialis 28S rDNA sequence to related species.
| Species | Voucher | Accession No. | Identities (bp) | Similarity (%) | Locality | Reference |
|---|---|---|---|---|---|---|
| C. socialis | SCORL 000358 | AB971264 | 608 | Samut Sakhon, Thailand | Present study | |
| C. socialis | F197 | AM503477 | 561/568 | 98.8 | KwaZulu-Natal, South Africa | [20] |
| C. socialis | F173 | AM503476 | 561/568 | 98.8 | Portland, Jamaica | [20] |
| C. socialis | F200 | AM503440 | 561/581 | 98.8 | Balboa, Panama | [20] |
| C. socialis | F225 | AM503478 | 560/567 | 98.8 | Rottnest Is., Australia | [20] |
| C. socialis | F228 | AM503479 | 559/566 | 98.8 | unknown | [20] |
Discussion
Recent molecular phylogenetic studies have proven that Cladophora is not a monophyletic genus and that most of the morphological distinctions are untenable [13, 17, 19, 30]. Based on our phylogenetic analysis for both18S rDNA and 28S rDNA, the present species belongs to the Siphonocladales lineage and not Cladophorales. These results support the assertion that some members of the order Cladophorales should be reassigned to new genera and families [13].
It was impossible to determine if the study species is Cladophora coelothrix or C. socialis from our homology results, because the most closely related species were different in respective parts of the nucleic acid sequence, i.e., the study species was highly homologous to C. coelothrix and C. socialis in 18S rDNA and to C. socialis in 28S rDNA. C. socialis and C. coelothrix also closely resemble each other in external morphology; however, C. socialis can be distinguished from C. coelothrix by its smaller cell diameter, which is approximately half as thick [31–34]. Although the diameter range of the main axes overlapped slightly, that of the apical cells of the study species was much thinner than that of C. coelothrix (Table 4). Additionally, pyrenoid ultrastructure and growth pattern in the study species was consistent with that of C. socialis (Table 4). Thus, based on its external morphology, ultrastructure, growth pattern, and the molecular sequencing analysis, we conclude that the study species is C. socialis.
Table 4. Outline of the major morphological characteristics and habitats of Cladophora socialis and related Cladophora species.
| Cladophora socialis | C. coelothrix | C. socialis | ||||||
|---|---|---|---|---|---|---|---|---|
| External morphology | ||||||||
| Apical cells | ||||||||
| Diameter (μm) | 28–43 | 55–140 | 57–75 | 65–120 | 24–54 | 30–40 | 25–50 | 35–55 |
| L/D | 7–17 | 5–16 | 5–15 | 2.5–16 | 7–18 | 3–5 | 10–40 | 4–22 |
| Shape of terminal portion | Round | Round | Round | Round | Round | |||
| Cell wall thickness (μm) | 0.9–2.6 | 1–3 | 0.5 | < 1 | 1 | 0.5–1 | ||
| Main axis cells | ||||||||
| Diameter (μm) | 60–99 | 60–220 | 75–120 | 65–170 | 44–80 | 40–60 (70) | 35–55 | 35–85 |
| Ratio of Length to Diameter | 3–7 | 2.5–12 | 1.5–12 | 2–20 | 4–5 | 3–20 | 3–15 | |
| Cell wall thickness (μm) | 1.7–4.6 | < 15 | < 4 | 2–4 | < 8 | < 3 | ||
| Ramification | ||||||||
| Angle | 24–70 (180) | < 90 | 20–50 | > 45 | 45–90 | 45–90 | > 45 | |
| Internal morphology | ||||||||
| Chloroplast | ||||||||
| Position | Peripheral reticulate | |||||||
| Morphological form | Plate | |||||||
| Nuclei | ||||||||
| No. in apical cells | 12–38 | |||||||
| No. in main axis cells | 10–66 | |||||||
| Diameter in apical cells (μm) | 3–6 | |||||||
| Diameter in main axis cells (μm) | 2–7 | |||||||
| Ultrastructure | ||||||||
| Pyrenoid | ||||||||
| Diameter (μm) | 1–3 | |||||||
| Shape | Spherical, Ellipsoidal | |||||||
| Type | Bilenticular, Zonal | Bilenticular | ||||||
| Growth pattern | ACD at terminal cells and lateral division at intermediate cells | ACD | Mainly ACD, ICD lower down | Mainly ACD, ICD in shorter cells | Mainly ACD, ICD in shorter cells | ICD | ICD | Mainly ACD, ICD |
| Ecology | ||||||||
| Habitat | Stagnant water | Wave-exposed to mangrove areas | Intertidal to subtidal area | Shady places at sublittoral | Wave-exposed to mangrove areas | Intertidal, tidal pools | Intertidal areas | Sublittoral |
| Form | FSA, CM | T | CM | ASP, FF, | ||||
| Distribution | Atlantic, Mediterranean, Indo-Pacific | Tropical to warm temperate | Tropical to warm temperate | Atlantic, Mediterranean, Indo-Pacific | Tahiti (type locality), Hawaiian Is., Southern Japan, Europe | Tropical to warm temperate | Tropical to warm temperate | |
| Reference | Present study | [34] | [32] | [31] | [34] | [33] | [32] | [31] |
To our knowledge, this is the first record of C. socialis having a floating spherical aggregation form, as previous reports have only described delicate felts a few mm high growing on rocks and crustose corallines, high mats and pompons growing on intertidal rocks, and fluffs floating in salt marsh and mangrove lagoons [31, 32].
Aegagropila linnaei can occur in several different forms, such as a mat-like attached form, a floating unattached mat form, free-living filaments, and spherical aggregations depending on environmental conditions [4–6, 35–37]. In A. linnaei the ball shape is the result of both mechanical factors, such as wave-induced rolling motion, and morphological features leading to entanglement of the filaments [1, 6, 35, 37]. In this study, different C. socialis growth forms were also observed in different water motion environments, e.g., floating spherical aggregations and mat-like forms with semi-spherical aggregations were observed on the shore on the downwind side where there was water movement. Externally, members of the study species were entangled in each other; consequently, branches growing in different directions were found within a clump. Similar to A. linnaei, our results suggest that water movement and morphological characteristics promote the formation of spherical aggregations in C. socialis (Fig 3). Detailed surveys of the physical underwater environment and laboratory culture experiments will be required in the future to clarify the mechanism underlying spherical aggregation formation.
Wind is one of the main factors causing movement in stagnant waters that have no regular in- or outflow. We suspect that wind direction does not influence water movement (which induces spherical aggregation) at the study site because the wind direction was same in July, when the spherical aggregations occurred, as it was in August when they disappeared (Fig 9). However, differences in wind velocity are more likely to influence water movement and spherical aggregation formation because wind velocity in July (11 m s−1) was different to that in August (8 m s−1). Additionally, wind velocity was fastest in July that year (Fig 9) [38]. Thus, faster wind velocity induces stronger water movement, and stronger water movement may promote spherical aggregation formation in this seaweed. However, according to the owner of this reservoir, mass occurrence of spherical aggregations does not occur every year and is a very rare phenomenon. Because their occurrence is sudden, short-term, and rare, it is unlikely that spherical aggregations only occur because of water movement promoted by wind; other physical conditions are most likely involved.
Fig 9. Monthly maximum velocity (m s−1) and wind direction in Central Thailand.

Graphs were drawn from raw data provided by the Thai Meteorological Department [38].
Although Cladophora is the largest green algae genus [13, 14], information on their internal morphology and ultrastructure is limited [22, 23, 39–44]. Cladophora pyrenoid ultrastructure is divided into four types; bilenticular, zonal, simple polypyramidal, and complex polypyramidal [23]. The bilenticular type, which is composed of a matrix traversed by a single thylakoid band, has been reported in C. socialis [31]. Additionally, a zonal type in which the pyrenoid matrix is divided by parallel thylakoid bands was observed by TEM for the first time in this study. Both of these pyrenoids are common in Cladophora species.
Morphological classification does not necessarily always reflect molecular phylogenetic analysis [20, 45]. For example, C. coelothrix is not a monophyletic taxon, and it is distributed in a number of different clades in a phylogenetic tree [13, 20]. Thus, it is occasionally difficult for species level identification in Cladophora using part of a gene, such as the 18S rDNA and 28S rDNA nucleic sequences, because of limitations in classification sensitivity. The authors consider it important to develop whole genome identification methodologies in the future, such as the amplified fragment length polymorphism (AFLP) method, for more exact Cladophora species identification.
Conclusion
A mass occurrence of spherical Cladophora aggregations appeared for a short time in a salt field reservoir during July 2013 in Central Thailand. Extensive populations of spherical aggregations floated, accumulated, and colonized the water surface offshore where there was water movement. Additionally, individual filaments within the aggregations were entangled in each other, consequently branches growing in different directions were found within a clump. We suggest that water movement and morphological characteristics promote the formation of the spherical aggregations. Based on its external morphology, ultrastructure, growth pattern, and the molecular sequencing analysis, we conclude that the species in question is C. socialis. This is the first record of C. socialis having a floating spherical aggregation form.
Supporting Information
S1 Movie. Floating Cladophora socialis spherical aggregations and mat-like forms with semi-spherical aggregations (underwater).
These forms were observed offshore on the downwind side of a salt field reservoir.
(AVI)
Acknowledgments
The authors would like to sincerely thank Dr. Kazuyo Matsuyama-Serisawa and Dr. Yukihiko Serisawa of Yamanashi University, Japan for useful comments on Cladophora morphology, and Dr. Isamu Wakana of The Akankohan Eco Museum Center, Japan for providing information on spherical aggregations. We are grateful to Mr. Wonelop Yasathon and Mrs. Raruen Yasathon, the pond owner, for informing us on the history of spherical aggregations in the study site. We thank the National Research Council of Thailand (NRCT) for providing useful advice to conduct this study in Thailand. The authors also greatly appreciate Dean Sakchai Choochote, and the Head of Fisheries group, Dr. Paveena Taveekijakarn, Faculty of Agricultural Technology, KMITL for their helpful advice. We thank Dr. Masayoshi Saito, Program Director, JIRCA for his constructive comments.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This study was funded and performed as part of the international research project “Development of aquaculture technologies for suitable and equitable production of aquatic products in tropical coastal areas” conducted by the Faculty of Agricultural Technology, KMITL and the Fisheries Division, JIRCAS. The funding bodies had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Boedeker C, Eggert A, Immers A, Smets E (2010) Global decline of and threats to Aegagropila linnaei, with special reference to the lake ball habit. BioScience 60: 187–198. [Google Scholar]
- 2.Mathieson AC, Dawes CJ (2002) Chaetomorpha balls foul New Hampshire, USA beaches. Algae 17: 283–292. [Google Scholar]
- 3.Berry S (29 Sep 2014) UFOs on Sydney beach. Yahoo News. Available: http://au.news.yahoo.com/nsw/a/25061829/ufos-on-sydney-beach/. Accessed 6 December 2014.
- 4.Togashi T, Sasaki H, Yoshimura J (2014) A geometrical approach explains lake ball (marimo) formations in the green alga, Aegagropila linnaei. Sci Rep 4: 1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Boedeker C, Immers A (2009) No more lake balls (Aegagropila linnaei Kützing, Cladophoraphyceae, Chlorophyta) in the Netherlands? Aquat Ecol 43: 891–902. [Google Scholar]
- 6.Kurogi M (1980) Lake ball “Marimo” in Lake Akan. Jap J Phycol 28: 168–169. [Google Scholar]
- 7.Hasegawa Y (1978) Occurrence of “Seawater Marimo” at seashore. Hokkaido Natl Fish Res Inst (Jpn) 19: 5. (in Japanese) [Google Scholar]
- 8.van den Hoek C, Ducker SC, Womersley HBS (1984) Wittrockiella salina Chapman (Cladophorales, Chlorophyceae), a mat and ball forming alga. Phycologia 23: 39–46. [Google Scholar]
- 9.Mathieson AC, Hehre EJ, Dawes CJ (2000) Aegagrophilous Desmarestia aculeata from New Hampshire. Rhodora 102: 202–207. [Google Scholar]
- 10.Burrows EM (1991) Seaweeds of the British Isles London: British Museum (Natural History). 238 p. [Google Scholar]
- 11.Bach S, Josselyn MN (1978) Mass blooms of the alga Cladophora in Bermuda. Mar Pollut Bull 9: 34–37. [Google Scholar]
- 12.Littler DS, Littler MM, Bucher KE, Norris JN (1989) Marine plants of the Caribbean Washington DC: Smithsonian Institution Press. 263 p. [Google Scholar]
- 13.Leliaert F, Boedeker C (2007) Cladophorales In: Brodie J, Maggs CA, John DM editors. The Green Seaweeds of Britain and Ireland. London: The British Phycological Society; pp. 131–183. [Google Scholar]
- 14.Guiry MD, Guiry GM (2014) AlgaeBase World-wide electronic publication, National University of Ireland, Galway: Available: http://www.algaebase.org. Accessed 8 October 2014. [Google Scholar]
- 15.Sakai Y (1964) The species of Cladophora from Japan and its vicinity Scientific papers of the Institute of Algalogical Research, Faculty of Science, Hokkaido University. 104 p. [Google Scholar]
- 16.Niiyama Y (1986) Morphological phenology of Cladophora glomerata (L.) Kützing in Hokkaido. Jap J Phycol 34: 216–224. [Google Scholar]
- 17.Bakker FT, Olsen JL, Stam WT, Hoek C van den (1994) The Cladophora complex (Chlorophyta): New views based on 18S rRNA gene sequences. Mol Phylogenet Evol 3: 365–382. [DOI] [PubMed] [Google Scholar]
- 18.Matsuyama K, Tanaka J (2000) Cladophoracean algae (Ulvophyceae, Chlorophyta) of the Imperial Palace, Tokyo. Mem Natn Sci Mus Tokyo 34: 139–146. [Google Scholar]
- 19.Leliaert F, Rousseau F, de Reviers B, Coppejans E (2003) Phylogeny of the Cladophorophyceae (Chlorophyta) inferred from partial LSU rRNA gene sequences; is the recognition of a separate order Siphonocladales justified? Eur J Phycol 38: 233–246. [Google Scholar]
- 20.Leliaert F, de Clerck O, Verbruggen H, Boedeker C, Coppejans E (2007) Molecular phylogeny of the Siphonocladales (Chlorophyta; Cladophorophyceae). Mol Phylogenet Evol 44: 1237–1256. [DOI] [PubMed] [Google Scholar]
- 21.Thiamdao S, Boo GH, Boo SM, Peerapornpisai Y (2012) Diversity of edible Cladophora (Cladophorales, Chlorophyta) in northern and northeastern Thailand, based on morphology and nuclear ribosomal DNA sequences. Chiang Mai J Sci 39: 300–310. [Google Scholar]
- 22.Ichihara K, Shimada S, Miyaji K (2013) Systematics of _Rhizoclonium_-like algae (Cladophorales, Chlorophyta) from Japanese brackish waters, based on molecular phylogenetic and morphological analysis. Phycologia 52: 398–410. [Google Scholar]
- 23.Matsuyama K, Matsuoka T, Miyaji K, Tanaka J, Aruga Y (1998) Ultrastructure of the pyrenoid in the family Cladophoraceae (Cladophorales, Chlorophyta). J Jpn Bot 73: 279–286. [Google Scholar]
- 24.Miyaji K (1999) A new type of pyrenoid in the genus Rhizoclonium (Cladophorales, Chlorophyta). Phycologia 38: 267–276. [Google Scholar]
- 25.Matsuyama-Serisawa K, Yasuda T, Nakano T. Serisawa Y (2009) Rediscovery of Aegagropila sauteri var. yamanakaensis Okada from Lake Yamanaka. Mount Fuji Research 3: 13–18. [Google Scholar]
- 26.Wittmann W (1965) Aceto-iron-haematoxylin-chloral hydrate for chromosome staining. Stain Tech 40: 161–164. [DOI] [PubMed] [Google Scholar]
- 27.Provasoli L (1966) Media and prospects for the cultivation of marine algae In: Watanabe A, Hattori A (ed) Culture and collections of algae. Japanese Society of Plant Physiologists, Tokyo: pp. 63–75. [Google Scholar]
- 28.Lewin J (1966) Silicon metabolism in diatoms. V. germanium dioxide, a specific inhibitor of diatom growth. Phycologia 6: 1–12. [Google Scholar]
- 29.Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignments through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673–4680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hanyuda T, Wakana I, Arai S, Miyaji K, Watano Y, Ueda K (2002) Phylogenetic relationships within Cladophorales (Ulvophyceae, Chlorophyta) inferred from 18S rRNA gene sequences, with special reference to Aegagropila linnaei. J Phycol 38: 564–571. [Google Scholar]
- 31.Hoek C van den, Chihara M (2000) A taxonomic revision of the marine species of Cladophora (Chlorophyta) along the coasts of Japan and the Russian far-east. Nat Sci Mus monographs 19: 242 pp. [Google Scholar]
- 32.Leliaert F, Coppejans E (2003) The marine species of Cladophora (Chlorophyta) from the South African east coast. Nova Hedwigia 76: 45–82. [Google Scholar]
- 33.Abbott IA, Huisman JM (2004) Marine green and brown algae of the Hawaiian Islands. Honolulu: Bishop Mus Bull Bot 4: 260. [Google Scholar]
- 34.Dawes CJ, Mathieson AC (2008) The seaweeds of Florida Florida: University Press of Florida. 592 p. [Google Scholar]
- 35.Niiyama Y (1989) Morphology and classification of Cladophora aegagropila (L) Rabenhorst (Cladophorales, Chlorophyta) in Japanese lake. Phycologia 28: 70–76. [Google Scholar]
- 36.Sano O, Arai S, Watanuki A, Ogino K, Wakana I, Yokohama Y (1994) Distribution of Cladophora aegagropila around Churui Island in Lake Akan and the environmental characteristic for development of the ‘Marimo’ aggregation. Marimo Res 3: 26–30. [Google Scholar]
- 37.Boedeker C (2010) The attached form of the endangered freshwater alga Aegagropila linnaei Kützing (Chlorophyta) is found in the Zuideindigerwiede, The Netherlands. Aquat Bot 92: 75–77. 10.3354/dao02263 [DOI] [PubMed] [Google Scholar]
- 38.Thai Meteorological Department (2014) Climatological data for the period 2013. Internal database (unpublished).
- 39.Gibbs SP (1962) The ultrastructure of the pyrenoids of green algae. J Ultrastructure Res 7: 262–272. [DOI] [PubMed] [Google Scholar]
- 40.Hori T, Ueda R (1967) Electron microscope studies on the fine structure of plasmids in siphonous green algae with special reference to their phylogenetic relationships. Sci Rep Tokyo Kyoiku Daigaku 12: 225–244. [Google Scholar]
- 41.Scott JL, Bullock KW (1976) Ultrastructure of cell division in Cladophora. Pregametangial cell division in the haploid generation of Cladophora flexuosa. Can J Bot 54: 1546–1560. [Google Scholar]
- 42.Chan KY, Ling Wong SL, Wong MH (1978) Observations on Chaetomorpha brachygona Harv. (Chlorophyta, Cladophorales). I Ultrastructure of the vegetative cells. Phycologia 17: 419–429. [Google Scholar]
- 43.Wang XY (1989) Observations on pyrenoid ultrastructure of Cladophora conchopheria (Chlorophyceae). Jap J Phycol 37: 57–59. [Google Scholar]
- 44.Boedeker C, O’Kelly CJ, Star W, Leliaert F (2012) Molecular phylogeny and taxonomy of the Aegagropila clade (Cladophorales, Ulvophyceae), including the description of Aegagropilopsis gen. nov. and Pseudocladophora gen. nov. J Phycol 48: 808–825. 10.1111/j.1440-1754.2011.02203.x [DOI] [PubMed] [Google Scholar]
- 45.Cocquyt E, Verbruggen H, Leliaert F, De Clark O (2010) Evolution and cytological diversification of the green seaweeds (Ulvophyceae). Mol Biol Evol 27: 2052–2061. 10.1093/molbev/msq091 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
S1 Movie. Floating Cladophora socialis spherical aggregations and mat-like forms with semi-spherical aggregations (underwater).
These forms were observed offshore on the downwind side of a salt field reservoir.
(AVI)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.