Ultraviolet light illuminates species-specific biofluorescent casque patterns in cassowaries (Casuarius) (original) (raw)
Introduction
Cranial ornaments are present in all major clades of vertebrates, yet their specific functional roles often remain unclear. Dinosaurs, including modern birds, are well-known for their ornate features composed of soft (feathers or flesh), hard (bone), or multiple tissue types1,2,3. These features include plumage coloration (e.g., dark feathers near bright regions4) and exaggerated anatomy (e.g., cranial ornaments1) which often feature in behavioral displays (e.g., courtship dances5). Compared to other vertebrates, modern birds exhibit an especially high degree of visual acuity6, which is thought to have influenced the evolution of prominent display structures. Not only do birds see into the color spectrum of humans (Homo sapiens; 400–700 nm), but many birds possess ultraviolet (UV)-capable, short-wavelength-sensitive type 1 opsin pigments and UV/V cones7or increased sensitivity of their rod vision8that also allow them to perceive UV light (320–400 nm9). It has been suggested that birds exploit this ability (see10and references therein) to locate prey (e.g., Falco tinnunculus11), aid in foraging [e.g., _Leptotila cassinii_12), and highlight themselves with UV-visible plumage or rhamphothecae (e.g., Fratercula arctica13; Cerorhinca monocerata14; Melopsittacus undulatus15; Asio otus7; Otus scops16; various Passeriformes17,18). Considering its presence in integumentary structures and the commonality of UV sensitivity among living birds (i.e., approximately half of all bird orders6,19,21), it is likely that some aspects of their visual prowess may have played an important role in the evolution of intraspecific signaling and other visual displays across archosaurs21.
Both UV biofluorescence (i.e., absorption of short-wavelength light that initiates excitation of a biomolecule and translates to emission of long wavelength light) and UV reflectance (i.e., bouncing of short-wavelength light off of a surface) can be important metrics when assessing organisms in an ecological context. Biofluorescence may allow researchers to find natural patterns that are otherwise incapable of being seen in the visible spectrum. For example, wing feathers of the northern saw-whet owl (Aegolius acadicus) and the American barn owl (Tyto furcata) are biofluorescent—the most intensely fluorescent feathers being those that are the most recently grown22. That has proved important for assessing molt patterns and determining the relative age of individual owls22. Although biofluorescence can be biologically relevant, it is also important to consider that many tissues can exhibit the property, and careful assessment should be considered in order to identify those patterns that are relevant to an organism’s life history23. In addition to potentially strong absorption of UV wavelengths, biofluorescence also weakly re-emits fluorescent wavelengths23. Such UV visual signals can be visualized in aquatic environments, but less so in many terrestrial environments, illustrating the importance of considering the background environment of organisms (see23and references therein). Despite biofluorescence studies of integumentary structures becoming more common, a number of avian studies (e.g.13,14), describe UV biofluorescence without considering the UV reflectance under in vivo conditions (see23,24). By considering reflectance, we gain a better understanding of the anatomical regions that are related to how UV wavelengths can reach the eyes of an observer. Because it is difficult to decipher whether UV visualization is biologically relevant to specific organisms, Marshall and Johnsen24established a checklist for investigators to determine the (1) visible location of a fluorescent compound, (2) excitation and emission ranges of the structure, (3) sensitivity ranges of observing organisms, (4) natural lighting conditions, and (5) behaviors that may benefit from a fluorescence structure. An investigation of facial and crown feathers of budgerigar (M. undulates) provides an example of results which satisfy these requirements15. Our understanding of how the UV reflectance of anatomical structures may play a role biologically or evolutionarily is reliant upon studies that provide a holistic look at individual avian taxa with such scrutiny considered.
Among extant birds, cassowaries (i.e., large palaeognathous birds native to Australasia) are especially notable for their brightly colored integument in the human-visible spectrum. These include vividly colored apteria (e.g., blue, red, yellow, orange, purple, pink, white), carunculated skin, and pendulous wattles (see25). These traits are visually distinct enough to differentiate the three modern cassowary species (dwarf, Casuarius bennetti; southern, C. casuarius; northern, C. unappendiculatus), and it is presumed that cassowaries recognize these differences in congeners as well25. Cassowaries also possess tall cranial ornamentations called casques, which feature prominently as a dorsal protrusion of the skull sheathed by keratin. Casques have also repeatedly been implicated in visual displays (e.g., dominance, sexual differentiation, species recognition25,26,28,43). However, their dull colors (e.g., green, brown, grey, black) compared to brightly colored apteria and wide variations in form (e.g.28,29), have undermined such conjecture in the eyes of researchers, leaving their potential display role as an enigma (Fig. 1;25,[30](/articles/s41598-026-40230-1#ref-CR30 "Green, T. L. & Gignac, P. M. Osteological comparison of casque ontogeny in palaeognathous and neognathous birds: implications for selecting modern analogs in the study of cranial ornaments from extinct archosaurs. Zool. J. Linn. Soc. https://doi.org/10.1093/zoolinnean/zlad016]
(2023).")). Recent studies have hypothesized the numerous ways in which the casque could be used as a signal structure via size or shape over ontogeny[35](/articles/s41598-026-40230-1#ref-CR35 "Farke, A. A., Chok, D. J., Herrero, A., Scolieri, B. & Werning, S. Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids. PeerJ 1, e182 (2013)."),[36](/articles/s41598-026-40230-1#ref-CR36 "Lü, J. et al. High diversity of the Ganzhou oviraptorid fauna increased by a new cassowary-like crested species. Sci. Rep. 7, 1–13 (2017)."), within species[27](/articles/s41598-026-40230-1#ref-CR27 "Crome, F. H. J. The cassowary’s casque. Emu 88, 123–124 (1988)."),[29](/articles/s41598-026-40230-1#ref-CR29 "Perron, R. M. Taxonomy of the Genus Casuarius: the Defined and Known Living Cassowary Species and Subspecies (United Kingdom: Quantum Conservation,, 2016)."), and between species[28](/articles/s41598-026-40230-1#ref-CR28 "Green, T. L., Kay, D. I. & Gignac, P. M. Intraspecific variation and directional casque asymmetry in adult Southern cassowaries (Casuarius casuarius). J. Anat. 241, 951–965 (2022).")rather than color alone. Nonetheless, speculation about the still relatively poorly understood cassowary casque biology has also fueled interpretations of cranial ornament functions in non-avian dinosaurs, such as oviraptorosaurs and hadrosaurs (e.g.[31](#ref-CR31 "Green, T. L. & Gignac, P. M. Osteological description of casque ontogeny in the Southern cassowary (Casuarius casuarius) using micro-CT imaging. Anat. Rec. 304, 461–479 (2021)."),[32](#ref-CR32 "Naish, D. & Perron, R. Structure and function of the cassowary’s casque and its implications for cassowary history, biology and evolution. Hist. Biol. 28, 507–518 (2016)."),[33](#ref-CR33 "Dodson, P. Taxonomic implications of relative growth in lambeosaurine hadrosaurs. Syst. Biol. 24, 37–54 (1975)."),[34](/articles/s41598-026-40230-1#ref-CR34 "Hone, D. W., Naish, D. & Cuthill, D. I. C. Does mutual sexual selection explain the evolution of head crests in pterosaurs and dinosaurs? Lethaia 45, 139–156 (2012).")). Cassowaries are the largest, extant tropical birds and live in the Wet Tropics of New Guinea and northeastern Australia. Generally found in the swaths of rainforest with thick foliage, they feed on the fruits of hundreds of local plants and are recognized ecologically as primary seed dispersers[37](#ref-CR37 "Stocker, G. C. & Irvine, A. K. Seed dispersal by cassowaries (Casuarius casuarius) in North Queensland’s rainforests. Biotropica, 170–176 (1983)."),[38](#ref-CR38 "Mack, A. L. Distance and non-randomness of seed dispersal by the Dwarf cassowary Casuarius bennetti. Ecography 18, 286–295 (1995)."),[39](#ref-CR39 "Webber, B. L. & Woodrow, I. E. Cassowary frugivory, seed defleshing and fruit fly infestation influence the transition from seed to seedling in the rare Australian rainforest tree, Ryparosa sp. nov. 1 (Achariaceae). Funct. Plant. Biol. 31, 505–516 (2004)."),[40](#ref-CR40 "Bradford, M. G. & Westcott, D. A. Consequences of Southern cassowary (Casuarius casuarius, L.) gut passage and deposition pattern on the germination of rainforest seeds. Austral Ecol. 35, 325–333 (2010)."),[41](/articles/s41598-026-40230-1#ref-CR41 "Bradford, M. G. & Westcott, D. A. Predation of cassowary dispersed seeds: is the cassowary an effective disperser? Integr. Zool. 6, 168–177 (2011)."). Generally solitary outside of the breeding season, cassowaries are notorious for their territoriality and exhibit both ritualized visual displays and aggressive behaviors[42](/articles/s41598-026-40230-1#ref-CR42 "Crome, F. H. J. Some observations on the biology of the cassowary in Northern Queensland. Emu 76, 8–14 (1976)."). Anatomical features and physical displays both have been suggested to be important visually to cassowary life history and their evolution (see[28](/articles/s41598-026-40230-1#ref-CR28 "Green, T. L., Kay, D. I. & Gignac, P. M. Intraspecific variation and directional casque asymmetry in adult Southern cassowaries (Casuarius casuarius). J. Anat. 241, 951–965 (2022).")).Fig. 1

The alternative text for this image may have been generated using AI.
Exemplar adult casque biofluorescence among live cassowaries. (a, b) Casuarius casuarius (rostrolateral), (c, d) Casuarius unappendiculatus rothchildi (right lateral), and (e, f) Casuarius unappendiculatus rufotinctus (left lateral) under (a, c, e) visible and (d) 365 nm and (b, f) 385–395 nm ultraviolet light with regions of keratin biofluorescence evident in light blue and green. To accurately depict in vivo biofluorescence visually, the contrast and color balance are accentuated with a yellow filter in figure images at 385–395 nm (note: no adjustments were made to images used for data collection and analysis). Photos: T.L. Green.
Like most birds, cassowaries and other palaeognaths (i.e., the ancient lineage of birds that includes tinamous and ratites) possess retinal pigments for UV sensitivity20, 44. One conjecture that has not been evaluated is whether cassowary cranial ornaments are fluorescent and reflective in the UV spectrum. Ultraviolet coloration of casque keratin would enable the casque to be as visibly distinct as other display traits, like apteria and wattles. Further, this would implicate the casque as serving a primary function on par with these more conventional visual traits. In this study, we test the hypothesis that if cranial ornaments do serve important conspecific or congeneric display functions, they should be as visually distinct as other brightly colored integumentary features. We, therefore, predict that cassowary casque keratin is UV biofluorescent and reflective, which would implicate a display function by rendering the casque easily identifiable by other cassowaries. Despite a clear reliance on visual and UV-based cues by birds, previous studies have not tested biofluorescent properties of the keratinous sheathing on bony cranial ornaments. Cassowaries serve an excellent example for this purpose because they have large casques with high surface areas, and they are routinely referenced as a modern analog for the cranial ornaments of non-avian dinosaurs due to their relatively large body mass, flightlessness, and taxonomic placement compared to other living birds (e.g.34, 35, 36, 37, 31). Indeed, addressing the potential for UV coloration in cassowary casques would establish an important biological baseline for the potential display capacities in other birds and extinct dinosaurs with keratin-sheathed bony ornaments. Here, we determine whether the cassowary keratinous casque sheathing is (1) UV biofluorescent, (2) indicative of UV fluorescence within a range visible to cassowaries, (3) UV reflective, and (4) whether this could be used to accentuate visually-based biological roles of casques.
Results
UV biofluorescence
Our results indicate that casque keratin in Casuarius is distinctly UV fluorescent at 365 nm and within 385–395 nm (Figs. 1 and 2). Significant differences in biofluorescent casque coverage are not detected when comparing these tested wavelengths. In addition, correlation analyses on percent coverage of UV fluorescence were conducted and showed strong correspondence between wavelengths (R2 > 0.994; p < 0.0001). Thus, we report only the values for 385–395 nm in this section. In addition, the biofluorescent casque coverage and patterns differ between and within species (Table 1, Supplementary Table 1, Fig. 3). Only one C. bennetti individual in our sample possessed (minimal) casque biofluorescence in lateral view (percent of total lateral coverage of 0.80% at 385–395 nm), while all others were non-fluorescent. All C. casuarius and C. unappendiculatus individuals exhibited biofluorescence in both lateral and rostral views ranging from 8.77 to 93.69%.
Fig. 2

The alternative text for this image may have been generated using AI.
Casque biofluorescence patterns differences between Casuarius species. (a) Casuarius bennetti, (b) Casuarius casuarius, (c) Casuarius unappendiculatus rothchildi, and (d) Casuarius unappendiculatus rufotinctus preserved dry specimens in left lateral view under 385–395 nm ultraviolet light (insets in upper left illustrate the animals under visible light; artwork: J. A. Campbell-Smith). (e) C. bennetti (pastel purple), (f) C. casuarius (pastel green), and (g, h) C. unappendiculatus (pastel yellow) cranial line diagrams with typical casque biofluorescence patterns (dark blue-green regions) in lateral (left) and rostral (right) views. To accurately depict in vivo biofluorescence visually, the contrast and color balance are accentuated with a yellow filter in figure images at 385–395 nm (note: no adjustments were made to images used for data collection and analysis). Scale bar, 10 cm.
Table 1 Means and Kruskal-Wallis test results for Casuarius casque UV biofluorescence coverage.
Fig. 3
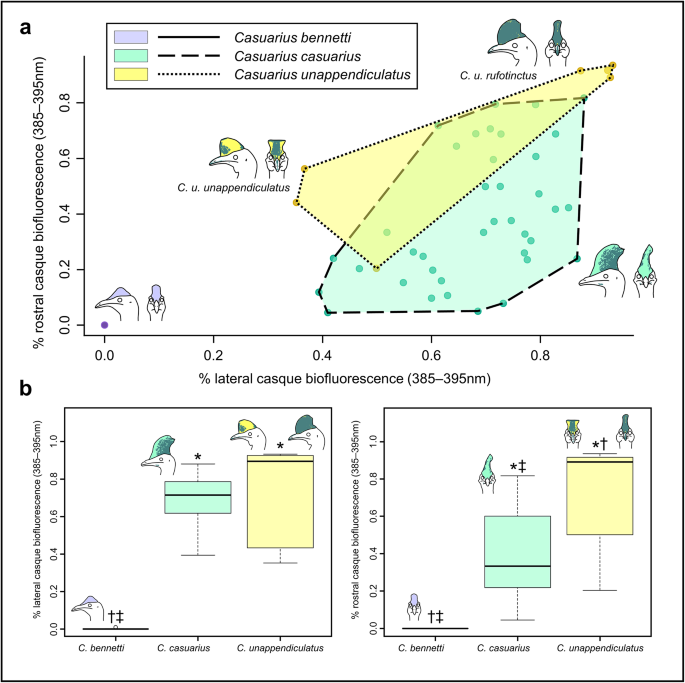
The alternative text for this image may have been generated using AI.
Casque biofluorescence comparison between Casuarius species. (a) Morphospace of casque biofluorescence percentage for lateral versus rostral views between Casuarius species (C. bennetti = pastel purple, C. casuarius = pastel green, and C. unappendiculatus = pastel yellow datapoints and polygons). Convex hulls illustrate substantial overlap in fluorescence coverage between C. casuarius (n = 38) and C. unappendiculatus (n = 7), and no overlap between C. bennetti (n = 30) and the other two species. The broad variation in the C. unappendiculatus convex hull is likely explained by differences in biofluorescent coverage between subspecies; the most distinctive being between C. u. unappendiculatus and C. u. rufotinctus. (b) Box and whisker plots specifying differences in lateral (left; C. bennetti, n = 32; C. casuarius, n = 45; C. unappendiculatus, n = 8) and rostral (right; C. bennetti, n = 30; C. casuarius, n = 39; C. unappendiculatus, n = 7) casque biofluorescence (385–395 nm) between species;* = significantly different from C. bennetti; † = significantly different from C. casuarius; ‡ = significantly different from C. unappendiculatus (significance based on Kruskal-Wallis test results, α = 0.05); means represented by solid black, horizontal lines.
We obtained relative casque size data to determine biofluorescence coverage of each casque surface, but did not observe any noticeable pattern in degree of biofluorescence coverage for absolute casque size in this adult-only sample. The average biofluorescent coverage and standard deviation at 385–395 nm in lateral view was 0.03 ± 0.14% for C. bennetti, 69.46 ± 12.30% for C. casuarius, and 72.24 ± 26.74% for C. unappendiculatus. In rostral view at 385–395 nm, coverage was 0.00 ± 0.00% for C. bennetti, 39.14 ± 23.35% for C. casuarius, and 69.51 ± 29.35% for C. unappendiculatus. We identify significant differences (p < 0.05) in these biofluorescence patterns between all three species (Table 1). Specifically, C. bennetti and C. casuarius differ in both lateral and rostral views, C. bennetti and C. unappendiculatus differ in both lateral and rostral views, and C. casuarius and C. unappendiculatus differ in only rostral view.
In addition to the percentage of biofluorescent casque coverage differing between species, the biofluorescence in specific casque regions differs interspecifically. The casques of C. bennetti are most frequently non-fluorescent (Figs. 2a and e and 3) with one individual (AMNH SKIN 268350) possessing a small patch of keratinous biofluorescence on the right caudolateral region of the casque. The casques of C. casuarius specimens exhibit biofluorescence most frequently on their caudolateral and caudal regions (Fig. 2b, f; Supplementary Fig. 2). We find two patterns exhibited by the casques of C. unappendiculatus specimens: (1) patchy distribution of biofluorescence on just the rostrolateral and dorsal regions, or (2) almost completely (i.e., > 80% coverage in lateral and rostral views) biofluorescent (Figs. 2c, d, g and h and 3). We describe the implications of these two patterns in the discussion section below.
We document biofluorescence in only specific regions of casque keratin and rhamphotheca that are either brown, green, or yellow under visible light. This suggests that the color of the keratin under visible light is a good indicator for which regions of the casque are UV fluorescent (Figs. 1 and 2). For example, casque keratin of C. bennetti individuals is typically black or dark grey under visible light (and non-fluorescent). The casque keratin of C. casuarius individuals under visible light is typically black or dark grey in the rostrolateral and dorsal portions (i.e., non-fluorescent), and brown and green in the caudolateral and caudal portions of the casque (i.e., fluorescent). The casque keratin of C. unappendiculatus individuals under visible light is green or brown in the rostrolateral and dorsal portions (i.e., fluorescent), and brown (i.e., fluorescent) or dark grey or black (i.e., non-fluorescent) in the caudolateral and caudal portions. In addition to the casque keratin, we observed biofluorescence in rhamphotheca in regions that were yellow, green, or brown under visible light for all three species (Fig. 1); however, this characteristic was highly variable among individuals. Prominently fluorescent under UV light, yellow (i.e., under visible light) malar patches of keratin on the proximolateral portions of the lower bill were observed in most C. casuarius individuals (Fig. 1b) and some C. unappendiculatus individuals (Fig. 1d).
In live cassowaries (Fig. 1), the casque biofluorescence coverages are congruent with the averages and patterns determined by the preserved specimen sample (e.g., average casque coverages of live birds fell within the range of those of preserved birds and patterns appeared similar; Supplementary Table 1). Live C. casuarius individuals possess average lateral biofluorescence coverages of 64.92 ± 24.75% (n = 6) and rostral biofluorescence coverages of 24.75 ± 16.98% (n = 3) under 385–395 nm light. Live C. unappendiculatus individuals possessed average lateral biofluorescence coverages of 55.43 ± 26.51% (n = 3) and rostral biofluorescence coverages of 61.85 ± 23.69% (n = 3) under 385–395 nm light. The keratinous rhamphotheca in live S. camelus and D. novaehollandiae did not exhibit biofluorescence (Supplementary Fig. 1).
UV reflectivity
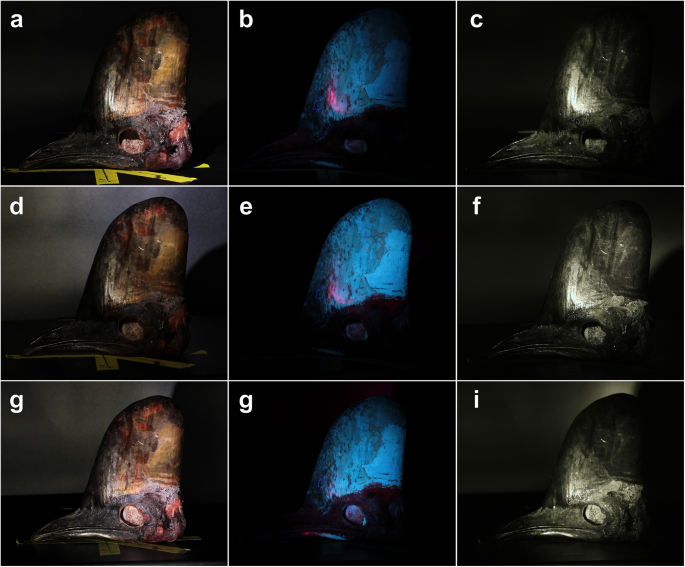
In addition to UV biofluorescence (Figs. 4b, e and h and 5b, e and h), we also find that the casque surfaces of the three C. casuarius individuals exhibit UV reflectance (Figs. 4c, f and i and 5c, f and i). The UV-biofluorescent patterns present with the non-modified camera are not evident in the UV-reflective areas of the casque with the UV modified camera. The 365 nm reflectance pattern of the casque surface was more similar to that which was observed under visible light.
Fig. 4

The alternative text for this image may have been generated using AI.
Adult southern cassowary (Casuarius casuarius) cranial specimen (eyes removed via dissection; TLG C069) in visible light (a, d, g) and 365 nm (b, c, e, f, h, i). Photos were taken with a visible light camera (a, b, d, e, g, h) and a UV-sensitive camera with a 365 nm band pass filter (c, f, i). The head is rotated at 90° (a–c), 45° (d–f), and 0° (g–i).
Fig. 5
The alternative text for this image may have been generated using AI.
Adult southern cassowary (Casuarius casuarius) cranial specimen (eyes removed via dissection; TLG C069) in visible light (a, d, g) and 365 nm (b, c, e, f, h, i). Photos were taken with a visible light camera (a, b, d, e, g, h) and a UV-sensitive camera with a 365 nm band pass filter (c, f, i). The head was illuminated at a light source angle of − 25° (a–c), − 10° (d–f), and 0° (gg–i).
Discussion
Casque UV biofluorescence and reflectance
In this study, we present the first qualitative and quantitative descriptions of casque keratin biofluorescence in Casuarius. Biofluorescent patterns at 365 nm and within 385–395 nm are unique in each recognized cassowary species. The opsin gene expressed in palaeognathous birds7 indicates that Casuarius vision can detect UV light within our tested wavelengths (365 nm), and the casque surface reflects UV light at this same wavelength. We find that species, and possibly some subspecies, characteristically differ in biofluorescent patterns (Fig. 3). These patterns appear unique between individuals; however, males and females (in C. casuarius, specifically) did not show sex-specific UV biofluorescence. There were not enough known sex individuals in our C. bennetti and C. unappendiculatus sample to formerly test dimorphism in these two species. If this were to support the assertion that UV reflectance served as a visual signaling role, then we would expect 365 nm UV wavelengths to also be reflected from the casque surface in similar configurations as the biofluorescent patterns, which we tested by isolating this UV wavelength via a 365 nm bandpass filter. We found that keratin does reflect 365 nm light, but does so more broadly from the casque surface. Specifically, the majority of the keratinous surface can reflect this wavelength, depending on the angle of the light source relative to the casque (Figs. 4c, f and i and 5c, f and i). The species-specific biofluorescent patterns do not match those of the 365 nm reflectivity detected by the UV-sensitive camera with a bandpass filter in our experiments. This means that while cassowaries may be able to see some UV light reflected from the casque surface, it is unclear whether they are able to perceive the species-specific patterns that we documented using biofluorescence.
This unexpected result begs the question: why do consistent UV biofluorescent patterns persist if cassowaries are unable to see them? We offer three potential answers to this question. First, some aspect of the physical structure of keratin or how it grows over the surface of the casque may explain specific patterns. The basis of keratin biofluorescence and reflectivity in cassowary casques is unknown. We recommend future studies determine casque keratin biochemistry, molecular scaffolding, and fluorescent compounds (like porphyrins or carotenoids), one or all of which may contribute, mechanistically, to biofluorescence. Second, even though they are clearly not random, it is possible that the species-specific biofluorescent patterns we observe are coincidental and, thus, do not play a role in any biological functions. In this case, our results would represent a cautionary tale not to over-interpret apparent signaling patterns that do not appear to be a result of selection. Third, we cannot rule out that such casque patterns are detectable by cassowaries, but that this phenomenon is not detectable by our experimental design. Perhaps we did not test specific UV wavelengths which provide the most intense reflectivity or the combination between visible and UV light reflectivity could cause these areas to appear visually brilliant to cassowaries. In addition, even if cassowaries were only able to see more broadly, UV reflective casques, they may aid in assessment of overall ornament size and shape in the rainforest environments cassowaries inhabit, as dense green surroundings can allow UV reflective structures to stand out due to most leaves absorbing many UV wavelengths (see22 and references therein). There are other types of functions that may be beneficial with a more fully UV reflective casque (e.g., parent-offspring communication, interspecific identification, assessing health or age). Exactly how visible signals are perceived by cassowaries in their environments remains unknown, and thus, direct behavioral observations that test whether they respond to UV cues naturally may prove particularly important in parsing out the behavioral utility of such optical abilities.
Previous hypotheses for the biological role of cassowary casques include physical ramming (Crome & Moore, 1988), vocal resonation and audition44, thermoregulation45, 46, and visual display25, 26, 28, 33, 34, 44. Of these, visual signaling is the most widely supported, particularly in the context of stretch displays—behaviors in which cassowaries elongate their necks to present their casque at the apex of their body profile, likely to assert dominance or avoid conflict during potentially dangerous24, 42, 47 interactions with other cassowaries26, 48.
Given this behavioral context, our finding that casque keratin reflects UV light raises the possibility that UV reflectance enhances the visibility or contrast of the casque during such displays. Although the unique biofluorescent patterns (Figs. 1 and 2) we documented are questionable to be visible to cassowaries, the presence of non-patterned UV reflectance (Figs. 4c, f and i and 5c, f and i)—especially under lighting conditions where UV is proportionally more abundant (e.g., dawn, dusk, or under canopy-filtered light49, 50) could accentuate the casque’s visual salience. In this way, UV reflectance may function alongside visible-spectrum coloration in head and neck skin24 to amplify visual signals used in social or reproductive contexts. This possibility aligns with previous hypotheses about casque evolution and warrants further investigation through behavioral and perceptual studies in cassowaries.
Importance of biofluorescence in cassowary conservation and museum studies
The biofluorescence simulated via UV-handheld light is vivid in living and preserved C. casuarius and C. unappendiculatus specimens. There was a lack of keratinous rhamphotheca biofluorescence in non-casqued palaeognaths (i.e., S. camelus, D. novaehollandiae; Supplementary Fig. 1), suggesting that this type of keratin UV biofluorescence is a unique characteristic within Casuarius. Our results suggest that (1) species differ based on mean coverage of UV casque biofluorescence and (2) patterns of biofluorescence are unique between individuals within the same species. Together, these two types of information can aid researchers in species recognition as well as individual identification.
Our results indicate that UV biofluorescence of casques for specimens preserved in museums (n = 86) accurately reflects the biofluorescence and degree of coverage also observed in the casques of live individuals (n = 9). This includes dried samples (study skins, mounted specimens), frozen specimens, and those fixed and maintained in liquid media (see Methods). A recent study described a geometric morphometric method to differentiate Casuarius species by casque outline shape, indicating species-specific casque shapes between taxa21. Because casque biofluorescence is retained regardless of preservation mode, precise taxonomic identification can be additionally be determined from cassowary individuals in historic collections with no postcrania (e.g., inability to see the number of wattles), poorly preserved apteria (e.g., taxonomically identifiable skin color that fades more and more over time), non-specific geographic data (e.g., regional range of specific species is unknown), or slightly damaged casque shapes (e.g., dorsalmost region of the casque that is the most diagnostic between species and most likely to be damaged in museum drawers) through this methodology. If museums possess cassowary specimens of unknown species, curators can use UV lights to determine percent biofluorescent casque coverage and biofluorescent casque regions and compare them to the results in this study to assess taxonomic identity (Fig. 3), at least on a preliminary basis. Few studies have investigated the genetic relationships within Casuarius25, 51, and we recommend that future studies should explore the phylogenetic reliability of such casque-based relationships. Until the taxonomic relationships in Casuarius can be reliably determined using molecular techniques, biofluorescent casque variation can be included with other morphological characteristics in phylogenetic studies. For example, the two patterns of UV casque biofluorescence we identified in C. unappendiculatus may be useful in our understanding of previously proposed subspecies relationships24, 29.
In the field, the dense and dark forests that cassowaries inhabit obscure light, which may impede researchers from properly photographing and identifying individual birds, particularly casque shape. In low-light conditions, casque biofluorescence is vividly evident in live birds at 365 and 385‒395 nm wavelengths and using mobile UV equipment; therefore, it could be a direct and safe field methodology for ecologists to differentiate cassowary individuals (Supplementary Fig. 2) or species (Fig. 2) at a distance. Perhaps, trail cameras could be modified with UV-sensitive detectors and flashes to capture casque anatomy via remote monitoring. Cassowaries are a challenging group of birds to study due to their rarity, elusiveness, and territorial aggression24, 42, 47. Unique, contrasted biofluorescent patterns would allow for more precise censusing of wild cassowaries similar to the tail fluke “fingerprinting” of cetaceans (e.g.52, 53). The contrasting UV keratin signatures (Figs. 1 and 2) would greatly assist automated, digital software54, 55 in this identification process as well.
Conclusions and future investigations
The current study is the first to describe casque biofluorescence and reflectivity within Casuarius while simultaneously raising questions regarding cassowary vision, casque function, and ornament evolution in archosaurs. Reviewing our aims listed in the Introduction section, we find that the casques of cassowaries (1) vary interspecifically from non-UV fluorescent to highly UV fluorescent (2) at wavelengths potentially visible to Casuarius (3) with some UV reflectivity, but without indication of species-specific UV biofluorescence patterns. Our experimental design, therefore, (4) does not directly support the role for these ornaments in conspecific and congeneric visual signaling. Biofluorescent properties may aid in the taxonomic and individual identification of these rare avians in museum and field studies. It is critical that subsequent studies focus on the intensity of UV reflectivity necessary for cassowaries to observe UV casque coloration in various natural lighting conditions (e.g., in direct sunlight, in dense forest canopy, in low-light dawn and dusk) to better understand whether a biological role in casque signaling is possible. In addition, we suggest the future examination of the biomolecular components and structural composition of casque keratin that is responsible for biofluorescence and reflectance. Such seemingly unique features within palaeognathous birds may shed light upon cassowary evolution in deep time.
Methods
Specimen sample
In total, 95 adult Casuarius individuals were analyzed for this study (Supplementary Table 1). Specifically, casque keratin from dried (C. bennetti, n = 32; C. casuarius, n = 43; C. unappendiculatus, n = 8), frozen (C. casuarius, n = 3), and living specimens (C. casuarius, n = 6; C. unappendiculatus, n = 3) was exposed to UV light (wavelength ranges of 365 nm and 385‒395 nm). In addition, live, adult ostriches (Struthio camelus, n = 22) and emus (Dromaius novaehollandiae, n = 6) were observed under the same UV wavelengths as outgroups to determine whether bill keratin (i.e., rhamphotheca) in closely related palaeognathous birds exhibits biofluorescence. Individuals were determined to be adults via museum databases or assessed based on plumage and apteria development (see28, 31). Sexes were determined via museum databases, dissections of postcrania, or observed behavior. Specimen data were collected from both captive and wild-caught, vouchered specimens housed at the American Museum of Natural History (AMNH; New York, NY, USA), Brevard Zoo (BVZ; Melbourne, FL, USA), Natural History Museum at Tring (NHMUK; Tring, GBR), Sedgwick County Zoo (SCZ; Wichita, KS, USA), and T. L. Green Research Collection (TLG; Denver, CO, USA) as well as live animals from the Cassowary Conservation Project (CCP; Fort Pierce, FL, USA), Laughing D Farm (LDF; Parker, CO, USA), and Longneck Ranch (LNR; Rose Hill, KS). We did not include data from fluid-fixed specimens, as many preservation liquids themselves fluoresce under UV wavelengths (T. L. G. pers. obs.); however, we note that fluid-preserved cassowary specimens did appear to possess similar casque biofluorescence patterns as other preservation types. Depending on casque condition, access to specific anatomical orientations, and unknown sex status, some specimens were not used in all analyses (see sex data below). All UV light experiments with living cassowaries were conducted humanely, in accordance with all relevant state and local regulations, and approved by the Institutional Animal Care and Use Committee at New York Institute of Technology College of Osteopathic Medicine (NYITCOM; Protocol #2021-TG-01A1). In addition, the authors complied with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines.
UV biofluorescence photographic data collection
Photographs for UV analyses of preserved specimens were collected by T. L. G. using a Canon EOS 90D, Canon EF 100–400 mm f/4.5–5.6L IS II USM Lens (Canon Inc., Tokyo, JPN). Photographs were taken at distances ≥ 1.0 m from the sample with solid-contrasting backgrounds to ensure casque visibility. To ensure the most optimal settings for deciphering UV patterns, we made substantial efforts to control the lighting environment for each specimen. When possible, vouchered museum specimens were photographed inside a portable light/dark box (27” in. Angler Port-a-Cube LED Light Tent with Dimmer II; Angler, Manhattan, NY, USA). All individuals were photographed under visible (400–700 nm) and two wavelengths of UV light (365 nm and 385‒395 nm). We sampled these wavelengths because the cones in cassowary eyes are sensitive to this range (Aidala et al., 2012), and these wavelengths are typically used to detect biological fluorescence (Aidala et al., 2012; Dunning et al., 2018). Pairs of portable ultraviolet lights (V3 black light UV flashlights; uvBeast, Portland, Oregon, USA) were used for each of the UV wavelengths to provide full coverage to the casque surface and to minimize shadows. Cassowary heads were photographed from lateral and rostral views, in accordance with the methods from Green et al. (2022). Living specimens were photographed by T.L.G. with a Panasonic DMC-ZS60, Leica DC Vario-Elmar 1:3.3–6.4/4.3–129 ASPH Lens (Panasonic Corporation, Kadoma, JP; Leica Microsystems, Wetzlar, GER) behind chain-link fencing at CCP, which served to ensure safety for the observer. Live animals were exposed to UV wavelengths for 15-second sessions. If needed, up to four exposures were conducted per session (one session = within a one-hour period), with a maximum of one session per day (maximum allowance of 60 seconds of exposure for each animal). If individual animals appeared to be uncomfortable with the procedure in any way (e.g., squinting of eyes, attempting to run away, hissing vocalizations, panting with mouth open) or if we were concerned an animal may injure itself, we planned to cease the light experiment and exclude them from the study; however, none of the live birds exhibited such complications. All live birds in this study were adults (i.e., sexually mature; Casuarius ≥ 4.0 years; Dromaius ≥ 3.0 years; Struthio ≥ 2.0 years). Sex was recorded for each of these individuals (see Supplementary Table 1), but weight was not collected as it was not feasible and would have been particularly stressful and dangerous for the birds and researchers. Ultraviolet protective eyeglasses (ANSI-Z87 rated with side protection) were used by all data collectors and individuals in the area as a preventative measure against UV light exposure.
Statistical analyses
Only preserved specimens (n = 86) were analyzed for our primary statistical analyses for determination of casque biofluorescence coverage (C. bennetti, n = 32; C. casuarius, n = 46; C. unappendiculatus, n = 8) because live individuals were photographed with a separate camera. Lateral- and rostral-view casque photographs were used to determine the percentage of relative casque area that is biofluorescent. The casques were scored in binary states of non-biofluorescence and biofluorescence by importing image files into ImageJ (v. 1.53c; US National Institutes of Health, Bethesda, MD, USA), splitting color channels (blue for 365 nm; green for 385–395 nm) due to the visual appearance of the keratin biofluorescence at 365 nm (light blue; Fig. 1d) versus 385–395 nm (light green; Fig. 1b, f), tracing the casque with the polygon and fit spline tools, adjusting the threshold to select areas of fluorescence, and measuring total relative casque area and total relative biofluorescence area. The percentage of casque biofluorescence in both lateral and rostral views was used for statistical comparisons between conspecifics and species.
The majority of the cassowaries in our preserved sample (n = 70) were photographed from the left lateral view; however, 16 could only be photographed from the right. Therefore, we analyzed 26 C. casuarius that were photographed from both the left and right to ensure consistency of right-specific versus left-specific patterns and provide a rationale for pooling lateral samples. We completed all statistical analyses in this study in RStudio56. Normality was assessed using Shapiro-Wilk tests, box plots, and outlier testing (see Supplementary R Code). Due to the non-normal nature of our data, we conducted non-parametric Wilcoxon signed-rank tests (paired, two-tailed). We found no significant differences (α = 0.05) in casque biofluorescence coverage between left and right views at either 365 nm wavelengths (p = 0.1153) or 385–395 nm (p = 0.1096). In addition, correlation analyses were conducted and showed strong correspondence between left and right sides for both wavelengths (correlation coefficient > 0.982; p < 0.0001). Thus, the two sides of the casque showed statistically comparable variation, justifying our inclusion of the 16 right-lateral-only specimens with the specimens photographed from the left lateral view.
The casque shapes of C. casuarius have previously been described as non-dimorphic between females and males (Green et al., 2022). To confirm that biofluorescent coverage is not sexually dimorphic, we compared intraspecific data between female (n = 11) and male (n = 14) C. casuarius because this was the species with the most known-sex individuals in our sample. Shapiro-Wilk tests, box plots, and outliers testing indicated that these data did not follow a normal distribution; therefore, non-parametric Kruskal-Wallis one-way analyses of variance were conducted on sex data (see Supplementary R Code). We found no significant differences (α = 0.05) in casque biofluorescence coverage between sexes in lateral (365 nm, p = 0.5820; 385–395 nm, p = 0.5430) or rostral views (365 nm, p = 0.8674; 385–395 nm, p = 0.9202) at either UV wavelength, justifying our inclusion of females and males in the same analyses.
We tested for interspecific differences (C. bennetti, n = 32; C. casuarius, n = 46; C. unappendiculatus, n = 8) in biofluorescence coverage using nonparametric Kruskal-Wallis one-way analyses of variance because data were determined to be nonnormal via Shapiro-Wilk tests, box plots, and outliers testing. Species means for biofluorescent casque coverage in lateral and rostral views were used as the basis of comparison for this dataset. In addition, we compared the species means from C. casuarius and C. unappendiculatus in this preserved specimen dataset with those from living individuals of these taxa (C. casuarius, n = 6; C. unappendiculatus, n = 3) to determine if the biofluorescent casque percentage of live individuals fell within the range of our larger, preserved specimen sample.
UV reflectivity photographic data collection
We performed light reflectivity experiments with three frozen C. casuarius individuals. All heads were frozen for less than six years in airtight polyethylene bags to prevent freeze-drying and represented the freshest tissue from non-living individuals in our study. These tests were not plausible to conduct on moving, living cassowaries. These frozen specimens (TLG C022, TLG C062, TLG C069) were photographed using a Canon EOS Rebel T2i (Canon Inc., Tokyo, JP), Yongnuo 50 mm f/1.8 Lens (Yongnuo, Shenzhen, CN), and a UV-modified (UV/IR cut sensor replaced with 100% UV passing glass) Canon EOS 2000D (Canon Inc., Tokyo, JP; Kolari Vision LLC, Raritan, New Jersey, USA), Yongnuo 50 mm f/1.8 Lens (Yongnuo, Shenzhen, CN) with a Kolari 52 mm UV Ready 365 nm bandpass filter (Kolari Vision LLC, Raritan, New Jersey, USA). Three treatments were implemented to photograph these specimens: (1) in visible light with the unmodified camera, (2) in light from a 365 nm handheld light (V3 black light UV flashlights; uvBeast, Portland, Oregon, USA) with the unmodified camera, and (3) in light from a 365 nm handheld light with the UV-modified camera and the 365 nm bandpass filter. Because the 365 nm light source emits visible blue light as well, the bandpass filter prevented these wavelengths from reaching the camera’s detector. In addition, the specimens were photographed at three light incident angles (0°, − 10°, − 25°) and three head rotations (0°, − 45°, − 90°).
Data availability
Morphological data that support the findings of this study have been deposited in Morphobank at http://morphobank.org/permalink/?P4639.
References
- Mayr, G. A survey of casques, frontal humps, and other extravagant bony cranial protuberances in birds. Zoomorphology 137, 457–472 (2018).
Article Google Scholar - Terrill, R. S. & Shultz, A. J. Feather function and the evolution of birds. Biol Rev (2022).
- Sereno, P.C. et al., New scimitar-crested Spinosaurus species from the Sahara caps stepwise spinosaurid radiation. Science, 391, 782–792 (2026).
- McCoy, D. E. & Prum, R. O. Convergent evolution of super black plumage near bright color in 15 bird families. J. Exp. Biol. 222, jeb208140 (2019).
Article PubMed Google Scholar - Bolwig, N. Agonistic and sexual behavior of the African ostrich (Struthio camelus). Condor 75, 100–105 (1973).
Article Google Scholar - Hill, G. E. & McGraw, K. J. Bird Coloration: Function and Evolution. Vol. 2 (Cambridge, Harvard University, 2006).
- Ödeen, A. & Håstad, O. The phylogenetic distribution of ultraviolet sensitivity in birds. BMC Evol. Biol. 13, 1–10 (2013).
Article Google Scholar - Höglund, J. et al. Owls lack UV-sensitive cone Opsin and red oil droplets, but see UV light at night: retinal transcriptomes and ocular media transmittance. Vis. Res. 158, 109–119 (2019).
Article PubMed Google Scholar - Rajchard, J. Ultraviolet (UV) light perception by birds: a review. Vet. Med. (Praha). 54, 351–359 (2009).
Article Google Scholar - Bennett, A. T. & Cuthill, I. C. Ultraviolet vision in birds: what is its function? Vis. Res. 34, 1471–1478 (1994).
Article CAS PubMed Google Scholar - Viitala, J., Korpimaki, E., Palokangas, P. & Koivula, M. Attraction of kestrels to vole scent marks visible in ultraviolet light. Nature 373, 425–427 (1995).
Article ADS CAS Google Scholar - Altshuler, D. L. Ultraviolet reflectance in fruits, ambient light composition and fruit removal in a tropical forest. Evol. Ecol. Res. 3, 767–778 (2001).
Google Scholar - Dunning, J. Photoluminescence in the bill of the Atlantic Puffin Fratercula Arctica. Bird. Study. 65, 570–573 (2018).
Article Google Scholar - Wilkinson, B. P., Johns, M. E. & Warzybok, P. Fluorescent ornamentation in the rhinoceros auklet Cerorhinca monocerata. Ibis 161, 694–698 (2019).
Article Google Scholar - Pearn, S. M., Bennett, A. T. & Cuthill, I. C. Ultraviolet vision, fluorescence and mate choice in a parrot, the Budgerigar Melopsittacus undulatus. Proc. Royal Soc. B. 268, 2273–2279 (2001).
Article CAS Google Scholar - Parejo, D., Avilés, J. M. & Rodríguez, J. Visual cues and parental favouritism in a nocturnal bird. Biol. Lett. 6, 171–173 (2010).
Article PubMed Google Scholar - He, Y. et al. Deep learning image segmentation reveals patterns of UV reflectance evolution in passerine birds. Nat. Commun. 13, 1–13 (2022).
ADS Google Scholar - Håstad, O., Victorsson, J. & Ödeen, A. Differences in color vision make passerines less conspicuous in the eyes of their predators. PNAS 102, 6391–6394 (2005).
Article ADS PubMed PubMed Central Google Scholar - Aidala, Z. et al. Ultraviolet visual sensitivity in three avian lineages: Paleognaths, parrots, and passerines. J. Comp. Physiol. 198, 495–510 (2012).
Article Google Scholar - Woodruff, D. C., Naish, D. & Dunning, J. Photoluminescent visual displays: an additional function of integumentary structures in extinct archosaurs? Hist. Biol. 33, 1718–1725 (2021).
Article Google Scholar - Weidensaul, C. S. et al. Use of ultraviolet light as an aid in age classification of owls. Wilson J. Ornithol. 123, 373–377 (2011).
Article Google Scholar - Nicolaï, M. P. The function and consequences of fluorescence in tetrapods. PNAS 121, e2318189121 (2024).
Article PubMed PubMed Central Google Scholar - Marshall, J. & Johnsen, S. Fluorescence as a means of colour signal enhancement. Philos. Trans. R Soc. B. 372, 20160335 (2017).
Article Google Scholar - Rothschild, A. A monograph of the genus Casuarius. Trans. Zool. Soc. Lond. 15, 109–148 (1900).
Article Google Scholar - Hone, D. W. E. & Naish, D. The ‘species recognition hypothesis’ does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs. J. Zool. 290, 172–180 (2013).
Article Google Scholar - Green, T. L., Kay, D. I., Watanabe, A. & Gignac, P. M. Species-specific casque shapes in the genus Casuarius and implications for visual display. Ornithol 142, ukaf026 (2025).
Article Google Scholar - Crome, F. H. J. The cassowary’s casque. Emu 88, 123–124 (1988).
Article Google Scholar - Green, T. L., Kay, D. I. & Gignac, P. M. Intraspecific variation and directional casque asymmetry in adult Southern cassowaries (Casuarius casuarius). J. Anat. 241, 951–965 (2022).
Article PubMed PubMed Central Google Scholar - Perron, R. M. Taxonomy of the Genus Casuarius: the Defined and Known Living Cassowary Species and Subspecies (United Kingdom: Quantum Conservation,, 2016).
- Green, T. L. & Gignac, P. M. Osteological comparison of casque ontogeny in palaeognathous and neognathous birds: implications for selecting modern analogs in the study of cranial ornaments from extinct archosaurs. Zool. J. Linn. Soc. https://doi.org/10.1093/zoolinnean/zlad016] (2023).
Article Google Scholar - Green, T. L. & Gignac, P. M. Osteological description of casque ontogeny in the Southern cassowary (Casuarius casuarius) using micro-CT imaging. Anat. Rec. 304, 461–479 (2021).
Article CAS Google Scholar - Naish, D. & Perron, R. Structure and function of the cassowary’s casque and its implications for cassowary history, biology and evolution. Hist. Biol. 28, 507–518 (2016).
Article Google Scholar - Dodson, P. Taxonomic implications of relative growth in lambeosaurine hadrosaurs. Syst. Biol. 24, 37–54 (1975).
Article Google Scholar - Hone, D. W., Naish, D. & Cuthill, D. I. C. Does mutual sexual selection explain the evolution of head crests in pterosaurs and dinosaurs? Lethaia 45, 139–156 (2012).
Article Google Scholar - Farke, A. A., Chok, D. J., Herrero, A., Scolieri, B. & Werning, S. Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids. PeerJ 1, e182 (2013).
Article PubMed PubMed Central Google Scholar - Lü, J. et al. High diversity of the Ganzhou oviraptorid fauna increased by a new cassowary-like crested species. Sci. Rep. 7, 1–13 (2017).
ADS Google Scholar - Stocker, G. C. & Irvine, A. K. Seed dispersal by cassowaries (Casuarius casuarius) in North Queensland’s rainforests. Biotropica, 170–176 (1983).
- Mack, A. L. Distance and non-randomness of seed dispersal by the Dwarf cassowary Casuarius bennetti. Ecography 18, 286–295 (1995).
Article ADS Google Scholar - Webber, B. L. & Woodrow, I. E. Cassowary frugivory, seed defleshing and fruit fly infestation influence the transition from seed to seedling in the rare Australian rainforest tree, Ryparosa sp. nov. 1 (Achariaceae). Funct. Plant. Biol. 31, 505–516 (2004).
Article PubMed Google Scholar - Bradford, M. G. & Westcott, D. A. Consequences of Southern cassowary (Casuarius casuarius, L.) gut passage and deposition pattern on the germination of rainforest seeds. Austral Ecol. 35, 325–333 (2010).
Article Google Scholar - Bradford, M. G. & Westcott, D. A. Predation of cassowary dispersed seeds: is the cassowary an effective disperser? Integr. Zool. 6, 168–177 (2011).
Article PubMed Google Scholar - Crome, F. H. J. Some observations on the biology of the cassowary in Northern Queensland. Emu 76, 8–14 (1976).
Article Google Scholar - Mullen, P. & Pohland, G. Studies on UV reflection in feathers of some 1000 bird species: are UV peaks in feathers correlated with violet-sensitive and ultraviolet‐sensitive cones? Ibis 150, 59–68 (2008).
Article Google Scholar - Mack, A. L. & Jones, J. Low-frequency vocalizations by cassowaries (Casuarius spp). Auk 120, 1062–1068 (2003).
Article Google Scholar - Phillips, P. K. & Sanborn, A. F. An infrared, thermographic study of surface temperature in three ratites: ostrich, Emu and double-wattled cassowary. J. Therm. Biol. 19, 423–430 (1994).
Article Google Scholar - Eastick, D. L., Tattersall, G. J., Watson, S. J., Lesku, J. A. & Robert, K. A. Cassowary casques act as thermal windows. Sci. Rep. 9, 1966 (2019).
Article ADS PubMed PubMed Central Google Scholar - Kofron, C. P. Attacks to humans and domestic animals by the Southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia. J. Zool. 249, 375–381 (1999).
Article Google Scholar - Bentrupperbaumer, J. Reciprocal Ecosystem Impact and Behavioural Interactions Between Cassowaries, Casuarius casuarius, and Humans, Homo sapiens: Exploring the Natural-Human Environment Interface and its Implications for Endangered Species Recovery in North Queensland, AustraliaPhD thesis, James Cook University. (1997).
- Henriksen, K., Stamnes, K. & Østensen, K. P. Measurements of solar UV, visible and near IR irradiance at 78 N. Atmos. Environ. 23, 1573–1579 (1989).
Article ADS CAS Google Scholar - Hogg, C. et al. Arctic reindeer extend their visual range into the ultraviolet. J. Exp. Biol. 214, 2014–2019 (2011).
Article PubMed Google Scholar - Mitchell, K. J. et al. Ancient DNA reveals elephant birds and Kiwi are sister taxa and clarifies ratite bird evolution. Science 344, 898–900 (2014).
Article ADS CAS PubMed Google Scholar - Katona, S. et al. Identification of humpback whales by fluke photographs. Behav. Mar. Animals Curr. Perspect. Res. 3, 33–44 (1979).
Article Google Scholar - Yoshikawa, T. et al. Identification of over one thousand individual wild humpback whales using fluke photos. VISIGRAPP 4, 957–967 (2022).
Google Scholar - Ranguelova, E., Huiskes, M. & Pauwels, E. J. Towards computer-assisted photo-identification of humpback whales. 2004 Int. Conf. Image Process. IEEE 3, 1727–1730 (2004).
Google Scholar - Bogucki, R. et al. Applying deep learning to right Whale photo identification. Conserv. Biol. 33, 676–684 (2019).
Article PubMed Google Scholar - Posit Team. RStudio: Integrated development environment for R. Posit Software, PBC, Boston, MA (2025).
Acknowledgements
Funding for this study was provided by: The Company of Biologists and sponsoring journal, Experimental Biology (Visiting Fellowship, JEBTF1903122; to T.L.G.), and the Western Interior Paleontological Society (2023 Karl Hirsch Memorial Grant; to T.L.G.). For specimen access and/or donation, the authors thank: American Museum of Natural History (Paul Sweet, Joel Cracraft, Thomas Trombone, Augie Kramer), Brevard Zoo (Trevor T. Zachariah, M. David Quavillon, Ellen Dreyer, Michelle Smurl), Cassowary Conservation Project (R. Glenn Hood, Scott Snedeker), Laughing D Farm (Karin Davidson, John Davidson), Longneck Ranch (Stan Barenberg, Samantha Potts), Natural History Museum at Tring (Mark Adams, Hein Van Grouw, Paul Kitching, Douglas Russell, Judith White, Joanne Cooper, Pete Key, Claire Walsh), and Sedgwick County Zoo (Scott Newland, Heather Arens, and Phillip Horvey). For additional assistance and discussions, the authors extend gratitude to Brian Beatty, Jason Bourke, Jennifer Campbell-Smith, Tim Carlton, Danielle Carroll, Adam Cossette, Dosha Cummins, AJ Dodd, Natarajan Ganesan, Jonathan Geisler, Jane Green, Richard Green, Simone Hoffmann, David Hone, David Ian Kay, Andrew Mack, Nicholas Mahler, Danny Martin, Matthew Mihlbachler, Julia Molnar, Darren Naish, Helian Naish, Haley O’Brien, Karen Poole, Viswanathan Rajagopalan, Amber Schlater, Nikos Solounias, Nathan Thompson, and Melody Young.
Funding
Funding for this study was provided by: The Company of Biologists and sponsoring journal, Experimental Biology (Visiting Fellowship, JEBTF1903122; to T.L.G.), and the Western Interior Paleontological Society (2023 Karl Hirsch Memorial Grant; to T.L.G.). For specimen access and/or donation, the authors thank: American Museum of Natural History (Paul Sweet, Joel Cracraft, Thomas Trombone, Augie Kramer), Brevard Zoo (Trevor T. Zachariah, M. David Quavillon, Ellen Dreyer, Michelle Smurl), Cassowary Conservation Project (R. Glenn Hood, Scott Snedeker), Laughing D Farm (Karin Davidson, John Davidson), Longneck Ranch (Stan Barenberg, Samantha Potts), Natural History Museum at Tring (Mark Adams, Hein Van Grouw, Paul Kitching, Douglas Russell, Judith White, Joanne Cooper, Pete Key, Claire Walsh), and Sedgwick County Zoo (Scott Newland, Heather Arens, and Phillip Horvey). For additional assistance and discussions, the authors extend gratitude to Brian Beatty, Jason Bourke, Jennifer Campbell-Smith, Tim Carlton, Danielle Carroll, Adam Cossette, Dosha Cummins, AJ Dodd, Troy Ford, Natarajan Ganesan, Jonathan Geisler, Jane Green, Richard Green, Simone Hoffmann, David Hone, David Ian Kay, Andrew Mack, Nicholas Mahler, Danny Martin, Matthew Mihlbachler, Julia Molnar, Darren Naish, Helian Naish, Haley O’Brien, Karen Poole, Viswanathan Rajagopalan, Amber Schlater, Nikos Solounias, Nathan Thompson, and Melody Young.
Author information
Authors and Affiliations
- Department of Biological and Anatomical Sciences, New York Institute of Technology College of Osteopathic Medicine at Arkansas State University, Wilson Hall, 2405 Aggie Road, Jonesboro, NY, 72401, USA
Todd L. Green - Department of Anatomy, New York Institute of Technology College of Osteopathic Medicine, Old Westbury, NY, 11568, USA
Akinobu Watanabe - Division of Paleontology, American Museum of Natural History, New York, NY, 10024, USA
Akinobu Watanabe & Paul M. Gignac - Life Sciences Department, Natural History Museum, London, SW7 5BD, UK
Akinobu Watanabe - Department of Biomedical Sciences, Baptist Health Sciences University College of Osteopathic Medicine, Memphis, TN, 38104, USA
Jonathan M. Berman - Department of Ecology and Evolutionary Biology, The University of Tennessee, Knoxville, Knoxville, TN, 37996, USA
Michael C. Granatosky - Department of Cellular and Molecular Medicine, University of Arizona College of Medicine, Tucson, AZ, 85308, USA
Paul M. Gignac - MicroCT Imaging Consortium for Research and Outreach (MICRO), Fayetteville, AR, 72701, USA
Paul M. Gignac
Authors
- Todd L. Green
- Akinobu Watanabe
- Jonathan M. Berman
- Michael C. Granatosky
- Paul M. Gignac
Contributions
T.L.G., P.M.G., A.W., and J.M.B. conceived and designed the study. T.L.G., P.M.G., A.W., J.M.B., and M.C.G. wrote and edited the manuscript and provided analysis and interpretation of data. T.L.G., P.M.G., and J.M.B. conducted museum specimen and collections work. T.L.G., M.C.G., and A.W. conducted live specimen observation and photography. A.W. and T.L.G. wrote and managed R code and constructed tables and graphs. T.L.G. designed and created figures.
Corresponding author
Correspondence toTodd L. Green.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article
Cite this article
Green, T.L., Watanabe, A., Berman, J.M. et al. Ultraviolet light illuminates species-specific biofluorescent casque patterns in cassowaries (Casuarius).Sci Rep 16, 10302 (2026). https://doi.org/10.1038/s41598-026-40230-1
- Received: 19 August 2025
- Accepted: 11 February 2026
- Published: 24 February 2026
- Version of record: 27 March 2026
- DOI: https://doi.org/10.1038/s41598-026-40230-1