Phylogeny, character evolution, and classification of Selaginellaceae (lycophytes) (original) (raw)
Abstract
Selaginella is the largest and most taxonomically complex genus in lycophytes. The fact that over 750 species are currently treated in a single genus makes Selaginellales/Selaginellaceae unique in pteridophytes. Here we assembled a dataset of six existing and newly sampled plastid and nuclear loci with a total of 684 accessions (74% increase of the earlier largest sampling) representing ca. 300 species to infer a new phylogeny. The evolution of 10 morphological characters is studied in the new phylogenetic context. Our major results include: (1) the nuclear and plastid phylogenies are congruent with each other and combined analysis well resolved and strongly supported the relationships of all but two major clades; (2) the Sinensis group is resolved as sister to S. subg. Pulviniella with strong support in two of the three analyses; (3) most morphological characters are highly homoplasious but some characters alone or combinations of characters well define the major clades in the family; and (4) an infrafamilial classification of Selaginellaceae is proposed and the currently defined Selaginella s.l. is split into seven subfamilies (corresponding to the current six subgenera + the Sinensis group) and 19 genera (the major diagnosable clades) with nine new species-poor genera. We support the conservation of Selaginella with a new type, S. flabellata, to minimize nomenclatural instability. We provide a key to subfamilies and genera, images illustrating their morphology, their morphological and geographical synopses, a list of constituent species, and necessary new combinations. This new classification will hopefully facilitate communication, promote further studies, and help conservation.
Keywords: Generic classification, Homoplasy, Lycophyte phylogeny, Megaspore types, Microspores, Nuclear 18S and 26S genes
Graphical abstract
Highlights
- •
684 accessions representing ca. 300 (40% of total) species of Selaginella are sampled. - •
Nuclear and plastid phylogenies are congruent with each other. - •
The evolution of 10 morphological characters is studied. - •
The Sinensis group is resolved as sister to S. subg. Pulviniella. - •
Selaginella s.l. is split into 7 subfamilies and 19 genera.
1. Introduction
Selaginella (Selaginellaceae) is the largest genus in lycophytes and known for its special phylogenetic position in vascular plants, its heterospores, its resurrection ability, and its pharmacological value (Banks et al., 2009). Selaginella is among the earliest diverging lineages of vascular plants and diverged from its closest living relative about 383 million years ago (Ma) in the Devonian Period (Klaus et al., 2017).
Following PPG I (2016), Selaginella alone constitutes the family Selaginellaceae and the order Selaginellales. However, it is unique for an order/family in pteridophytes with 700 or more species (Jermy, 1990; Tryon and Lugardon, 1991; Zhou and Zhang, 2015) to contain only one genus, considering the most recent classifications of pteridophytes, for example, ca. 380 species in 17 genera in Lycopodiales (Chen et al., 2022; Zhang and Zhou, 2022), the sister order of Selaginellales + Isoëtales, ca. 265 species in 25 genera in Blechnaceae (PPG I, 2016; de Gasper et al., 2016; González et al., 2020), ca. 1200 species in 37 genera in Thelypteridaceae (Fawcett and Smith, 2021), and ca. 900 species in 42 genera in Polypodiaceae subfam. Grammitidoideae (Yang et al., 2023; Zhou et al., 2023).
In fact, morphological heterogeneity within the broadly defined Selaginella or Selaginellaceae has been observed for a long time and a number of studies to recognize several genera in Selaginellaceae have been published. Splitting Selaginella into several genera dates back to Lamark and Mirbel (1803) and Palisot de Beauvois (1804) when they described and recognized Didiclis P. Beauv. ex Mirb., Diplostachyum P. Beauv., Gymnogynum P. Beauv., and Stachygynandrum P. Beauv. ex Mirb. Kuntze (1891) transferred a number of species of Selaginella to Lycopodioides but he actually did not split the genus into two, because he recognized Lycopodioides only. Börner (1912) separated Heterophyllium Hieron. ex Börner from Selaginella based on Selaginella helvetica, although this generic name is a later homonym of Heterophyllium (Schimp.) Müll. Hal. ex Kindb. and a nomenclatural synonym of Diplostachyum. Rothmaler (1944) recognized three genera in Selaginellaceae: Didiclis with a single macrosporangium at the base of strobili surrounded by sterile sporophylls, Lycopodioides with multiple macrosporangia at the base of strobili without any sporophylls, and Selaginella with uniform and spirally arranged leaves. Kung (1988) divided Selaginella s.l. into Lycopodioides and Selaginella s.s. and restricted the latter to Selaginella selaginoides (L.) P. Beauv. ex Schrank & Mart. and its allies that have no rhizophores and have loosely spirally arranged sterile leaves and cylindric strobili (S. subg. Selaginella sensu Zhou and Zhang, 2015). Selaginella sensu Kung (1988) has the same circumscription as Rothmaler's (1944). Satou (1997) and Tzvelev (2004) adopted Rothmaler's (1944) and Kung's (1988) classifications. Soják (1992) accepted Selaginella sensu Rothmaler (1944) and Kung (1988) and split Lycopodioides sensu Kung (1988) into the narrowly defined Lycopodioides and Bryodesma Soják; the latter was based on Selaginella rupestris (L.) Spring and included those species with spirally arranged sterile leaves and decussate sporophylls. Most recently, Weakley (2012) has recognized three genera in the family, Bryodesma, Lycopodioides, and Selaginella s.s., for the Flora of the Southern and Mid-Atlantic States. Weakley (2022) and Weakley and Southeastern Flora Team (2022) included one more genus in the family, Gymnogynum, in the Flora of the Southeastern United States.
One of the major reasons that the broadly defined Selaginella was often adopted (e.g., Jermy, 1990; Valdespino, 1993; Zhang et al., 2013; PPG I, 2016) is that the current type of the genus, S. selaginoides, is a member of a two-species subunit with no rhizophores at all, dramatically different from all other species in the genus. Recognizing multiple genera would result in name changes of all but the two species without rhizophores, although the name Selaginella may be proposed for conservation with a conserved type (Turland et al., 2018).
The second major reason that the broadly defined Selaginella was often adopted is that the phylogenetic relationships within the genus had been largely unclear in spite of earlier phylogenetic studies (Korall et al., 1999; Korall and Kenrick, 2002, 2004; Arrigo et al., 2013) and the morphological homoplasy had not been adequately assessed in an appropriate phylogenetic context. These have been improved greatly in the past few years. Weststrand and Korall (2016a) and Zhou et al. (2016 available online in 2015) published large-scale phylogenies based on plastid and nuclear markers. Based on plastid rbcL and nuclear ITS data of 394 accessions representing ca. 200 species (ca. 25% of the extant diversity of the genus) of Selaginella, Zhou et al. (2016) identified six deep-level clades and 20 major clades with strong resolution of the relationships among most of these clades. Zhou and Zhang (2015) thus recognized these six deep-level clades as six subgenera of Selaginella (S. subg. Selaginella, S. subg. Boreoselaginella Warb., S. subg. Pulviniella Li Bing Zhang & X.M. Zhou, “S. subg. Ericetorum Jermy” (= S. subg. Gymnogynum (P. Beauv.) Weststrand & Korall), S. subg. Heterostachys Baker, and S. subg. Stachygynandrum (P. Beauv. ex Mirb.) Baker) and three of subgenera (S. subg. Ericetorum, S. subg. Heterostachys, and S. subg. Stachygynandrum) were further classified into six, five, and seven sections, respectively. However, some relationships, e.g., those of the Selaginella sanguinolenta clade, were not strongly supported in Zhou et al. (2016) and the enigmatic Sinensis group, which had caused poor phylogenetic resolution in the previous study (Korall and Kenrick, 2002), was not sampled in their study. Based on plastid rbcL and nuclear pgiC and SQD1 data of 340 accessions representing 223 species of Selaginella including 10 species of the Sinensis group, Weststrand and Korall (2016a) resolved the Sinensis group as sister to those taxa (S. subg. Pulviniella, S. subg. Ericetorum, S. subg. Heterostachys, and S. subg. Stachygynandrum sensu Zhou and Zhang (2015)) with rhizophores clearly originated on the ventral side of the stem with moderate support (PP = 0.97) in Bayesian inference (BI). The relationships of five out of the seven major clades identified by Weststrand and Korall (2016a) were not resolved. Depending on different DNA markers, the relationships of S. subg. Boreoselaginella [including Selaginella nummularifolia Ching, Selaginella rossii (Baker) Warb., S. sanguinolenta (L.) Spring, etc.)] were resolved differently (Weststrand and Korall, 2016a; Zhou et al., 2016). Based on plastid genomes of 26 species of Selaginella, Zhang et al. (2019) resolved S. subg. Boreoselaginella as sister to the superclade with ventral rhizophores with strong support, same as the position β found by Weststrand and Korall (2016a). Zhang et al. (2020, 2022) then speculated that high GC content, extensive RNA editing sites, and elevated substitution rate of plastid genomes maybe strongly shook the stability of phylogenetic topology of Selaginella. A recent phylogenomic analysis based on 59 plastomes failed to well resolve the phylogenetic relationships in Selaginella with confidence, highlighting the difficulty in resolving the phylogeny and evolution of this particularly important land plant lineage (Zhou et al., 2022).
Morphologically, state of rhizophores, arrangement of vegetative leaves and sporophylls, and spore morphology are traditionally used for the classification of Selaginella (e.g., Jermy, 1986; Zhou and Zhang, 2015; Weststrand and Korall, 2016b). However, because of the homoplastic nature of some morphological characteristics, some taxa are very difficult to define and identify clearly (Zhou et al., 2015; Weststrand and Korall, 2016b). For example, rosette-forming habit is present in three subgenera: S. subg. Stachygynandrum [e.g., S. pallescens (C. Presl) Spring), S. subg. Gymnogynum [e.g., Selaginella lepidophylla (Hook. & Grev.) Spring, S. novoleonensis Hieron.), and S. subg. Pulviniella [e.g., S. pulvinata (Hook. & Grev.) Maxim., S. tamariscina (P. Beauv.) Spring), and dimorphic sporophylls appear in S. subg. Stachygynandrum [e.g., S. flagellata Spring, S. moritziana Spring, S. radiata Baker, S. simplex Baker] and S. subg. Heterostachys [e.g., S. leptophylla Baker, S. nipponica Franch. & Sav., S. repanda (Desv. ex Poir.) Spring]. Similarly, some morphological character states, e.g., rhizophores borne on dorsal sides of the stem, reticulate surfaces of megaspores, stele with more than two vascular bundles, and diverse sporangial arrangement on the strobili, are also present in more than one section and/or subgenus.
Selaginella is heterosporous. A number of studies on the morphology of megaspores and microspores (especially megaspores) of Selaginella have been published (e.g., Zhou et al., 2015; Valdespino et al., 2015; Valdespino et al., 2018a; Valdespino et al., 2018b; Valdespino, 2015, 2017a, 2020; Bauer et al., 2016; Wang et al., 2018). These studies showed that Selaginella holds extremely high diversity in spore morphology. Spore data have provided very important and useful taxonomic information which has been incorporated in the recent classifications of the genus (Zhou and Zhang, 2015; Weststrand and Korall, 2016a, 2016b). In addition to spore ornamentation, the size of both megaspores and microspores also shows great variation in Selaginella. Megaspore sizes range from 150 to 1500 μm and microspore sizes from 15 to 50 μm (Mickel and Hellwig, 1969; Zhou et al., 2015; Bauer et al., 2016). Taxonomically, species of S. subg. Gymnogynum often have larger (500–1000 μm) megaspores, whereas those of S. subg. Heterostachys and S. subg. Pulviniella usually have smaller (200–400 μm) megaspores.
Plastome studies (Tsuji et al., 2007; Smith, 2009; Wicke et al., 2011; Jansen and Ruhlman 2012; Ruhlman and Jansen 2018; Mower et al., 2019; Xu et al., 2018; Zhang et al., 2019a, Zhang et al., 2019b; Kang et al., 2020; Xiang et al., 2022; Zhou et al., 2022) showed that plastomes of Selaginellaceae and their infrafamilial lineages known so far have a number of unique and diverse features: (a) plastomes of most plant lineages are highly conserved with quadripartite structure composed by a large single copy (LSC), a small single copy (SSC), and two inverted repeats (IRa and IRb), whereas plastomes of Selaginellaceae exhibit DR structure (also can be R, DR, IR, or DR-IR coexisting types) with small to medium repeats existed in SC, and plastome conformations ranged from one to 24 (Zhou et al., 2022); (b) plastome sizes in most land plants are 120–160 kb but those in Selaginellaceae are 78–190 kb; (c) a plastome in other vascular plants usually contains approximately 120–130 genes but those of Selaginellaceae contain 36–128 genes; (d) accD/cemA/infA/psaM/rpl20/rpl21/rpl32/rpl33/rps12/rps15/rps16/ycf66/ycf94 and most of the rRNA, tRNA, and introns are generally lost or pseudogenetized in Selaginellaceae; (e) GC content in land plant plastomes ranges from 34% to 40%, but plastomes of Selaginellaceae often are extremely GC-rich (>50%); and (f) overall distinctions of plastomes among subgenera even among sections in Selaginella are much greater than those among orders/families/subfamilies/genera in other vascular plants.
In this study our goals included: (1) better resolving the relationships within Selaginella with expanded taxon sampling by including the elusive Sinensis group and with expanded character sampling by including two highly conserved nrDNA markers, 18S and 26S, to be sampled for the first time for Selaginella; (2) for the first time evaluating the evolution of gross morphology and micromorphology of megaspores and microspores and identifying morphological synapomorphies of various clades in the context of the new phylogeny; and (3) proposing an infrafamilial classification of Selaginellaceae based on new molecular and morphological results.
2. Materials and methods
2.1. Taxon sampling
Silica gel-dried materials and herbarium samples either were collected in the fields or herbaria (CDBI, KUN, MO, and PYU). Taxon sampling for this study consisted of 686 accessions representing ca. 300 species in Selaginella including all subgenera/sections recognized by Zhou and Zhang (2015) and Weststrand and Korall (2016b). For morphological analysis, 291 accessions to represent ca. 275 species of Selaginella were included.
2.2. DNA extraction, amplification and sequencing
Total genomic DNA was extracted from silica-dried material or sometimes from herbarium specimens using a TIANGEN plant genomic DNA extraction kit (TIANGEN Biotech., Beijing, China) following the manufacturers’ protocols. In total six genes, one plastid marker (rbcL), two single-copy nuclear genes (pgiC and SQD1), and three nuclear rDNA markers (18S, 5.8S, and 26S), were used in the phylogenetic study. The primer sequences and PCR conditions are listed in Table S1. PCR products were purified and sequenced by TSINGKE Biological Technology (Chengdu, China). Sequencher 4.14 (Gene Codes Corp., Ann Arbor, MI, USA) was used to assemble and edit contiguous sequences.
2.3. Sequence alignment and phylogenetic analysis
Sequences for each marker were initially aligned with MAFFT v.7 (Katoh and Standley, 2013) and manually adjusted in BioEdit (Hall, 1999). The ambiguously aligned areas were deleted. jModeltest 2.1.7 (Darriba et al., 2012) was used to select the best-fitting likelihood model for Bayesian analyses. The Akaike information criterion (Akaike, 1974) was used to select among models instead of the hierarchical likelihood ratio test, following Pol (2004) and Posada and Buckley (2004). The best-fitting models and parameter values are provided in Table 2. The alignment has been deposited in FigShare with study number #22730093.
Table 2.
Best-fitting models and parameter values for separate (rbc L, 18S, 26S, 5.8S, pgiC, and SQD1), simultaneous nuclear, and simultaneous nuclear + plastid datasets and in this study. “G” = gamma distribution shape parameter (Yang, 1994). “GTR” = general-time-reversible model (Tavaré, 1986). “I” = proportion of invariable sites. “pInv” = proportion of invariable sites. “Simultaneous-1” = 18S + 26S + 5.8S + pgiC + SQD1. “Simultaneous-2” = 18S + 26S + 5.8S + pgiC + SQD1 + rbcL.
| Region | Model (AIC) | Base frequencies | Substitution model (rate matrix) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | C | G | T | A–C | A–G | A–T | C–G | C–T | G–T | pInv | G | ||
| rbcL | GTR + I + G | 0.2857 | 0.2433 | 0.2255 | 0.2455 | 0.4631 | 3.9209 | 0.2720 | 0.5053 | 2.4318 | 1.0000 | 0.3140 | 1.0406 |
| 18S | GTR + I + G4 | 0.2781 | 0.2738 | 0.2168 | 0.2313 | 0.6532 | 6.6774 | 1.4011 | 0.3697 | 2.5304 | 1.0000 | 0.6271 | 0.5937 |
| 26S | GTR + I + G | 0.2181 | 0.3153 | 0.2171 | 0.2496 | 0.8700 | 9.0898 | 1.9289 | 0.3311 | 3.0681 | 1.0000 | 0.5842 | 0.7472 |
| 5.8S | TIM2ef + I + G4 | 0.2500 | 0.2500 | 0.2500 | 0.2500 | 1.8204 | 4.9269 | 1.8204 | 1.0000 | 11.7664 | 1.0000 | 0.3311 | 0.5161 |
| pgiC | TVM + I + G4 | 0.2169 | 0.2487 | 0.2353 | 0.2990 | 1.5604 | 8.0577 | 1.8589 | 1.6658 | 8.0577 | 1.0000 | 0.4050 | 1.2491 |
| SQD1 | TIM3+I + G | 0.2077 | 0.2875 | 0.3089 | 0.1959 | 1.3327 | 3.7081 | 1.0000 | 1.3327 | 5.8799 | 1.0000 | 0.4093 | 1.0064 |
| Simultaneous-1 | GTR + F + I + G4 | 0.2410 | 0.2790 | 0.2310 | 0.2480 | 0.9487 | 5.8027 | 1.1042 | 0.8629 | 4.0430 | 1.0000 | 0.5810 | 0.6100 |
| Simultaneous-2 | GTR + I + G4 | 0.2488 | 0.2598 | 0.2411 | 0.2503 | 0.7940 | 5.0369 | 0.7221 | 0.7775 | 3.7732 | 1.0000 | 0.4813 | 0.6299 |
Equally weighted maximum parsimony (MP) jackknife (JK) analyses (Farris et al., 1996) for each locus and the combined sequence data were conducted using PAUP∗ v.4.0b10 (Swofford, 2002), with insertions and deletions coded as missing data, the removal probability set to approximately 37%, and “jac” resampling emulated analyses. One thousand replicates were performed with 10 TBR searches per replicate and a maximum of 100 trees held per TBR search.
For the concatenated analysis of all nucleotide characters, maximum likelihood (ML; Felsenstein, 1973) tree searches and ML bootstrapping (BS) were conducted using the web server RAxML-HPC2 on TG v.7.2.8 on CIPRES web server (Miller et al., 2010), with 1000 rapid bootstrap analyses and -m GTRCAT as the default model followed by a search for the best-scoring tree in a single run (Stamatakis et al., 2008).
Bayesian inference (BI) was conducted using MrBayes 3.2.7 (Ronquist and Huelsenbeck, 2003) on CIPRES (Miller et al., 2010), transition/transversion rate ratio set as a free parameter, and GTR + I+Γ model (lset nst = 6 rates = invgamma) for all partitions, all model parameters were unlinked across four partitions (unlink statefreq = [all], revmat = [all], shape = [all], pinvar = [all]), and all partitions were allowed to have different rates (prset applyto = [all] ratepr = variable). Two independent runs, each with four chains (one cold, three heated), were conducted, each beginning with a random tree and sampling one tree every 1000 generations of 10,000,000 generations. Convergence among generations and stationarity were assessed using Tracer v.1.4 (Rambaut and Drummond, 2007) and the first 25% of the trees was discarded as burn-in to ensure that stationarity in log-likelihood had been reached. The remaining trees were used to calculate a 50% majority-rule consensus topology and posterior probabilities (PP).
2.4. Morphological studies
Morphological data (habit, sterile leaves, states of rhizophores, strobilus morphology, sporophylls, etc.) were obtained from field observations, herbarium investigations (specimens at CDBI, MO, NY, PYU, SING, and UC), and literature studies. Megaspores and microspores for some representative species were observed and studied using scanning electron microscope (SEM) (QUANTA 200 Scanning Electron Microscope, FEI Co., USA) at Yunnan University, Kunming, China.
2.5. Character evolution analysis
Based on previous morphological works and our studies of the morphology of Selaginella species worldwide, 10 morphological characters were analyzed: presence of rhizophores (absent, present), positions of rhizophores (dorsal side of stems and/or branches, ventral side of stems or/and branches, uncertain), dimorphism of overall sterile leaves (dimorphic, monomorphic), dimorphism of sterile leaves at the base of main stems (dimorphic, monomorphic), arrangement of sterile leaves [(spiral, four rows, decussate (at least on stem)], number of megasporophylls on strobili (only one to few at base, several on dorsal or/and ventral side), dimorphism of sporophylls on strobili (dimorphic, monomorphic), resupination of strobili with dimorphic sporophylls (resupinate, non-resupinate, N/A), surfaces of megaspores (reticulate, non-reticulate), reticulate megaspores [the _Bryodesma_ type (with prominent and dense muri: Fig. 5Q), the Gymnogynum type (with often high and strong or wing-like muri and slightly regular meshes: Fig. 5N–Q), the Stachygynandrum type (with more or less open and irregular meshes: Fig. 10H, K–N), the Tetragonostachyae type-1 (with very fine muri and open meshes: Fig. 9M and N), and the Tetragonostachyae type-2 (with fine muri formed by verrucate ornamentation: Fig. 9L), N/A]. Similar to megaspores, microspores of Selaginella also showed high variation (e.g., Valdespino et al., 2015; Zhou et al., 2015; Wang et al., 2018), but microspore data were often unavailable and thus were not included in our analysis. Mesquite v.2.75 (Maddison and Maddison, 2011) was used to optimize morphological features on the most likely ML tree. The ancestral states were estimated based on parsimony criterion. For most species with multiple samples, only one or two samples of each species were selected on the tree for analysis.
Fig. 5.

Morphology of subfamily Gymnogynoideae 关节卷柏亚科 (Selaginella subg. Gymnogynum “ = subg. _Ericetorum_”; Zhou and Zhang, 2015). A, G, H, Q and V. Bryodesma vardei (A. Habit; G. Strobilus tetragonal and sporophylls monomorphic; H. Plant; Q. Megaspore; V. Microspore). B & J. Gymnogynum kraussianum (B, Plant; J. Strobili with only one megasporophyll at base). C, P and U. Ericetorum gracillimum (C. Plants; P. Megaspore; U. Microspore). D, L and W. Lepidoselaginella lepidophylla (D. Rosset plants; L. Strobili tetragonal and sporophylls monomorphic; W. Microspore). E, F, O and T. Megaloselaginella exaltata (E. Plant; F. Rhizophore bore on dorsal side of stem and stem swollen on branch; O. Megaspore; T. Microspore). I. Gymnogynum remotifolium (I. Rhizophore bear on dorsal side of stem and stem swollen on branch). K. Ericetorum lyallii (K, Deccuately arranged leaves on stem). **M and R. Gymnogynum marginatum (**M. Microspore; R. Megaspore). N and S. Afroselaginella myosurus (N, Megaspore; S. Microspore).
Fig. 10.

Morphology of Chuselaginella 朱氏卷柏属in subfamily Selaginelloideae (Selaginella subg. Stachygynandrum; Zhou and Zhang, 2015). A, K, and O. Chuselaginella petelotii (A. Habit; K. Megaspore; O. Microspore); B, H, M, and Q. C. doederleinii (B. Habit; H. Strobili; M. Megaspore; Q. Microspore); C, D, I, and J. C. frondosa (C. Habit; D. Sterile leaves on stem; I. Megaspore; J. Microspore); E. C. intermedia (Habit); F, G, L, and P. C. rolandi-principis (F. Habit; G. Sterile leaves and strobili; L. Megaspore; P. Microspore); N and R, C. trachophylla (N. Megaspore; R. Microspore).
Fig. 9.

Morphology of subfamily Lycopodioidoideae 异穗卷柏亚科 (Selaginella subg. Heterostachys; Zhou and Zhang, 2015). A. Hypopterygiopsis monospora (A. Strobilus), B and C. H. ciliaris (B. Dorsal side of strobilus; C. Ventral side of strobilus), D, K and O. H. repanda (D. Strobilus; K. Megaspore; O. Microspore); E. Lycopodioides helvetica (E. Strobilus); F and G. L. nipponica (F. Dorsal side of strobilus; G. Ventral side of strobilus); H, T and X. Valdespinoa douglosii (H. Strobilus; T. Proximal surface of megaspore; X. Distal surface of megaspore), I. Didiclis bisulcata (I. Strobilus); J. D. braunii (J. Strobilus); L and P. H. neocaledonica (L. Megaspore; P. Microspore); M and Q. H. arbuscula (M. Megaspore; Q. Microspore); N and R. H. ciliaris (N. Megaspore; R. Microspore); S and W. H. effusa (S. Megaspore; W. Microspore); U and Y. Didiclis vogelii (U. Megaspore; Y. Microspore); V and Z. D. siamensis (V. Megaspore; Z. Microspore).
3. Results
3.1. Molecular phylogeny
A total of 1551 sequences including 665 sequences of rbcL, 128 of pgiC, 143 of SQD1, 187 of 18S, 284 of 5.8S, and 144 of 26S were used in the study, of which 420 (ca. 30% of total sequences) sequences were newly sequenced for this study. An additional 1131 sequences were downloaded from GenBank. Voucher information and GenBank accession numbers for all sequences are listed in Table S1. Characteristics of alignments are summarized in Table 1. The tree topologies using maximum likelihood, Bayesian inference, and maximum parsimony analyses were generally concordant when using the concatenated dataset.
Table 1.
Data matrices and tree statistics for each of the dataset. PI chars. include the outgroups.
| Region | # Accessions | # Characters | # PI chars. (%) |
|---|---|---|---|
| rbcL | 665 | 1323 | 715 (54.0%) |
| 18S | 187 | 1637 | 392 (23.9%) |
| 26S | 144 | 909 | 222 (24.4%) |
| 5.8S | 284 | 166 | 63 (38.0%) |
| pgiC | 128 | 480 | 293 (61.0%) |
| SQD1 | 143 | 543 | 338 (62.2%) |
| 18S + 26S + ITS + pgiC + SQD1 | 292 | 3735 | 1308 (35.0%) |
| 18S + 26S + ITS + pgiC + SQD1+rbcL | 665 | 5058 | 2023 (40.0%) |
A comparison of the trees resulting from MPJK analyses of the plastid marker, individual nuclear markers, combined nuclear markers, and combined plastid and nuclear dataset did not identify any well-supported conflicts in MP analyses (MPJK >70%; Mason-Gamer and Kellogg, 1996; Zhang and Simmons, 2006). The ML tree based on the plastid marker is shown in Fig. S1, that based on the combined nuclear markers is in Fig. S2, that based on combine plastid and nuclear markers is in Fig. S3, and the simplified Fig. S3 is in Fig. 1. The results described below were based on combined plastid and nuclear markers.
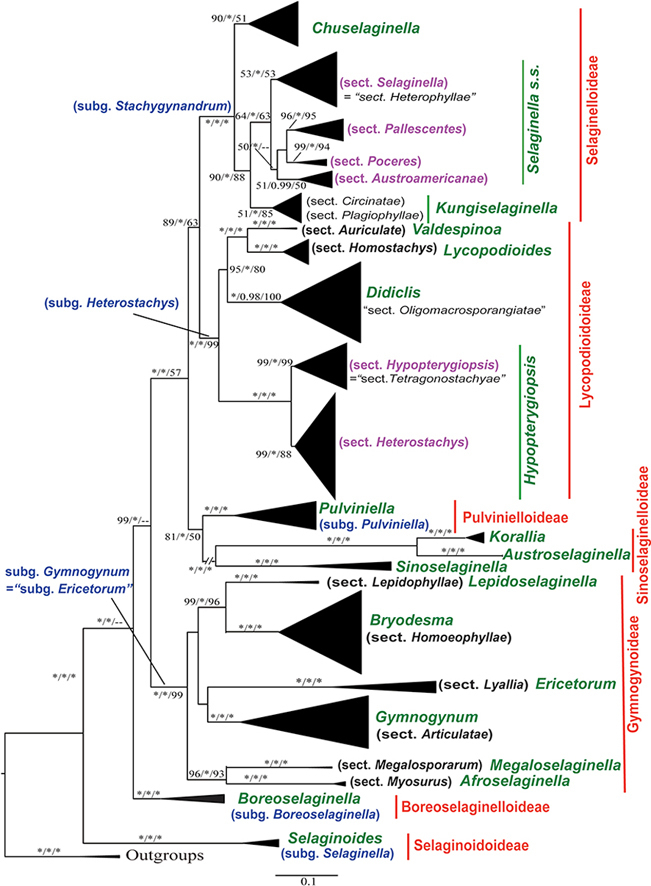
Fig. 1.

Simplified maximum likelihood phylogeny of Selaginellaceae based on plastid rbcL and five nuclear markers. The sizes of black triangles are in proportion to the sample sizes of individual clades. Support values (maximum likelihood bootstrap support, maximum parsimony jackknife support, and posterior probability) are shown along the branches. “∗” indicates a 100% support value. Red and green names indicate the 7 subfamilies and 19 genera, respectively, recognized in the current classification. Blue and black Latin names indicate the subgenera and sections recognized by Zhou and Zhang (2015). Black Chinese names indicate Chinese vernacular names of the genera. Purple names indicate the 6 sections recognized by Zhou and Zhang (2015) and this study at the same rank.
The monophyly of Selaginella was strongly supported (MLBS: 100%; MPJK: 100; BIPP: 1.00). Within Selaginella, seven deep-level clades corresponding to S. subg. Selaginella, S. subg. Boreoselaginella, S. subg. Gymnogynum (“S. subg. _Ericetorum_” sensu Zhou and Zhang, 2015), S. subg. Pulviniella, S. subg. Heterostachys, S. subg. Stachygynandrum sensu Zhou and Zhang (2015), and the Sinensis group were strongly supported (MLBS: 100%; MPJK ≥99; BIPP: 1.00).
Selaginella subg. Selaginella, S. subg. Boreoselaginella, and S. subg. Gymnogynum were resolved as the first, second, and third diverging lineages, respectively, all with strong ML and BI support. Selaginella subg. Pulviniella was resolved as sister to the Sinensis group with strong ML and BI support. They together were sister to S. subg. Heterostachys + S. subg. Stachygynandrum. The MPJK values were generally low for the inter-subgeneric relationships. Within the Sinensis group, three deep geographically differentiated clades were identified: one with Asian and northern African species, one with Selaginella australiensis, and one with species from Indian Ocean islands. Within S. subg. Gymnogynum, six major clades corresponding six sections sensu Zhou and Zhang (2015) were strongly supported as monophyletic: sect. Megalosporarum was strongly supported as sister to S. sect. Myosurus, and S. sect. Lepidophyllae was strongly supported as sister to S. sect. Homoeophyllae, but other inter-sectional relationships were not strongly supported. Within S. subg. Heterostachys, five sections defined by Zhou and Zhang (2015) were all strongly supported as monophyletic (MLBS ≥99%; MPJK ≥88; BIPP: 0.98) and their relationships were well resolved and strongly supported (MLBS ≥95%; MPJK ≥80; BIPP: 1.00), but same as those in Zhou et al. (2016). Within S. subg. Stachygynandrum, S. sect. Ascendentes was resolved as sister to the rest, followed by S. sect. Circinatae and S. sect. Plagiophyllae which formed a clade sister to the remaining four sections sensu Zhou and Zhang (2015).
3.2. Morphological evolution
Ten common morphological characters and their states are shown in Fig. 2A–J. It took one step for the parsimony reconstruction of rhizophores (Fig. 2A); three steps for that of positions of rhizophores (Fig. 2B), four steps for that of dimorphism of overall sterile leaves (Fig. 2C); 20 steps for that of dimorphism of sterile leaves at the base of main stems (Fig. 2D); three steps for that of arrangement of sterile leaves (Fig. 2E); four steps for that of number of megasporophylls on strobili (Fig. 2F); 15 steps for that of dimorphism of sporophylls on strobili (Fig. 2G); 15 steps for that of resupination of strobili with dimorphic sporophylls (Fig. 2H); seven steps for that of surfaces of megaspores (Fig. 2I); and eight steps for that of reticulate megaspores (Fig. 2J).
Fig. 2.




Morphological characters of Selaginellaceae optimized onto the maximum likelihood tree. A. Presence of rhizophores. B. Position of rhizophores. C. Dimorphism of overall sterile leaves. D. Dimorphism of sterile leaves at the base of main stems. E. Arrangement of sterile leaves. F. Number of megasporophylls on strobili. G. Dimorphism of sporophylls on strobili. H. Resupination of strobili with dimorphic sporophylls. I. Surfaces of megaspores. J. Reticulate ornamentation of megaspores.
4. Discussion
4.1. Major new findings in the phylogeny
With the largest phylogeny so far in terms of the number of species sampled, our study made a number of new findings in comparison with earlier such works (Weststrand and Korall, 2016a; Zhou et al., 2016). These new findings might provide foundation for future studies of interspecific relationships and discoveries of hidden diversity. Below we focus on and update the major relationships in the genus. In order to compare our current results with previous ones, we use subgeneric and sectional names in the Zhou and Zhang (2015) classification except the Sinensis group, which was not treated in our earlier classification, and S. subg. Ericetorum which is changed to S. subg. Gymnogynum here (Weststrand and Korall, 2016a).
Overall, the major new findings include: (1) The Sinensis group is resolved as sister to S. subg. Pulviniella with strong support in two of the three analyses; (2) S. sect. Circinatae and S. sect. Plagiophyllae formed a clade with strong support in two of the three analyses; (3) S. sect. Megalosporarum and S. sect. Myosurus are resolved as sister to each other; (4) S. sect. Ascendentes is resolved as sister to the rest of S. subg. Stachygynandrum with strong support; and (5) S. sect. Ascendentes is found to contain African species. We will discuss them in detail below.
4.2. Deep relationships in Selaginella
4.2.1. Selaginella subg. Selaginella (Fig. 3)
Fig. 3.

Morphology of subfamily Selaginoidoideae 圆穗卷柏亚科 (Selaginella subg. Selaginella; Zhou and Zhang, 2015). A–C, G and L–O. Selaginoides spinulosa (A. Habit; B. Showing plant without rhizophore; C and G. Showing densely and spirally arranged sporophylls. L and M. Spiny or verrucate ornamentation on megaspore surface. N and O. Spiny ornamentation on microspore surface (N. Distal surface in tetrad and O. Proximal surface). D, E, F, and H–K. S. deflexa (A. Habit; D and E. Densely and spirally arranged sporophylls; F. Spirally arranged sterile leaves on stem; H and I. Spiny ornamentation on megaspore surface; J and K. Spiny ornamentation on microspore (J. proximal surface and K. distal surface).
Selaginella subg. Selaginella is the first diverging lineage in Selaginella. It contains only two species, S. selaginoides and S. deflexa Brack. Its relationships have been quite consistent (Korall et al., 1999; Korall and Kenrick, 2002, 2004; Arrigo et al., 2013; Weststrand and Korall, 2016a; Zhou et al., 2016). This clade is well characterized by spiral and monomorphic sporophylls and absence of rhizophores (Fig. 2, Fig. 3).
4.2.2. Selaginella subg. Boreoselaginella (Fig. 4)
Fig. 4.

Morphology of subfamily Boreoselaginelloideae 北方卷柏亚科 (Selaginella subg. Boreoselaginella; Zhou and Zhang, 2015). A, D, E, I and L. Boreoselaginella sanguinolenta (A. Habit; D. Rhizophore bore on dorsal side of stem; E. Strobilus tetragonal and sporophylls monomorphic; I. Megaspore; L. Microspore). B, C, F, G, J and M. B. nammularifolia (B. Habit; C. Plant; F and G. Strobilus (F. Dorsal side and G. Ventral side); J. Megaspore; M. Microspore). H, K and N. B. rossii (H. Plant; K. Megaspore; N. Microspore).
It corresponds to the Sanguinolenta group (Weststrand and Korall, 2016a). The resolutions of Selaginella subg. Boreoselaginella have been controversial. Depending on datasets and analysis methods, there have been three resolutions: (1) sister to the remaining taxa with rhizophores [e.g., BI based on _rbcL_, _SQD1_, or _rbcL_ + _SQD1_: Weststrand and Korall (2016a); BI and ML based on rbcL + ITS: Zhou et al. (2016)]; (2) sister to a clade containing S. subg. Pulviniella + S. subg. Heterostachys + S. subg. Stachygynandrum sensu Zhou and Zhang (2016) based on pgiC data (Weststrand and Korall, 2016a) and based on plastome data (Zhou et al., 2022); and (3) sister to the ventral-rhizophore clade with strong support based on plastid gnomes of 26 species of Selaginella but only a few species of Selaginella were sampled and did not include the Sinensis group (Zhang et al., 2020).
Morphologically, Selaginella subg. Boreoselaginella has large number of inexplicable features in Selaginella, e.g., nearly monomorphic sterile leaves, rhizophores borne on dorsal side of stems and branches, xerophytic habit, monomorphic sporophylls, larger megaspores, and rugate surface of microspore. It is similar to S. sect. Articulatae, S. sect. Megalosporarum, and S. sect. Myosurus in having large spore size, dimorphic sterile leaves, and rhizophores borne on dorsal side of stems and branches. It is similar to S. sect. Homoeophyllae in having xerophytic habit, rhizophores borne on dorsal side, and rugate surface of microspores (Fig. 4L–N). Some species (S. sanguinolenta and S. nummularifolia) with nearly monomorphic sterile leaves are similar to S. sect. Lepidophyllae. However, this subgenus also has some characters (e.g., strobili with more than one megasporophylls, sterile leaves dimorphic and in four rows) similar to S. subg. Heterostachys and S. subg. Stachygynandrum sensu Zhou and Zhang (2015). In particular, except rhizophores borne on dorsal side and strobili with more than one megasporophyll, this subgenus is strongly similar to some species (e.g., S. albocinata) of the Sinensis group in Asia in having xerophytic habit and monomorphic sporophylls, and they usually share same distribution area in Asia.
In our current study Selaginella subg. Boreoselaginella is well supported as the second diverging lineage in our ML and BIanalyses (second resolution above), although the support value is <50% in our MP analysis (Fig. 1). Being the second diverging lineage has so far most frequently supported.
4.2.3. Selaginella subg. Gymnogynum (Fig. 5)
We used Selaginella subg. Ericetorum in our earlier papers (Zhou and Zhang, 2015; Zhou et al., 2016) which turned out to be a synonym of S. subg. Gymnogynum (Weststrand and Korall, 2016a). This subgenus is strongly supported as the third diverging lineage in our current study, consistent with our earlier results (Zhou et al., 2016). It shares a synapomorphy of rhizophores borne on dorsal side of stems; this character state is undetermined in some species in S. sect. Lyallia, one of the six sections in the subgenus (Zhou and Zhang, 2015). Reticulate megaspore surfaces are a synapomorphy of this subgenus (see below). Within this subgenus, six well-supported clades/sections, S. sect. Lyallia, S. sect. Megalosporarum, S. sect. Myosurus, S. sect. Articulatae, S. sect. Homoeophyllae, and S. sect. Lepidophyllae are well identified. Selaginella sect. Homoeophyllae and S. sect. Lepidophyllae are strongly supported as sister to each other (Weststrand and Korall, 2016a; Zhou et al., 2016; our Figs. 1 and S3). Selaginella sect. Megalosporarum and S. sect. Myosurus are resolved as sister to each other, which was not found in our earlier study (Zhou et al., 2016), but consistent with the results of Weststrand and Korall (2016a). Other internal relationships of this subgenus are poorly resolved as previous studies (Weststrand and Korall, 2016a; Zhou et al., 2016) and more studies are needed.
Selaginella subg. Exaltatae sensu Weststrand and Korall (2016b) includes our S. sect. Megalosporarum and S. sect. Myosurus (Zhou and Zhang, 2015). These two sections, not recognized by Weststrand and Korall (2016b), have different geographical distributions (the former in America and the latter in Africa) and specialized gross morphology and microspores (Fig. 5). Selaginella sect. Megalosporarum has microspores with rod-like ornamentation on surfaces, but S. sect. Myosurus has microspores with equatorial ring and verrucous ornamentation on surfaces (Fig. 5S vs. 5T). In addition, the divergence between them has been dated as ca. 200 million years ago (Klaus et al., 2017).
4.2.4. The Sinensis group (Fig. 6)
Fig. 6.

Morphology of subfamily Sinoselaginelloideae 中华卷柏亚科 (the Sinensis group; Weststrand and Korall, 2016b). A, B, I, and M. Sinoselaginella sinensis (A. Habit; B. Showing part of plant with ventral rhizophore bore on ventral side and strobilus with only one megasporophyll on base; I. Megaspore; M, Microspore). C, D, J, and N. S. albocincta (C. Part of plant; D. Strobilus with only one megasporophyll at base; J. Megaspore; N. Microspore). E, F and T. Austroselaginella australiensis (E. Ventral side of part of plant; F. Strobilus with only one megasporophyll (megasporangium) at base; T. Microspore). K and O. Sinoselaginella yemensis (K. Megaspore; O. Microspore). L, P and S. Korallia cataphracta (L and P. Megaspore; S. Microspore).
Weststrand and Korall (2016a) resolved the Sinensis group as sister to the rest of their Selaginella subg. Stachygynandrum (= Selaginella subg. Pulviniella + S. subg. Heterostachys + S. subg. Stachygynandrum sensu Zhou and Zhang (2015)). In our current study, the Sinensis group is for the first time resolved as sister to S. subg. Pulviniella with relatively strong support in ML and BI analyses, although only 50% support value in MP analysis (Figs. 1 and S3). The monophyly of the Sinensis group is strongly supported. Morphologically, the Sinensis group can be easily distinguished from other taxa in Selaginella in having rhizophores borne on the ventral side of stems, only one (to a few) megasporophyll at the base of strobili, large megaspores (usually >600 μm in diam.) (Fig. 2, Fig. 6). Within the Sinensis group, three well-supported subclades (the Selaginella sinensis subclade, the S. australiensis subclade, and the S. fissidentoides subclade) are identified, consistent with results by Weststrand and Korall (2016a). These subclades can be easily distinguished from one another by gross morphology, microspores, megaspores, and geographical distributions (see below). Although strobili with only one megasporophyll (megasporangium) at the base are also present in S. subg. Gymnogynum, the latter has rhizophores borne on dorsal side of stems, reticulate ornamentation on megaspore surfaces, and often spiny ornamentation on microspore surfaces (Fig. 5).
Recent plastome study has shown that 48 species of Selaginella sampled had high GC content (>50%) but all three species in the Sinensis group had ∼ 30% GC content in plastomes, similar to most vascular plants; the Sinensis group has the smallest plastomes in land plants except some parasitic plants, and their plastomes do not contain any tRNAs (Zhou et al., 2022).
4.2.5. Selaginella subg. Pulviniella (Fig. 7)
Fig. 7.

Morphology of subfamily Pulvinielloideae 垫状卷柏亚科 (Selaginella subg. Pulviniella; Zhou and Zhang, 2015). A, B, D, E, and I. Pulviniella pulvinata (A and B. Habit; D. Strobili tetragonal and sporophylls monomorphic; E. Megaspore; I. Microspore). **C, F, and J. P. digitata (C. Plants; F. Megaspore; J. Microspore). G and K. P. imbricata (**G. Megaspore; K. Microspore). H and L. P. stauntoniana (H. Megaspore; L. Microspore).
Selaginella subg. Pulviniella was resolved as sister to S. subg. Heterostachys + S. subg. Stachygynandrum sensu Zhou and Zhang (2015) with moderate support in our earlier study (Zhou et al., 2016) but the Sinensis group was not sampled. Weststrand and Korall (2016a) sampled the Sinensis group but only BI analysis was presented. Our current study resolved S. subg. Pulviniella as sister to the Sinensis group with relatively strong support in ML and BI analysis, although only 50% support value in MP analysis (Figs. 1 and S3). This resolution has not been found before. Morphologically and ecologically, this subgenus comprises species mostly with rosette-forming habit and extremely drought habitat. This subgenus and the Sinensis group share drought habitat, but morphological synapomorphies to support their sister relationship are unclear.
4.2.6. Selaginella subg. Heterostachys (Fig. 8, Fig. 9)
Fig. 8.

Growth forms of subfamily Lycopodioidoideae 异穗卷柏亚科 (Selaginella subg. Heterostachys; Zhou and Zhang, 2015). A. Hypopterygiopsis bodinieri; B. H. heterostachys; C. H. monospora; D. H. ciliaris; E. H. repanda; F. H. vaginata; G. Lycopodioides nipponica; H. Didiclis bisulcata; I. D. delicatula; J. D. mairei; K. D. siamensis; L. D. willdenowii.
Consistent with our previous results (Zhou et al., 2016), our current study resolved Selaginella subg. Heterostachys as sister to S. subg. Stachygynandrum with strong support and identified five strongly supported clades within the subgenus corresponding to the five sections proposed by Zhou and Zhang (2015): S. sect. Auriculate, S. sect. Homostachys, S. sect. Heterostachys, Didiclis (“S. sect. _Oligomacrosporangiatae_”), and S. sect. Tetragonostachyae. Although Weststrand and Korall (2016a, b) did not recognize this subgenus and our sections, their results were consistent with ours with five sections sensu Zhou and Zhang (2015) strongly supported. This subgenus, strongly supported as monophyletic (Weststrand and Korall, 2016a; Zhou et al., 2016; our Figs. 1 and S3) and characterized by non-reticulate megaspores (Fig. 2I), mainly comprises species with dimorphic sporophylls of strobili in Asia and those species with verrucate ornamentation on their microspore surfaces but this reversed to finely reticulate ornamentation (Tetragonostachyae type-1 and type-2) in some pacific island species of S. sect. Tetragonostachyae (Fig. 2I and J).
4.2.7. Selaginella subg. Stachygynandrum (Fig. 10)
This subgenus is also always strongly supported as monophyletic (Korall and Weststrand, 2016a; Zhou et al., 2016; our Figs. 1 and S3). Reticulate megaspore surfaces are a synapomorphy of S. subg. Stachygynandrum (see below). In our earlier study, we recognized seven sections/clades in this subgenus: S. sect. Ascendentes, S. sect. Austroamericanae, S. sect. Circinatae, S. sect. Heterophyllae, S. sect. Pallescentes, S. sect. Plagiophyllae, and S. sect. Poceres (Zhou and Zhang, 2015) and resolved two paraphyletic clades, S. sect. Circinatae and S. sect. Plagiophyllae, as the first diverging lineages with weak support (Zhou et al., 2016). In our current study, these two clades formed a clade strongly supported as monophyletic in two of the three analyses and as the second diverging lineage in the subgenus with strong support. Selaginella sect. Ascendentes is resolved as sister to the rest of the subgenus. These results are consistent with those in Korall and Weststrand (2016a). Selaginella sect. Ascendentes was only known to be distributed in Asia and Pacific islands (Zhou et al., 2016), but now an African species (Selaginella versicolor) is also resolved in this clade (Korall and Weststrand, 2016a; our Figs. 1 and S3). Korall and Weststrand (2016b) recognized this subgenus with a very broad circumscription of 600+ species including three of our subgenera: S. subg. Heterostachys, S. subg. Pulviniella, and S. subg. Stachygynandrum, plus the Sinensis group. They recognized none of our sections in this subgenus either, although five out of our seven sections were strongly supported and the remaining two formed a strongly supported monophyletic clade in their study (our Table 3).
Table 3.
A comparison of this classification to two recent classifications [Zhou and Zhang's (2015) and Weststrand and Korall's (2016b)] and Weststrand and Korall's (2016a) phylogeny.
| This classification | Zhou and Zhang (2015) | Weststrand and Korall (2016b) | Weststrand and Korall (2016a) |
|---|---|---|---|
| subfam. I. Selaginoidoideae [_Selaginoides_] | subg. Selaginella | subg. Selaginella | Selaginella clade (pp = 1.00) |
| subfam. II. Boreoselaginelloideae [**Boreoselaginella**] | subg. Boreoselaginella | in subg. Stachygynandrum | the Sanguinolenta group clade (pp = 1.00) |
| subfam. III. Gymnogynoideae | subg. Ericetorum | subg. Rupestrae +Gymnogynum/+Exaltatae +Lepidophyllae +Ericetorum | Rupestrae +Lepidophyllae+Gymnogynum+Exaltatae+Ericetorum clade (pp = 1.00) |
| Afroselaginella | sect. Myosurus | in subg. Exaltatae | the S. congoensis to S. myosurus E clade (pp = 1.00) |
| Bryodesma | sect. Homoeophyllae | subg. Rupestrae | Rupestrae clade (pp = 1.00) |
| Ericetorum | sect. Lyallia | subg. Ericetorum | Ericetorum clade (pp = 1.00) |
| Gymnogynum | sect. Articulatae | subg. Gymnogynum | Gymnogynum clade (pp = 1.00) |
| Lepidoselaginella | sect. Lepidophyllae | subg. Lepidophyllae | Lepidophyllae clade (pp = 1.00) |
| Megaloselaginella | sect. Megalosporarum | subg. Exaltatae | the S. exaltata A-B clade (pp = 1.00) |
| subfamily IV. Sinoselaginelloideae | Unsampled | in subg. Stachygynandrum | the S. yemensis A to S. schellarum clade (pp = 1.00) |
| Austroselaginella | Unsampled | in subg. Stachygynandrum | the S. australiensis A-B clade (pp = 1.00) |
| Korallia | Unsampled | in subg. Stachygynandrum | the S. distachya to S. sechellarum clade (pp = 1.00) |
| Sinoselaginella | Unsampled | in subg. Stachygynandrum | the S. yemensis A to S. sinensis clade (pp = 1.00) |
| subfam. V. Pulvinielloideae [_Pulviniella_] | subg. Pulviniella | in subg. Stachygynandrum | the S. nubigena to S. digitata A-C clade (pp = 1.00) |
| subfam. VI. Lycopodioidoideae | subg. Heterostachys | in subg. Stachygynandrum | the S. douglasii A to S. arbuscula clade (pp = 1.00) |
| Didiclis | sect. Oligomacrosporangiatae | in subg. Stachygynandrum | the S. siamensis to S. mayeri B clade (pp = 1.00) |
| Hypopterygiopsis | sect. Heterostachys + sect. Tetragonostachyae | in subg. Stachygynandrum | the S. monospora A to S. arbuscula clade (pp = 1.00) |
| sect. Heterostachys | sect. Heterostachys | in subg. Stachygynandrum | the S. monospora A to S. labordei E clade (pp = 1.00) |
| sect. Hypopterygiopsis | sect. Tetragonostachyae | in subg. Stachygynandrum | the S. repanda to S. arbuscula clade (pp = 1.00) |
| Lycopodioides | sect. Homostachys | in subg. Stachygynandrum | the S. tama-montana to S. helavetica B clade (pp = 1.00) |
| Valdespinoa | sect. Auriculate | in subg. Stachygynandrum | the S. douglasii clade (pp = 1.00) |
| subfamily VII. Selaginelloideae | subg. Stachygynandrum | in subg. Stachygynandrum | the clade S. versicolor A-C to S. contigua clade (pp = 1.00) |
| Chuselaginella | sect. Ascendentes | in subg. Stachygynandrum | the S. versicolor A-C to S. roxburghii C clade (pp = 1.00) |
| Kungiselaginella | sect. Circinatae + sect. Plagiophyllae | in subg. Stachygynandrum | the S. moellendorffii A-B to S. biformis B clade (pp = 1.00) |
| Selaginellas.s. | in subg. Stachygynandrum | in subg. Stachygynandrum | the S. versicolor A-C to S. contigua clade (pp = 1.00) |
| sect. Austroamericanae | sect. Austroamericanae | in subg. Stachygynandrum | the S. hirsuta to S. revoluta C clade (pp = 0.99) |
| sect. Pallescentes | sect. Pallescentes | in subg. Stachygynandrum | the S. huehuetenangensis A-B to S. pulcherrima clade (pp = 1.00) |
| sect. Poceres | sect. Poceres | in subg. Stachygynandrum | the S. novae-hollandiae B to S. oaxacana B clade (pp = 1.00) |
| sect. Selaginella | sect. Heterophyllae | in subg. Stachygynandrum | the S. cladorrhizans to S. contigua clade (pp = 0.99) |
4.3. Morphological evolution in Selaginella
4.3.1. Character 1_—Presence of rhizophores_ (Fig. 2A)
Two species of Selaginella, S. selaginoides and S. deflexa, have no rhizophores (Fig. 3C), and all other species have. These two species are the only member of S. subg. Selaginella (Zhou and Zhang, 2015). The ancestral state of rhizophores in Selaginella is ambiguous because the character state in Isoëtes is unknown or not applicable. Absence of rhizophores can be a synapomorphy or a symplesiomorphy of S. subg. Selaginella. Presence of rhizophores can also be a synapomorphy or a symplesiomorphy of the rest of Selaginella excluding S. subg. Selaginella (Fig. 2A).
4.3.2. Character 2_—Position of rhizophores_ (Fig. 2B)
The rhizophores in Selaginella can be borne on dorsal or ventral side of stems or/and branches. The rhizophores borne on dorsal side are the ancestral state of rhizophores in Selaginella (Fig. 2B). The rhizophores of S. sect. Lyallia are strictly restricted to the base of erect stems and the dorsal/ventral position of rhizophores is unknown (Fig. 5C). The ventrally borne rhizophores evolved once in Selaginella and are a synapomorphy of S. subg. Pulviniella + S. subg. Heterostachys (Fig. 8D, G) + S. subg. Stachygynandrum + the Sinensis group (Fig. 6B).
4.3.3. Character 3_—Dimorphism of overall sterile leaves_ (Fig. 2C)
The sterile leaves of Selaginella can be monomorphic or dimorphic. Although only about 8% of the species have monomorphic sterile leaves in Selaginella, monomorphic sterile leaves have been frequently used to identify and define some taxa, e.g., S. subg. Selaginella (Fig. 3A–D), S. sect. Homoeophyllae Spring sensu Zhou and Zhang (2015) (Fig. 5A, H), and S. subg. Ericetorum sensu Jermy (1986) (Fig. 5C, K). Our morphological analysis suggests that the ancestral state of sterile leaves in Selaginella is ambiguous. The widespread dimorphic sterile leaves evolved only once from monomorphic state in Selaginella. Dimorphic sterile leaves evolved to monomorphic leaves once independently each in S. sect. Homoeophyllae and S. sect. Lyallia.
4.3.4. Character 4_—_ D_imorphism of sterile leaves at the base of main stems_ (Fig. 2D)
Except all species of Selaginella subg. Selaginella and S. sect. Homoeophyllae, and some of S. sect. Lyallia with monomorphic sterile leaves throughout plants in Character 3, those species with erect stems and dimorphic sterile leaves on branches can have monomorphic sterile leaves at the base of the main stem. Most species have dimorphic sterile leaves at the base of the main stem (Fig. 3, Fig. 4, Fig. 5, Fig. 6, Fig. 7, Fig. 8, Fig. 9, Fig. 10) in Selaginella and this dimorphism is a morphological synapomorphy or ancestral state of Selaginella excluding S. subg. Selaginella. Monomorphic sterile leaves at the base of the main stem have evolved at least 18 times from dimorphic sterile leaves independently in the genus. Monomorphic sterile leaves at the base of the main stem (Fig. 5C) appear to be a synapomorphy of S. sect. Lyallia and it is reversed to dimorphic sterile leaves on lower parts of main stem independently.
4.3.5. Character 5_—Arrangement of sterile leaves_ (Fig. 2E)
Arrangement of sterile leaves in Selaginella can be spiral (Fig. 3, Fig. 5A, H), in four rows (Fig. 4, Fig. 5, 6A–H, 7A–D, 8A–L and 10A–C, F), or decussate (at least on stem; Fig. 5C, K). The ancestral state of arrangement of sterile leaves in Selaginella is ambiguous and can be either spiral or in four rows. Sterile leaves in four rows can be a synapomorphy or a symplesiomorphy of Selaginella excluding S. subg. Selaginella. Sterile leaves in four rows evolved to spiral sterile leaves once in S. sect. Homoeophyllae. Decussate sterile leaves evolved from those in four rows and are a synapomorphy of S. sect. Lyallia. Decussate sterile leaves on branches are restricted to S. sect. Lyallia, while some other species with decussate sterile leaves on main stems have sterile leaves in four rows on branches. We hypothesize that decussate sterile leaves in S. sect. Lyallia are homologous, but more developmental and anatomical studies are necessary.
4.3.6. Character 6_—Number of megasporophylls on strobili_ (Fig. 2F)
Number of megasporophylls on strobili can be only one (to a few) at base of strobili (Figs. 5G and 6D–F) or more than two on dorsal side or/and ventral side of strobili (Figs. 3G, 4E–G, 5G, L, 7D, 9A–J and 10D–F). Having several megasporophylls on strobili is a synapomorphy of Selaginella s.l. Only one (to a few) megasporophyll on strobili is a morphological synapomorphy of the Sinensis group. The ancestral state of the clade containing S. sect. Articulatae + S. sect. Megalosporarum + S. sect. Myosurus + S. sect. Lyallia is ambiguous.
4.3.7. Character 7_—_ D_imorphism of sporophylls on strobili_ (Fig. 2G)
Sporophylls on strobili in Selaginella can be dimorphic (Figs. 8B, D, 9A–D, F, G, I, and 10D, E) or monomorphic (Figs. 3A, B, D, E, G, 4E–G, 5G, H, J, L, 6D, F, 7D, 9E, H, J and 10F). Traditionally, based on the dimorphic sporophylls in Selaginella, S. subg. Heterostachys sensu Jermy (1986) was widely accepted. However, phylogenetic analyses showed that these species with dimorphic sporophylls in Selaginella are polyphyletic (Weststrand and Korall, 2016a; Zhou et al., 2016; our Figs. 1 and S1–S3). Our morphological analysis suggests that monomorphic sporophylls on strobili are the ancestral state of Selaginella. Dimorphic sporophylls do not appear in other subgenera or the Sinensis group but only evolved in the two latest-diverging subgenera, S. subg. Heterostachys and S. subg. Stachygynandrum. Dimorphic sporophylls evolved at least 14 times in these two subgenera, but are dominant in S. subg. Heterostachys. The ancestral state of dimorphism of sporophylls on strobili in S. subg. Heterostachys is ambiguous.
4.3.8. Character 8_—R_ esupination of strobili with dimorphic sporophylls (Fig. 2H)
Based on the relative position between dimorphic sporophylls and sterile leaves, strobili with dimorphic sporophylls (those taxa with dimorphic sporophylls in character 7) in Selaginella can be resupinate (Figs. 8B, D, 9A–D, I and 10D, E) or non-resupinate (Fig. 9F and G). Non-resupinate strobili evolved from monomorphic sporophylls and is a synapomorphy of S. sect. Homostachys. Selaginella sect. Auriculate, the sister of S. sect. Homostachys, has monomorphic sporophylls. Resupinate strobili evolved once or twice in S. subg. Heterostachys depending on the ancestral state and evolved at least 10 times in S. subg. Stachygynandrum. The ancestral state of resupination of strobili in S. subg. Heterostachys is ambiguous.
4.3.9. Character 9_—Surfaces of megaspores_ (Fig. 2I)
Based on the main elements of ornamentation, surfaces of megaspores in Selaginella can be reticulate (Figs. 5N–R and 9L–N) or non-reticulate (Figs. 3H, L, 4J–K, 6I–L, 7E–H, 9K, S–V, X). Our morphological analysis suggests that non-reticulate surfaces are the ancestral state of megaspores in Selaginella. Reticulate megaspores in the genus are not homologous but could have evolved only three times in Selaginella independently. Reticulate surfaces of megaspores are unambiguously a synapomorphy of S. subg. Stachygynandrum and a synapomorphy of S. subg. Gymnogynum as well. The ancestral state of S. sect. Tetragonostachyae of S. subg. Heterostachys is ambiguous. Reticulate megaspores reversed to non-reticulate megaspores once in S. subg. Stachygynandrum and at least once in S. sect. Tetragonostachyae. Reticulate megaspores in S. sect. Tetragonostachyae can be a symplesiomorphy. Based on the morphology and size of muri, five types of reticulate ornamentation can be identified (see below).
4.3.10. Character 10_—Reticulate megaspores_ (Fig. 2J)
Based on the density, size, and shape of muri of reticulate ornamentation, reticulate megaspores can be divided into the Gymnogynum type (with often high and strong or wing-like muri and slightly regular meshes: Fig. 5N–P, R), the Bryodesma type (with prominent and dense muri: Fig. 5Q), the Stachygynandrum type (with more or less open and irregular meshes: Fig. 10H, K–N), the Tetragonostachyae type-1 (with fine muri and open meshes: Fig. 9M−N), and the Tetragonostachyae type-2 (with fine muri formed by connected verrucate ornamentation, Fig. 9L). The Stachygynandrum type of megaspores is a synapomorphy of S. subg. Stachygynandrum. The Gymnogynum type of megaspores is shared by S. subg. Gymnogynum excluding S. sect. Homoeophyllae, which supports a close relationship, not resolved by our molecular data, among S. sect. Articulatae, S. sect. Lyallia, S. sect. Megalosporarum, and S. sect. Myosurus. The Bryodesma type of megaspores is a synapomorphy of S. sect. Homoeophyllae (Bryodesma). The Tetragonostachyae type-1 and -2 of megaspores characterize S. sect. Tetragonostachyae but the ancestral state of the section is ambiguous.
4.4. Splitting Selaginella into seven subfamilies and 19 genera
With our new molecular and morphological results and results from previous studies (Zhou and Zhang, 2015; Weststrand and Korall, 2016a, 2016b; Zhou et al., 2016), it appears that the major monophyletic clades in the currently defined Selaginella s.l. or Selaginellaceae can be morphologically defined by some characters alone or combinations of characters. We propose to recognize these major clades as genera. The question is how many genera should be recognized. Jermy (1986) recognized four subgenera, Zhou and Zhang (2015) accepted six subgenera excluding the Sinensis group and three subgenera were further classified into six, five, and seven sections, respectively, and Weststrand and Korall (2016b) recognized seven subgenera with 600+ species in one subgenus and excluding the Sanguinolenta group. The major differences between the two recent classifications by Zhou and Zhang (2015) and by Weststrand and Korall (2016b) lie mainly in how to treat S. subg. Stachygynandrum. Weststrand and Korall (2016b) combined S. subg. Stachygynandrum sensu Zhou and Zhang (2015), S. subg. Heterostachys sensu Zhou and Zhang (2015), S. subg. Pulviniella, and the Sinensis group based on their largely unresolved relationships. We prefer to divide their S. subg. Stachygynandrum further for two reasons: (a) their S. subg. Stachygynandrum contains 600+ species with heterogeneous morphologies and distributions and is not informative; and (b) the relationships within it are now well resolved and strongly supported (at least in the nuclear phylogeny) in our current study (Figs. 1 and S3).
Here we propose to recognize our six earlier subgenera and the Sinensis group as seven subfamilies and the 19 major clades in our phylogenies as 19 genera in Selaginellaceae (list of subfamilies and genera below) for the following reasons:
- (1)
Although recognizing 19 genera for 700–800 species in Selaginellaceae/Selaginellales may be viewed as dramatic deviation from the current single-genus classification, there is a history of dividing Selaginellaceae into more than one genus (Palisot de Beauvois, 1804; Kuntze, 1891; Börner, 1912; Rothmaler, 1944; Sakurai, 1943; Kung, 1988; Soják, 1992; Satou, 1997; Tzvelev, 2004; Weakley, 2012, 2022; Weakley and Southeastern Flora Team, 2022). Schuettpelz et al. (2018) argued that there are too few fern genera. Our current classification is also in agreement with recent classifications of pteridophytes, e.g., 25 genera for ca. 265 species in Blechnaceae (PPG I, 2016; de Gasper et al., 2016; González et al., 2020), 17 genera for ca. 380 species in Lycopodiales (Øllgaard, 2012; PPG I, 2016; Chen et al., 2022; Zhang and Zhou, 2022), 37 genera for ca. 1200 species in Thelypteridaceae (Fawcett et al., 2021; Fawcett and Smith, 2021), and 42 genera for ca. 900 species in Polypodiaceae subfam. Grammitidoideae (Yang et al., 2023; Zhou et al., 2023). Many of these recent generic segregates are not necessarily clearer phylogenetically or morphologically than the genera of Selaginellaceae we recognize here. - (2)
The relationships in Selaginellaceae, especially those within Selaginella subg. Stachygynandrum sensu Weststrand and Korall (2016b), which were not well resolved in Weststrand and Korall (2016a), have been well resolved (Zhou et al., 2016), even with the Sinensis group sampled (Figs. 1 and S3). The genus-level units are well-supported phylogenetically, such that the circumscription of the genera won't need to change in the near future as more phylogenetic information becomes available. - (3)
These 19 genera are all deeply diverged lineages in Selaginellaceae and mostly among the oldest lineages in vascular plants. Molecular dating by Klaus et al. (2017) estimated the 15 out of the 19 genera to have diverged from their sisters (stem ages) about 120–373 Ma and the remaining four about 75–80 Ma. The stem ages of the 15 genera are older than any polypod families and genera and the 19 genera we recognize here are older than those of most genera of pteridophytes as estimated by Schuettpelz and Pryer (2009) and Du et al. (2021). Notably, all 42 grammitid fern genera recognized in recent studies diverged from their sisters in the past 30 million years (Sundue et al., 2014; Yang et al., 2023; Zhou et al., 2023). Lu et al. (2020), Zhang et al. (2020), and Zhang and Zhang (2022) used relatively old ages as an argument for generic recognition in Woodsiaceae, Lepisoreae (Polypodiaceae), and Ophioglossaceae, respectively. - (4)
Most of the 19 genera and seven subfamilies are morphologically definable, although combinations of morphological characters are sometimes needed (Zhou and Zhang, 2015; Weststrand and Korall, 2016a, b; Zhou et al., 2016, Fig. 2, Fig. 3, Fig. 4, Fig. 5, Fig. 6, Fig. 7, Fig. 8, Fig. 9, Fig. 10; Table 4, Table 5). In fact, as shown below, five out of the seven subfamilies (except Lycopodioidoideae and Selaginelloideae) and 16 out of the 19 genera (except Didiclis, Hypopterygiopsis, Selaginella s.s.) can be distinguished from one another using macromorphological features only. Spore features and distribution information are useful in distinguishing Lycopodioidoideae from Selaginelloideae, and are critical in distinguishing Didiclis from Hypopterygiopsis and in distinguishing Selaginella s.s. from Chuselaginella and Kungiselaginella. - (5)
Most of the 19 genera are geographically coherent: Afroselaginella is endemic to Africa; Austroselaginella is endemic to Australia; Boreoselaginella, Chuselaginella, and Kungiselaginella occurs in the Old World; Bryodesma occurs in Africa, East Asia, and North America; Didiclis occurs in Africa, Asia, and Australasia; Ericetorum occurs in Africa and Australia; Gymnogynum occurs in North to South Americas, and rarely in Asia and Africa; Hypopterygiopsis occurs in Asia and Pacific islands and a few in Africa and Madagascar; Korallia is endemic to Madagascar and adjacent islands; Lepidoselaginella is endemic to North America; Lycopodioides occurs in Eurasia; Megaloselaginella is endemic to Central to South America; Pulviniella occurs in Africa and Asia and rarely in Americas; Selaginella s.s. is nearly endmics to Americas except one or a few species in Africa; Selaginoides is endemic to the boreal area and Hawaii; Sinoselaginella occurs in China extending to the Arabic area and eastern Africa (Ethiopia, Kenya, Somalia); and Valdespinoa is endemic to North America. - (6)
This 19-genus classification is largely in line with our earlier classification (Zhou and Zhang, 2015), albeit the taxa have different ranks (Fig. 1). We recognized six subgenera and three of them were further divided into five, six, and seven sections, respectively (Zhou and Zhang, 2015). Fifteen out of our 21 subgenera/sections are recognized as genera here, whereas our S. sect. Heterostachys and S. sect. Tetragonostachyae are combined to form Hypopterygiopsis, and S. subg. Stachygynandrum sensu Zhou and Zhang (2015) collectively forms Selaginelloideae. The major difference is that the Sinensis group was not treated by Zhou and Zhang (2015) and is divided into three genera here. This 19-genus classification is in fact not contradictory with the one proposed by Weststrand and Korall (2016b) in which seven subgenera were recognized. Five of their subgenera, S. subg. Ericetorum, S. subg. Gymnogynum, S. subg. Lepidophyllae, S. subg. Rupestrae, S. subg. Selaginella, are circumscribed exactly the same as the corresponding five of our 19 genera. Their S. subg. Exaltatae is divided into our Afroselaginella and Megaloselaginella. Their unplaced Sanguinolenta group is our Boreoselaginella. The biggest difference is that we recognize 11 genera in their S. subg. Stachygynandrum, which formed an unresolved polytomy in their phylogeny (Weststrand and Korall, 2016a), but is well resolved in our current study, especially in the nuclear phylogeny (Figs. 1 and S1–S3). - (7)
Although segregating these 19 clades will cause disruption of a number of species names, in parallel with this paper we have published a proposal to conserve Selaginella with a new type (S. flabellata (L.) Spring) in the most species-rich major clade in Stachygynandrum (Wan et al., 2023), which will largely reduce nomenclatural instability. In fact, most American species belong to the newly defined Selaginella s.s. The current type of Selaginella, S. selaginoides, is resolved in the Selaginoides clade which contains only two species. There is already a generic name available, Selaginoides, for this clade. In addition to the names Selaginella and Selaginoides, five generic names, Bryodesma, Didiclis, Gymnogynum, Hypopterygiopsis, and Lycopodioides, are available and a number of species have already been transferred to these genera (Kuntze, 1891; Sakurai, 1943; Rothmaler, 1944; Kung, 1988; Soják, 1992; Satou, 1997; Tzvelev, 2004; Weakley, 2022). Three generic names, Boreoselaginella, Ericetorum, and Pulviniella, are raised from existing subgeneric names (Warburg, 1900; Jermy, 1986; Zhou and Zhang, 2015; Weststrand and Korall, 2016b). Therefore, only nine new generic names are needed and they mostly contain only one to a few species and will not cause mass disruption of names: Afroselaginella for S. sect. Myosurus with about four species, Austroselaginella for S. australiensis and additional three species, Chuselaginella for S. sect. Ascendentes with ca. 70 species, Kungiselaginella for S. sect. Circinatae and S. sect. Plagiophyllae with about two dozen species, Lepidoselaginella for S. sect. Lepidophyllae with about three species, Megaloselaginella for S. sect. Megalosporarum with about two species, Korallia for S. sect. Fissidentoides with about 15 species, and Sinoselaginella for portion of the Sinensis group with five species, and Valdespinoa for S. sect. Auriculate with about one species. - (8)
Subdividing Selaginella s.l. into homogeneous and manageable genera is good for the purposes of analysis, communication, and conservation. It will be easier for botanists to undertake monographic work, for example, on the subunits, and to identify and communicate meaningful phylogenetic/evolutionary changes. Treating Selaginella as a single genus devalues it as a triage level for judging biodiversity value and ignores the ancient diversity of its deeply diverged clades. It has been argued that there are too few genera in pteridophytes, which does not foster more precise and efficient communication, promote additional research, or facilitate herbarium curation (Schuettpelz et al., 2018).
Table 4.
Comparison of gross morphology of 7 subfamilies and 19 genera in Selaginellaceae recognized in the current classification.
| Subfamilies/Genera | Growth forms | Rhizophores borne on stem/branch | Strobili | Dimorphism of sporophylls | Arrangement of sporophylls | Dimorphism of sterile leaves | Arrangement of sterile eaves | Numbers of megasporophylls on strobili |
|---|---|---|---|---|---|---|---|---|
| Subfamily I Selaginoidoideae [_Selaginoides_] | Erect | Absent | Cylindrical | Monomorphic | Spiral | Monomorphic | Spiral | Several on dorsal or/and ventral sides |
| Subfamily II Boreoselaginelloideae [_Boreoselaginella_] | Creeping | Dorsal | Tetragonal | Monomorphic | Four rows | Dimorphic to nearly monomorphic | Four rows | Several on dorsal or/and ventral sides |
| Subfamily III Gymnogynoideae | Creeping, ascending, erect, rosette-forming | Dorsal or restricted to base of stem | Tetragonal | Monomorphic | Four rows | Monomorphic to dimorphic | Four rows | Several on dorsal or/and ventral sides; or only one (to a few) at base |
| Gymnogynum | Creeping, ascending, and erect | Dorsal | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Only one (to a few) at base |
| Bryodesma | Creeping | Dorsal | Tetragonal | Monomorphic | Four rows | Monomorphic | Spiral | Several on dorsal or/and ventral sides |
| Lepidoselaginella | Rosette-forming | Dorsal | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Several on dorsal or/and ventral sides |
| Ericetorum | Erect | Restricted to base of stem | Tetragonal | Monomorphic | Four rows | Monomorphic (rarely dimorphic on branches) | Decussate (some four rows on branches) | Several on dorsal or/and ventral sides; or only one (to a few) at base |
| Megaloselaginella | Erect | Dorsal | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Only one (to a few) at base |
| Afroselaginella | Creeping | Dorsal | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Only one (to a few) at base |
| Subfamily IV Sinoselaginelloideae | Creeping, ascending | Ventral | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Only one (to a few) at base |
| Austroselaginella | Creeping | Ventral | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Only one (to a few) at base |
| Korallia | Creeping, ascending | Ventral | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Only one (to a few) at base |
| Sinoselaginella | Creeping | Ventral | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Only one (to a few) at base |
| Subfamily V Pulvinielloideae [_Pulviniella_] | Rosette-forming or erect | Ventral | Tetragonal | Monomorphic | Four rows | Dimorphic (rarely monomorphic on stems) | Four rows | Several on dorsal or/and ventral sides |
| Subfamily VI Lycopodioidoideae | Ascending, creeping, erect, prostrate, scandent | Ventral | Complanate to tetragonal | Dimorphic to slightly dimorphic (resupinate) or monomorphic | Four rows | Dimorphic | Four rows | Several on dorsal or/and ventral sides |
| Valdespinoa | Prostrate | Ventral | Tetragonal | Monomorphic | Four rows | Dimorphic | Four rows | Several on dorsal or/and ventral sides |
| Hypopterygiopsis | Creeping, ascending, erect, or prostrate | Ventral | Complanate to tetragonal | Dimorphic to slightly dimorphic (resupinate) or monomorphic | Four rows | Dimorphic | Four rows | Several on dorsal or/and ventral sides |
| Lycopodioides | Creeping, or prostrate | Ventral | Complanate but lax | Dimorphic (non-resupinate), rarely monomorphic) | Four rows | Dimorphic | Four rows | Several on dorsal or/and ventral sides |
| Didiclis | Ascending, creeping, erect, or scandent | Ventral, rarely restricted to base of stem | Tetragonal (only complanate in the Bisulcata group) | Monomorphic [only dimorphic (resupinate) in the Bisulcata group] | Four rows | Dimorphic (rarely monomorphic on stem) | Four rows | Several on dorsal or/and ventral sides |
| Subfamily VII Selaginelloideae | Erect, ascending, or creeping (rarely rosette-forming) | Ventral, or restricted to base of stem | Tetragonal, rarely complanate | Monomorphic [rarely dimorphic (resupinate) in America] | Four rows | Dimorphic (or monomorphic on stem) | Four rows | Several on dorsal or/and ventral sides |
| Chuselaginella | Mainly ascending, some erect | Ventral, rarely restricted to base of stem | Tetragonal | Monomorphic | Four rows | Dimorphic (or monomorphic on stem) | Four rows | Several on dorsal or/and ventral sides |
| Kungiselaginella | Mainly erect, few creeping | Restricted to base of stem, few ventral | Tetragonal | Monomorphic | Four rows | Dimorphic (or monomorphic on stem) | Four rows | Several on dorsal or/and ventral sides |
| Selaginella s.s. | Erect, ascending, or creeping (rarely rosette-forming) | Ventral or restricted to base of stem | Tetragonal, rarely complanate | Monomorphic or few dimorphic (resupinate) | Four rows | Dimorphic (or monomorphic on stem) | Four rows | Several on dorsal or/and ventral sides |
Table 5.
Comparison of spore (megaspore and microspore) morphology, distribution, and constituent species of 7 subfamilies and 19 genera in the current classification of Selaginellaceae.
| Subfamilies/Genera | Megaspores | Microspores | Distribution | Constituent species |
|---|---|---|---|---|
| Subfamily I Selaginoidoideae [_Selaginoides_] | Baculate to verrucate | Spiny | Circumboreal areas and Hawaii | 2 species (Selaginoides deflexa; S. selaginoides) |
| Subfamily II Boreoselaginelloideae [_Boreoselaginella_] | With densely contiguous tubercles and verrucae | Rugulate, tuberculate, or verrucate | Eastern to western Asia, and extending to Russia. | 6 species (B. aitchisonii, B. borealis, B. jacquemontii, B. nummularifolia, B. rossii, B. sanguinolenta) |
| Subfamily III Gymnogynoideae | Gymnogynum type or Bryodesma type | Rugulate, spiny, or verrucate | Worldwide | All species with dorsal rhizophores excluding Boreoselaginella; all species with sterile leaves decussately arranged on the stem |
| Afroselaginella | Gymnogynum type | With an equatorial flange | Southern Africa | Except S. kraussiana, all African species with rhizophores born on dorsal side of stems and branches (Afroselaginella chevalieri, A. congoensis, A. myosurus, A. volubilis) |
| Bryodesma | Bryodesma type | Rugulate and verrucate | Americas, Africa and Asia | All species with dorsal rhizophores + spirally arranged monomorphic sterile leaves |
| Ericetorum | Gymnogynum type | Rugulate or verrucate | Australasia and Afro-Malagasy | All species with rhizophores restricted to the base of stems and sterile leaves decussately arranged on stem |
| Gymnogynum | Gymnogynum type | Spiny | Americas (a few in Africa and Asia) | All species in Americas with dorsal rhizophores + only one (to a few) megasporophyll at the base of strobili + African Gymnogynum kraussianum and Asian G. remotifolium |
| Lepidoselaginella | Gymnogynum type | Several curved ridges | Mexico and USA | 3 rosette-forming species (L. lepidophylla, L. novoleonensis, L. ribae). |
| Megaloselaginella | Gymnogynum type | Columned | Central and South Americas | 2 species (Megaloselaginella exaltata, M. gigantea) with extremely large plants |
| Subfamily IV Sinoselaginelloideae | Verrucate, tuberculate, lamellate | Rugulate, tuberculate, verrucate | Old World | Only one (to a few) megasporophyll at the base + rhizophores borne on ventral side of stems or branches |
| Austroselaginella | Verrucate, tuberculate | Coarse | Queensland in Australia | All species in Queensland of Australia with strobili with only one (to a few) megasporophyll at the base + rhizophores borne on ventral side of stems or branches |
| Korallia | Verrucate, tuberculate | Verrucate | Madagascar and adjacent Indian Ocean islands | All species in Madagascar and adjacent Indian Ocean islands with strobili with one (to a few) megasporophyll at the base + rhizophores borne on ventral side of stems or branches |
| Sinoselaginella | Verrucate, tuberculate | Rugulate, tuberculate, verrucate | China to the Arabic area and eastern Africa | All species in China (extending to the Arabic area and eastern Africa) with strobili with only one (to a few) megasporophyll at the base + rhizophores borne on ventral side of stems or branches |
| Subfamily V Pulvinielloideae [_Pulviniella_] | Coarse or small verrucate or granular | Rugulate, verrucate, coarse | Africa, Asia, and North and Central Americas | All rosette-forming species except Lepidoselaginella and two species of Selaginella (S. pallescens and S. nothohybrida); some species with extremely xerophytic habit and erect plants (e.g., Pulviniella helioclada, P. stauntoniana) |
| Subfamily VI Lycopodioidoideae | Tuberculate to verrucate | Tuberculate, verrucate or granular (rarely baculate) | Old World (a few in Americas) | All Old-World species with resupinate strobili and dimorphic to slightly dimorphic sporophylls; all species in the Old World with strictly monomorphic sporophylls + sterile leaves in four rows + non-reticulate megaspore surfaces; two American species: Didiclis hoffmannii and Valdespinoa douglasii |
| Didiclis | Tuberculate to verrucate, often interconnected; or smooth | Verrucate, blunt-spiny, cristate, and lamellate | Asia and Africa (a few in Americas) | All Old-World species with strictly monomorphic sporophylls + sterile leaves four rows + non-reticulate megaspore surfaces; species with dimorphic sporophylls in the Bisulcata group: Didiclis bisulcata, D. obovata, D. opaca, D. pennata, and D. soyauxii; and American D. hoffmannii) |
| Hypopterygiopsis | Verrucate, tuberculate, Tetragonostachyae type-1 or -2 | Granular, verrucate, smooth | Asia, Africa, and south Pacific islands | All species with resupinate strobili + dimorphic or slightly dimorphic sporophylls in the Old World except the Bisulcata group of Didiclis (D. bisulcata, D. obovata, D. opaca, D. pennata, and D. soyauxii) in South and Southeast Asia |
| Lycopodioides | Tuberculate and/or vermiculate or ridged | Verrucate | Europe to South and East Asia (only 1 sp. in Java, Indonesia) | All species with non-resupinate strobili + dimorphic sporophylls (but including Lycopodioides denticulata and L. helvetica) |
| Valdespinoa | Granular to verrucate | Surfaces smooth but fossulate | USA | Only Valdespinoa douglasii |
| Subfamily VII Selaginelloideae | Stachygynandrum type; rarely verrucate | Baculate, blunte spiny, some tubercare, verrucate, papillate, or coarse etc. | Asia, Americas, Pacific islands, a few in Africa | All species with rhizophores born on ventral side of stems and branches or restricted at the base of stem + many megasporophylls born on dorsal or ventral side of strobili + no rosette-forming habit (except S. pallescens, S. nothohybrida) + often reticulate megaspore surfaces + often baculate microspores surfaces |
| Chuselaginella | Stachygynandrum type | Baculate | Asia, Pacific islands, and a few in Africa | All Asian species with a zonal on the proxiaml surface of megaspores + microspores with baculate ornamentation, and African Chuselaginella versicolor, etc. |
| Kungiselaginella | Stachygynandrum type | Blunte spiny | Asia | All Asian species without a zonal on the proxiaml surface of megaspores + microspores with blunte or tubercate ornamentation |
| Selaginella s.s. | Stachygynandrum type or few tubercate on surfaces | Baculate, blunte spiny, verrucate, papillate, or coarse etc. | Americas and one (or a few) in Africa | All species with rhizophores born on ventral side of stems and branches or restricted at the base of stem + many megasporophylls born on dorsal or ventral side of strobili + no rosette-forming habit (except S. pallescens, S. nothohybrida), and the African S. cathedrifolia. |
In a very parallel case, Thelypteris Schmidel was reduced to a tiny (2 species) genus, with all other species being placed in “new” genera (Fawcett et al., 2021; Fawcett and Smith, 2021). The same could be done in Selaginella, but we prefer to conserve Selaginella with a new type to maximally stabilize the names (Wan et al., 2023). Thelypteris had been to a much greater degree already treated and split into many genera, whereas, despite some movement over the centuries towards splitting Selaginella, much less had been done and segregates had been much less discussed, used, combinations made, in Selaginella until the modern era of gene sequencing and phylogenetic trees.
Treating these 19 genera at the rank of subgenus or section within Selaginella is an alternative (Jermy, 1986, 1990; Zhou and Zhang, 2015; Weststrand and Korall, 2016b), but those ranks are almost always ignored except by professional plant systematists, and there is nothing like the name of a species (incorporating the genus) to emphasize less relationship (different genus names) or more relationship (same genus name; A. Weakley, pers. comm.).
It will be more inconvenient for today's botanists to adopt a dramatically different classification than to use the existing one with all 750+ species together in one genus, but in the long run the next-generation botanists will find it informative to distinguish the deeply diverged clades at the generic rank because of their molecular, morphological, and geographical distinctiveness. Treating all 750+ species in a single genus is comparable to lumping all then 33 genera of Polypodiaceae subfam. Grammitidodeae into Grammitis Sw. or lumping all 24 genera of cheilanthoid ferns into Hemionitis L. (Pteridaceae) as done by Christenhusz et al. (2018).
5. Taxonomic treatment
Our current phylogeny based on the largest taxon and character (nuclear and plastid) sampling corroborated earlier findings (Weststrand and Korall, 2016a; Zhou et al., 2016) that Selaginella is well resolved into a number of major clades. Our morphological analysis of 10 important characters showed that the major phylogenetic clades can be defined morphologically (Weststrand and Korall, 2016a; our Fig. 2). Our earlier classification (Zhou and Zhang, 2015) recognized six subgenera and three of them further into five, six, and seven sections, respectively, excluding the Sinensis group. Our new results confirm the monophyly of all but two sections (the relationships between S. sect. Circinatae and S. sect. Plagiophyllae are not well resolved but they together are monophyletic). Considering all evidence available, earlier classifications of Selaginella (Mirbel (in Lamark and Mirbel, 1803); Kuntze, 1891; Walton and Alston, 1938; Sakurai, 1943; Rothmaler, 1944; Jermy, 1986, 1990; Kung, 1988; Soják, 1992; Satou, 1997; Tzvelev, 2004; Weakley, 2012, 2022; Zhou and Zhang, 2015; Weststrand and Korall, 2016b), and recent trend of pteridophyte classifications, we here propose to split the currently defined Selaginella s.l. into seven subfamilies corresponding our earlier six subgenera (Zhou and Zhang, 2015) and the Sinensis group, three of which are monogeneric and four of which are further divided into 3, 3, 4, and 6 genera, respectively. The 7 subfamilies and 19 genera include: Boreoselaginelloideae (Boreoselaginella), Gymnogynoideae (Afroselaginella, Bryodesma, Ericetorum, Gymnogynum, Lepidoselaginella, Megaloselaginella), Lycopodioidoideae (Didiclis, Hypopterygiopsis, Lycopodioides, Valdespinoa), Pulvinielloideae (Pulviniella), Selaginelloideae (Chuselaginella, Kungiselaginella, Selaginoides s.s.), Selaginoidoideae (Selaginoides), and Sinoselaginelloideae (Austroselaginella, Korallia, Sinoselaginella). A comparison of our current and earlier classifications is listed in Table 3 and Figs. 1 and S1–S3.
We provide a key to subfamilies and genera below. For each subfamily and genus, we provide morphological, phylogenetic, and geographical synopses, constituent species, and the necessary new combinations. For these, we provide basionyms and other recent combinations. For additional synonymy see Tryon (1955), Kung (1988), Soják (1992), Valdespino (1993), Jermy and Holmes (1998), Mickel and Smith (2004), Chu (2006), Roux (2009), Zhang et al. (2013), Fraser-Jenkins et al. (2015, 2017), Valdespino and Zimmer (2016), and Hassler (2022). We list all species of each genus currently known to us. Species lacking sufficient evidence are listed at the end of “Taxonomic treatment” without combinations pending further studies. We provide Chinese vernacular names for all subfamilies, genera, and sections in the family and for all species occurring in China.
5.1. Key to subfamilies and genera of Selaginellaceae
- Rhizophore absent; sterile leaves loosely spirally arranged; strobili cylindric (circumboreal areas and Hawaii; 2n = 18) [subfam. I. Selaginoidoideae 圆穗卷柏亚科] .......................... ...................................................................... Selaginoides 圆穗卷柏属
- Rhizophore present; sterile leaves densely and spirally, in four rows, or decussately arranged; strobili tetragonal or dorsiventrally complanate (2n = 16–60) .................................. 2
- Rhizophores borne on dorsal side of stems and/or branches; if rhizophores strictly restricted to the base of stem, sterile leaves at least decussately arranged on stem or throughout plants ...................................................................................................... 3
- Rhizophores borne on ventral side of stems and/or branches; if rhizophores strictly restricted to the base of stem, sterile leaves on stem never decussately arranged .............................. 9
- Plants xerophytic, non-rosette-forming, creeping; sterile leaves in 4 rows, strobili with many megasporangia on dorsal and/or ventral side; megaspore surfaces densely contiguous-tuberculate, non-reticulate [subfam. II. Boreoselaginelloideae 北方卷柏亚科].................... Boreoselaginella 北方卷柏属
- Plants non-xerophytic (if xerophytic, plants rosette-forming or with spirally monomorphic leaves on sterile stem and branch), erect, suberect, ascending, scandent, or creeping (if creeping, plants with monomorphic leaves throughout the plant or strobili with only one megasporangium at base); megaspore surfaces reticulate (the Gymnogynum type and the Bryodesma type) [subfam. III. Gymnogynoideae 关节卷柏亚科] ................ 4
- Sterile leaves strictly monomorphic throughout the plant or at least on stem .................................................................................... 5
- Sterile leaves dimorphic or slightly monomorphic throughout the plant ......................................................................... 6
- Sterile leaves monomorphic and spirally arranged throughout the plant .............................. Bryodesma 同形卷柏属
- Sterile leaves monomorphic and decussately arranged throughout the plant or at least at the base of stems ................. ......................................................................... Ericetorum 对叶卷柏属
- Plants rosette-forming ............. Lepidoselaginella 鳞叶卷柏属
- Plants non-rosette-forming ............................................................ 7
- Stems inarticulate; microspores with an equatorial flange and surfaces verrucate .................. Afroselaginella 非洲卷柏属
- Stems often articulate; microspores without an equatorial flange and surfaces echinate or pillared ..................................... 8
- Plants extremely large (up to 1 m tall); stem’s vascular system actino-plectostele; megaspores ca. 1.5 mm; microspore surfaces pillared ........................ Megaloselaginella 大孢卷柏属
- Plants normally shorter than 80 cm; stem's vascular system haplostele, actinostele, or plectostele; megaspores smaller than 1 mm; microspore surfaces often echinate........................... .................................................................... **Gymnogynum**关节卷柏属
- Plants xerophytic; often forming rosettes, rarely stems erect, rhizophores strictly restricted to the base of stem and stems tufted and curling inward when dry; megaspore surfaces coarse without clear ornamentation; microspores spherical, surface scabrate or verrucate [subfam. V. Pulvinielloideae垫状卷柏亚科] .................................................. _Pulviniella_垫状卷柏属
- Plants growing in wet habitats or rarely slightly xerophytic; stems erect, suberect, ascending, creeping, or scandent, rarely rosette-forming but megaspore surfaces reticulate, and microspores often hemispherical ....................................... 10
- Sporophylls monomorphic [if dimorphic, then megaspore surfaces with typical reticulate ornamentation (only in Americas)]; megaspore surfaces reticulate (the Stachygynandrum type) [if non-reticulate, megaspores larger than 500 μm; Sinoselaginelloideae]; microspore surfaces often verrucate to baculate on distal surfaces (Old and New World) ....................... 16
- Sporophylls dimorphic [if monomorphic, megaspore surfaces verrucate, low-rugate, tuberculate, finely reticulate (or fenestrate)], megaspore surfaces often non-reticulate [if reticulate, megaspores are of the Tetragonostachyae type-1 & -2]; microspore surfaces verrucate, blunt spiny, lamellate, cristate, plain but with globules or spherules, or rarely baculate on distal surfaces (Old World, only Didiclis hoffmannii in the New World] [subfam. VI. Lycopodioidoideae 异穗卷柏亚科] .......................................................................................... 11
- Sporophylls dimorphic, non-resupinate or resupinate (if monomorphic, strobili strongly loose and not distinct from sterile parts) ........................................................................................ 12
- Sporophylls monomorphic or submonomorphic, and strobili distinct from sterile parts ................................................................ 14
- Strobili non-resupinate (sporophylls dimorphic) or rarely slightly lax (monomorphic); sporophylls loosely arranged ................................................................... Lycopodioides 疏穗卷柏属
- Strobili resupinate (sporophylls dimorphic); sporophylls densely arranged .............................................................................. 13
- Dorsal leaves obovate ............................................................................ ...................................Didiclis (the D. bisulcata group) 瘤孢卷柏属
- Dorsal leaves non-obovate ........ Hypopterygiopsis 异穗卷柏属
- Plants small, ca. 10 cm, strictly creeping forming loose mats ...................................................................... Valdespinoa 韦氏卷柏属
- Plants medium-to large-sized, more than 20 cm, erect, scandent, rarely creeping, not forming loose mats ................ 15
- Megaspore surfaces tuberculate and usually interconnected to form reticula-like structure; microspore surfaces blunt spiny, laminate, or cristate ............................ Didiclis 瘤孢卷柏属
- Megaspore surfaces finely reticulate (Tetragonostachyae type-1 and -2); microspore surfaces low-verrucate or smooth with irregular spherules ............ Hypopterygiopsis 异穗卷柏属
- Strobili with several megasporophylls (sporangia) on the ventral and/or dorsal side; megaspores smaller than 400 μm, surfaces reticulate (the Stachygynandrum type) [subfamily VII. Selaginelloideae 同穗卷柏亚科]...................... 17
- Strobili with only one (to a few) megasporophyll (sporangium) at the base; megaspores larger than 500 μm, surfaces non-reticulate [subfamily IV. Sinoselaginelloideae 中华卷柏亚科] ....................................................................................................... 19
- Microspore surfaces coarse, baculate, blunt spiny, papillate, or verrucate, rarely ridged (Americas, except S. cathedrifolia in Africa) ............................................... Selaginella s.s. 同穗卷柏属
- Microspore surfaces often baculate or with blunt spines (Old World) ................................................................................................... 18
- Plants often ascending to suberect, a few strictly erect; sides of veins of ventral leaves with two light-colored bands; megaspores without a zonal structure on proximal surface; microspore surfaces blunt spiny (Asia) .............................................................. Kungiselaginella 孔氏卷柏属
- Plants often strictly erect, a few creeping; sides of veins of ventral leaves without light-colored bands; megaspores with a zonal structure on proximal surface; microspore surfaces baculate with extending tips (Asia, Pacific islands to Africa) .................................................................. Chuselaginella 朱氏卷柏属
- Plants xerophytic; microspore surfaces prominently tuberculate (East and South Asia extending to Arabic area and northern Africa)................................. Sinoselaginella 中华卷柏属
- Plants non-xerophytic; microspore surfaces verrucate or coarse (Australia, Madagascar and adjacent islands) .......... 20
- Ventral leaves usually auriculate at basiscopic and acroscopic bases; microspores with three holes on proximal surfaces (Madagascar and adjacent islands) .......................................................................... Korallia 科氏卷柏属
- Ventral leaves non-auriculate at base; microspore surfaces without holes on proximal surfaces (Australia) ............................................................ Austroselaginella 澳洲卷柏属
5.2. Subfamily I. Selaginoidoideae
Subfamily I. Selaginoidoideae Li Bing Zhang & X.M. Zhou, subfam. nov. 圆穗卷柏亚科(新拟) – Type: Selaginoides Ség.
Plants erect, with no rhizophores; sterile leaves monomorphic and spirally arranged; sporophylls monomorphic; megaspore surfaces blunt echinate to tuberculate; microspore surfaces with echinate (Fig. 3); 2n = 18 (Takamiya, 1993).
This subfamily corresponds to Selaginella subg. Selaginella sensu Zhou and Zhang (2015) and Weststrand and Korall (2016b).
It contains one genus: Selaginoides, occurring in circumboreal areas and Hawaii.
5.2.1. Selaginoides
Selaginoides Ség., Pl. Veron. 3: 51. 1754. 圆穗卷柏属(新拟) ≡ Mirmau Adans., Fam. Pl. (Adanson) 2: 491. (1763) ≡ Polycocca Hill, Gener. Nat. Hist., ed. 2, 2(Hist. Pl.): 116 (1773) – Type: Lycopodium selaginoides L. ≡ Selaginella selaginoides (L.). P. Beauv. ex Schrank & Mart., Hort. Reg. Monac. 3 (1829) = Selaginoides spinulosa (A. Braun ex Döll) Li Bing Zhang & X.M. Zhou ≡ Selaginella spinulosa A. Braun ex Döll, Rhein. Fl.: 38 (1843) ≡ Lycopodina spinulosa (A. Braun ex Döll) Bubani, Fl. Pyren. 4: 445 (1901) = Selaginella subg. Homoeophyllum Hieron. & Sadeb., Engler & Prantl, Nat. Pflanzenfam. l(4): 669. 1902 – Type: Selaginella selaginoides (L.) P. Beauv. ex Mart. & Schrank_._
Selaginoides is resurrected here. Selaginella has been proposed to be conserved with a conserved type, Selaginella flabellata (L.) Spring (Wan et al., 2023; also see below). Selaginella spinosa P. Beauv. (Prodr. Aethéogam.: 112. 1805) is an illegitimate name – the epithet ‘_selaginoides_’ (from Lycopodium selaginoides L., Sp. Pl.: 1101. 1753) was available for use by Palisot de Beauvois in Selaginella. The epithet ‘_selaginoides_’ from L. selaginoides would create a tautonym in Selaginoides (Art. 23.4 of the ICN). The earliest available epithet in Selaginoides is ‘_spinulosa_’.
Selaginoides is the earliest diverging lineage in Selaginellaceae (Korall et al., 1999; Korall and Kenrick, 2004; Zhou et al., 2016, 2022; Weststrand and Korall, 2016a).
Morphologically, this genus differs from the rest of species in the family in having no rhizophores and monomorphic and spirally arranged leaves (Fig. 2A, E).
The genus contains two species: Selaginoides spinulosa (circumboreal area) and S. deflexa (Hawaii) (Fig. 3).
Members:
Selaginoides deflexa (Brack.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella deflexa Brack., U.S_._ Expl. Exped., Filic. 16: 332, no. 3, t.45, f.3 (1854) ≡ Lycopodioides deflexa (Brack.) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Selaginoides spinulosa (A. Braun ex Döll) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella spinulosa A. Braun ex Döll, Rhein. Fl.: 38 (1843) ≡ L. spinulosa (A. Braun ex Döll) Bubani, Fl. Pyren. 4: 445 (1901).
5.3. Subfamily II. Boreoselaginelloideae
Subfamily II. Boreoselaginelloideae Li Bing Zhang & X.M. Zhou, subfam. nov. 北方卷柏亚科(新拟) – Type: Boreoselaginella (Warb.) Li Bing Zhang & X.M. Zhou.
Plants xerophytic, creeping (Fig. 4A–C); sterile leaves in four rows, nearly monomorphic (Fig. 4D) or dimorphic (Fig. 4H), axillary leaves present; rhizophores borne on dorsal side of stem (Fig. 4D); sporophylls monomorphic; megaspore surfaces with dense contiguous tubercles; microspore surfaces rugate to verrucate; 2n = 30 (Takamiya, 1993).
This subfamily corresponds to Selaginella subg. Boreoselaginella sensu Zhou and Zhang (2015). It is likely the second earlier diverging lineage in the family (Zhou et al., 2016; our Figs. 1 and S3).
Plastome conformation of Boreoselaginelloideae known so far (Zhou et al., 2022) was DR structure with only one conformation. In addition, these genes (psaM/rpl20/rpl21/rpl33) tend to be lost or pseudogenized, but are preserved in other taxa of Selaginellaceae. Furthermore, genes (psaM/trnI-CAU/trnP-GGG/trnS-GCU/trnS-UGA) only existed in Boreoselaginelloideae. Except ndhE, all ndh genes were lost or pseudogenized in Boreoselaginelloideae (Zhang et al., 2022; Zhou et al., 2022).
It contains one genus: Boreoselaginella, occurring from eastern to western Asia, and extending to Russia.
5.3.1. Boreoselaginella
Boreoselaginella (Warb.) Li Bing Zhang & X.M. Zhou, stat. nov. 北方卷柏属(新拟) – Basionym: Selaginella subg. Boreoselaginella Warb., Monsunia 1: 100. 1900 – Lectotype (designated by Zhou and Zhang, 2015: 1129): Boreoselaginella borealis (Kaulf.) Li Bing Zhang & X.M. Zhou ≡ Selaginella borealis (Kaulf.) Spring (= Selaginella sanguinolenta (L.) Spring).
Boreoselaginella has rhizophores borne on the dorsal side of the stem, monomorphic and spirally arranged sterile leaves throughout, and non-reticulate megaspores_._ This combination of the morphology is unique in the family.
It contains about six species distributed from eastern to western Asia and extending to Russia.
Members:
Boreoselaginella aitchisonii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella aitchisonii Hieron., Engl. & Prantl, Nat. Pfl. 1 (4) 674 (1902) ≡ Lycopodioides aitchisonii (Hieron.) Tzvelev, Novosti Sist. Vyssh. Rast. 36: 25 (2004) ≡ Selaginella sanguinolenta f. aitchisonii (Hieron.) Alston, Proc. Nat. Inst. Sci. India 11: 215 (1945).
Boreoselaginella borealis (Kaulf.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium boreale Kaulf., Enum. Fil. 17–18 (1842) ≡ Selaginella borealis (Kaulf.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10 (1): 141, no. 40 (1843) ≡ Lycopodioides borealis (Kaulf.) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 北方卷柏(新拟)
Boreoselaginella jacquemontii (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella jacquemontii Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 226, no. 104 (1843).
Boreoselaginella nummularifolia (Ching) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella nummularifolia Ching, Fl. Xizangica 1: 21 (1983). 钱叶北方卷柏(新拟)
Boreoselaginella rossii (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella mongholica var. rossii Baker, J. Bot. 21: 45 (1883) ≡ Selaginella rossii (Baker) Warb., Monsunia 1: 101 (1900) ≡ Lycopodioides rossii (Baker) Tzvelev, Novosti Sist. Vyssh. Rast. 36: 25 (2004). 鹿角北方卷柏(新拟)
Boreoselaginella sanguinolenta (L.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium sanguinolentum L., Sp. Pl. 2: 1100 (1753) ≡ Stachygynandrum sanguinolentum (L.) P. Beauv., Prodr. aethéogam. 114 (1805) ≡ Selaginella sanguinolenta (L.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 135, no. 2 (1843) ≡ Lycopodioides sanguinolenta (L.) Kuntze, Rev. Gen. Pl. 1: 827 (1891). 红枝北方卷柏(新拟)
5.4. Subfamily III. Gymnogynoideae
Subfamily III. Gymnogynoideae Li Bing Zhang & X.M. Zhou, subfam. nov. 关节卷柏亚科 (新拟) – Type: Gymnogynum P. Beauv.
Plants erect, ascending, or creeping, rarely scandent or rosette-forming (Fig. 5A–E), with rhizophores borne on dorsal side of stems and/or branches (Fig. 5F, H–L); a few species with rhizophores strictly restricted to the base of stem (Fig. 5C); sterile leaves monomorphic and decussately arranged at least in lower portion of plants (Ericetorum) (Fig. 5C, K); sporophylls monomorphic, with only one megasporophyll at the base of strobili (Fig. 5J); reticulate megaspore surfaces with wing-like and highly convoluted laesurae forming a complex mass close to the pole (Ericetorum) (Fig. 5P); megaspore surfaces reticulate (mainly Gymnogynum type with high or wing-like muri and often closed meshes) (Fig. 5N–R); microspore surfaces verrucate to rugate or echinate (Fig. 5S–W; Zhou et al., 2015); 2n = 18, 20 (Jermy et al., 1967; Takamiya, 1993; Marcon, 2003).
The plastome conformations of Gymnogynoideae known so far (Zhou et al., 2022) have all four types of structures (IR, DR, IR-DR coexisting, and NR). In addition, genes ndh and matK tend to lose or pseudogenize except in Gymnogynum. The rpl14 (except pseudogenetization in Lepidoselaginella) and _ycf_3 intron2 (except preserved in Megaloselaginella) have been lost in all genera.
This subfamily corresponds to “Selaginella subg. _Ericetorum_” (= S. subg. Gymnogynum) sensu Zhou and Zhang (2015). Weststrand and Korall (2016b) recognized five subgenera in this clade.
It contains about 130 species in six genera, Afroselaginella, Bryodesma, Ericetorum, Gymnogynum, Lepidoselaginella, and Megaloselaginella, occurring in Africa, Australasia, the Caribbeans, and Americas.
5.4.1. Afroselaginella
Afroselaginella Li Bing Zhang & X.M. Zhou, gen. nov. 非洲卷柏属(新拟) – Type: Afroselaginella myosurus (Sw.) Li Bing Zhang & X.M. Zhou [Lycopodium myosurus Sw.; Selaginella myosurus (Sw.) Alston].
= Selaginella sect. Myosurus Li Bing Zhang & X.M. Zhou, Taxon 64 (6): 1133 (2015) – Type: Selaginella myosurus (Sw.) Alston.
Etymology: “_Afro-_” derived from Africa, referring to the African distribution of the genus.
Plants creeping, stems inarticulate, with a single large megasporangium per strobilus, reticulate megaspores with extremely wide and high muri (Fig. 5N), microspores with an equatorial ring (Fig. 5S).
Afroselaginella has the smallest plastome size (ca. 100 kb) in Gymnogynoideae known so far (Zhou et al., 2022). Plastome structure of Afroselaginella has only SC region (single copy region) but lacking repeat region and has one ribosomal operon (Zhou et al., 2022).
Afroselaginella circumscribed here corresponds to Selaginella sect. Myosurus sensu Zhou and Zhang (2015) and part of S. subg. Exaltatae sensu Weststrand and Korall (2016b).
It contains about four species in Africa.
Members:
Afroselaginella chevalieri (Hieron. ex Bonap.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella chevalieri Hieron. ex Bonap., Notes pterid. 1: 124 (1915).
Afroselaginella congoensis (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella congoensis Alston, Mém. Inst. Fr. Afr. Noire 50: 30, t. 6, f. 1–8 (1957).
**Afroselaginella myosurus (**Sw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium myosurus Sw., Schrad., J. Bot. (1800) (2): 118 (1801) ≡ Selaginella myosurus (Sw.) Alston, J. Bot. 70: 64, no. 6 (1932). J. Bot. 70: 64, no. 6 (1932).
Afroselaginella volubilis (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella volubilis Alston, Bol. Soc. Broter. Ser. 2, 30: 25 (1956).
5.4.2. Bryodesma
Bryodesma Soják, Preslia 64(2): 154 (1993) 同形卷柏属(新拟) – Type: Bryodesma rupestre (L.) Soják, Preslia 64(2): 155 (1993) ≡ Lycopodium rupestre L., Sp. Pl. 2: 1101 (1753) ≡ Selaginella rupestris (L.) Spring, Flora 21(1): 182. 1838.
= Selaginella sect. Homoeophyllae Spring in Martius, Fl. Bras_._ 1(2): 118 (1840) ≡ Selaginella sect. Homotropae A. Broun, App. Ind. Sem. Hort. Berol. 11. ([1857] 1858) – Type: Selaginella rupestris (L.) Spring.
= Selaginella subg. Tetragonostachys Jermy, Fern Gaz. 13: 118. 1986, non Hook. & Grev. in Bot. Misc. 2: 382 (1831) – Type: Selaginella rupestris (L.) Spring.
Bryodesma corresponds to Selaginella sect. Homoeophyllae sensu Zhou and Zhang (2015) and to S. subg. Rupestrae sensu Weststrand and Korall (2016b), and approximates “S. subg. _Tetragonostachys_” sensu Jermy (1986). It also approximates S. sect. Tetragonostachys sensu Tryon (1955) who recognized four series (S. ser. Arenicolae Tryon, S. ser. Sartorii Tryon, S. ser. Rupestres Tryon, and S. ser. Eremophilae Tryon) based on the habit and leaf morphology. The relationships of the four series in our phylogeny are unresolved (Fig. S3). Two plastid genomes of species from this genus [Selaginella vardei H. Lév. and S. indica (Milde) R.M. Tryon] were studied and extremely rare short dispersed repeats have been found (Zhang et al., 2019).
Morphologically, Bryodesma can be easily distinguished in having xerophytic habit (Fig. 5A), rhizophores borne on dorsal side of the stem (Fig. 5H), sterile leaves spirally arranged throughout (Fig. 5A–H), and axillary leaves absent (Fig. 5H). The proximal surfaces of microspores always present rough and irregularly rugate ornamentation (Fig. 5V), which is also found in these xerophytic species of S. subg. Boreoselaginella (Zhou et al., 2015).
Plastome structures of nearly all species in Bryodesma known so far (Zhou et al., 2022) are DR (direct repeats) structure and without small or medium repeats existed in SC (single-copy region).
Bryodesma contains about 60 species, about 40 species in Americas (nearly all in North America), about 12 species in Africa and Madagascar and adjacent islands, and about five species in Asia (Tryon and Lugardon, 1991; Valdespino, 1993a; Roux, 2009; Weakley, 2012, 2022; Arrigo et al., 2013; Fraser-Jenkins et al., 2015, 2017; Zhou and Zhang, 2015; Weststrand and Korall, 2016b; Hassler, 2022). Bryodesma corallinum (Riddell) Weakley has recently been transferred to the genus (Weakley, 2022).
Members:
Bryodesma acanthonota (Underw.) Skoda, Preslia 68(4): 343 (1997). Basionym: Selaginella acanthonota Underw., Torreya 2: 172 (1902).
Bryodesma aethiopicum (Bizzarri) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella caffrorum var. aethiopica Bizzarri, Webbia 29: 556 (1975).
Bryodesma arenaria (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella arenaria Baker, J. Bot. 21: 82, no. 23 (1883).
Bryodesma arenicola (Underw.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella arenicola Underw., Bull. Torrey Bot. Club 25: 541 (1898).
Bryodesma arizonicum (Maxon) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella arizonica Maxon, Smithsonian Misc. Collect. 72: 5–6 (1920).
Bryodesma arsenei (Weath.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella arsenei Weath., J. Arnold Arbor. 25: 417, t. 2, f. 8 (1944).
Bryodesma asprellum (Maxon) Soják; Preslia 64(2): 154 (1992 publ. 1993) Basionym: Selaginella asprella Maxon, Smithsonian Misc. Collect. 72: 6 (1920).
Bryodesma balansae (A. Braun) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella balansae (A. Braun) Hieron., Hedwigia 39: 318 (1900).
Bryodesma bigelovii (Underw.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella bigelovii Underw., Bull. Torrey Bot. Club 25: 130, no. 6 (1898).
Bryodesma caffrorum (Milde) Soják; Preslia 64(2): 15,4 (1992 publ. 1993). Basionym: Selaginella caffrorum (Milde) Hieron., Hedwigia 39: 313 (1900).
Bryodesma basipilosum (Valdespino) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella basipilosa Valdespino, Brittonia 44(3): 314(1992 publ. 1993).
Bryodesma carinatum (R.M. Tryon) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella carinata R.M. Tryon, Ann. Mo. Bot. Gard. 42: 50, f. 25–26, map 31 (1955).
Bryodesma carnerosanum (T. Reeves) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella carnerosana T. Reeves, Brittonia 32(3): 365 (1980).
Bryodesma cinerascens (A.A. Eaton) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella cinerascens A.A. Eaton; Fern Bull. 7: 33 (1899).
Bryodesma corallinum (Riddell) Weakley; J. Bot. Res. Inst. Texas 16(2): 405 (2022). Basionym: Lycopodium corallinum Riddell; New Orleans Med. Surg. J. 9: 617 (1853).
Bryodesma densum (Rydb.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella densa Rydb.; Mem. New York Bot. Gard. 1: 7 (1900).
Bryodesma dregei (C. Presl) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Lycopodium dregei C. Presl; Abh. Königl. Böhm. Ges. Wiss., Math.-Naturw. Cl. V, 3 (1845), reimpr. in Bot. Bemerk. (C. Presl) 153 (1846).
Bryodesma echinatum (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella echinata Baker, J. Linn. Soc. Bot. 22: 536 (1887).
Bryodesma emodi (Fraser-Jenk.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella emodi Fraser-Jenk., in Fraser-Jenkins, Kandel & Pariyar, Ferns Fern-Allies Nepal 1: 67–68 (2015). 印度同形卷柏(新拟)
Bryodesma eremophilum (Maxon) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella eremophila Maxon; Smithsonian Misc. Collect. 72(5): 3, t. 2 (1920).
Bryodesma extensum (Underw.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella extensa Underw.; Bull. Torrey Bot. Club 25: 131 (1898).
Bryodesma griseum (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella grisea Alston, J. Bot. 77: 222 (1939).
Bryodesma hansenii (Hieron.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella hansenii Hieron., Hedwigia 39: 301 (1900).
Bryodesma landii (Greenm. & N. Pfeiff.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella landii Greenm. & N. Pfeiff., Ann. Mo. Bot. Gard. 5: 205 (1918).
Bryodesma macratherum (Weath.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella macrathera Weath., J. Arnold Arbor. 24: 326 (1943).
Bryodesma niveum (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella nivea Alston, Dansk Bot. Ark. 7: 194, t. 78(1–4) (1932), nom. nud., et Cat. Pl. Madagasc., Pterid. 71 (1932).
Bryodesma humbertii (Stefanov. & Rakotondr.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella nivea subsp. humbertii Stefanov. & Rakotondr., Novon 6(2): 207 (1996).
Bryodesma leucobryoides (Maxon) Skoda; Preslia 68(4): 343 (1997). Basionym: Selaginella leucobryoides Maxon, Smithsonian Misc. Collect. 72(5): 8, t. 5 (1920).
Bryodesma limitaneum (Weath.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella mutica var. limitanea Weath., J. Arnold Arbor. 25: 414 (1944) ≡ Bryodesma muticum var. limitaneum (Weath.) Skoda, Preslia 68(4): 343 (1997).
Bryodesma muticum (D.C. Eaton ex Underw.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella mutica D.C. Eaton ex Underw., Bull. Torrey Bot. Club 25: 128 (1898).
Bryodesma ×neomexicanum (Maxon) Skoda; Preslia 68(4): 343 (1997). Basionym: Selaginella ×neomexicana Maxon, Smithsonian Misc. Collect. 72: 2 (1920).
Bryodesma njamnjamense (Hieron.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella njamnjamensis Hieron., Hedwigia 39: 312 (1900).
Bryodesma oreganum (D.C. Eaton) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella oregana D.C. Eaton; Wats., Bot. California 2: 350 (1880).
Bryodesma parishii (Underw.) Soják; Preslia 64(2): 154 (1992 publ. 1993). Basionym: Selaginella parishii Underw., Bull. Torrey Bot. Club 33: 202 (1906).
Bryodesma peruvianum (Milde) Soják; Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella peruviana (Milde) Hieron., Hedwigia 39: 307 (1900).
**Bryodesma phillipsianum (**Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella wightii var. phillipsiana Hieron., Hedwigia 39: 320 (1900) ≡ Selaginella phillipsiana (Hieron.) Alston, J. Bot. 77: 222 (1939).
Bryodesma proximum (R.M. Tryon) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella proxima R.M. Tryon, Ann. Mo. Bot. Gard. 42: 56, f. 33–35, map 37 (1955).
Bryodesma rupestre (L.) Soják; Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella rupestris (L.) Spring; Flora 21: 149 and 182, no. 12 (1838).
Bryodesma rupincola (Underw.) Soják; Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella rupincola Underw.; Bull. Torrey Bot. Club 25: 129 (1898).
Bryodesma sartorii (Hieron.) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella sartorii Hieron.; Hedwigia 39: 304 (1900).
Bryodesma scopulorum (Maxon) Skoda & Holub, Preslia 68(4): 343 (1997). Basionym: Selaginella scopulorum Maxon; Am. Fern J. 11: 36 (1921).
Bryodesma sellowii (Hieron.) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella sellowii Hieron., Hedwigia 39: 306 (1900).
Bryodesma shabaense (Bizzarri) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella shabaensis Bizzarri, Webbia 36(1): 204 (1982).
Bryodesma shakotanense (Franch. ex Takeda) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella shakotanensis (Franch. ex Takeda) Miyabe & Kudô; Fl. Hokkaido 63 (1930). 北海道同形卷柏(新拟)
Bryodesma sibiricum (Milde) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella sibirica (Milde) Hieron.; Hedwigia 39: 290, no. 1 (1900). 西伯利亚同形卷柏(新拟)
Bryodesma standleyi (Maxon) Skoda; Preslia 68(4): 343 (1997). Basionym: Selaginella standleyi Maxon; Smithsonian Misc. Collect. 72(5): 9 (1920).
Bryodesma steyermarkii (Alston) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella steyermarkii Alston, Ann. Mag. Nat. Hist. XII, 7: 638, t. XII.9, f. a–c. (1954).
Bryodesma tortipilum (A. Braun) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella tortipila A. Braun, Ann. Sci. Nat., Bot., sér. V, 3: 271 (1865).
Bryodesma underwoodii (Hieron.) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella underwoodii Hieron. in Engl. & Prantl, Nat. Pflanzenfam. 1: 714 (1901).
Bryodesma utahense (Flowers) Skoda & Holub, Preslia 68(4): 343 (1997). Basionym: Selaginella utahensis Flowers, Am. Fern J. 39: 83 (1949).
Bryodesma vardei (H. Lév.) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella vardei H. Lév., Cat. Pl. Yun-Nan 172, f. 41 (1915)-17. 细瘦同形卷柏(新拟)
Bryodesma viridissimum (Weath.) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella viridissima Weath., J. Arnold Arbor. 24: 326 (1943).
Bryodesma wallacei (Hieron.) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella wallacei Hieron., Hedwigia 39: 297 (1900).
Bryodesma watsonii (Underw.) Soják; Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella watsonii Underw., Bull. Torrey Bot. Club 25: 127 (1898).
Bryodesma weatherbianum (R.M. Tryon) Soják, Preslia 64(2): 155 (1992 publ. 1993). Basionym: Selaginella weatherbiana R.M. Tryon, Am. Fern J. 40: 69 (1950).
Bryodesma wightii (Hieron.) Soják, Preslia 64(2): 155 (1992). Basionym: Selaginella wightii Hieron., Hedwigia 39: 319 (1900).
Bryodesma wrightii (Hieron.) Soják; Preslia 64(2): 155 (1992). Basionym: Selaginella wrightii Hieron, Hedwigia 39: 298–299 (1900).
5.4.3. Ericetorum
Ericetorum (Jermy) Li Bing Zhang & X.M. Zhou, stat. nov. 对叶卷柏属(新拟) – Basionym: Selaginella subg. Ericetorum Jermy, Fern Gaz. 13(2): 117. 1986 – Type: Ericetorum uliginosa (Labill.) Li Bing Zhang & X.M. Zhou (≡ Selaginella uliginosa (Labill.) Spring).
= Didiclis sect. Lyallia Rothm., Feddes Repert. Spec. Nov. Regni Veg. 54: 70 (1944) ≡ Selaginella sect. Lyallia (Rothm.) Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1133 (2015) – Type: Selaginella lyallii (Hook. & Grev.) Spring.
= Selaginella (unranked) Tetrastichae A. Braun, App. Ind. Sem. Hort. Berol. 11. [1857] 1858 – Lectotype (designated by Zhou and Zhang 2015: 1133): Selaginella uliginosa (Labill.) Spring.
= Selaginella subser. Pleiostelicae Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. L(4): 707. 1902, non Hieronymus and Sadebeck, 1901 (1902: 700, 710) – Lectotype (designated by Zhou and Zhang 2015: 1133): Selaginella lyallii (Hook. & Grev.) Spring.
Ericetorum circumscribed here corresponds to Selaginella sect. Lyallia (Rothm.) Li Bing Zhang & X.M. Zhou (2015) and to S. subg. Ericetorum sensu Weststrand and Korall (2016b). Its geographical distribution and constituent species are same as those reported in previous studies (Zhou and Zhang, 2015; Weststrand and Korall, 2016b).
Morphologically, Ericetorum can be easily identified in having erect plant, solenostelic rhizome (Weststrand and Korall, 2016a), rhizophores strictly restricted to the base of stem (Fig. 5C), sterile leaves monomorphic and decussately arranged (or at least on stem) (Fig. 5K; Stefanović et al., 1997; Schulz et al., 2013). Megaspore surfaces are reticulate and have convoluted laesurae forming a complex mass close to the pole on proximal surfaces (Fig. 5P; Stefanović et al., 1997; Schulz et al., 2013). Microspore surfaces are gemmate, foveolate, verrucate to rugate ornamentation or scabrate often covered with microstructure being spines or gemmae (Fig. 5U; Stefanović et al., 1997; Schulz et al., 2013).
The plastome of Ericetorum known so far (Zhou et al., 2022) has DR structure, the lowest GC content (ca. 50%), and the smallest LSC (ca. 45 kb) in Gymnogynoideae.
Eight species are currently known in this genus containing four Australasian and four Afro-Malagasy species (Stefanović et al., 1997; Roux, 2009; Schulz et al., 2013).
Members:
Ericetorum aboriginale (C. Schulz & Homberg) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella aboriginalis C. Schulz & Homberg, Syst. Bot. 38(1): 11 (2013).
Ericetorum gracillimum (Kunze) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium gracillimum Kunze, Lehm., Pl. Preiss. 2: 109 (1846–47) ≡ Lycopodioides gracillima (Kunze) Kuntze, Rev. Gen. Pl. 1: 825 (1891) ≡ Selaginella gracillima (Kunze) Spring ex Salomon; Nomencl. 353 (1883).
Ericetorum lyallii (Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium lyallii Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 2: 387, no. 120 (1831) ≡ Selaginella laevigata var. lyallii (Hook. & Grev.) Baker, J. Bot. 23: 116 (1885) ≡ S. lyallii (Hook. & Grev.) Spring; Bull. Acad. Roy. Soc. Bruxelles 10: 146, no. 84 (1843).
Ericetorum moratii (W. Hagemann & Rauh) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella moratii W. Hagemann & Rauh; Pl. Syst. Evol. 176(3–4): 205 (1991).
Ericetorum pygmaeum (Kaulf.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium pygmaeum Kaulf., Enum. Fil. 9 (1824) ≡ Lycopodium pumilum var. pygmaeum (Kaulf.) Schltdl., Adumbr. Fl. Aethiop. 6 (1825) ≡ Lycopodioides pygmaeum (Kaulf.) Kuntze, Rev. Gen. Pl. 1: 825 (1891) ≡ Selaginella pygmaea (Kaulf.) Alston; J. Bot. (London) 69: 257 (1931).
Ericetorum pectinatum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pectinata Spring, Bull. Acad. Roy. Sci. Bruxelles 10(1): 146. 1843, nom. nov. for Lycopodium pectinatum Willd., Sp. Pl., ed. 4. 5(1):44. 1810, nom. illeg., non Lamarck (1792: 651). – Selaginella polymorpha Badre’, Fl. Madagasc. fam. 14:25. 1997, a nom. nov. but superfl. for L. pectinatum Willd. – Lectotype (designated by Smith et al., 2016): “America meridionale?” [protologue], “Habitat in Madagascar” [specimens], B–W 19400-02 0.
Ericetorum royenii (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella royenii Alston; Nova Guinea ser. 2, 7: 2 (1956).
Ericetorum uliginosum (Labill.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium uliginosum Labill., Nov. Holl. pl. spec. 2: 154, t. 251, f. 1 (1806) ≡ Lycopodioides uliginosa (Labill.) Kuntze, Rev. Gen. Pl. 1: 827 (1891) ≡ L. uliginosum Labill., Nov. Holl. pl. spec. 2: 154, t. 251, f. 1 (1806) ≡ Selaginella uliginosa (Labill.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 136 (1843).
5.4.4. Gymnogynum
Gymnogynum P. Beauv., Mag. Encycl. 9(5): 480 (1804) 关节卷柏属(新拟) ≡ Lycopodium subg. Gymnogynum (P. Beauv.) Rchb., Consp. Regn. Veg.: 78 (1828) ≡ Didiclis sect. Gymnogynum (P. Beauv.) Rothm., Feddes Repert. Spec. Nov. Regni Veg. 54: 70 (1944) – Type: Gymnogynum domingense P. Beauv., Mag. Encycl. 9(5): 480 (1804) [= Selaginella plumosa (L.) C. Presl ≡ Lycopodium plumosum L.].
= Selaginella sect. Articulatae (Spring) Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1132 (2015) ≡ Selaginella (unranked) Articulatae Spring in Mém. Acad. Sci. Belg. 24: 53 (1850) ≡ S. ser. Articulatae (Spring) Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. 1(4): 707 (1902) ≡ Didiclis sect. Articulatae (Spring) Rothm. in Feddes Repert. Spec. Nov. Regni Veg. 54: 71. (1944) – Lectotype (designated by Rothmaler, 1944: 71): Didiclis sulcata (Desv. ex Poir.) Rothm. (= Selaginella sulcata (Desv. ex Poir.) Spring ex Mart.).
= Selaginella (unranked) Ascendentes A. Braun in App. Ind. Sem. Hort. Berol. 11 ([1857] 1858) non A. Braun (1858: 11) – Lectotype (designated by Zhou and Zhang, 2015: 1132): Selaginella galeottii Spring (= Selaginella stellata Spring).
= Selaginella (unranked) Repentes A. Braun in App. Ind. Sem. Hort. Berol. 12. [1857] 1858, non A. Braun (1858: 11) – Lectotype (designated by Zhou and Zhang 2015: 1132): Selaginella hortensis Mett. (= Selaginella kraussiana (G. Kunze) A. Braun).
= Selaginella (unranked) Caulescentes A. Braun in App. Ind. Sem. Hort. Berol. 12 ([1857] 1858), non A. Braun (1858: 11) – Type: Selaginella asperula Spring.
Selaginella subser. Pleiostelicae Hieron. & Sadeb. in Engler. & Prantl, Nat. Pflanzenf. 1(4): 710 (1902) non Hieronymus and Sadebeck, 1901 (1902: 700, 707) – Type: Selaginella kraussiana (G. Kunze) A. Braun.
= Selaginella subser. Monostelicae Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. 1(4): 708 (1902) non Hieronymus and Sadebeck, 1901 (1902: 673, 704) – Type: Selaginella remotifolia Spring.
Gymnogynum corresponds to Selaginella sect. Articulatae sensu Zhou and Zhang (2015) and to S. subg. Gymnogynum sensu Weststrand and Korall (2016b). Gymnogynum is always well supported as monophyletic in morphology (Mickel et al., 2004) and phylogeny (Weststrand and Korall, 2016a; Zhou et al., 2016; our Figs. 1 and S1–S3).
Morphologically, Gymnogynum has articulation (presenting swellings or nodules: Fig. 5I) on stems and branches (Lopes et al., 2020), strobilus with only one large megasporophyll at the base (Fig. 5J), rhizophores borne on dorsal side of stem or/and branch (Fig. 5B, F–I), megaspores large and with reticulate ornamentation on surfaces (Korall and Taylor, 2006; Zhou et al., 2015; Fig. 5R), and microspore surfaces usually spiny or echinate (Fig. 5M; Zhou et al., 2015; Valdespino and Lòpez, 2020).
The ndh gene of plastomes in Gymnogynum is preserved (Zhou et al., 2022).
This genus contains about 50 species mainly in North to South Americas and a few in Africa and Asia. Gymnogynum kraussianum is native to Africa and has been cultivated throughout the world and has been recently transferred to Gymnogynum by Weakley (2022) as G. kraussianum (Kunze) Weakley.
Members:
Gymnogynum atirrense (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella atirrensis Hieron., Nat. Pflanzenf. 1(4): 711 (1901).
Gymnogynum anaclastum (Alston ex Crabbe & Jermy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella anaclasta Alston ex Crabbe & Jermy, Am. Fern J. 63(3): 135 (1973).
Gymnogynum angustifolium (A. Braun) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella sericea var. angustifolia A. Braun; Ann. Sci. Nat., Bot., sér. V, 3: 299 (1865).
Gymnogynum arthriticum (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella arthritica Alston, Arch. Bot. (Forli) 11: 43 (1935).
**Gymnogynum articulatum (**Kunze) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium articulatum Kunze, Linn. 9: 10 (1834 publ. 1835) ≡ Selaginella articulata (Kunze) Spring, Flora 21: 182 (1838) ≡ Lycopodioides articulata (Kunze) Kuntze, Rev. Gen. Pl. 1: 825 (1891).
Gymnogynum asperulum (Mart. ex Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium asperulum C.Mart. ex Spring in Mart. Fl. Bras. 1(2): 127, no. 15 (1840) ≡ Lycopodioides asperula (Mart. ex Spring) Kuntze, Rev. Gen. Pl. 1: 825 (1891) ≡ Selaginella asperula Mart. ex Spring, Mart., Fl. Bras. 1(2): 127 (1840).
Gymnogynum asplundii (Alston ex Crabbe & Jermy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella asplundii Alston ex Crabbe & Jermy, Fern Gaz. 11(4): 257 (1976).
Gymnogynum buchtienii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella buchtienii Hieron., Med. Rijks Herb. Leiden, no. 27: 2 (1915).
Gymnogynum caluffii (Shelton) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella caluffii Shelton, Willdenowia 33(1): 159 (2003).
Gymnogynum coarctatum (Mart. ex Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella coarctata Mart. ex Spring, Mart., Fl. Bras. 1(2): 126, no. 14 (1840) ≡ Lycopodioides coarctata (Mart. ex Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Gymnogynum conduplicatum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella conduplicata Spring, Mart., Fl. Bras. 1(2): 129 (1840) ≡ Selaginella geniculata subsp. conduplicata (Spring) Hieron. in Engl. & Prantl, Nat. Pflanzenf. 1(4): 712 (1901 publ. 1902) ≡ Selaginella geniculata var. conduplicata (Spring) A. Braun, Ann. sc. nat. V, 3: 303 (1865).
Gymnogynum decompositum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella decomposita Spring, Mart., Fl. Bras. 1(2): 123, no. 10 (1840).
**Gymnogynum diffusum (**C. Presl) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium diffusum C. Presl, Rel. Haenk. 1(1): 78 (1825) (nom. illeg., non R. Br. (1810)) ≡ Selaginella diffusa (C. Presl) Spring, Bull. Acad. Roy. Soc. Bruxelles 10 (1): 143 (1843).
Gymnogynum eurynotum (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella eurynota A. Braun, Ann. sc. nat. V, 3: 293 (1865) ≡ Lycopodioides eurynota (A. Braun) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Gymnogynum expansum (Sodiro) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella expansa Sodiro, Rec. Cr. vasc. quit. 95, no. 12 (1883) ≡ Lycopodioides expansa (Sodiro) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Gymnogynum flabellum (Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella flabellum (Desv.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 234, no. 157 (1843).
Gymnogynum fragile (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella fragilis A. Braun, Ann. sc. nat. V, 5, 3: 305 (1865).
Gymnogynum filicaule (Sodiro) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella filicaulis Sodiro, Rec. Cr. vasc. quit. 600 (1893), et Anal. Univ. Centr. Ecuador 12: 413 (1895).
Gymnogynum fuertesii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella fuertesii Hieron., Urban, Symb. Ant. 7: 164 (1912).
**Gymnogynum geniculatum (**C. Presl) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium geniculatum C. Presl, Rel. Haenk. 1(1): 80 (1825) ≡ Selaginella geniculata (C. Presl) Spring, Bull. Acad. Roy. Soc. Bruxelles 10(1): 230 (1843) ≡ Lycopodioides geniculata (C. Presl) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Gymnogynum germinans (Valdespino & C. López) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella germinans Valdespino & C. López, Botany Letters 165(3–4): 488 (2018).
Gymnogynum horizontale (C. Presl) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium horizontale C. Presl, Rel. Haenk. 1(1): 78 (1825) ≡ Selaginella horizontalis (C. Presl) Spring; Bull. Acad. Roy. Soc. Bruxelles 10(1): 226 (1843).
Gymnogynum humboldtianum (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella humboldtiana A. Braun; Ann. Sci. Nat., Bot., sér. V, 5, 3: 293 (1865).
Gymnogynum ivanii (Shelton & Caluff) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella ivanii Shelton & Caluff, Willdenowia 33(1): 162 (2003).
Gymnogynum kraussianum (Kunze) Weakley; J. Bot. Res. Inst. Texas 16(2): 405 (2022). Basionym: Lycopodium kraussianum Kunze; Linnaea 18: 114 (1844). 小翠云 (栽培)
Gymnogynum kunzeanum (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kunzeana A. Braun, Ann. sc. nat. V, 3: 296–297, no. 35 (1865) ≡ Lycopodioides kunzeana (A. Braun) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Gymnogynum lingulatum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lingulata Spring, Mém. Acad. Roy. Sci. Belg. 24: 224 (1849) ≡ Lycopodioides lingulata (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
**Gymnogynum marginatum (**Humb. & Bonpl. ex Willd.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium marginatum Humb. & Bonpl. ex Willd., Sp. Pl. 5: 41, no. 58 (1810) ≡ Selaginella marginata (Humb. & Bonpl. ex Willd.) Spring, Flora 21: 194 (1838) ≡ Lycopodioides marginata (Kunth) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Gymnogynum microtus (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella microtus A. Braun, Ann. sc. nat. V, 3: 293, no. 32 (1865).
Gymnogynum moranianum (Valdespino & C. López) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella moraniana Valdespino & C. López, Brittonia 72(1): 24 (2019).
**Gymnogynum parkeri (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium parkeri Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 2: 388, no. 123 (1831) ≡ Selaginella parkeri (Hook. & Grev.) Spring, Bull. Acad. Roy. Sci. Bruxelles 10: 146 (1843) ≡ Lycopodioides parkeri (Hook. & Grev.) Kuntze, Rev. Gen. Pl. 2: 827 (1891).
Gymnogynum parviarticulatum (W.R. Buck) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella parviarticulata W.R. Buck, Brittonia 38(1): 45 (1986).
Gymnogynum pedatum (Klotzsch) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pedata Klotzsch, Linnaea 17: 521 (1844).
Gymnogynum plumosum (L.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium plumosum L., Sp. Pl. 2: 1100 (1753) ≡ Stachygynandrum plumosum (L.) P. Beauv., Fl. D'Oware, 1: 10 (1805) ≡ Lycopodioides plumosa (L.) Kuntze, Rev. Gen. Pl. 1: 825 & 827 (1891) ≡ Selaginella plumosa (L.) C. Presl, Abh. (K.) Böhm. Ges. Wiss., Math.-Naturw. Cl. V, 3: 583 (1845).
Gymnogynum poeppigianum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium poeppigianum Hook. & Grev., Enum. Filic. J. Bot. (Hook.) Kew Misc. 2: 392–393, no. 143 (1831). ≡ Selaginella poeppigiana (Spring) Spring, Flora 21: 185 (1838). ≡ Lycopodioides poeppigiana (Spring.) Kuntze, Revis. Gen. Pl. 2: 827 (1891).
Gymnogynum remotifolium (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella remotifolia Spring, Miq. Pl. Jungh. 3: 276, no. 5 (1854) ≡ Lycopodioides remotifolia (Spring) H.S. Kung, Fl. Sichuan. 6: 65–67, t. 19, 10–15 (1988). 疏叶关节卷柏 (新拟)
Gymnogynum schizobasis (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella schizobasis Baker, J. Bot. 21: 333, no. 86 (1883) ≡ Lycopodioides schizobasis (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Gymnogynum sericeum (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sericea A. Braun, Ann. sc. nat. V, 3: 298–299, no. 37 (1865) ≡ Lycopodioides sericea (A. Braun) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Gymnogynum sertatum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sertata Spring, Mém. Acad. Roy. Sci. Belg. 24: 104 (1849) ≡ Lycopodioides sertata (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Gymnogynum silvestre (Aspl.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella silvestris Aspl., Ark. för Bot. 20A(7): 30, f. 3–5 (1926).
Gymnogynum stellatum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella stellata Spring, Flora 21: 188 (1838) ≡ Selaginella parkeri var. stellata (Spring) Baker, J. Bot. 23: 120 (1885) ≡ Selaginella stolonifera var. stellata Spring, Flora 21: 194 (1838).
**Gymnogynum suave (**Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sulcata ssp. suavis Spring, Flora 21: 185 (1838) ≡ Selaginella suavis (Spring) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 229 (1843) ≡ Lycopodioides suavis (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
**Gymnogynum sulcatum (**Desv. ex Poir.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium sulcatum Desv. ex Poir., Lam., Encycl. suppl. 3: 549, no. 74. (1813 publ. 1814) ≡ Selaginella sulcata (Desv. ex Poir.) Spring ex Mart., Flora 20 (2): 126 (1837) ≡ D. sulcata (Desv.) Rothm., Fedde, Repert. Spec. Nov. 54: 71 (1944) ≡ Lycopodioides sulcata (Desv.) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Gymnogynum tardum (Mickel & Beitel) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tarda Mickel & Beitel, Mem. New York Bot. Gard. 46: 353, f. 128H–N (1988).
Gymnogynum tenuissimum (Fée) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tenuissima Fée, Cr. vasc. Br. 2: 98, no. 6/2, t. 108 (1873) ≡ Lycopodioides tenuissima (Fée) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Gymnogynum tomentosum (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tomentosa Spring, Mém. Acad. Roy. Sci. Belg. 24: 231 (1849) ≡ Selaginella geniculata var. tomentosa (Spring) Baker, J. Bot. 23: 121 (1885).
Gymnogynum trisulcatum (Aspl.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella trisulcata Aspl., Ark. för Bot. 20A(7): 34, f. 6 (1926).
Gymnogynum versatile (A.R.Sm.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella versatilis A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 270 (1990).
5.4.5. Lepidoselaginella
Lepidoselaginella Li Bing Zhang & X.M. Zhou, gen. nov. 鳞叶卷柏属(新拟) – Type: Lepidoselaginella lepidophylla (Hook. & Grev.) Li Bing Zhang & X.M. Zhou ≡ Lycopodium lepidophyllum Hook. & Grev., Icon. Filic. 2(9): 162 (1830) ≡ Selaginella lepidophylla (Hook. & Grev.) Spring, Fl. Bras. (Martius) 1(2): 126 (1840).
= Selaginella sect. Lepidophyllae Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1133 (2015) – Type: Selaginella lepidophylla (Hook. & Grev.) Spring.
Etymology: “_Lepido-_” derived from lepidophylla, referring to the scales-looking leaves of the type species.
Lepidoselaginella corresponds to Selaginella sect. Lepidophyllae sensu Zhou and Zhang (2015) and S. subg. Lepidophyllae sensu Weststrand and Korall (2016b). It is strongly supported as sister to Bryodesma (Weststrand and Korall, 2016a; Zhou et al., 2016; our Figs. 1 and S1–S3).
Morphologically, Lepidoselaginella has rosette habit (Fig. 5D) and rhizophores borne on dorsal side of the stem. Although some taxa in other genera (e.g., Selaginella pallescens, S. nothohybrida, and most members of Pulviniella) also have rosette habit, they have rhizophores borne on ventral side of the stem_. Lepidoselaginella_ has megaspores with ridges on the distal surface (Korall and Tryon, 2006) and microspores with striped ornamentation which are different from rest of species in the family (Fig. 5W).
The plastome of Lepidoselaginella has IR structure (Zhou et al., 2022).
Lepidoselaginella contains about three species occurring in Mexico and USA (Mickel and Smith, 2004; Weststrand and Korall, 2016a; Zhou et al., 2016).
Members:
**Lepidoselaginella lepidophylla (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium lepidophyllum Hook. & Grev., Icon. Fil. 2(9): 162 (1830), et Enum. Fil. J. Bot. (Hook.) Kew Misc. 3: 106 (1833) ≡ Selaginella lepidophylla (Hook. & Grev.) Spring, Mart., Fl. Bras. 1(2): 126 (1840) ≡ Lycopodioides lepidophylla (Hook.) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Lepidoselaginella novoleonensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella novoleonensis Hieron. in Engl. & Prantl, Nat. Pflanzenf. 1(4): 676, no. 56. (1901 publ. 1902).
Lepidoselaginella ribae (Valdespino) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella ribae Valdespino, Mem. New York Bot. Gard. 88 (Pterid. Mexico): 591–592 (2004).
5.4.6. Megaloselaginella
Megaloselaginella Li Bing Zhang & X.M. Zhou, gen. nov. 大孢卷柏属(新拟) – Type: Megaloselaginella exaltata (Kunze) Li Bing Zhang & X.M. Zhou (≡ Lycopodium exaltatum Kunze ≡ Selaginella exaltata (Kunze) Spring).
= Selaginella sect. Megalosporarum Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1133 (2015 - Type: S. exaltata Kunze.
Etymology: “_Megalo-_” derived from Megalosporarum, referring to the extremely large megaspores (ca. 1.5 mm).
Megaloselaginella corresponds to Selaginella sect. Megalosporarum sensu Zhou and Zhang (2015) and part of S. subg. Exaltatae sensu Weststrand and Korall (2016b).
Megaloselaginella is very similar to Gymnogynum in having rhizophores borne on dorsal side of the stem and only one megasporophyll at the base of strobili (Fig. 5F). However, Megaloselaginella has large erect plants (up to 1 m) (Fig. 5E), several steles in a special actino-plectoste (a sort of three-lobed plectostele) (Lopes et al., 2020), largest megaspores in the family (ca. 1.5 mm; Mickel and Hellwig, 1969, Fig. 5O), and pillared to baculate ornamentation on microspore surfaces (Fig. 5T).
The plastome of Megaloselaginella known so far (Zhou et al., 2022) has DR-IR coexisting structure and the intron2 of _ycf_3 (Zhou et al., 2022).
This genus contains about three species in Central and South Americas and Africa (Steyermark et al., 1986; Zhou et al., 2016; Hassler, 2022).
Members:
**Megaloselaginella exaltata (**Kunze) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium exaltatum Kunze, Linn. 9: 8 (1834 [1835]) ≡ Selaginella exaltata (Kunze) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 234 (1843) ≡ Lycopodioides exaltata (Kunze) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Megaloselaginella gigantea (Steyerm. & A.R. Sm.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella gigantea Steyerm. & A.R.Sm., Ann. Mo. Bot. Gard. 73(1): 209 (1986).
Megaloselaginella grallipes (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella grallipes Alston, Mem. Soc. Linn. Normandie Bot. 1(3): 80 (1938).
5.5. Subfamily IV. Sinoselaginelloideae
Subfamily IV. Sinoselaginelloideae Li Bing Zhang & X.M. Zhou, subfam. nov. 中华卷柏亚科 (新拟) – Type: Sinoselaginella Li Bing Zhang & X.M. Zhou.
Plants erect, ascending, or creeping (Fig. 6A, G), with ventral rhizophores (Fig. 6B); sterile leaves dimorphic (Fig. 6A–C, E, G, H); sporophylls monomorphic (Fig. 6D, F); strobili with only one (to a few) megasporophyll at the base (Quanansah, 1988; Zhao et al., 2001); megaspore surfaces verrucate, prominent elements composed by long curved baculate (Fig. 6P) or rope microstructure (Zhou et al., 2015); microspore surfaces rugate (Fig. 6M−O), coarse (Fig. 6Q–T), sometimes with holes on proximal surfaces (Fig. 6Q–S).
The plastomes of Sinoselaginelloideae known so far (Zhou et al., 2022) have the smallest size (78–90 kb) in Selaginellaceae and the plastome structure is more like “network” rather than looping. Except rrn16 and rrn23, most of the plastid tRNA and rRNA genes are lost in Sinoselaginelloideae. Sinoselaginelloideae have the least protein-coding genes in Selaginellaceae (ca. 27) (Zhou et al., 2022).
Some species of this subfamily like most of land plant lineages have a low GC content (ca. 30%).
This subfamily corresponds to the Sinensis group sensu Korall and Kenrick (2004) and Weststrand and Korall (2016a).
It contains about 24 species in three genera: Austroselaginella, Korallia, and Sinoselaginella, occurring in Africa, Asia, Australia, and Indian Ocean islands.
5.5.1. Austroselaginella
Austroselaginella Li Bing Zhang & X.M. Zhou, gen. nov. 澳洲卷柏属(新拟) – Type: Austroselaginella australiensis (Baker) Li Bing Zhang & X.M. Zhou (≡ Selaginella australiensis Baker, J. Bot. 21: 144, no. 55 (1883)).
Austroselaginella has creeping plants, monomorphic strobili, and only one (to a few) megasporophyll at the base of strobili (Fig. 6F). A. australiensis is very similar to Gymnogynum remotifolium and G. kraussianum, but the former has the rhizophores borne on ventral side and non-articulate stems. Austroselaginella is also similar to some species with creeping plants in Korallia, but the former has the base of ventral leaves not auriculate (Fig. 5E vs. 5H) and microspores without holes on proximal surfaces (Fig. 5T vs. 5Q–S). Austroselaginella has very large megaspores (600–750 μm in diam.). Although Korall and Taylor (2006) reported that the megaspore surfaces of A. australiensis were strongly reticulate, their recent study (Weststrand and Korall, 2016) clarified that the “reticulate surfaces of megaspores” was probably based on a misidentified specimen. Jermy and Holmes (1998) reported that the megaspore surfaces of A. australiensis are faintly reticulate. We did not examine the megaspores of A. australiensis but its microspore surfaces are somewhat smooth (Fig. 6T).
It contains about four species distributed in rain-forested regions of north-eastern Queensland in Australia (Jermy and Holmes, 1998).
Members:
Austroselaginella andrewsii (Jermy & J.S. Holmes) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella andrewsii Jermy & J.S. Holmes, Fl. Australia 48: 705 (1998).
Austroselaginella australiensis (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella australiensis Baker, J. Bot. 21: 144, no. 55 (1883) ≡ Lycopodioides australiensis (Baker) Kuntze, Rev. Gen. Pl. 1: 825 (1891).
Austroselaginella brisbanensis (F.M. Bailey) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella brisbanensis F.M. Bailey, Queensl. Fl. Suppl. 62 (1886) ≡ Lycopodioides brisbanensis (F.M. Bailey) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Austroselaginella leptostachya (F.M. Bailey) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella leptostachya F.M. Bailey, Proc. Roy. Soc. Queensl. 1: 13 (1884) ≡ Selaginella australiensis var. leptostachya (F.M. Bailey) Domin, Bibl. Bot. 85: 237 (1913).
5.5.2. Korallia
Korallia Li Bing Zhang & X.M. Zhou, gen. nov. 科氏卷柏属(新拟) – Type: Korallia fissidentoides (Hook. & Grev.) Li Bing Zhang & X.M. Zhou (≡ Lycopodium fissidentoides Hook. & Grev., Enum. Fil. Hook. Bot., Misc. 2: 305, no. 151 (1831)).
= Selaginella sect. Oligomacrosporangiatae Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. 1(4): 704 (1902) ≡ Didiclis subg. Oligomacrosporangiatae (Hieron. & Sadeb.) Rothm. in Feddes Repert. Spec. Nov. Regni Veg. 54: 70 (1944) ["Oligomacrosporangiata"] – Lectotype (designated by Rothmaler, 1944: 70): Selaginella fissidentoides (Hook. & Grev.) Spring.
= Selaginella subser. Monostelicae Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. 1(4): 704 (1902) non Hieronymus and Sadebeck, 1901 (1902: 673, 708) – Lectotype (designated by Zhou and Zhang, 2015: 1135): Selaginella fissidentoides (Hook. & Grev.) Spring.
Etymology:— From _korall_-, in honor of Prof. Petra Korall of Uppsala University, Sweden, for her contributions to the study of ferns in general and that of Selaginella in particular (e.g., Korall et al., 1999; Korall and Kenrick, 2002, 2004; Weststrand and Korall, 2016a, b).
In our earlier classification (Zhou and Zhang, 2015), we thought Selaginella fissidentoides should be resolved in Didiclis and misapplied S. sect. Oligomacrosporangiatae to Didiclis. Korallia is a member of the Sinensis group and sister to Austroselaginella (Weststrand and Korall, 2016; our Figs. S1–S3).
Species of Korallia have erect, creeping or ascending plants and often a biauriculate base of ventral leaves (Fig. 6H). The megaspore surfaces are often covered with dense and thin thorns (Fig. 6P). Microspores have a special hole on each proximal district of the proximal surfaces (Fig. 6Q–S)—such structure was never found or reported in other genera in the family.
The master plastome of Korallia known so far (Zhou et al., 2022) can mediate 12 isomers through repeats in SC. Almost all genes do not have copies (except rrn16/23 and ndhE) and the GC content is larger than 50% in Korallia. Korallia is the only genus with two copies of ndhE in Selaginellaceae.
Korallia contains about 15 species distributed in Madagascar and adjacent Indian Ocean islands (e.g., Mauritius, Mayotte, Reunion, and Seychelles; Badré, 2008).
Members:
Korallia amphirrhizos (A. Braun ex Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella amphirrhizos A. Braun ex Hieron., Nat. Pflanzenf. 1(4): 705 (1902) ≡ S. fissidentoides var. amphirrhizos (A. Braun ex Hieron.) Stefanov. & Rakotondr., Novon 6(2): 208 (1996).
Korallia balfourii (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella balfourii Baker, Fl. Maurit. 522, no. 2 (1877) ≡ Lycopodioides balfourii (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
**Korallia cataphracta (**Willd.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium cataphractum Willd., Sp. Pl. 5: 43, no. 62 (1810) ≡ Selaginella cataphracta (Willd.) Spring, Flora 21: 209 (1839).
**Korallia concinna (**Sw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium concinnum Sw., Syn. Fil. 182, 408, no. 52 (1806) ≡ Selaginella concinna (Sw.) Spring, Flora 21: 188, no. 15 (1838).
Korallia distachya (Cordem.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella distachya Cordem., Bull. Soc. Sc. Arts La Réunion,104 (1890–91).
Korallia fissidentoides (Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium fissidentoides Hook. & Grev., Enum. Fil. Hook. Bot., Misc. 2: 305, no. 151 (1831) ≡ Selaginella fissidentoides (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 8(12): 142 (1841) ≡ Didiclis fissidentoides (Hook. & Grev.) Rothm., Fedde, Repert. Spec. Nov. 54: 70 (1944) ≡ Lycopodioides fissidentoides (Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
**Korallia fruticulosa (**Bory ex Willd.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium fruticulosum Bory ex Willd., Sp. Pl. 5: 41, no. 59 (1810) ≡ Selaginella fruticulosa (Bory) Spring, Flora 21: 202 (1838).
Korallia obtusa (P. Beauv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium obtusum (P. Beauv.) Desv. ex Poir., Lam., Encycl. suppl. 3: 548. (1813 publ. 1814) ≡ Selaginella obtusa (P. Beauv.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 228 (1843) ≡ Lycopodioides obtusa (P. Beauv.) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Korallia rodriguesiana (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rodriguesiana Baker, Fl. Maurit. 523, no. 4 (1877) ≡ Lycopodioides rodriguesiana (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Korallia sechellara (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sechellarum Baker, Fl. Maurit. 523, no. 523 (1877) ≡ Lycopodioides sechellarum (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Korallia serrulata (Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium serrulatum Desv. ex Poir., Lam., Encycl. suppl. 3: 550, no. 78. (1813 publ. 1814) ≡ Selaginella serrulata (Desv.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 228, no. 113 (1843).
**Korallia sinuosa (**Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium sinuosum Desv. ex Poir., Lam., Encycl. suppl. 3: 558. (1813 publ. 1814) ≡ Selaginella sinuosa (Desv.) Alston, J. Bot. 72: 230 (1934).
**Korallia sparsifolia (**Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium sparsifolium Desv. ex Poir., Lam., Encycl. suppl. 3: 553, no. 95. (1813 publ. 1814) ≡ Selaginella sparsifolia (Desv.) Badré, Fl. Mascar. Ptérid. 1: 45 (2008).
**Korallia tereticaulis (**Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium tereticaulon Desv. ex Poir., Lam., Encycl. suppl. 3: 551, no. 85. (1813 publ. 1814) ≡ Selaginella tereticaulis (Desv.) Spring, Flora 21: 210 (1838).
**Korallia viridula (**Bory) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium viridulum Bory ex Willd., Sp. Pl. 5: 37, no. 50 (1810) ≡ Selaginella viridula (Bory) Spring, Flora 21: 190 (1838).
5.5.3. Sinoselaginella
Sinoselaginella Li Bing Zhang & X.M. Zhou, gen. nov. 中华卷柏属(新拟) – Type: S. sinensis (Desv.) Li Bing Zhang & X.M. Zhou [≡ Lycopodium sinense Desv., Mém. Soc. Linn. Paris 6: 189 (1827) ≡ Selaginella sinensis (Desv.) Spring, Bull. Acad. Roy. Sci. Bruxelles 10(1): 137 (1843)].
Etymology:— “_Sino-_” derived from sinensis, referring to the Chinese distribution of the type species.
Sinoselaginella is part of the Sinensis group. It is sister to Austroselaginella + Korallia. Morphologically, Sinoselaginella is xerophytic and has creeping stems (Fig. 6A), rhizophores borne on ventral side in axils of branches (Fig. 6B), intervals throughout the main stem (Fig. 6A and B), only one megasporophyll on the ventral side of the base of strobili (Fig. 6F), large and usually verrucate and tuberculate ornamentation on megaspore surfaces and this prominent elements comprised by ropes (Fig. 6I–K; Zhou et al., 2015), and microspore surfaces with prominent verrucae, and/or somewhat ridges (Fig. 6M−N). Species of Sinoselaginella are very similar to some species of Boreoselaginella, especially Sinoselaginella albocincta and B. sanguinolenta, two species with extremely similar appearance (Kung, 1981) and overlapping distribution (Zhang et al., 2013), but B. sanguinolenta has strobili with several megasporophylls rather than one (Fig. 4E–G) and rhizophores borne on the dorsal side of stems (Fig. 4D).
Nearly all species in Sinoselaginella known so far (Zhou et al., 2022) have protein-coding genes with two or three copies. GC content of the plastome is ca. 30% in Sinoselaginella which is different from other taxa in Selaginellaceae (Zhou et al., 2022).
Sinoselaginella contains about five species (Sinoselaginella adunca, S. albocincta, S. chuweimingii, S. sinensis, and S. yemensis) distributed in China extending to the Arabic area and eastern Africa (Somalia, Ethiopia, Kenya) (Zhang et al., 2013; Fraser-Jenkins et al., 2015, 2017; Zhou et al., 2015).
Members:
Sinoselaginella adunca (A. Braun ex Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella adunca A. Braun ex Hieron. in Engl. & Prantl, Nat. Pflanzenf. 1(4): 674 (1901 publ. 1902).
Sinoselaginella albocincta (Ching ex H.S. Kung) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella albocincta Ching ex H.S. Kung, Acta Bot. Yunnan. 3(2): 251–252, f. 1: 9–14 (1981) ≡ Selaginella adunca ssp. albocincta (Ching) Fraser-Jenk., Indian Fern J. 25(1–2): 8 (2008 publ. 2009) ≡ Lycopodioides albocincta (Ching ex H.S. Kung) H.S. Kung, Fl. Sichuan. 6: 61 (1988). 白边中华卷柏(新拟)
Sinoselaginella chuweimingii (X.M. Zhou, Z.R. He, Liang Zhang & Li Bing Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella chuweimingii X.M. Zhou, Z.R. He, Liang Zhang & Li Bing Zhang, Phytotaxa 231(3): 284 (2015). 维明中华卷柏(新拟)
**Sinoselaginella sinensis (**Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium sinense Desv., Mém. Soc. Linn. Paris 6: 189 (1827) ≡ Selaginella sinensis (Desv.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 137, no. 19 (1843) ≡ Lycopodioides sinensis (Desv.) Satou, Hikobia 12(3): 269 (1997). 中华卷柏(新拟)
**Sinoselaginella yemensis (**Sw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium yemense Sw., Syn. Fil. 182, 407, no. 49, t. 4, f. 4 (1806) ≡ Selaginella yemensis (Sw.) Spring ex Decne., Arch. Mus. Paris 2: 191 (1941–42) ≡ Lycopodioides yemensis (Sw.) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
5.6. Subfamily V. Pulvinielloideae
Subfamily V. Pulvinielloideae Li Bing Zhang & X.M. Zhou, subfam. nov. 垫状卷柏亚科(新拟) – Type: Pulviniella (Li Bing Zhang & X.M. Zhou) Li Bing Zhang & X.M. Zhou.
Plants often rosette-forming (Fig. 7A and B), or erect and tufted when dry (Fig. 7C), with ventral rhizophores; sterile leaves dimorphic, four rows (Fig. 7A); sporophylls monomorphic (Fig. 7D); megaspores surface coarse, usually without obvious ornamentation (Fig. 7E–H); microspores globose, surfaces coarse or non-prominent verruca and granule (Fig. 7I–L); 2n = 20.
The plastome of Pulvinielloideae known so far (Zhou et al., 2022) has DR structure and only one conformation. Genes _rpl_16 tend to pseudogenize, and _trnC_-GCA and rps18 are lost in Pulvinielloideae. All ndh genes are lost or pseudogenized in Pulvinielloideae.
This subfamily corresponds to Selaginella subg. Pulviniella sensu Zhou and Zhang (2015).
It contains about 17 species in one genus, Pulviniella, occurring in Africa, Asia, and North and Central Americas.
5.6.1. Pulviniella
Pulviniella (Li Bing Zhang & X.M. Zhou) Li Bing Zhang & X.M. Zhou, stat. nov. 垫状卷柏属 (新拟) – Basionym: Selaginella subg. Pulviniella Li Bing Zhang & X.M. Zhou, Taxon 64 (6): 1133 (2015) – Type: Pulviniella pulvinata (Hook. & Grev.) Li Bing Zhang & X.M. Zhou ≡ S. pulvinata (Hook. & Grev.) Maxim.
Pulviniella corresponds to Selaginella subg. Pulviniella Li Bing Zhang & X.M. Zhou and our data resolved it as sister to the Sinensis group (Figs. 1 and S3). However, Weststrand and Korall's (2016a) study found it to be sister to a clade including S. subg. Heterostachys sensu Zhou and Zhang (2015) and S. subg. Stachygynandrum sensu Zhou and Zhang (2015).
Most species of Pulviniella have rosette habit. The rosette habit evolved at least three times in Selaginellaceae: once in Pulviniella (e.g., S. pulvinata), once in Lepidoselaginella (e.g., L. lepidophylla and L. novoleonensis), and once in Selaginella s.s. (e.g., the S. pallescens + S. nothohybrida clade), respectively. However, except those in Pulviniella, all species with rosette habit in Selaginellaceae have reticulate ornamentation. 2n = 20 or 24 (Jermy et al., 1967; Takamiya,1993; Marcon et al., 2005).
Pulviniella and some species of Sinoselaginella often overlap in distribution and share xerophyte habit in China. For example, the _S. chuweimingii_-P. pulvinata pair and the _S. sinensis_-P. tamariscana pair, respectively, usually co-occur in a same habitat in China (Chu et al., 2006; Zhang et al., 2013; our field observations), but they are not sister to each other, which is consistent with morphology.
Pulviniella contains ca. 17 species occurring through Africa, Asia, and North & Central Americas (Zhang et al., 2013; Fraser-Jenkins et al., 2015, 2017; Zhou et al., 2015; Yang et al., 2023a).
Members:
**Pulviniella algida (**Jie Yang bis & X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella algida Jie Yang bis & X.C. Zhang, Taxon 72: 14–15 (2023). 高寒垫状卷柏(新拟)
**Pulviniella bryopteris (**L.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium bryopteris L., Sp. Pl. 2: 1103 (1753) ≡ Selaginella bryopteris (L.) Baker, J. Bot. 22: 376 (1884) ≡ Lycopodioides bryopteris (L.) Kuntze, Rev. Gen. Pl. 1: 825 (1891) ≡ Stachygynandrum bryopteris P. Beauv., Prodr. aethéogam. 109 (1805).
**Pulviniella convoluta (**Arn.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium convolutum Arn., Mem. Wern. Nat. Hist. Soc. 5: 199 (1824) ≡ Lycopodioides convoluta (Arn.) Kuntze, Rev. Gen. Pl. 1: 826 (1891) ≡ Selaginella convoluta (Arn.) Spring, Mart., Fl. Bras. 1 (2): 131 (1840).
Pulviniella digitata (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella digitata Spring, Mém. Acad. Roy. Sci. Belg. 24: 75 (1850) ≡ Lycopodioides digitata (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
**Pulviniella graniticola (**Jie Yang bis & X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella graniticola Jie Yang bis & X.C. Zhang, Taxon 72: 15. 2023. 花岗岩垫状卷柏(新拟)
Pulviniella gypsophila (A.R.Sm. & T. Reeves) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella gypsophila A.R.Sm. & T. Reeves, Sida 10(3): 211 (1984).
Pulviniella helioclada (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella helicoclada Alston, Dansk Bot. Ark. 7: 195, t. 78(10–12) (1932), nom. nud., et ex C. Chr., Perrier Cat. 71 (1932).
**Pulviniella imbricata (**Forssk.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium imbricatum Forssk., Fl. Aegypt.-Arab. 125: 1 (1775) ≡ Selaginella imbricata (Forssk.) Spring ex Decne., Arch. Mus. Hist. Nat. 2: 193, t. 7 (1841) ≡ Lycopodioides imbricata (Forssk.) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Pulviniella iridescens (X.C. Zhang & Y.R. Wang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella iridescens X.C. Zhang & Y.R. Wang, Taxonomy 1: 305. (2021). 彩虹垫状卷柏(新拟)
Pulviniella nubigena (J.P. Roux) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella nubigena J.P. Roux, Bothalia 38(2): 154 (2008).
**Pulviniella orientali-chinensis (**Ching & C.F. Zhang ex Hao Wei Wang & W.B. Liao) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella orientali-chinensis Ching & C.F. Zhang ex Hao Wei Wang & W.B. Liao, Acta Sci. Nat. Univ. Sunyatsenia 61(2): 305 (2022). 华东垫状卷柏(新拟)
Pulvinae pilifera (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pilifera A. Braun, Sitz. Ges. Naturf. Freunde, Jan. 1857 ex Index 1866, p. 2 ≡ Lycopodioides pilifera (A. Braun) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Pulviniella pseudotamariscina (X.C. Zhang & C.W. Chen) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pseudotamariscina X.C. Zhang & C.W. Chen, Guihaia: 42 (10): 1635 (2022).
**Pulviniella pulvinata (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium pulvinatum Hook. & Grev., J. Bot. (Hook.) Kew Misc. 2: 381 (1831) ≡ Selaginella pulvinata (Hook. & Grev.) Maxim., Mém. Ac. Imp. Sci. St. Pétersb. 9: 335 (1859) ≡ Lycopodioides pulvinata (Hook. & Grev.) H.S. Kung, Fl. Sichuan. 6: 64, t. 18, 1–3 (1988) ≡ Selaginella tamariscina var. pulvinata (Hook. & Grev.) Alston, Bull. Fan Mem. Inst. Biol. Bot. 5: 271 (1934). 垫状卷柏(新拟)
Pulviniella qinbashanica (Jie Yang bis & X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella pulvinata subsp. qinbashanica Jie Yang bis & X.C. Zhang, Taxon 72: 16–17 (2023). 秦巴垫状卷柏(新拟)
Pulviniella stauntoniana (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella stauntoniana Spring, Mém. Acad. Roy. Sci. Belg. 24: 71 (1850) ≡ Lycopodioides stauntoniana (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891). 旱生垫状卷柏(新拟)
**Pulviniella tamariscina (**P. Beauv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Stachygynandrum tamariscinum P. Beauv., Mag. Encycl. 9(5): 483 (1804), also Prodr. aethéogam. 106 (1805) ≡ Lycopodium tamariscinum (P. Beauv.) Desv. ex Poir., Lam., Encycl. suppl. 3: 540. (1813 publ. 1814) ≡ Selaginella tamariscina (P. Beauv.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 136, no. 9 (1843) ≡ Lycopodioides tamariscina (P. Beauv.) H.S. Kung, Fl. Sichuan. 6: 62–64, t. 18, 4–6 (1988). 卷柏
5.7. Subfamily VI. Lycopodioidoideae
Subfamily VI. Lycopodioidoideae Li Bing Zhang & X.M. Zhou, subfam. nov. 异穗卷柏亚科(新拟) – Type: Lycopodioides Boehm.
Plants creeping (Fig. 8B, D–H, K), prostrate, ascending (Fig. 8E), scandent (Fig. 8L) or erect (Fig. 8A, I, J), with ventral rhizophores(Fig. 8D, G); sterile leaves dimorphic (Fig. 8), four rows; sporophylls monomorphic (Fig. 9E, H, J) or dimorphic [non-resupinate (Fig. 9F and G) or resupinate (Fig. 9B–D, I)]; megaspore surfaces often verrucate to tuberculate, few reticulate, smooth (Fig. 9K–N, S–V); microspore surfaces can be cristate, verrucate, granulate, laminate, blunt spiny, baculate, or smooth, depending on different genera (Fig. 9O–R, W, Y, Z); 2n = 16, 18, 20, 32 (Jermy, 1967; Loyal and Kumar, 1984; Takamiya, 1993; Mukhopadhyay and Goswami, 1996; Marcon et al., 2005).
This subfamily corresponds to Selaginella subg. Heterostachys sensu Zhou and Zhang (2015).
Lycopodioidoideae comprise those species with dimorphic or slightly dimorphic and resupinate sporophylls (Hypopterygiopsis), dimorphic and non-resupinate sporophylls (most species of Lycopodioides), or monomorphic sporophylls (Valdespinoa and nearly all species of Didiclis).
Traditionally, Selaginella subg. Heterostachys only contains those species with dimorphic and resupinate sporophylls in Selaginella (e.g., Baker, 1883, 1887; Warburg, 1900; Walton and Alston, 1938; Jermy, 1986, 1990) before our classification (Zhou and Zhang, 2015). However, morphological analysis shows that species with dimorphic sporophylls are not monophyletic (Zhou et al., 2016) and dimorphic sporophylls have evolved at least 13 times in the family (Fig. 2G).
Based on our study on the phylogeny, gross morphology, spore morphology, and geographic distribution, all species with dimorphic sporophylls and resupinate strobili in the Old World (Asia, Pacific islands, Africa, and Madagascar) should belong to Lycopodioidoideae. However, based on the morphology of megaspores and microspores and molecular data, those species with dimorphic sporophylls and resupinate strobili in the New World (mainly in South and Central Americas) are members of Selaginelloideae (see below). The dimorphic sporophylls and resupinate strobili evolved many times independently in Lycopodioidoideae and Selaginelloideae (Fig. 2G and H).
Lycopodioidoideae known so far (Zhou et al., 2022) have the most complicated and diverse plastome structures in Selaginellaceae. Two genera (Lycopodioides and Valdespinoa) have three ribosomal operon copies and the largest plastome size in Selaginellaceae. All of known plastome structures of Lycopodioidoideae are DR-IR coexisting structure (Zhou et al., 2022).
Lycopodioidoideae contain over 260 species in the Old World except one American species (Didiclis hoffmannii) in four genera, Didiclis, Hypopterygiopsis, Lycopodioides, and Valdespinoa, occurring in the Old World and New World.
5.7.1. Didiclis
Didiclis P. Beauv. ex Mirb., Hist. Nat. Vég. 3: 477, 4: 314 (1802) 瘤孢卷柏属(新拟) – Type: Didiclis ornithopodioides (L.) P. Beauv. ex J. St.-Hil (Selaginella ornithopodioides (L.). Spring, Flora 21(14): 216 (1838)).
= Selaginella ser. Suberosae Warb. in Monsunia 1: 110 (1900) – Type: Selaginella suberosa Spring_._
= Selaginella (unranked) Erectae A. Braun in App. Ind. Sem. Hort. Berol. 11 ([1857] 1858) – Type: Selaginella inaequalifolia Spring.
= Selaginella (unranked) Scandentes A. Braun in App. Ind. Sem. Hort. Berol. 11 ([1857] 1858) ≡ Selaginella ser. Scandentes (A. Braun) Baker in J. Bot. Lond. 21: 4 (1883) – Lectotype (designated here): Selaginella laevigata Spring (= S. willdenowii (Desv. ex Poir.) Baker).
= Selaginella ser. Sarmentosae Baker in J. Bot. Lond. 21: 4 (1883) – Lectotype (designated by Zhou and Zhang, 2015: 1135): Selaginella inaequalifolia Spring.
= Selaginella subser. Pleurophyllae Warb. in Monsunia 1: 106 (1900) – Lectotype (designated by Zhou and Zhang, 2015: 1135): Selaginella willdenowii (Desv. ex Poir.) Baker.
= Selaginella ser. Bisulcatae Warb. in Monsunia 1: 108 (1900) – Type: Selaginella bisulcata Spring.
= Selaginella ser. Pleiostelicae Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. 1(4): 700. (1902) ≡ Lycopodioides sect. Pleiostelica (Hieron. & Sadeb.) Rothm. in Feddes Repert. Spec. Nov. Regni Veg. 54: 69 (1944), “_Pleiostele_” – Lectotype (designated by Rothmaler, 1944: 69): Selaginella uncinata (Desv. ex Poir.) Spring.
Didiclis corresponds to “Selaginella sect. _Oligomacrosporangiatae_” sensu Zhou and Zhang (2015). The type of Selaginella sect. Oligomacrosporangiatae, S. fissidentoides, turned out to be a member of the Sinensis group (now Korallia) shown by Weststrand and Korall (2016a). The type shares one megasporangium only at the base of strobili with the Sinensis group (Quansah, 1988; Stefanović et al., 1997; also see above).
Although species of Didiclis show high diversity of gross morphology, they share tuberculate or verrucate megaspores and blunt-spiny to lamellate or cristate microspores (Zhou et al., 2015; Wang et al., 2018).
Comparing with other subfamilies and genera, Didiclis known so far has the most diverse master conformations ranging from 2 to 10 in their plastomes (Zhou et al., 2022).
Didiclis contains ca. 80 species from Asia and some species from Africa (e.g., Didiclis pervillei) and one species from America (D. hoffmannii) in the subfamily (Fig. S3).
Based on the morphology and phylogeny, Didiclis can be divided into six well identifiable groups (the Braunii group, the Bisulcata group, the Delicatula group, the Pervillei group, the Siamensis group, and the Willedenowii group) corresponding to six subclades in the phylogeny. These six groups each are distinguishable from Hypopterigyopsis.
- (1)
The Braunii group: This group is characterized by xerophytic and erect plants with monomorphic sterile leaves on stem, broadly ovate sporophylls, and sterile leaves usually involute when dry. Based on our field observations and specimen examination, this group contains about five species (Didiclis braunii, D. fulcrata, D. mairei, D. ostenfieldii, and D. pubescens) in South and Southeast Asia. - (2)
The Delicatula group: This group is similar to the Willedenowii group in having nearly entire leaves (except D. hoffmannii from America), erect and suberect plants, but has blunt-spiny ornamentation on microspores_._ This group contains more than seven species (Didiclis caudata, D. delicatula, D. picta, D. plana, D. mayeri, D. stipulata, and D. wallichii). - (3)
The Bisulcata group: This group is characterized by resupinate and extremely complanate strobili with dimorphic sporophylls, obovate dorsal leaves, and nearly smooth surfaces on megaspores_._ Dimorphic sporophylls and resupinate strobili evolved independently or reversed in this group. This group contains about five species (Didiclis bisulcata, D. obovata, D. opaca, D. pennata, and D. soyauxii) in South and Southeast Asia. - (4)
The Pervillei group: This group is very similar to the Braunii group in having xerophytic habit, erect and pubescent plants, and involute leaves when dry, but species of this group have verrucate ornamentation on microspores and are only distributed in Madagascar and South Africa. This group contains about three species (Didiclis eublepharis, D. pervillei, and D. vogelii). - (5)
The Siamensis group: This group has long-creeping, ascending or scandent plant_s._ It is similar to some species in the Willedenowii group, but has leaf margins ciliolate (v_s._ entire in the Willedenowii group), only one vascular bundle (vs. usually 3 vascular bundles in the Willedenowii group), and slightly xerophytic habit. This group contains only one species, Didiclis siamensis, in Southeast Asia. - (6)
The Willedenowii group: This group is easily distinguished in having a large plant size (up to 1 m), scandent habit, entire leaves, and lamellate to cristate ornamentation on microspores_._ This group contains ca. 60 species, the most of species in Didiclis (e.g., D. bamleri, D. helferi, D. limbata, D. pseudopaleifera, D. uncinata, D. viridangula, D. willdenowii, etc.) (Chen et al., 2017; Zhang et al., 2013).
These groups might deserve some taxonomical rank, but more samples need to be included and detailed morphological studies are necessary.
Members:
Didiclis axillifolia (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella axillifolia Alderw., Bull. Jard. Bot. Buitenz. II, 11: 37 (1913).
Didiclis bamleri (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella bamleri Hieron., Engl. Bot. Jahrb. 56: 239, no. 51 (1920).
Didiclis bisulcata (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella bisulcata Spring, Mém. Acad. Roy. Sci. Belg. 24: 259 (1850) ≡ Lycopodioides bisulcata (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 双沟瘤孢卷柏(新拟)
Didiclis braunii (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella braunii Baker, Gard. Chr. (1867): 783, 1120, no. 24 ≡ Lycopodioides braunii (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 布朗瘤孢卷柏(新拟)
Didiclis buergersiana (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella buergersiana Hieron., Engl. Bot. Jahrb. 56: 231, no. 30 (1920).
**Didiclis canaliculata (**L.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium canaliculatum L., Sp. Pl. 2: 1105 (1753) ≡ Stachygynandrum canaliculatum (L.) P. Beauv., Prodr. aethéogam. 110 (1805) ≡ Selaginella canaliculata (L.) Spring, Flora 21: 201, no. 21 (1838) ≡ Lycopodioides canaliculata (L.) Kuntze, Rev. Gen. Pl. 1: 824 (1891).
Didiclis caudata (Desv.S) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium caudatum Desv., Lam., Encycl. suppl. 3: 558 (1813 publ. 1814) ≡ Selaginella caudata (Desv.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 144 (1843) ≡ Selaginella canaliculata var. caudata (Desv.) Warb., Monsunia 1: 107, 122 (1900).
Didiclis commersonii (Spring) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella caudata var. commersonii Spring; Mém. Acad. Roy. Sci. Belgique 24 [Monogr. Lyc. 2]: 140 (1849).
Didiclis conferta (T. Moore) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella conferta T. Moore, Proc. Roy. Hort. Soc. 1: 133 (1861) ≡ Lycopodioides conferta (T. Moore) Kuntze, Rev. Gen. Pl. 1: 826 (1891)
Didiclis decurrens (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella decurrens Hieron., Engl. & Prantl, Nat. Pfl. 1(4): 703, no. 7 (1901 publ. 1902).
**Didiclis delicatula (**Desv. ex Poir.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium delicatulum Desv. ex Poir., Lam., Encycl. suppl. 3: 554, no. 99. (1813 publ. 1814) ≡ Selaginella delicatula (Desv. ex Poir.) Alston, J. Bot. 70(838): 282 (1932) ≡ Lycopodioides delicatula (Desv. ex Poir.) H.S. Kung, Fl. Sichuan. 6: 67–68, t. 20, 1–6 (1988). 薄叶瘤孢卷柏(新拟)
Didiclis dixitii (Madhus. & S. Nampy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella dixitii Madhus. & S. Nampy, Nordic J. Bot. 14(5): 527 (1994).
Didiclis dolichoclada (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella dolichoclada Alston, J. Bot. 70: 64, no. 5 (1932).
Didiclis engleri (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella engleri Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 704, no. 374. (1901 publ. 1902).
Didiclis eublepharis (A. Braun ex Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella eublepharis A. Braun ex Hieron., Nat. Pfl. 1 (4): 677 (1902).
Didiclis finium (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella finium Alderw., Bull. Jard. Bot. Buitenz. II, 16: 52 (1914).
**Didiclis fulcrata (**Buch.-Ham. ex D. Don) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium fulcratum Buch.-Ham. ex D. Don, Prodr. Fl. Nepal. 17 (1825) ≡ Selaginella fulcrata (Buch.-Ham. ex D. Don) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 231 (1843) ≡ Lycopodioides fulcrata (Buch.-Ham. ex D. Don) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Didiclis fulvicaulis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella fulvicaulis Hieron., Hedwigia 50: 29, no. 21 (1910).
Didiclis furcillifolia (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella furcillifolia Hieron., Hedwigia 50: 31, no. 22 (1910).
Didiclis gastrophylla (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella gastrophylla Warb., Monsunia 1: 107, 121, no. 103, t. 4, f. D (1900).
Didiclis gaudichaudiana (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella gaudichaudiana Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 145, no. 72 (1843).
Didiclis gracilis (T. Moore) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella gracilis T. Moore, Gard. Chr. 25, 1: 752, 2: 407 (1886).
Didiclis hainanensis (X.C. Zhang & Noot.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hainanensis X.C. Zhang & Noot., Bot. J. Linn. Soc. 148(3): 323 (2005). 琼海瘤孢卷柏(新拟)
Didiclis helferi (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella helferi Warb., Monsunia 1: 107, 121, no. 106 (1900). 攀缘瘤孢卷柏(新拟)
Didiclis hewittii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hewittii Hieron., Hedwigia 51: 262, no. 12 (1911 publ. 1912).
Didiclis hindsii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hindsii Hieron., Engl. Bot. Jahrb. 50: 2, 43, no. 18 (1913).
Didiclis hochreutineri (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hochreutineri Hieron., Ann. Jard. Cons. Bot. Genève 15–16: 228–230 (1912).
Didiclis hoffmannii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hoffmannii Hieron., Hedwigia 41: 184 (1902).
**Didiclis inaequalifolia (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium inequalifolium Hook. & Grev., J. Bot. (Hook.) Kew Misc. 2. 391 (1831) ≡ Selaginella inaequalifolia (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 145 (1843) ≡ Lycopodioides inaequalifolia (Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Didiclis ingens (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella ingens Alston, J. Bot. 72: 229 (1934).
Didiclis kittyae (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kittyae Alderw., Bull. Jard. Bot. Buitenz. II, 7: 35 (1912).
Didiclis lacerata (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lacerata Warb., Monsunia 1: 106, 120, no. 97 (1900).
Didiclis latupana (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella latupana Alderw., Bull. Jard. Bot. Buitenz. II, 16: 54 (1914).
Didiclis limbata (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella limbata Alston, J. Bot. 70(831): 62 (1932). 具边瘤孢卷柏(新拟)
Didiclis lobbii (Veitch ex A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lobbii Veitch ex A. Braun, Ind. Sem. Hort. Berol., 20 (1858) ≡ Lycopodioides lobbii (Veitch ex A. Braun) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Didiclis mairei (H. Lév.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella mairei H. Lév., Sert. Yunnan. 299 (1916). 狭叶瘤孢卷柏(新拟)
Didiclis maxima (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella maxima Alderw., Bull. Jard. Bot. Buitenz. II, 16: 53 (1914).
Didiclis mayeri (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella mayeri Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 700, no. 343. (1901 publ. 1902).
Didiclis megalura (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella megalura Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 702, no. 358 (1901 publ. 1902).
Didiclis megastachya (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella megastachya Baker, J. Bot. 23: 20, no. 202 (1885) ≡ Lycopodioides megastachya (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Didiclis obovata (S.Y. Dong) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella obovata S.Y. Dong, Syst. Bot. 47(1): 88 (2022). 倒卵叶瘤孢卷柏(新拟)
Didiclis opaca (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella opaca Warb., Monsunia 1: 108, 122, no. 112 (1900).
**Didiclis ornithopodioides (**L.) Spring Basionym: Lycopodium ornithopodioides L., Sp. Pl. 2: 1105, no. 23 (1753) ≡ Selaginella ornithopodioides (L.) Spring, Flora 21: 216, no. 25 (1838) ≡ Lycopodioides ornithopodioides (L.) Kuntze, Rev. Gen. Pl. 1: 824 & 827 (1891) ≡ Stachygynandrum ornithopodioides (L.) P. Beauv., Prodr. aethéogam. 110 (1805).
Didiclis ostenfeldii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella ostenfeldii Hieron., Bull. Herb. Boiss. II, 5: 721, no. 98 (1905).
Didiclis padangensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella padangensis Hieron., Hedwigia 50: 34, no. 23 (1910).
**Didiclis pennata (**D. Don) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium pennatum D. Don, Prodr. Fl. Nepal. 18 (1825) ≡ Selaginella pennata (D. Don) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 232 (1843) ≡ Lycopodioides pennata (D. Don) Kuntze, Rev. Gen. Pl. 2: 827 (1891). 拟双沟瘤孢卷柏(新拟)
Didiclis permutata (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella permutata Hieron., Hedwigia 50: 24, no. 19 (1910).
Didiclis pervillei (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pervillei Spring, Mém. Acad. Roy. Sci. Belg. 24: 169 (1850) ≡ Lycopodioides pervillei (Spring) Kuntze, Rev. Gen. Pl. 1: 825 (1891).
**Didiclis picta (**Griff.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium pictum Griff. ex Baker, J. Bot. 23: 19 (1885) ≡ Selaginella picta (Griff.) A. Braun ex Baker, J. Bot. 23(265): 19 (1885) ≡ Lycopodioides picta (Griff.) Kuntze, Rev. Gen. Pl. 1: 827 (1891). 黑顶瘤孢卷柏(新拟)
**Didiclis plana (**Desv. ex Poir.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium planum Desv. ex Poir., Lam., Encycl. suppl. 3: 554, no. 98. (1813 publ. 1814) ≡ Selaginella plana (Desv. ex Poir.) Hieron., Nat. Pfl. 1(4): 703 (1901).
**Didiclis polystachya (**Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella wallichii var. polystachya Warb., Monsunia 1: 106 (1900) ≡ Selaginella polystachya (Warb.) Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 702, no. 362. (1901 publ. 1902).
Didiclis praetermissa (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella praetermissa Alston, J. Bot. 70: 65 (1932).
Didiclis pseudopaleifera (Hand.-Mazz.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pseudopaleifera Hand.-Mazz., Sitzungsber. Akad. Wiss. Wien, 61: 82 (1924). 耳叶瘤孢卷柏(新拟)
**Didiclis pubescens (**Wall. ex Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium pubescens Wall., Num. List no. 123 (1829), nomen, ex Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 1: 383, no. 103 (1831) ≡ Selaginella pubescens (Wall. ex Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 225 (1843). 二歧瘤孢卷柏(新拟)
Didiclis rechingeri (Hieron. ex Rech.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rechingeri Hieron. ex Rech., Denkschr. Kaiserl. Akad. Wiss., Wien. Math.-Naturwiss. Kl. 89: 486 (1914).
Didiclis retroflexa (v.A.v.R.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella axillifolia var. retroflexa v.A.v.R., Bull. Jard. Bot. Buit. II, 11: 38. (1913).
Didiclis schlechteri (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella schlechteri Hieron., Bot. Jahrb. Syst. 50: 2, 41, no. 17 (1913).
**Didiclis semicordata (**Wall. ex Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium semicordatum Wall., (Cat. n. 126-3, 11 & 13 (1829), nom. nud.) ex Hook. & Grev., J. Bot. (Hook.) Kew Misc. 2: 396 (1831) ≡ Lycopodioides semicordata (Wall. ex Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 827 (1891) ≡ Selaginella semicordata (Wall. ex Hook. & Grev.) Spring, Mart., Fl. Bras. 1 (2): 122 (1840).
Didiclis siamensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella siamensis Hieron., Bot. Tidsskr. 24: 113 (1901). 泰国瘤孢卷柏(新拟)
Didiclis soyauxii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella soyauxii Hieron., Engl. & Prantl, Nat. Pfl. 1(4): 697, no. 305. (1901 publ. 1902).
Didiclis stipulata (Blume) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium stipulatum Blume, Enum. Pl. Jav. 2: 268, no. 18 (1830) ≡ Selaginella stipulata (Blume) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 145, no. 70 (1843).
Didiclis subbiflora (Hieron ex Brause) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella gracilis var. subbiflora Hieron ex Brause, Engl. Bot. Jahrb. 56: 238, no. 47. (1920).
Didiclis trichoclada (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella trichoclada Alston, J. Bot. 70: 63, no. 3 (1932). 毛枝瘤孢卷柏(新拟)
Didiclis tylophora (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tylophora Alderw., Bull. Jard. Bot. Buitenz. II, 16: 50 (1914).
**Didiclis uncinata (**Desv. ex Poir.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium uncinatum Desv. ex Poir., Lam., Encycl. Suppl. 3: 558 (1814) ≡ Lycopodioides uncinata (Desv.) Kuntze, Rev. Gen. Pl. 1: 825 (1891) ≡ Selaginella uncinata (Desv. ex Poir.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 141 (1843). 翠云草
Didiclis usterii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella usterii Hieron., ex Usteri, Beitr. Phil. Veg., 135 (1905), et Leafl. Phil. Bot. 6: 2055 (1913).
Didiclis velutina (Ces.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella velutina Ces., Rec. Ac. Sci. Nap. 16: 28, 31 (1877).
Didiclis victoriae (J.W. Moore) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella victoriae J.W. Moore, Cat. n. 143: 8, t. 6 (1878) ≡ Lycopodioides victoriae (J.W. Moore) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Didiclis viridangula (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella viridangula Spring, v. Heurck, Pl. Nov., 29 (1870) ≡ Lycopodioides viridangula (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Didiclis vogelii (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella vogelii Spring, Monogr. Lyc. 2: 170, no. 111 (1850).
**Didiclis wallichii (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium wallichii Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 2: 384, no. 106 (1831) ≡ Selaginella wallichii (Hook. & Grev.) Spring, Mart., Fl. Bras. 1 (2): 124 (1840). 瓦氏瘤孢卷柏(新拟)
**Didiclis willdenowii (**Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium willdenovii Desv. ex Poir., Lam., Encycl. suppl. 3: 540, 552, no. 87. (1813 publ. 1814) ≡ Selaginella willdenowii (Desv.) Baker, Gard. Chr. 783, 950 (1867) ≡ Lycopodioides willdenowii (Desv.) Kuntze, Rev. Gen. Pl. 1: 827 (1891). 藤瘤孢卷柏(新拟)
5.7.2. Hypopterygiopsis
Hypopterygiopsis Sakurai, Bot. Mag. (Tokyo) 57: 255 (1943) 异穗卷柏属(新拟) – Type: Hypopterygiopsis reptans Sakurai, Bot. Mag. (Tokyo) 57: 255 (1943) (≡ Selaginella sakuraii H.A. Mill.).
= Lycopodium (unranked) Platystachya Hook. & Grev. in Bot. Misc. 2: 380 (1831) ≡ Selaginella (unranked) Platystachyae (Hook. & Grev.) Spring in Mém. Acad. Sci. Belg. 24: 53 (1850) – Lectotype (designated by Zhou and Zhang, 2015: 1134): Selaginella chrysocaulos (Grev. & Hook.) Spring.
= Selaginella ser. Brachystachyae Warb. in Monsunia 1: 110 (1900) – Type: Selaginella brachystachya (Hook. & Grev.) Spring.
= Lycopodium (unranked) Planifolia Hook. & Grev., Bot. Misc. 2: 382 (1831) ≡ Selaginella (unranked) Planifoliae (Hook. & Grev.) Spring in Martius, Fl. Bras_._ 1(2): 118. (1840) – Lectotype (designated by Zhou and Zhang, 2015: 1135): Lycopodium tetragonostachyum Wall. ex Grev. & Hook.
= Selaginella (unranked) Pronae A. Braun, App. Ind. Sem. Hort. Berol. 12 ([1857] 1858) – Lectotype (designated by Zhou and Zhang, 2015: 1135): Selaginella ciliaris Spring.
= Selaginella ser. Proniflorae Warb., Monsunia 1: 108 (1900) – Type: Selaginella proniflora (L.) Baker.
The type of Hypopterygiopsis was described as a “moss” species (Sakurai, 1943) but it turned out to be a Selaginellaceae (Miller, 1967). Our examination of the type confirmed this_._ The type is not sampled in our molecular work and is assigned here based on the morphology.
Plant without creeping rhizome (vs. with creeping rhizome in the Didiclis braunii group and the D. pervillei group); stem with single vascular bundle (vs. often with three vascular bundles in the D. willedenowii group and the D. delicatula group); stem creeping to ascending, sometimes erect, never scandent (vs. scandent in the D. willedenowii group and the D. siamensis group); dorsal leaves rarely obovate (vs. obovate in the D. bisulcata group); sporophylls dimorphic or submonomorphic, resupinate, rarely strictly monomorphic (vs. always strictly monomorphic in Didiclis, except for the D. bisulcata group; often dimorphic, non-resupinate in Lycopodioides); megaspore surfaces often not interconnected verrucate to tuberculate, or finely reticulate (Tetragonostachyae type-1 and type-2) (vs. megaspore surfaces often interconnected verrucate to tuberculate and never reticulate in Didiclis); microspore surfaces often verrucate to spherulate, rarely smooth to coarse or baculate (vs. microspore surfaces blunt-spiny, lamellate, or cristate in the D. braunii group, the D. delicatula group, the D. siamensis group, and the D. willedenowii group).
This genus contains about 170 species in Africa and Asia. Based on the synapomorphy of strobili with dimorphic sporophylls, we combined Selaginella sect. Heterostachys and S. sect. Tetragonostachyae sensu Zhou and Zhang (2015) to form Hypopterygiopsis here. Within Hypopterygiopsis, we recognize two sections.
Hypopterygiopsis sect. Hypopterygiopsis 藓状异穗卷柏组(新拟)
= Selaginella (unranked) Tetragonostachyae (Hook. & Grev.) Spring in Mém. Acad. Sci. Belg. 24: 53 (1850). Basionym: Lycopodium (unranked) Tetragonostachya Hook. & Grev. in Bot. Misc. 2: 382 (1831) ≡ S. sect. Tetragonostachyae (Hook. & Grev.) Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. 1(4): 669 (1902) ≡ S. sect. Tetragonostachyae (Hook. & Grev.) Rothm. in Feddes Repert. Spec. Nov. Regni Veg. 54: 68 (1944), “_Tetragonostachya_” ≡ Lycopodioides subg. Tetragonostachya (Hook. & Grev.) Tzvel. in Novosti Sist. Vyssh. Rast. 36: 25 (2004) – Type: L. tetragonostachyum Wall. ex Grev. & Hook. [≡ Hypopterygiopsis tetragonostachya (Wall. ex Grev. & Hook.) Li Bing Zhang and X.M. Zhou].
This section corresponds to Selaginella sect. Tetragonostachyae sensu Zhou and Zhang (2015).
Based on the evolution of strobilus morphology, monomorphic sporophylls are reversed in this section. Although some species from Pacific islands have monomorphic to submonomorphic sporophylls, they have same megaspores as those species with dimorphic sporophylls in Asia. Species of this section have megaspore surfaces reticulate (Tetragonostachyae type-1 and type-2 with very fine muri and open meshes: Fig. 9L–N) and usually microspore surfaces smooth (Fig. 9P and Q), verrucate (Fig. 9O, R) or baculate (S. ciliaris; Zhou et al., 2015).
This section contains two well-supported clades in our phylogenetic analysis_._ One clade contains species with monomorphic sporophylls (e.g., Hypopterygiopsis arbuscula, H. kanehirae, H. whitmeei) mainly from Pacific islands (Kato, 2008; Chen et al., 2017), and the other contains species with strongly (H. ciliaris and H. xipholepis) or slightly dimorphic (H. repanda and H. vaginata) sporophylls mainly in Asia and some in Africa and Madagascar (Figs. S1–S3) (Quansah and Thomas, 1985; Stefanović, 1997; Zhang et al., 2013).
Hypopterygiopsis sect. Heterostachys (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. 异穗卷柏组(新拟) – Basionym: Selaginella subg. Heterostachys Baker in J. Bot. Lond. 21: 3 (1883) ≡ Selaginella sect. Heterostachys (Baker) Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1134 (2015) – Lectotype (designated by Jermy, 1986: 118): Hypopterygiopsis heterostachys (Baker) Li Bing Zhang & X.M. Zhou (≡ Selaginella heterostachys (Baker).
This section corresponds to Selaginella sect. Heterostachys sensu Zhou and Zhang (2015). This section contains the most species with resupinate strobili and dimorphic sporophylls from Asia and Africa (Quansah and Thomas, 1985; Stefanović, 1997; Zhang et al., 2013).
Morphologically, Selaginella sect. Heterostachys has often creeping (Fig. 8B and C), sometimes ascending or erect plants (Fig. 8A); resupinate strobili and dimorphic sporophylls (Fig. 9A); megaspore surfaces papillate, tuberculate, and verrucate (Fig. 9S, U; Zhou et al., 2015), sometimes elements connected (Fig. 9U); and microspore surfaces often verrucate, gemmulate, and spherulate (Fig. 9W–Y), rarely few tuberculate and rugate (Zhou et al., 2015).
Although dimorphic sporophylls and resupinate strobili also exist in Hypopterygiopsis sect. Hypopterygiopsis and some America species of Selaginella s.s. (see below), these species of the latter usually have megaspores with reticulate ornamentation (Fig. 9, Fig. 10; Korall and Taylor 2006; Bauer et al., 2016; Valdespino, 2017c).
Members of the genus:
**Hypopterygiopsis abyssinica (**Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella abyssinica Spring, Monogr. Lyc. 2: 99, no. 44 (1850) ≡ Selaginella goudotiana var. abyssinica (Spring) Bizzarri, Webbia 29: 585 (1975) ≡ Lycopodioides abyssinica (Spring) Kuntze, Rev. Gen. Pl. 1: 825 (1891).
Hypopterygiopsis albociliata (P.S. Wang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella albociliata P.S. Wang, J. Arnold Arbor. 71(2): 269 (1990). 白毛异穗卷柏(新拟)
Hypopterygiopsis alutacea (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella alutacea Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 233, no. 154 (1843) ≡ Lycopodioides alutacea (Spring) Kuntze, Rev. Gen. Pl. 1: 825 (1891).
Hypopterygiopsis amblyphylla (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella amblyphylla Alston, Bull. Fan Mem. Inst. Biol. Bot.5(6): 287–288 (1934). 钝叶异穗卷柏(新拟)
Hypopterygiopsis angustifolia (Hieron. ex Jeanpert) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella usta var. angustifolia Hieron. ex Jeanpert, Bull. Mus. Paris 17: 579 (nomen) (1911).
Hypopterygiopsis antimonanensis (B.C. Tan & Jermy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella antimonanensis B.C. Tan & Jermy, Fern Gaz. 12(3): 170 (1981).
Hypopterygiopsis apoensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella apoensis Hieron., Elmer, Leafl. Phil. Bot., 6: 2023, no. 18 (1913).
**Hypopterygiopsis arbuscula (**Kaulf.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium arbusculum Kaulf., Enum. Fil. 19 (1824) ≡ Selaginella arbuscula (Kaulf.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 227 (1843) ≡ Lycopodioides arbuscula (Kaulf.) Kuntze, Rev. Gen. Pl. 1: 825 (1891).
Hypopterygiopsis arbusculoides (J.W. Moore) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella arbusculoides J.W. Moore, Bull. Bernice P. Bishop Mus. 102: 15 (1933).
Hypopterygiopsis aristata (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella aristata Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 232, no. 152 (1843) ≡ Selaginella circinalis var. aristata C. Presl, Bot. Bem. 153 (1845).
Hypopterygiopsis atimonanensis (B.C. Tan & Jermy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella atimonanensis B.C. Tan & Jermy, Fern Gaz. 12(3): 170 (1981).
Hypopterygiopsis auquieri (Bizzarri) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella auquieri Bizzarri, Bull. Jard. Bot. Belg. 51(1–2): 220 (1981).
Hypopterygiopsis austro-orientalis (H.J. Wei & X.M. Zhou) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella austro-orientalis H.J. Wei & X.M. Zhou; Phytotaxa 579(2): 91 (2023). 东南异穗卷柏(新拟)
Hypopterygiopsis bankii (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella banksii Alston; Bull. Bernice P. Bishop Mus. 93: 15, 83 (1932).
Hypopterygiopsis beccariana (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella beccariana Baker, J. Bot. 23: 154, no. 254 (1885) ≡ Lycopodioides beccariana (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis behrmanniana (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella behrmanniana Hieron., Engl. Bot. Jahrb. 56: 233 (1920).
Hypopterygiopsis bemarahensis (Stefanović & Rakotondr.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella bemarahensis Stefanović & Rakotondr., Novon 6(2): 203 (1996).
Hypopterygiopsis blepharophylla (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella blepharophylla Alston, Mém. Inst. Fr. Afr. Noire 50: 40, t. 6, ff. 9–15 (1957).
Hypopterygiopsis bodinieri (Hieron. ex Christ) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella bodinieri Hieron. ex Christ, Bull. Ac. Géogr. Bot. Mans 11: 273, no. 109 (1902), nomen, et Hedwigia 43: 6, no. 25 (1904). 大叶异穗卷柏(新拟)
Hypopterygiopsis boninensis (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella boninensis Baker, J. Bot. 23 (270): 178 (1885) ≡ Lycopodioides boninensis (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 小笠原异穗卷柏(新拟)
**Hypopterygiopsis brachystachya (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium brachystachyum Hook. & Grev., J. Bot. (Hook.) Kew Misc. 3:107 (1833) ≡ Selaginella brachystachya (Hook. & Grev.) Spring, Bull. Acad. Roy. Sci. Bruxelles 10: 232 (1843) ≡ Lycopodioides brachystachya (Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis buchholzii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella buchholzii Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 696, no. 272 (1901 publ. 1902).
Hypopterygiopsis burbidgei (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella burbidgei Baker, J. Bot. 23: 154, no. 253 (1885).
Hypopterygiopsis busuensis (Alderw.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella darmandvillei var. busuensis Alderw.; Bull. Jard. Bot. Buitenzorg, sér. 2, 16. 50 (1914).
Hypopterygiopsis calcicola (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella calcicola Hieron., Hedwigia 51: 258, no. 9 ((1911) 1912).
**Hypopterygiopsis calostachya (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium calostachyum Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 3: 108 (1833) ≡ Selaginella calostachya (Hook. & Grev.) Alston, J. Bot. 70: 65, no. 8 (1932).
Hypopterygiopsis cataractarum (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella cataractarum Alston, Proc. Nat. Inst. Sci. India 11: 228 (1945).
Hypopterygiopsis chaetoloma (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella chaetoloma Alston, J. Bot. 70: 67, no. 12 (1932). 毛边异穗卷柏(新拟)
Hypopterygiopsis chaii (Jermy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella chaii Jermy, Kew Bull. 41(3): 551 (1986).
Hypopterygiopsis chingii (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella chingii Alston, J. Bot. 70: 66, no. 10 (1932). 秦氏异穗卷柏(新拟)
**Hypopterygiopsis chrysocaulos (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium chrysocaulon Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 2: 401, no. 182 (1831) ≡ Selaginella chrysocaulos (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 232 (1843) ≡ Lycopodioides chrysocaulos (Hook. & Grev.) H.S. Kung, Fl. Sichuan. 6: 78 (1988). 块茎异穗卷柏(新拟)
Hypopterygiopsis chrysorrhizos (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella chrysorrhizos Spring, Mém. Acad. Roy. Sci. Belg. 24 (2): 251 (1850) ≡ Lycopodioides chrysorhiza (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
**Hypopterygiopsis ciliaris (**Retz.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium ciliare Retz., Obs. Bot. 5: (1789) ≡ Selaginella ciliaris (Retz.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 231 (1843) ≡ Lycopodioides ciliaris (Retz.) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 睫毛异穗卷柏(新拟)
Hypopterygiopsis ciliata (Alston) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella selangorensis var. ciliata Alston, Gard. Bull. Straits Settlem. 8: 44 (1934).
Hypopterygiopsis cochleata (Hook. & Grev.) Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium cochleatum Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 2: 395, no. 153 (1831) ≡ Lycopodioides cochleata (Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 826 (1891) ≡ Selaginella cochleata (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 143, no. 55 (1843).
Hypopterygiopsis compta (Hand.-Mazz.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella compta Hand.-Mazz., Symb. Sin. 6: 9, pl. 2. (1929). 缘毛异穗卷柏(新拟)
Hypopterygiopsis coonooriana (R.D. Dixit) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella coonooriana R.D. Dixit, Bull. Bot. Surv. India 25(1–4): 223 (1983 publ. 1985).
Hypopterygiopsis coriaceifolia (X.M. Zhou, N.T. Lu & Li Bing Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella coriaceifolia X.M. Zhou, N.T. Lu & Li Bing Zhang, Phytotaxa 453(2): 125 (2020).
Hypopterygiopsis crassipes (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella crassipes Spring, Monogr. Lyc. 2: 243, no. 181 (1850) ≡ Lycopodioides crassipes (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis curtisii (Ridl.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella curtisii Ridl., J. Roy. As. Soc., Str. Br., No. 80: 148, no. 2 (1919).
Hypopterygiopsis daozhenensis (Li Bing Zhang, Q.W. Sun & Jun H. Zhao) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella daozhenensis Li Bing Zhang, Q.W. Sun & Jun H. Zhao, Phytotaxa 207(2): 187 (2015). 道真异穗卷柏(新拟)
Hypopterygiopsis darmandvillei (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella darmandvillei Alderw., Bull. Jard. Bot. Buitenz. II, 1: 23–24 (1911).
Hypopterygiopsis decipiens (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella decipiens Warb., Monsunia 1: 127 (1900). 拟大叶异穗卷柏(新拟)
Hypopterygiopsis densiciliata (X.M. Zhou, Liang Zhang & Bo Xu) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella densiciliata X.M. Zhou, Liang Zhang & Bo Xu, PhytoKeys 227: 135–149 (2023). 密毛异穗卷柏(新拟)
Hypopterygiopsis devolii (H.M. Chang, P.F. Lu & W.L. Chiou) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella devolii H.M. Chang, P.F. Lu & W.L. Chiou, Blumea 56(1): 21 (2011). 棣氏异穗卷柏(新拟)
Hypopterygiopsis dianzhongensis (X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella dianzhongensis X.C. Zhang, PhytoKeys 118: 77 (2019). 滇中异穗卷柏(新拟)
Hypopterygiopsis drepanophylla (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella drepanophylla Alston, J. Bot. 70(831): 66–67 (1932). 镰叶异穗卷柏(新拟)
Hypopterygiopsis dulongjiangensis (W.M. Chu) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella effusa var. dulongjiangensis W.M. Chu, Fl. Yunnan. 20: 718 (2006). 独龙江异穗卷柏(新拟)
Hypopterygiopsis effusa (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella effusa Alston, J. Bot. 70: 65, no. 9 (1932). 疏松异穗卷柏(新拟)
Hypopterygiopsis elegantissima (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella elegantissima Warb., Monsunia 1: 111, no. 175 (1900).
Hypopterygiopsis eschscholzii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella eschscholzii Hieron., Elmer, Leafl. Phil. Bot. 6: 2041, no. 26 (1913).
Hypopterygiopsis exasperata (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella exasperata Warb., Monsunia 1: 109, 126, no. 139 (1900).
Hypopterygiopsis firmula (A. Braun ex Kuhn) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella firmula A. Braun ex Kuhn; Verh. K. K. Zool.-Bot. Ges. Wien 19: 585, no. 132 (1869).
Hypopterygiopsis goudotiana (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella goudotiana Spring, Bull. Acad. Roy. Soc. Bruxelles 8 (12): 140 (1841) ≡ Lycopodioides goudotiana (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis heterostachys (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella heterostachys Baker, J. Bot. 23: 177, no. 264 (1885) ≡ Lycopodioides heterostachya (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 异穗卷柏.
Hypopterygiopsis hildebrandtii (A. Braun ex Kuhn) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hildebrandtii A. Braun ex Kuhn, v. Decken, Reis. III, 3, Bot.: 71, no. 88 (1879).
Hypopterygiopsis hollrungii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hollrungii Hieron. Engl. Bot. Jahrb. 50: 35, no. 14 (1913).
Hypopterygiopsis intertexta (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella intertexta Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 233, no. 156 (1843).
Hypopterygiopsis jainii (R.D. Dixit) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella jainii R.D. Dixit, Bull. Bot. Surv. India 25: 225, f. 2 A–H (1985).
Hypopterygiopsis kaernbachii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kaernbachii Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 700, no. 335 (1901 publ. 1902).
Hypopterygiopsis kalbreyeri (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kalbreyeri Baker, J. Bot. 22: 276, no. 157 (1884).
Hypopterygiopsis kanehirae (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kanehirae Alston, J. Bot. 72: 227 (1934).
Hypopterygiopsis keralensis (R.D. Dixit) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella keralensis R.D. Dixit, Bull. Bot. Surv. India 27 (1–4): 123–124. 1987 (“1986”).
Hypopterygiopsis ketra-ayam (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella ketra-ayam Alderw., Bull. Jard. Bot. Buitenz. II, 1: 24–25 (1911).
Hypopterygiopsis kivuensis (Bizzarri) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kivuensis Bizzarri, Bull. Jard. Bot. Belg. 53(1–2): 174 (1983).
Hypopterygiopsis kouytcheensis (H. Lév.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kouytcheensis H. Lév., Fedde, Repert. 9 (222–226): 451 (1911). 贵州异穗卷柏(新拟)
Hypopterygiopsis kurzii (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kurzii Baker, J. Bot. 23: 249, no. 283 (1885) ≡ Lycopodioides kurzii (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 缅甸异穗卷柏(新拟)
Hypopterygiopsis labordei (Hieron. ex Christ) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella labordei Hieron. ex Christ, Bull. Ac. Inst. Géogr. Bot. III, 11: 272, no. 168 (1902) ≡ Lycopodioides labordei (Hieron. ex Christ) H.S. Kung, Fl. Sichuan. 6: 77–78 (1988). 细叶异穗卷柏(新拟)
Hypopterygiopsis lanceolata (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lanceolata Warb., Monsunia 1: 108, 123, no. 117 (1900).
Hypopterygiopsis lauterbachii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lauterbachii Hieron., Bot. Jahrb. Syst. 50: 31, no. 12 (1913).
Hypopterygiopsis laxa (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella laxa Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 233, no. 153 (1843) ≡ Lycopodioides laxa (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis leoneensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella leoneensis Hieron., Engl. & Prantl, Nat. Pfl. 1(4): 697(1901 publ. 1902).
Hypopterygiopsis leptophylla (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella leptophylla Baker, J. Bot. 23 (269): 157 (1885) ≡ Lycopodioides leptophylla (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 膜叶异穗卷柏(新拟)
Hypopterygiopsis lewalleana (Bizzarri) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lewalleana Bizzarri, Bull. Jard. Bot. Belg. 51(1–2): 222 (1981).
Hypopterygiopsis lindhardtii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lindhardtii Hieron., Bull. Herb. Boiss. II, 5: 723, no. 99 (1905).
Hypopterygiopsis llanosii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella llanosii Hieron., Elmer, Leafl. Phil. Bot. 6: 2039, no. (1913).
Hypopterygiopsis longiciliata (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella longiciliata Hieron., Engl., Bot. Jahrb. 50: 2, 33, no. 13 (1913).
Hypopterygiopsis loriai (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella loriai Hieron., Engl. Bot. Jahrb. 50: 2, 27, no. 10 (1913).
Hypopterygiopsis lutchuensis (Koidz.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lutchuensis Koidz., Pl. Nov. Amamio-Oshimensis, Ins. Adjasc., 4 (1928), et Acta Phytotax. Geobot. 1: 165 (1932). 琉球异穗卷柏(新拟)
Hypopterygiopsis macroblepharis (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym:Selaginella macroblepharis Warb., Monsunia 1: 108, 124, no. 126, t. 3C (1900).
Hypopterygiopsis mannii (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella mannii Baker, J. Bot. 23: 180, no. 277 (1885) ≡ Lycopodioides mannii (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis marinii (Stefanović & Rakotondr.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella marinii Stefanović & Rakotondr., Novon 6(2): 205 (1996).
Hypopterygiopsis megaphylla (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella megaphylla Baker, J. Bot. 23: 180 (1885) ≡ Lycopodioides megaphylla (Baker) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 宽叶异穗卷柏(新拟)
**Hypopterygiopsis menziesii (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium menziesii Hook. & Grev., Enum. Fil. 1: 2, no. 131 (1831) ≡ Selaginella menziesii (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 227, no. 109 (1843) ≡ Lycopodioides menziesii (Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 826 (1891) ≡ Selaginella arbuscula var. menziesii (Hook. & Grev.) Skottsb., Medd. Göteborgs Bot. Trädgard, 15: 145 (1944).
**Hypopterygiopsis miniatospora (**Dalzell) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium miniatosporum Dalzell, J. Bot. (Hook.) 4: 114 (1852) ≡ Selaginella miniatospora (Dalzell) Baker, J. Bot. 23: 249 (1885) ≡ Lycopodioides miniatospora (Dalzell) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis minutifolia (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella minutifolia Spring, Monogr. Lyc. 2: 239, no. 176 (1850) ≡ Lycopodioides minutifolia (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891). 小叶异穗卷柏(新拟)
Hypopterygiopsis mittenii (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella mittenii Baker, J. Bot. (London) 21: 18 (1883).
Hypopterygiopsis molleri (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella molleri Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 697, no. 292. (1901 publ. 1902).
Hypopterygiopsis molliceps (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella molliceps Spring, Mém. Acad. Roy. Sci. Belg. 24: 257 (1850) ≡ Lycopodioides molliceps (Spring) Kuntze, Rev. Gen. Pl. 1: 826 (1891).
Hypopterygiopsis monodii (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella monodii Alston, Bull. Inst. Franc. Afr. Noire, A, 21: 440 (1959).
Hypopterygiopsis monospora (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium monosporum (Spring) Hook., Bot. Misc. 9: 362 (1857) ≡ Selaginella monospora Spring, Mém. Acad. Roy. Sci. Belg. 24: 135 (1850) ≡ Selaginella plumosa var. monospora (Spring) Baker, J. Bot. 21: 145 (1883). 单子异穗卷柏(新拟)
Hypopterygiopsis morganii (Zeiller) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella morganii Zeiller, Bull. Soc. Bot. France 32: 78 (1985).
**Hypopterygiopsis myosuroides (**Kaulf.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium myosuroides Kaulf., Enum. Fil. 19 (1824) ≡ Selaginella myosuroides (Kaulf.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 232, no. 150 (1843) ≡ Lycopodioides myosuroides (Kaulf.) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis nairii (R.D. Dixit) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella nairii R.D. Dixit, Bull. Bot. Surv. India 26: 106 (1985).
Hypopterygiopsis namdaphaensis (Sarn. Singh & Panigrahi) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella namdaphaensis Sarn. Singh & Panigrahi, Ferns & Fern-Allies of Arunachal Pradesh 1: 64. (2005).
**Hypopterygiopsis nana (**Desv.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium nanum Desv. ex Poir., Lam., Encycl. suppl. 3: 554, no. 96. (1813 publ. 1814) ≡ Selaginella nana (Desv.) Spring, Bull. Acad. Roy. Sci. Bruxelles 10: 232, no. 151 (1843) ≡ Lycopodioides nana (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis nayarii (R.D. Dixit) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella nayarii R.D. Dixit, Bull. Bot. Surv. India 27(1–4): 123 (1987).
Hypopterygiopsis neocaledonica (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella neocaledonica Baker, J. Bot. 22: 245, no. 143 (1884) ≡Lycopodioides neocaledonica (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis nummularia (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella nummularia Warb., Monsunia 1: 108, 123, no. 121 (1900).
**Hypopterygiopsis ornata (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium ornatum Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 3: 108 (1883) ≡ Selaginella ornata (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 232 (1843) ≡ Selaginella brachystachya var. ornata (Hook. & Grev.) Baker, J. Bot. 23: 180 (1885). 微齿异穗卷柏(新拟)
Hypopterygiopsis panchghaniana (R.D. Dixit) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella panchghaniana R.D. Dixit, Bull. Bot. Surv. India 25: 226 (1985).
Hypopterygiopsis panigrahii (R.D. Dixit) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella panigrahii R.D. Dixit, Bull. Bot. Surv. India 25: 226, f. 4. A–H (1985).
Hypopterygiopsis parachrysocaulos (M.H. Zhang & X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella parachrysocaulos M.H. Zhang & X.C. Zhang, Taxon 71(6): 1167 (2022). 拟块茎异穗卷柏(新拟)
Hypopterygiopsis perpusilla (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella perpusilla Baker, J. Bot. 23: 292 (1885) ≡ Lycopodioides perpusilla (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis phanotricha (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella phanotricha Baker, J. Bot. 23: 156, no. 261 (1885) ≡ Lycopodioides phanotricha (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis philippina (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella philippina Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 140 (1843) ≡ Lycopodioides philippina (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis pricei (B.C. Tan & Jermy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pricei B.C. Tan & Jermy, Fern Gaz. 12(3): 170 (1981).
**Hypopterygiopsis proniflora (**Lam.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium proniflorum Lam., Encycl. 3: 652, no. 25. (1789 publ. 1791) ≡ Selaginella proniflora (Lam.) Baker, J. Bot. 22: 156 (1885) ≡ Stachygynandrum proniflorum (Lam.) P. Beauv., Prodr. aethéogam. 110 (1805) ≡ Lycopodioides proniflora (Lam.) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis protensa (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella protensa Alston, Mém. Inst. Fr. Afr. Noire 50: 41 (1957).
Hypopterygiopsis qingchengshanensis (Li Bing Zhang & X.M. Zhou) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella qingchengshanensis Li Bing Zhang & X.M. Zhou, Phytotaxa 522(4): 286 (2021). 青城山异穗卷柏(新拟)
**Hypopterygiopsis radicata (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium radicatum Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 2: 397, no. 160 (1831) ≡ Selaginella radicata (Hook. & Grev.) Spring, Mém. Acad. Roy. Sci. Belg. 24 (2) 114 (1850) ≡ S. plumosa var. radicata (Hook. & Grev.) Warb., Monsunia 1: 102 (1900).
Hypopterygiopsis raiateensis (J.W. Moore) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella raiateensis J.W. Moore, Bull. Bernice P. Bishop Mus. 102: 15 (1933).
Hypopterygiopsis rajasthanensis (Gena, Bhardwaja & A.K. Yadav) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rajasthanensis Gena, Bhardwaja & A.K. Yadav, Am. Fern J. 69: 119 (1979).
Hypopterygiopsis rasoloheryi (Rakotondr.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rasoloheryi Rakotondr., Candollea 71(1): 144 (2016).
Hypopterygiopsis reineckei (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella reineckei Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 678, no. 83. (1901 publ. 1902).
**Hypopterygiopsis repanda (**Desv. & Poir.) Spring Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium repandum Desv. ex Poir., Lam., Encycl. suppl. 3: 558. (1813 publ. 1814) ≡ Selaginella repanda (Desv. & Poir.) Spring, Gaudich., Voy. Bonite Bot. 1: 329 (1846). 高雄异穗卷柏(新拟)
Hypopterygiopsis reptans Sakurai, Bot. Mag. (Tokyo) 57: 255 (1943) Basionym: Selaginella sakuraii H. Miller, Taxon 16: 70 (1967).
**Hypopterygiopsis reticulata (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium reticulatum Hook. & Grev., J. Bot. (Hook.) Kew Misc. 2: 402 (1831) ≡ Selaginella reticulata (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 233 (1843).
Hypopterygiopsis robinsonii (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella robinsonii Alderw., Phil. J. Sci. Bot., 11C: 118 (1916).
Hypopterygiopsis roesickeana (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella roesickeana Hieron., Engl. Bot. Jahrb. 56: 235, no. 36 (1920).
Hypopterygiopsis sambiranensis (Stefanov. & Rakotondr.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sambiranensis Stefanov. & Rakotondr., Adansonia ser. 3, 3: 19(1): 166, f. 1–2 (1997).
Hypopterygiopsis scabrida (Ridl.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella scabrida Ridl., J. Roy. As. Soc., Str. Br., No. 80: 159, no. 32 (1919) ≡ Selaginella alutacea var. scabrida (Ridl.) Alston, Gard. Bull. Str. Settl. 8: 56 (1934).
Hypopterygiopsis schefferi (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella schefferi Hieron., Engl. Bot. Jahrb. 50: 2, 24, no. 8 (1913).
Hypopterygiopsis selangorensis (Bedd. ex Ridl.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella selangorensis Bedd. ex Ridl., J. As. Soc., Str. Br., No. 80: 148, no. 3 (1919).
Hypopterygiopsis sespillifolia (Brownlie) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sespillifolia Brownlie, Fl. Nouv.-Caléd. 3: 32 (1969).
Hypopterygiopsis setchellii (O.C. Schmidt) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella setchellii O.C. Schmidt, Fedde, Repert, 20: 158 (1924).
Hypopterygiopsis sichuanica (H.S. Kung) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sichuanica H.S. Kung, Acta Bot. Yunnan. 3(2): 252 (1981). 四川异穗卷柏(新拟)
Hypopterygiopsis singalanensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella singalanensis Hieron., Hedwigia 50: 18, no. 12 (1910).
Hypopterygiopsis societatis (J.W. Moore) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella societatis J.W. Moore; Bull. Bernice P. Bishop Mus. 102: 13 (1923).
Hypopterygiopsis spinulosovena (G.Q. Gou & P.S. Wang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella spinulosovena G.Q. Gou & P.S. Wang, Acta Bot. Yunnan. 27(2): 145–146 (2005). 刺脉异穗卷柏(新拟)
Hypopterygiopsis squarrosa (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella squarrosa Baker, J. Bot. 23: 180, no. 276 (1885).
Hypopterygiopsis stolleana (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella stolleana Hieron., Engl. Bot. Jahrb. 56: 236, no. 37 (1920).
Hypopterygiopsis strigosa (Bedd.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella strigosa Bedd., Kew Bull., 192, no. 600 (1911).
Hypopterygiopsis subcordata (A. Braun ex Kuhn) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subcordata A. Braun ex Kuhn; Fil. Afr. 193 (1868) ≡ Lycopodioides subcordata (A. Braun) Kuntze, Rev. Gen. Pl. 1: 827 (1891) ≡ Stachygynandrum subcordatum (A. Braun) Carruth., Cat. Afr. Pl. coll. by Dr. F. Welwitsch in 1853–1861, 2(2): 262 (1901).
Hypopterygiopsis subeffusa (Shalimov et X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subeffusa Shalimov & X.C. Zhang, Turczaninowia 25(3): 58 (2022). 近疏松异穗卷柏(新拟)
**Hypopterygiopsis subdiaphana (**Wall. ex Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium subdiaphanum Wall., (Cat. n. 136 (1829), nom. nud.) ex Hook. & Grev., Enum. Fil. J. Bot. (Hook.) Kew Misc. 2: 401, no. 183 (1831) ≡ Selaginella subdiaphana (Wall. ex Hook. & Grev.) Spring, Bull. Acad. Roy. Sci. Bruxelles 10: 232 (1843) ≡ Lycopodioides subdiaphana (Wall. ex Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 825 (1891). 近透明异穗卷柏(新拟)
Hypopterygiopsis subisophylla (Jermy) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subisophylla Jermy, Brit. Fern Gaz. 10: 30, f. 1–9 (1968).
Hypopterygiopsis subchaetoloma (X.M. Zhou, Li Bing Zhang & Z.R. He) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subchaetoloma X.M. Zhou, Li Bing Zhang & Z.R. He, Phytotaxa 603(3): 225 (2023). 近毛边异穗卷柏(新拟)
Hypopterygiopsis submonospora (Shalimov & X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella submonospora Shalimov & X.C. Zhang, Turczaninowia 25(1): 154 (2022). 拟单子异穗卷柏(新拟)
Hypopterygiopsis subspinulosa (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subspinulosa Spring, Miq. Pl. Jungh. 3: 277, no. 10 (1854).
Hypopterygiopsis subvaginata (X.C. Zhang & Shalimov) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subvaginata X.C. Zhang & Shalimov, J. Sp. Res. 9(3): 222 (2020). 拟鞘舌异穗卷柏(新拟)
Hypopterygiopsis tectissima (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tectissima Baker, J. Bot. 22: 89, no. 119 (1884).
Hypopterygiopsis temehaniensis (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella temehaniensis J.W. Moore; Bull. Bernice P. Bishop Mus. 102: 13 (1933).
**Hypopterygiopsis tenera (**Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium tenerum Hook. & Grev., J. Bot. (Hook.) Kew Misc. 2: 400 (1831) ≡ Selaginella tenera (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 232 (1843) ≡ Lycopodioides tenera (Hook. & Grev.) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis tenerrima (A. Braun ex Kuhn) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tenerrima A. Braun ex Kuhn, Fil. Afr. 193 (1868) ≡ Lycopodioides tenerrima (A. Braun) Kuntze, Rev. Gen. Pl. 1: 827 (1891) ≡ Stachygynandrum tenerrimum (A. Braun) Carruth., Cat. Afr. Pl. coll. by Dr. F. Welwitsch in 1853–1861, 2(2): 262 (1901).
Hypopterygiopsis tenuifolia (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tenuifolia Spring, Mém. Acad. Roy. Sci. Belg. 24(2): 253 (1850) ≡ Lycopodioides tenuifolia (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis thomensis (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella thomensis Alston, Exell, Cat. Vasc. Pl. S. Tomé, 97, f. 3 (1944).
Hypopterygiopsis trichophylla (K.H. Shing) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella trichophylla K.H. Shing, Vasc. Pl. Hengduan Mount. 1: 11 (1993), without Latin descr. et Acta Phytotax. Sin. 31(6): 569 (1993) ≡ Selaginella monospora ssp. trichophylla (K.H. Shing) X.C. Zhang, Fl. Reipubl. Popularis Sin. 6(3): 189 (2004). 毛叶异穗卷柏(新拟)
Hypopterygiopsis unilateralis (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella unilateralis Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 232, no. 140 (1843) ≡ Lycopodioides unilateralis (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis usta (Vieill. ex Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym:Selaginella usta Vieill. ex Baker, Jour. Bot. 23: 23, no. 212. 1885.
Hypopterygiopsis vanderystii (Bizzarri) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym:Selaginella vanderystii Bizzarri, Bull. Jard. Bot. Belg. 53(1–2): 171 (1983).
Hypopterygiopsis vaginata (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella vaginata Spring, Mém. Acad. Roy. Sci. Belg. 24: 87 (1850) ≡ Lycopodioides vaginata (Spring) Kuntze, Rev. Gen. Pl. 1: 827 (1891). 鞘舌异穗卷柏(新拟)
Hypopterygiopsis vieillardii (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella vieillardii Warb., Monsunia 1: 125, no. 138 (1900).
Hypopterygiopsis wangpeishanii (Li Bing Zhang, H. He & Q.W. Sun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella wangpeishanii Li Bing Zhang, H. He & Q.W. Sun, Phytotaxa 164(3): 195 (2014). 培善异穗卷柏(新拟)
Hypopterygiopsis wattii (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella wattii Baker, Handb. Fern-Allies 109 (1887) ≡ Lycopodioides wattii (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis weinlandii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella weinlandii Hieron., Engl. Bot. Jahrb. 50: 21, 29, no. 11 (1913).
Hypopterygiopsis whitmeei (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella whitmeei Baker, J. Bot. 23: 24, no. 215 (1885) ≡ Lycopodioides whitmeei (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891).
Hypopterygiopsis wuyishanensis (K.W. Xu, X.M. Zhou & Y.F. Duan) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella wuyishanensis K.W. Xu, X.M. Zhou & Y.F. Duan, PhytoKeys 202: 111 (2022). 武夷山异穗卷柏(新拟)
Hypopterygiopsis xichouensis (W.M. Chu) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella xichouensis W.M. Chu, Fl. Yunnan. 20: 720 (2006). 西畴异穗卷柏(新拟)
Hypopterygiopsis xipholepis (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella xipholepis Baker, J. Bot. 23: 155, no. 258 (1885) ≡ Lycopodioides xipholepis (Baker) Kuntze, Rev. Gen. Pl. 1: 827 (1891). 剑叶异穗卷柏(新拟)
Hypopterygiopsis xishuiensis (G.Q. Gou & P.S. Wang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella xishuiensis G.Q. Gou & P.S. Wang, Acta Phytotax. Sin. 43(1): 71 (2005). 习水异穗卷柏(新拟)
Hypopterygiopsis yunckeri (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella yunckeri Alston, Bull. Bernice P. Bishop Mus. 220: 44 (1959).
Hypopterygiopsis zechii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella zechii Hieron., Engl. & Prantl, Nat. Pfl. 1(4): 697, no. 298 (1901 publ. 1902).
Hypopterygiopsis zhangii (S.Y. Dong) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella zhangii S.Y. Dong, Syst. Bot. 47(1): 91 (2022). 宪春异穗卷柏(新拟)
Hypopterygiopsis zollingeriana (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella zollingeriana Spring, Miq., Pl. Jungh. 3: 278, no. 11 (1854).
5.7.3. Lycopodioides
Lycopodioides Boehm., Def. Gen. Pl. (ed. 3). 485. 1760. 疏穗卷柏属(新拟) – Isonym: Lycopodioides Kuntze, Revis_._ Gen. Pl. 2: 824 (1891) ≡ Trispermium Hill, Gener. Nat. Hist., ed. 2, 2(Hist. Pl.): 112 (1773) – Type: Lycopodium denticulatum L., Sp. Pl. 2: 1106 (1753) ≡ Selaginella denticulata (L.) Spring, Flora 21(10): 149 (1838) ≡ Lycopodioides denticulata (L.) Kuntze, Revis_._ Gen. Pl. 1–2: 824 (1891).
= Diplostachyum P. Beauv., Mag. Encycl. 5: 481. (1804) (‘Diplostachium’). Lycopodioides sect. Diplostachyum (P. Beauv.) Rothm. Lycopodium sect. Diplostachyum (P. Beauv.) Rchb. ≡ Selaginella sect. Diplostachyum (P. Beauv.) T. Moore ≡ Heterophyllium Hieron. ex Börner. Lectotype (designated by Pfeiffer, Nom. 1: 1100. 1874): Lycopodium helveticum L., Sp. Pl. 2: 1104 (1753) ≡ Bernhardia helvetica (L.) Gray, Nat. Arr. Brit. Pl. 2: 23 (1821) ≡ Selaginella helvetica (L.) Spring, Flora 21(1): 149 (1838) ≡ Lycopodioides helvetica (L.) Kuntze, Revis. Gen. Pl. 2: 826 (1891) ≡ Heterophyllium helveticum (L.) Börner, Fl. Deut. Volk 285 (1912).
Lycopodioides is the most frequently recognized genus in addition to Selaginella in the family, but our circumscription is different from those in earlier literature and corresponds to Selaginella sect. Homostachys sensu Zhou and Zhang (2015). The latter has been recognized in earlier classifications of Selaginella (e.g., Walton and Alston, 1938; Warburg, 1900). Walton and Alston (1938) included Selaginella rothertii, a species from high mountain in Java (Alston, 1935) as a member of S. subg. Homostachys. S. rothertii might be the southernmost species in subg. Homostachys, whereas the remaining species are distributed from East Asia to Europe (Zhang et al., 2021). Warburg (1900) recognized S. subg. Homostachys, but included S. ciliaris and S. pallidissima in it. Although S. ciliaris has creeping plant, similar to those species in subg. Homostachys, this species has dimorphic but resupinate strobili and reticulate megaspore and is a member of Hypopterygiopsis sect. Hypopterygiopsis.
This genus is characterized by creeping stems (Fig. 8G), dimorphic sporophylls (Fig. 8F and G) (except for Selaginella denticulata Spring and S. helvetica (L.) Spring with nearly monomorphic but loose sporophylls (Fig. 9E)), loose and non-resupinate (with the smaller ones in the same plane as the median leaves and the larger ones in the same plane as the ventral leaves) strobili (Fig. 8F and G). Species in Lycopodioides usually grow in wet habitat. Although megaspore surfaces with papillate, tuberculate, and verrucate ornamentation are similar to those of Hypopterygiopsis sect. Hypopterygiopsis, but the laesurae of megaspores in Lycopodioides are usually disconnected at the pole (Zhou et al., 2015; Zhou and Zhang, 2015). 2n = 18 (Reeve, 1935; Jermy et al., 1967; Loyal et al., 1984; Takamiya, 1993).
Lycopodioides known so far (Zhou et al., 2022) has large plastome size (ca. 187 kb) with the most genes (128) in Selaginellaceae. Lycopodioides has three ribosomal operon copies, and its master conformation of plastomes can mediate eight isomers (Zhou et al., 2022).
This genus contains about 12 species mainly occurring in Asia and Europe (Zhang et al., 2021).
Members:
Lycopodioides denticulata (L.) Kuntze in Revis. Gen. Pl. 2: 824. 1891. Basionym: L. denticulatum L., Sp. Pl. 2: 1106 (1753).
Lycopodioides helvetica (L.) Kuntze, Revis. Gen. Pl. 2: 826. 1891. Basionym: L. helveticum L., Sp. Pl. 2: 1104–1105 (1753). 小疏穗卷柏(新拟)
Lycopodioides jiulongensis H.S. Kung, Li Bing Zhang & X.S. Guo in Acta Bot. Yunnan. 17(4): 420, fig. 1 (1995). 九龙疏穗卷柏(新拟)
Lycopodioides laxistrobila (K.H. Shing) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella laxistrobila K.H. Shing, Acta Phytotax. Sin. 31(6): 569 (1993). 疏穗卷柏(新拟)
Lycopodioides longistrobilina (P.S. Wang, X.Y. Wang & Li Bing Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella longistrobilina P.S. Wang, X.Y. Wang & Li Bing Zhang, Novon 22: 260. 2012. 长穗疏穗卷柏(新拟)
Lycopodioides nipponica (Franch. & Sav.) Kuntze, Revis. Gen. Pl. 2: 827. 1891 (‘nipponicum’). Basionym: Selaginella nipponica Franch. & Sav., Enum. Pl. Jap. 2(2): 199, 615 (1879). 伏地疏穗卷柏(新拟)
Lycopodioides pallidissima (Spring) Kuntze; Revis. Gen. Pl. 2: 827 (1891). Selaginella pallidissima Spring in Bull. Acad. Roy. Sci. Bruxelles 10: 231 (1843). 平疏穗卷柏(新拟)
Lycopodioides prostrata H.S. Kung, Fl. Sichuan. 6: 76 (1988). 地疏穗卷柏(新拟)
Lycopodioides pseudonipponica (Tagawa) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pseudonipponica Tagawa, Acta Phytotax. Geobot. 25: 177 (1973) ≡ Selaginella helvetica subsp. pseudonipponica (Tagawa) H.M. Chang, W.L. Chiou & J.C. Wang, Fl. Taiwan, Selaginellaceae 38 (2012). 拟伏地疏穗卷柏(新拟)
Lycopodioides rothertii (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rothertii Alderw., Bull. Jard. Bot. Buitenz. II, 7: 30 (1912).
Lycopodioides shensiensis (Christ) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella shensiensis Christ; Nuov. Giorn. Bot. Ital. n. s. 4: 102 (1897). 陕西疏穗卷柏(新拟)
Lycopodioides tama-montana (Seriz.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella tama-montana Seriz., J. Jpn. Bot. 53(8): 242 (1978). 高山疏穗卷柏(新拟)
5.7.4. Valdespinoa
Valdespinoa Li Bing Zhang & X.M. Zhou, gen. nov. 韦氏卷柏属(新拟) – Type: Valdespinoa douglasii (Hook. & Grev.) Li Bing Zhang & X.M. Zhou (≡ Selaginella douglasii (Hook. & Grev.) Spring).
Etymology: In honor of Prof. Iván A. Valdespino currently based at PMA for his contributions to the study of Selaginella (e.g., Valdespino, 1993a, 2015, 2016, 2017b, 2020; Valdespino et al., 2015, 2018a, b).
Valdespinoa corresponds to Selaginella sect. Auriculate sensu Zhou and Zhang (2015).
Valdespinoa is sister to Lycopodioides with strong support. However, Valdespinoa has monomorphic sporophylls and is only distributed in America. We recognize the monospecific Valdespinoa containing V. douglasii only. Hieronymous and Sadebeck (1902) recognized the Douglasii group which contains S. accharata, S. delicatissma, S. douglasii, and S. reflexa. Phylogenetic analyses showed that S. reflexa is member of our Selaginella s.s (Weststrand and Korall, 2016a; Zhou et al., 2016). Based on the morphology, S. delicatissma (including S. accharata) might be members of Valdespinoa, but more studies are necessary.
Valdespinoa has the largest plastome size (>187 kb) known in Selaginellaceae. Its plastome has three ribosomal operon copies and its master conformation can mediate 24 isomers (Zhou et al., 2022).
Valdespinoa currently contains V. douglasii in USA only (Valdespino, 1993a).
Member:
Valdespinoa douglasii (Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium douglasii Hook. & Grev. Bot. Misc. 2: 396 (1831) ≡ Selaginella douglasii (Hook. & Grev.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 138 (1843).
5.8. Subfamily VII. Selaginelloideae
Subfamily VII. Selaginelloideae Li Bing Zhang & X.M. Zhou, subfam. nov. 同穗卷柏亚科(新拟) – Type: Selaginella P. Beauv.
Plants erect, suberect, ascending, or creeping, rarely rosette-forming, with ventral rhizophores borne on ventral side of stems and/or branches; sterile leaves dimorphic, decussate; sporophylls monomorphic or dimorphic (resupinate); megaspore surfaces reticulate (Stachygynandrum type with low and open muri; Fig. 2Z), rarely verrucate; microspore surfaces often baculate, sometimes verrucate to blunt echinate; 2n = 18, 20, 22, 36, 36–40, or 50–60 (Jermy et al., 1967; Love and Love, 1976; Takamiya, 1993; Marcon et al., 2005).
This subfamily corresponds to Selaginella subg. Stachygynandrum sensu Zhou and Zhang (2015).
Morphologically, Selaginelloideae are characterized by creeping, ascending or erect (never scandent) plants (Figs. 10A–F and 11A–D), monomorphic sporophylls (Fig. 10, Fig. 11B, E, F) (but including all these species with dimorphic or subdimorphic sporophylls in new world) (Fig. 11G and H). Megaspores usually have more or less a slightly developed zona at the end of laesurae (Figs. 10K–N) or obviously equatorial flange (e.g., Kungiselaginella involvens: Fig. 11I) and surfaces with reticulate ornamentation usually with often open mesh (Figs. 10I, K–N and 11J, M−P). Microspore surfaces are usually baculate (Fig. 10J, O–R), blunt spiny (Fig. 10 Q–R), papilary (K–L), or verrucate (Fig. 10S and T) (Valdespino, 2015, 2016, 2017b, c, 2020; Valdespino et al., 2015, 2018b; Zhou et al., 2015).
Fig. 11.

Morphology of Kungiselaginella 孔氏卷柏属 and Selaginella s.s. 同穗卷柏属 in subfamily Selaginelloideae (Selaginella subg. Stachygynandrum; Zhou and Zhang, 2015). A, F, I and K. Kungiselaginella involvens (A. Habit; F. Strobilus; I. Megaspore; K. Microspore); B and C. K. davidii (B. Strobili; C. Habit); D, E, J, L. K. moellendorffii (D. Habit; E. Strobilus; J. Megaspore; L. Microspore); G, H, M and Q. Selaginella flagellata (G. Dorsal side of strobilus; H. Ventral side of strobilus; M. Megaspore; Q. Microspore); N & R. S. porphyrospora (N. Megaspore; R. Microspore); O and S. S. pallescens (O. Megaspore; S. Microspore); P & T. S. apoda (P. Megaspore; T. Microspore).
Following our current classification, nearly all New World species with ventral rhizophores, except Didiclis hoffmannii, Pulviniella, and Valdespinoa douglasii, should be members of this subfamily. Selaginelloideae are mainly pantropical distribution, and some species can extend to subtropical and temperate Asia and Americas (Valdespino, 1993a; Zhang et al., 2013).
In our previous analysis (Zhou et al., 2015, 2016) and classification (Zhou and Zhang, 2015), megaspore data have been well explored and used and their taxonomic significance was intensively discussed (Korall and Taylor, 2006; Zhou and Zhang, 2015; Zhou et al., 2015, 2016; Weststrand and Korall, 2016a). In contrast, the microspore morphology of Selaginelloideae was rarely observed and reported except that of the Old World (especially Asian) species (e.g., Zhou et al., 2015). In recent years, as a large number of new species from the New World have been described and microspore data have been provided by Valdespino and co-authors (e.g., Valdespino, 2015, 2016, 2017b, c; 2020; Valdespino et al., 2015, 2018b), we now know that New-World species have more diverse microspore surfaces than those of the Old-World species (Zhou et al., 2015; unpublished data).
Jermy (1986) defined Selaginella subg. Stachygynandrum as containing only those species with monomorphic sporophylls, but phylogenetic analyses showed that some species with dimorphic strobili in the New World are also mixed with species with monomorphic sporophylls in the tree and the dimorphism of sporophylls have evolved 10 times (Zhou et al., 2016; Weststrand and Korall, 2006a; our Fig. 2G). Based on previous studies and our observations, species with dimorphic sporophylls from the New World also have same reticulate ornamentation as those species with monomorphic sporophylls in S. subg. Stachygynandrum sensu Jermy (1986) [e.g., Selaginella hyalogramma_ Valdespino (Valdespino, 2017a) S. mucronata (Valdespino et al., 2015); S. pellucidopunctata_ Valdespino (Valdespino et al., 2015); and our observations for S. flagellata and S. simplex_]. These are entirely different from species with dimorphic sporophylls in Lycopodioides and Didiclis which do not occur in the New World except D. hoffmannii. The dimorphism of sporophylls in those species from the Old World and that in those from the New World independently evolved (Fig. 2G). Evidence so far showed that the New-World species with dimorphic sporophylls are typically reticulate on megaspores and baculate on microspores._ It is clear that reticulate megaspores and baculate or verrucate microspores are diagnosing features of Selaginelloideae newly defined here.
In addition, tuberculate or verrucate microspores were also observed in some New-World species in the subfamily [(e.g., _Selaginella pubimarginata_ Valdespino (2020); S. brigitteana Valdespino (Valdespino, 2019); S. neospringiana Valdespino (2015c); S. mucugensis Valdespino; S. saltuicola Valdespino; S. sematophylla Valdespino (Valdespino et al., 2015)]. Also, some morphological characters, e.g., dimorphic or monomorphic sporophylls, reticulate or non-reticulate megaspores, baculate or non-baculate microspores, etc., reversed and/or independently evolved many times in the New-World Selaginelloideae (Fig. 2J).
The plastomes of Selaginelloideae known so far (Zhou et al., 2022) are extremely conserved and have the same gene order. Species of Selaginelloideae have no more than two pairs of small repeats in SC and their plastomes have up to five conformations. Those of Selaginella moellendorffii and its close relatives have IR and a pair of repeats existed in SC with four conformations, whereas those of the remaining species in the subfamily have DR or DR-IR coexisting.
This subfamily contains about 330 species in three genera, Chuselaginella, Kungiselaginella, and Selaginella s.s., the former two occurring in the Old World only, and the latter in Americas only except one (a few) species in Africa. Chuselaginella and Kungiselaginella are morphologically distinguishable from each other (see below and our key) and have slightly different distributions: Asia to Pacific islands and Africa for Chuselaginella, and Asia for Kungiselaginella. K. moellendorffii (= S. moellendorffii) in America is introduced from the Old World. Selaginella s.s. is morphologically nearly indistinguishable from the other two. Alternatively, these three genera could be combined into a broadly defined Selaginella, but we prefer to separate them because of their molecular divergence (especially in nuclear phylogeny; Fig. S2), geographical coherence, and more manageable size.
5.8.1. Chuselaginella
Chuselaginella Li Bing Zhang & X.M. Zhou, gen. nov. 朱氏卷柏属(新拟) – Type: Chuselaginella alopecuroides (Baker) Li Bing Zhang & X.M. Zhou (= Selaginella alopecuroides Baker, J. Bot. British & Foreign 19: 368, no. 83. (1881)).
≡ Selaginella sect. Ascendentes (Baker) Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1136. 2015 ≡ S. ser. Ascendentes Baker, J. Bot. Lond. 21: 3 (1883) – Lectotype (designated by Zhou and Zhang, 2015: 1136): S. alopecuroides Baker.
≡ S. ser. Radicantes Warb., Monsunia 1: 102 (1900) – Lectotype (designated by Zhou and Zhang, 2015: 1136): S. alopecuroides Baker.
Etymology:—From _Chu_-, in honor of the late Prof. Wei-Ming Chu of Yunnan University (herbarium PYU), China, for his contributions to the study of ferns in general and those of Selaginella of Yunnan in particular (Chu et al., 2006).
Chuselaginella corresponds to Selaginella sect. Ascendentes sensu Zhou and Zhang (2015). This genus is supported as monophyletic in all of our analyses (Figs. 1 and S1–S3). Species of this genus have plants often ascending (a few strictly erect), microspores always baculate with expanded tips and megaspores with reticulum and a unique zonal structure at the ends of laesurae (Fig. 10K–N) (e.g., Liu et al., 1989 for Selaginella doederleinii Hieron.; Liu et al., 2001 for S. frondosa Warb.; Zhao et al., 2006 for S. commutata Alderw.; Zhou et al., 2015, for S. trichophylla K.H. Shing and S. scabrifolia Ching & Chu H. Wang; and Korall and Taylor, 2006 for additional species).
Chuselaginella contains more than 70 species from Africa, Asia, Australia, and South Pacific islands.
Members:
Chuselaginella aenea (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: _Selaginella aene_a Warb., Monsunia 1: 104, 115, no. 58 (1900).
Chuselaginella agusanensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella agusanensis Hieron., Leafl. Philipp. Bot. 6: 1998.
Chuselaginella alligans (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella alligans Hieron., Elmer, Leafl. Phil. Bot. 6: 1998, 2012, no. 15 (1913).
Chuselaginella alopecuroides (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella alopecuroides Baker, J. Bot. 19: 368, no. 83 (1881).
Chuselaginella auriculata (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella auriculata Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 142, no. 147 (1843).
Chuselaginella bluuensis (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella bluuensis Alderw., Bull. Jard. Bot. Buitenz. II, 11: 29 (1913).
Chuselaginella borneensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella frondosa var. borneensis Hieron., Bot. Jahrb. Syst. 44: 511 (1910).
Chuselaginella brachyblepharis (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella brachyblepharis Alderw., Bull. Jard. Bot. Buitenzorg, sér. 2, 11: 24 (1913)
Chuselaginella brevipes (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella brevipes A. Braun, App. Ind. Sem. Hort. Berol. 1 (1867)
Chuselaginella breynioides (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella breynioides Baker, J. Bot. 23: 45, no. 222 (1885).
Chuselaginella brooksii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella brooksii Hieron., Hedwigia 51: 252, no. 6 (1912).
Chuselaginella burkei (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella burkei Hieron., Engl. Bot. Jahrb. 50: 2, 16, no. 5 (1913).
Chuselaginella carnea (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella carnea Alderw., Bull. Jard. Bot. Buitenzorg, sér. 2, 24: 7 (1917).
Chuselaginella cesatii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella cesatii Hieron., Hedwigia 50: 6, no. 5 (1910).
Chuselaginella ciliata (Alderw.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella frondosa var. ciliata Alderw., Bull. Jard. Bot. Buitenzorg, sér. 2, 16: 48 (1914).
Chuselaginella commutata (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella commutata Alderw., Bull. Jard. Bot. Buitenz. II, 28: 46 (1918). 长芒朱氏卷柏(新拟)
Chuselaginella copelandii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella copelandii Hieron. Repert. Spec. Nov. Regni Veg. 10: 106 (1911).
Chuselaginella cumingiana (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella cumingiana Spring Bull. Acad. Roy. Soc. Bruxelles 10: 146, no. 81 (1843).
Chuselaginella cuprea (Ridl.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella cuprea Ridl.; J. Straits Branch Roy. Asiat. Soc. 80: 152, no. 14 (1919)
Chuselaginella cupressina (Willd.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium cupressinum Willd., Sp. Pl. 5: 43 (1810).
Chuselaginella dielsii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella dielsii Hieron.; Hedwigia 51: 254, no. 7 (1911 publ. 1912).
Chuselaginella distans (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella distans Warb., Monsunia 1: 106, 120, no. 86 (1900).
Chuselaginella doederleinii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella doederleinii Hieron., Hedwigia 43(1): 41–42 (1904). 深绿朱氏卷柏(新拟)
Chuselaginella dolichocentrus (K.M. Wong) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella intermedia var. dolichocentrus K.M. Wong, Gard. Bull. Singapore 35(2): 125 (1982 publ. 1983)
Chuselaginella elmeri (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella elmeri Hieron., Repert. Spec. Nov. Regni Veg. 10: 46, no. 5 (1911).
Chuselaginella fenixii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella fenixii Hieron. Repert. Spec. Nov. Regni Veg. 10: 98, no. 10 (1911).
Chuselaginella frondosa (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella frondosa Warb. Monsunia 1: 105, 117, no. 75 (1900). 繁叶朱氏卷柏(新拟)
Chuselaginella grabowskyi (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella grabowskyi Warb., Monsunia 1: 107, 122, no. 109a (1900).
Chuselaginella grandis (T. Moore) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella grandis T. Moore, Gard. Chron. 18(2): 40, t. 8 (1882).
Chuselaginella griffithii (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella griffithii Spring Bull. Acad. Roy. Soc. Bruxelles 10: 145, no. 80 (1843).
Chuselaginella guihaia (X.C. Zhang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella guihaia X.C. Zhang PhytoKeys 80: 44 (2017). 桂海朱氏卷柏(新拟)
Chuselaginella hordeiformis (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hordeiformis Baker J. Bot. 23: 47, no. 229 (1885).
Chuselaginella hosei (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hosei Hieron.; Hedwigia 51: 243, no. 3. (1911 publ. 1912).
Chuselaginella intermedia (Blume) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium intermedium Blume, Enum. Pl. Javae 2: 269, no. 20 (1830). ≡ Selaginella intermedia (Blume) Spring Bull. Acad. Roy. Soc. Bruxelles 10: 144 (1843).
Chuselaginella jagorii (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella jagorii Warb. Monsunia 1: 104, 116, no. 68 (1900).
Chuselaginella kerstingii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kerstingii Hieron. Engl. Bot. Jahrb. 50: 2, 21, no. 7 (1913).
Chuselaginella kusaiensis (Hosok.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella kusaiensis Hosok. Trans. Nat. Hist. Soc. Formosa 25 (147): 440 (1935).
Chuselaginella latifrons (Warb. non Hort. ex Williams) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella latifrons Warb. non Hort. ex Williams Monsunia 1: 106, 120, no. 89 (1900).
Chuselaginella lebongtandaiana (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lebongtandaiana Alderw. Bull. Jard. Bot. Buitenz. II, 23: 23 (1916).
Chuselaginella leveriana (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella leveriana Alston J. Bot. 77: 225 (1939).
Chuselaginella longiaristata (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella longiaristata Hieron. Hedwigia 50: 16, no. 11 (1910).
Chuselaginella longipinna (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella longipinna Warb. Monsunia 1: 105, 119 (1900).
Chuselaginella longirostris (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella longirostris Alderw. Bull. Jard. Bot. Buitenz. II, 11: 25 (1913).
Chuselaginella lonko-batu (Hieron. & Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella lonko-batu Hieron. & Alderw. Bull. Jard. Bot. Buitenz. II, 16: 42 (1914).
Chuselaginella louisiadensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella burkei var. louisiadensis Hieron., Bot. Jahrb. Syst. 50: 2, 18 (1913).
Chuselaginella luzonensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella luzonensis Hieron. Engl. & Prantl, Nat. Pfl. 1(4): 681, no. 113. (1901 publ. 1902).
Chuselaginella magnifica (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella magnifica Warb. Monsunia 1: 103, 114, no. 45, t. 3A (1900).
Chuselaginella melanesica (Kuhn) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella melanesica Kuhn, Forschungsr. Gazelle 4, Bot: 17 (1889).
Chuselaginella moszkowskii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella moszkowskii Hieron. Bot. Jahrb. Syst. 50: 2, 14, no. 4 (1913).
Chuselaginella opaca (Seriz.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella doederleinii var. opaca Seriz., J. Phytogeogr. Taxon. 30(1): 44 (1982).
Chuselaginella parvifolia (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella parvifolia Alderw.; Bull. Jard. Bot. Buitenzorg, sér. 2, 11: 30 (1913).
Chuselaginella paxii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella paxii Hieron.; Bot. Jahrb. Syst. 44: 512 (1910).
Chuselaginella pentagona (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella pentagona Spring Mém. Acad. Roy. Sci. Belg. 24(2): 150 (1850).
Chuselaginella petelotii (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella petelotii Alston H. Lecomte, Fl. Indo-Chine 7: 562, t. 66, f. 1–5 (1951).
Chuselaginella poperangensis (Hieron. ex Rech.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella poperangensis Hieron. ex Rech. Denkschr. Kaiserl. Akad. Wiss., Wien. Math.-Naturwiss. Kl. 89: 483, t.7, f.13C (1914).
Chuselaginella posewitzii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella posewitzii Hieron. Hedwigia 51: 241, no. 1. (1911 publ. 1912).
Chuselaginella procera (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella procera Alston J. Bot. 72: 227 (1934).
Chuselaginella puberulipes (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella puberulipes Alderw., Nova Guinea, Rés. d. Expéd. á Nouv. Guin. 14 (Bot.): 66 (1924).
Chuselaginella quadrivenulosa (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella quadrivenulosa Alderw. Nova Guinea, Rés. d. Expéd. á Nouv. Guin. 14 (Bot.): 65 (1924).
Chuselaginella ramosii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella ramosii Hieron. Repert. Spec. Nov. Regni Veg. 10: 52, no. 8 (1911).
Chuselaginella rivalis (Ridl.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rivalis Ridl. Bull. Misc. Inform. Kew 1924: 266 (1924).
Chuselaginella rolandi-principis (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rolandi-principis Alston J. Bot. 72: 228–229 (1934). 海南朱氏卷柏(新拟)
Chuselaginella roxburghii ((Hook. & Grev.) Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella roxburghii (Hook. & Grev.) Spring Bull. Acad. Roy. Soc. Bruxelles 10: 228, no. 115 (1843).
Chuselaginella rugulosa (Ces.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella rugulosa Ces. Felci Borneo, Atti della R. Acad. Sc. 7 (8): 35 (1876).
Chuselaginella sambasensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sambasensis Hieron. Hedwigia 50: 9, no. 7 (1910).
Chuselaginella sarawakensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sarawakensis Hieron. Hedwigia 50: 13, no. 10 (1910).
Chuselaginella scabrifolia (Ching & Chu H. Wang) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella scabrifolia Ching & Chu H. Wang Acta Phytotax. Sin. 8(2): 157 (1959). 糙叶朱氏卷柏(新拟)
Chuselaginella schatteburgiana (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella schatteburgiana Hieron. Engl. Bot. Jahrb. 56: 229, no. 29 (1920).
Chuselaginella sepikensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella sepikensis Hieron. Bot. Jahrb. Syst. 56: 243, no. 58 (1920).
Chuselaginella spanielema (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella spanielema Alston, J. Bot. 72: 229 (1934) (spanioclema, sphalm.)
Chuselaginella strigosa (Ridl.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella trichobasis var. strigosa Ridl., J. Straits Branch Roy. Asiat. Soc. 80: 153 (1919). ≡ Selaginella roxburghii var. strigosa (Ridl.) K.M. Wong, Gard. Bull. Singapore 35(2): corrigendum slip [131 (1982 publ. 1983).
Chuselaginella subalpina (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subalpina Alderw. Bull. Jard. Bot. Buitenz. II, 20: 26 (1915).
Chuselaginella subcalcarata (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subcalcarata Alderw. Bull. Jard. Bot. Buitenz. II, 16: 49 (1914).
Chuselaginella subpedalis (Alderw.) Li Bing Zhang & X.M. Zhou, comb. & stat. nov. Basionym: Selaginella paxii var. subpedalis Alderw., Bull. Jard. Bot. Buitenzorg, sér. 2, 11: 30 (1913).
Chuselaginella subserpentina (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella subserpentina Alderw. Bull. Jard. Bot. Buitenz. II, 1: 17 (1911).
Chuselaginella suffruticosa (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella suffruticosa Alderw.; Bull. Jard. Bot. Buitenzorg, sér. 2, 1: 22 (1911).
Chuselaginella superba (Alston) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella superba Alston, J. Bot. 70: 63–64, no. 4, t. 600, f. A–F (1932). 粗茎朱氏卷柏(新拟)
Chuselaginella thurnwaldiana (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella thurnwaldiana Hieron. Engl. Bot. Jahrb. 56: 227, no. 28 (1920).
Chuselaginella trachyphylla (A. Braun) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella trachyphylla A. Braun Sitz. Ges. Naturf. Freunde Berl. Berl. 8 (1863). 粗叶朱氏卷柏(新拟)
Chuselaginella versicolor (Spring) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella versicolor Spring Bull. Acad. Roy. Soc. Bruxelles 10: 143, no. 57 (1843).
**Chuselaginella vestita (**Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella vestita Alderw. Bull. Jard. Bot. Buitenz. II, 28: 54 (1918).
Chuselaginella vonroemeri (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella vonroemeri Alderw. Bull. Jard. Bot. Buitenz. II, 24: 7 (1917).
Chuselaginella wariensis (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella wariensis Hieron. Engl. Bot. Jahrb. 50: 2, 19, no. 6 (1913).
Chuselaginella zahnii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella zahnii Hieron. Engl. Bot. Jahrb. 50: 2, 37, no. 15 (1913).
5.8.2. Kungiselaginella
Kungiselaginella Li Bing Zhang & X.M. Zhou, gen. nov. 孔氏卷柏属(新拟) – Type: Kungiselaginella involvens (Sw.) Li Bing Zhang & X.M. Zhou (= Lycopodium involvens Sw., Syn. Fil. 182, no. 50 (1806)).
≡ Selaginella sect. Circinatae (Hook. & Grev.) Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1136. 2015 ≡ Lycopodium (unranked) Circinatae Hook. & Grev., Bot. Misc. 2: 380. 1831 ["Circinata"] ≡ S. (unranked) Circinatae (Hook. & Grev.) Spring in Mart., Fl. Bras. 1(2): 118 (1840) – Lectotype (designated by Zhou and Zhang, 2015: 1136): S. involvens (Sw.) Spring.
≡ S. (unranked) Caulescentes A. Braun, App. Ind. Sem. Hort. Berol. 11. [1857] 1858 ≡ S. ser. Caulescentes (A. Braun) Baker, J. Bot. Lond. 21: 3 (1883) – Type: S. caulescens Spring [= S. involvens (Sw.) Spring].
≡ S. (unranked) Rosulatae A. Braun, App. Ind. Sem. Hort. Berol. 11 ([1857] 1858), nom. illeg. ≡ S. ser. Rosulatae (A. Braun) Baker, J. Bot. Lond. 2: 21: 3 (1883) ≡ Lycopodioides sect. Rosulatae (A. Braun) Tzvel., Novosti Sist. Vyssh. Rast. 36: 25 (2004) – Type: S. involvens (Sw.) Spring.
= S. sect. Plagiophyllae (Warb.) Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1137 (2015) ≡ S. subser. Plagiophyllae Warb., Monsunia 1: 103 (1900) – Lectotype (designated by Zhou and Zhang, 2015: 1137): Selaginella biformis A. Braun ex Kuhn.
Etymology:—From _Kung_-, in honor of the late Prof. Hsian-Shiu Kung of Chengdu Institute of Biology, Chinese Academy of Sciences (herbarium CDBI), for his contributions to the study of ferns in general and those of Sichuan in particular (e.g., Kung, 1988). He was one of the early authors who recognized more than one genus in Selaginellaceae.
This genus circumscribed here corresponds to Selaginella sect. Circinatae and S. sect. Plagiophyllae sensu Zhou and Zhang (2015) combined. The genus is strongly supported as monophyletic in two of the three analyses based on concatenated data and strongly supported in nuclear tree (Fig. 1 and S2).
Plants of Kungiselaginella are strictly erect (a few creeping). Ventral leaves of members of this genus usually have two light-colored bands on the sides of veins (Chu, 2006). Their microspores are either slightly baculate (e.g., K. davidii (Franch.) Li Bing Zhang & X.M. Zhou, and K. moellendorffii (Hieron.) Li Bing Zhang & X.M. Zhou) or tuberculate to blunt-spiny (e.g., K. involvens) (Zhou et al., 2015). Two forms of habit and usually sterile microspores are present in S. biformis (Zhou et al., 2015).
This genus contains more than two dozen species in Asia.
Members:
Kungiselaginella argentea (Wall. ex Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium argenteum Wall. ex Hook. & Grev., Bot. Misc. 2: 384 (1831).
Kungiselaginella biformis (A. Braun ex Kuhn) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella biformis A. Braun ex Kuhn Fil. Afr. 189 (1868), nomen, ex Baker, J. Bot. 21: 145 (1883). 二形孔氏卷柏(新拟)
Kungiselaginella davidii (Franch.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella davidii Franch., Pl. David. 1: 344 (1884). 蔓生孔氏卷柏(新拟)
Kungiselaginella hellwigii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hellwigii Hieron., Bot. Jahrb. Syst. 50: 2, 12, no. 3 (1913).
Kungiselaginella hieronymiana (Alderw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella hieronymiana Alderw. Bull. Jard. Bot. Buitenz. II, 7: 31 (1912).
Kungiselaginella involvens (Sw.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium involvens Sw., Syn. Fil. 182, no. 50 (1806). 兖州孔氏卷柏(新拟)
Kungiselaginella moellendorffii (Hieron.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella moellendorffii Hieron. in Engl. & Prantl, Nat. Pfl. 1(4): 680, no. 102 (1901 publ. 1902). 江南孔氏卷柏(新拟)
Kungiselaginella muelleri (Baker) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella muelleri Baker, J. Bot. 23: 122, no. 123 (1885).
Kungiselaginella pallida (Hook. & Grev.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Lycopodium pallidum Hook. & Grev.; Enum. Filic. J. Bot. (Hook.) Kew Misc. 2: 389, no. 130 (1831) (non Beyr.) 灰白孔氏卷柏(新拟)
Kungiselaginella peltata (C. Presl) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella peltata C. Presl Abh. Königl. Böhm. Ges. Wiss., ser. 5.
Kungiselaginella polyura (Warb.) Li Bing Zhang & X.M. Zhou, comb. nov. Basionym: Selaginella polyura Warb., Monsunia 1: 104, 116, no. 64, t. VI, f. A. (1900).
5.8.3. Selaginella
Selaginella P. Beauv., Mag. Encycl. 9(5): 478 (1804), nom. cons_._ 同穗卷柏属(新拟) – Type: S. flabellata (L.) Spring. conserved type. prop (Wan et al., 2023).
= Selago P. Browne, Civ. Nat. Hist. Jamaica 82. 1756 – Lectotype (designated by Pichi Sermolli, Webbia 26: 158 (1971)): Lycopodium serpens Desv. ex Poir. non L. (1753).
≡ Stachygynandrum P. Beauv. ex Mirb. in Lamarck & Mirbel, Hist. Nat. Vég. 3: 477 (1803) ≡ Selaginella subg. Stachygynandrum (P. Beauv. ex Mirb.) Baker, J. Bot. Lond. 21: 3. 1883 ≡ Lycopodium subg. Stachygynandrum (P. Beauv. ex Mirb.) Spreng., Anleit., ed. 2, 2(1): 109 (1817) ≡ L. (unranked) Stachygynandrum (P. Beauv. ex Mirb.) Hook. & Grev., Bot. Misc. 2: 380 (1831) ≡ L. (unranked) Stachygynandrum (P. Beauv. ex Mirb.) Kunze, Linnaea 9: 8. 1834, “_Stachygynandra_” ≡ Didiclis subg. Stachygynandrum (P. Beauv. ex Mirb.) Rothm., Feddes Repert. Spec. Nov. Regni Veg. 54: 70 (1944) – Lectotype (designated by Pichi Sermolli, Webbia 26: 164 (1971)): Selaginella flabellata (L.) Spring.
≡ Selaginella subg. Heterophyllum Hieron. & Sadeb. in Engler & Prantl, Nat. Pflanzenf. l(4): 673 (1902) – Lectotype (designated by Zhou and Zhang, 2015: 1136): Selaginella flabellata (L.) Spring.
Selaginella s.s. in this study corresponds to S. subg. Stachygynandrum Zhou and Zhang (2015) excluding S. sect. Ascendentes, S. sect. Circinatae, and S. sect. Plagiophyllae. The monophyly of the genus is strongly supported with nuclear data but its relationships are unresolved with plastid data. The combined dataset supported its monophyly strongly in BI analysis but weakly in ML and MP analyses.
Species of Selaginella s.s. have erect, suberect, creeping, asscending (a few rosette-forming) plants. Megaspore surfaces are often reticulate (Fig. 11J, M−P). Microspore surfaces are baculate, blunt spiny, verrucate, papillate, coarse, or ridge (Fig. 11Q–T).
The newly defined genus Selaginella s.s. contains about 230 species nearly endemic to Americas except one (to a few) species in Africa. Within Selaginella s.s., four sections, S. sect. Austroamericanae, S. sect. Heterophyllae, S. sect. Poceres, and S. sect. Selaginella (= S. sect. Pallescentes sensu Zhou and Zhang (2015)), can be recognized.
Section 1. Selaginella sect. Selaginella 同穗卷柏组(新拟).
= Selaginella sect. Heterophyllae Spring in Mart., Fl. Bras_._ 1(2): 118 (1840) – Lectotype (designated by Zhou and Zhang, 2015: 1137): Selaginella flexuosa Spring.
= S. (unranked) Enodes Spring in Mart., Fl. Bras. 1(2): 118 (1840) – Lectotype (designated by Zhou and Zhang, 2015: 1136): S. flexuosa Spring.
= S. (unranked) Continuae Spring, Mém. Acad. Sci. Belg. 24: 53 (1850) ≡ S. ser. Continuae (Spring) Hieron. & Sadeb. in Engl. & Prantl, Nat. Pflanzenf. 1(4): 704 (1902) – Lectotype (designated by Zhou and Zhang, 2015: 1137): Selaginella microphylla (Kunth) Spring.
= S. (unranked) Pusillae Spring, Mém. Acad. Sci. Belg. 24: 53 (1850) – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. microphylla (Kunth) Spring.
= S. (unranked) Ascendentes A. Braun, App. Ind. Sem. Hort. Berol. 11 ([1857] 1858) – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. flexuosa Spring.
= S. (unranked) Persistentes A. Braun, App. Ind. Sem. Hort. Berol. 11 ([1857] 1858) – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. flexuosa Spring.
= S. sect. Dichotropae A. Braun, App. Ind. Sem. Hort. Berol., 11. ([1857] 1858). nom. illeg. – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. flexuosa Spring.
= S. (unranked) Resupinatae A. Braun, App. Ind. Sem. Hort. Berol. 12. ([1857] 1858) – Lectotype (designated by Zhou and Zhang, 2015: 1137): Selaginella stenophylla A. Braun.
= S. ser. Decumbentes Baker, J. Bot. Lond. 21: 3 (1883) – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. microphylla (Kunth) Spring.
= S. sect. Pleiomacrosporangiatae Hieron. & Sadeb. in Engl. & Prantl, Nat. Pflanzenf. 1(4): 673. (1902) – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. flexuosa Spring.
= S. ser. Monostelicae Hieron. & Sadeb. in Engl. & Prantl, Nat. Pflanzenf. 1(4): 673 (1902) – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. flexuosa Spring.
≡ S. sect. Stachygynandrum (P. Beauv. ex Mirb.) T. Moore, Ind. Fil. Cxxviii (1857) ≡ Lycopodioides sect. Stachygynandrum (P. Beauv. ex Mirb.) Tzvel., Novosti Sist. Vyssh. Rast. 36: 25 (2004) – Type: S. flabellata (L.) Spring.
Because the genus has been proposed to be conserved with a conserved type, Selaginella flabellata (Wan et al., 2023), our S. sect. Heterophyllae sensu Zhou and Zhang (2015) now becomes a synonym of the autonym.
This section is weakly supported as monophyletic in two but strongly in one of our three analyses based on the concatenated data (Fig. 1) and is composed of New-World (South & Central Americas) species only_._ They are suberect, rarely creeping or erect (if plants erect, main stems with monomorphic leaves), and have dimorphic leaves and various chromosome numbers: 2n = 18 (S. simplex Baker) (Graustein, 1930; Marcon et al., 2005), 2n = 20 [S. erythropus Spring and S. flabellata (L.) Spring] (Jermy et al., 1967).
Section 2. Selaginella sect. Austroamericanae Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1136 (2015) 南美同穗卷柏组(新拟) – Type: S. hartwegiana Spring.
= S. (unranked) Redivivae A. Braun, App. Ind. Sem. Hort. Berol. 11 ([1857] 1858) – Lectotype (designated by Zhou and Zhang, 2015: 1136): S. ciliata (Willd.) A. Braun [ = Selaginella novae-hollandiae (Sw.) Spring].
The monophyly of this section is weakly supported (Fig. 1). Species of this section have stem nearly erect, different from those species with dimorphic sporophylls from America that often have creeping stems_._ They have dimorphic sterile leaves on stem, different from those species with erect stem and monomorphic sporophylls species from America that have monomorphic sterile leaves on stem (Valdespino, 1993b). Moreover, at least Selaginella hartwegiana tend to have monomorphic leaves below first branch of the stems.
About 13 species of this section are sampled in our phylogenetic analysis (Fig. S3). This section mainly occurs in North to South America and a few possibly occur in Africa (currently only Selaginella cathedrifolia known).
Section 3. Selaginella. sect. Pallescentes Li Bing Zhang & X.M. Zhou 白变同穗卷柏组(新拟), Taxon 64(6): 1137 (2015) – Type: S. pallescens (C. Presl) Spring.
Members of this section have monomorphic sporophylls [_Selaginella apoda_ (L.) C. Morren has slightly dimorphic sporophylls, but generally its sporophylls are considered to be monomorphic; Valdespino, 1993a], but variable numbers of chromosomes, e.g., S. apoda: 2n = 18; S. pallescens (C. Presl) Spring: 2n = 20, 22; Selaginella pulcherrima Liebm.: 2n = 20 (Tschermak-Woes and Doležal-Janish, 1959; Jermy et al., 1967).
About eight species of this section were sampled in our phylogenetic analysis (Fig. S3). However, it is necessary that more morphological and molecular studies are needed to confirm which species in Americas should be placed in this section.
Section 4. Selaginella sect. Proceres (Spring) Li Bing Zhang & X.M. Zhou, Taxon 64(6): 1137 (2015) 高贵同穗卷柏组(新拟) ≡ S. (unranked) P r oceres Spring, Mem. Acad. Sci. Belg. 24: 53 (1850) – Lectotype (designated by Zhou and Zhang, 2015: 1137): S. oaxacana Spring.
This section is strongly supported as monophyletic and resolved as sister to Selaginella sect. pallescentes (Fig. 1). Species of this section have monomorphic sporophylls and reticulate megaspore surfaces (S. martensii Spring: Giorgi et al., 1997; Korall and Taylor, 2006; S. oaxacana Spring: Hellwig, 1969]. Only three species were included in our phylogenetic analysis (Fig. S3). More studies are needed.
Members of the genus:
Selaginella acanthostachys Baker, J. Bot. 21: 99, no. 40 (1883).
Selaginella achotalensis Shelton & Caluff, Willdenowia 33(1): 163 (2003).
Selaginella aculeatifolia Valdespino, Nordic J. Bot. 34(3): 372 (2016).
Selaginella acutifolia (Stolze) Valdespino, Mem. New York Bot. Gard. 88 (Pterid. Mexico): 558 (2004).
Selaginella agioneuma Valdespino & C. López, PeerJ 6(e4708): 4 (2018).
Selaginella alampeta M. Kessler & A.R.Sm., Edinburgh J. Bot. 63(1): 86 (2006).
Selaginella albolineata A.R.Sm., Ann. Mo. Bot. Gard. 77: 262, f. 7A–E (1990).
Selaginella alstonii G. Heringer, Salino & Valdespino, PhytoKeys 50: 65 (2015).
Selaginella altheae Valdespino, PhytoKeys 91: 15 (2017).
Selaginella amazonica Spring, Mart., Fl. Bras. 1(2): 124 (1840).
Selaginella anceps (C. Presl) C. Presl, Abh. Königl. Böhm. Ges. Wiss., Math.-Naturw. Cl. V, 3: 581 (repr. 151) (1845).
Selaginella apoda (L.) Spring, Mart., Fl. Bras. 1(2): 119 (1840).
Selaginella applanata A. Braun, Ann. Sci. Nat., Bot., sér. V, 3: 274 (1865).
Selaginella armata Baker, J. Bot. 22: 90, no. 125 (1884).
Selaginella arrecta A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 264 (1990).
Selaginella arsiclada Valdespino, Am. Fern J. 84(3): 99 (1994).
Selaginella ayitiensis Valdespino, Willdenowia 49(1): 71–80 (2019).
Selaginella bahiensis Spring, Mart., Fl. Bras. 1(2): 124, no. 12 (1840).
Selaginella bahiensis subsp. manausensis (Bautista) Jermy & J.M. Rankin, Bull. Brit. Mus. (Nat. Hist.) Bot. 9(4): 260 (1981).
Selaginella barnebyana Valdespino, Mem. New York Bot. Gard. 88 (Pterid. Mexico): 560–561 (2004).
Selaginella beitelii A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 264 (1990).
Selaginella bernoullii Hieron., Hedwigia 41: 192, no. 15 (1902).
Selaginella blepharodella Valdespino, PhytoKeys 50: 68 (2015).
Selaginella bombycina Spring, Mém. Acad. Roy. Sci. Belg. 24: 191 (1849).
Selaginella boomii Valdespino, Brittonia 67(4): 329 (2015).
Selaginella bracei Hieron. ex O.C. Schmidt, Repert. Spec. Nov. Regni Veg. 20: 156 (1924).
Selaginella breedlovei Valdespino, Mem. New York Bot. Gard. 88 (Pterid. Mexico): 563 (2004).
Selaginella brevifolia Baker, J. Bot. 21: 21(3): 83 (1883).
Selaginella breweriana A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 266 (1990).
Selaginella breynii Spring, Mart., Fl. Bras. 1(2): 121 (1840).
Selaginella brigitteana Valdespino, Willdenowia 49(1): 71–80 (2019).
Selaginella bryophila M. Kessler & A.R.Sm., Edinburgh J. Bot. 63(1): 91 (2006).
Selaginella cabrerensis Hieron., Hedwigia 43: 29 (1904).
Selaginella calosticha Spring, Mém. Acad. Roy. Sci. Belgique 24 [Monogr. Lyc. 2]: 206, no. 145 (1849).
Selaginella cardiophylla Valdespino, Brittonia 44(2): 199 (1992).
Selaginella carioi Hieron., Engl. & Prantl, Nat. Pflanzenf. 1(4): 688 (1901 publ. 1902).
Selaginella cathedrifolia Spring, Mém. Acad. Roy. Sci. Belgique 24 [Monogr. Lyc. 2]: 112, no. 58 (1849).
Selaginella cavernaria Caluff & Shelton, Willdenowia 44(3): 311 (2014).
Selaginella cavifolia A. Braun, Ann. Sci. Nat., Bot., sér. V, 5, 3: 272 (1865).
Selaginella cheiromorpha Alston, Jermy & J. M. Rankin, Bull. Brit. Mus. (Nat. Hist.) Bot. 9(4): 257 (1981).
Selaginella chiapensis A.R. Sm., Am. Fern J. 70(1): 25 (1980).
Selaginella chionoloma Alston ex Crabbe & Jermy, Am. Fern J. 63: 137 (1973).
Selaginella chrysoleuca Spring, Bull. Acad. Roy. Soc. Bruxelles 10(1): 226 (1843).
Selaginella confusa Spring, Flora 21: 218 (1838).
Selaginella contigua Baker, J. Bot. 22: 295, no. 162 (1884).
Selaginella cordifolia (Desv. ex Poir.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 228 (1843).
Selaginella correae Valdespino, Brittonia 45(4): 315 (1993).
Selaginella corrugis Mickel & Beitel, Pterid. Fl. Oaxaca (Mem. New York Bot. Gard. 46): 336 (1988).
Selaginella crinita Valdespino, PhytoKeys 50: 74 (2015).
Selaginella cristalensis Shelton & Caluff, Willdenowia 33(2): 435 (2003).
Selaginella cruciformis Alston ex Crabbe & Jermy, Fern Gaz. 11(4): 257 (1976).
Selaginella culverwellii N.R. Crouch, Ferns S. Afr. Compreh. Guide: 746 (2011).
Selaginella cuneata Mickel & Beitel, Pterid. Fl. Oaxaca (Mem. New York Bot. Gard. 46: 336 (1988).
Selaginella cyclophylla A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 266 (1990).
Selaginella dasyloma Alston, J. Bot. 72: 228 (1934).
Selaginella delicatissima Linden, Cat. n. 11: 20 (1856), nomen, ex A. Braun, App. Ind. Sem. Hort. Berol. 13 (1857).
Selaginella dendricola Jenman, Gard. Chron., ser. 3, 2: 99 (1887).
Selaginella densifolia Spruce ex Hook., Hook., 2nd Cent., t. 85 (1861) (non Klotzsch, 1844).
Selaginella denudata (Willd.) Spring, Flora 21: 212 (1839).
Selaginella eatonii Hieron. ex Small, Ferns trop. Florida 67 (1918).
Selaginella eclipes W.R. Buck, Canad. J. Bot. 55: 366 (1977).
Selaginella epipubens Caluff & Shelton, Willdenowia 39(1): 116 (2009).
Selaginella epirrhizos Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 229, no. 126 (1843).
Selaginella erectifolia Spring, Mém. Acad. Roy. Sci. Belgique 24 [Monogr. Lyc. 2]: 92, no. 36 (1849).
Selaginella erythropus (Mart.) Spring, Mart., Fl. Bras. 1(2): 125 (1840).
Selaginella euclimax Alston ex Crabbe & Jermy, Fern Gaz. 11: 259 (1976).
Selaginella falcata (P. Beauv.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 225, no. 91 (1843).
Selaginella finitima Mickel & Beitel, Pterid. Fl. Oaxaca (Mem. New York Bot. Gard. 46: 338 (1988).
Selaginella flabellata (L.) Spring, Flora 21: 198, no. 19 (1838).
Selaginella flacca Alston, Jermy & J. M. Rankin, Bull. Brit. Mus. (Nat. Hist.) Bot. 9(4): 269 (1981).
Selaginella flagellata (L.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10(1): 228 (1843).
Selaginella flexuosa Spring, Flora 21: 197, no. 18 (1838).
Selaginella fragillima Silveira, Bol. Comm. Geogr. Geol. Est. Minas Gerais 5: 127, t. 11 (1898).
Selaginella gioiae Valdespino, PhytoKeys 159: 74 (2020).
Selaginella glossophylla Crabbe & Jermy, Fern Gaz. 11: 259 (1976).
Selaginella guatemalensis Baker, J. Bot. 21: 243 (1883).
Selaginella gynostachya Valdespino, Fern Gaz. 18(2): 42 (2007 publ. 2008).
Selaginella haematodes (Kunze) Spring, in Mart., Fl. Bras. 1(2): 126 (1840).
Selaginella haenkeana Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 225 (1843).
Selaginella harrisii Underw. & Hieron., Symb. Ant. 7: 162 (1912).
Selaginella hartii Hieron., Urban, Symb. Ant. 3: 525 (1903).
Selaginella hartwegiana Spring, Mém. Acad. Roy. Sci. Belg. 24: 188 (1849).
Selaginella hemicardia Valdespino, Brittonia 44(2): 201 (1992).
Selaginella heterodonta (Desv. ex Poir.) Hieron., Symb. Ant. 9: 392 (1925).
Selaginella hirsuta Alston ex Crabbe & Jermy, Am. Fern J. 63: 138 (1973).
Selaginella hirtifolia Valdespino, Mem. New York Bot. Gard. 88 (Pterid. Mexico): 573–574 (2004).
Selaginella hispida (Willd.) A. Braun ex Urb., Symb. Ant. 9: 394 (1925).
Selaginella homaliae A. Braun, Ann. Sci. Nat., Bot., sér. V, 3: 274 (1865).
Selaginella huehuetenangensis Hieron., Hedwigia 43: 32 (1904).
Selaginella hyalogramma Valdespino, Am. Fern J. 107(2): 74 (2017).
Selaginella idiospora Alston, Bull. Brit. Mus. (Nat. Hist.) Bot. 1(8): 246, t. 6, f. A–E (1955).
Selaginella illecebrosa Alston, Bull. Brit. Mus. (Nat. Hist.) Bot. 1(8): 239, t. 5, f. A–E (1955).
Selaginella imbricans A.R.Sm., Ann. Mo. Bot. Gard. 77: 266, f. 9K–N (1990).
Selaginella jungermannioides (Gaudich.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 143 (1843).
Selaginella karowtipuensis Valdespino, Fern Gaz. 18(2): 46 (2007 publ. 2008).
Selaginella kochii Hieron., Koch-Grünberg Reisen in Nordwest-Brasilien, 2: 361 (1910).
Selaginella kriegeriana L.A. Góes, Syst. Bot. 43(4): 921 (2018).
Selaginella krugii Hieron., Symb. Ant. 3: 526 (1903).
Selaginella laxifolia Baker ex Krug ex Urb., Bot. Jahrb. Syst. 24: 151 (1897).
Selaginella lechleri Hieron., Engl. & Prantl, Nat. Pflanzenf. 1(4): 683 (1901).
Selaginella leonardii O.C. Schmidt, Fedde, Repert. 20: 155 (1924).
Selaginella leucoloma Alston, Crabbe & Jermy, Fern Gaz. 11(4): 262 (1976).
Selaginella lindenii Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 142, no. 52 (1843).
Selaginella lineolata Mickel & Beitel, Pterid. Fl. Oaxaca (Mem. New York Bot. Gard. 46): 343 (1988).
Selaginella longissima Baker, J. Bot. 19: 208 (1881).
Selaginella ludoviciana (A. Braun) A. Braun, Ann. Sci. Nat., Bot., sér. 4, 13: 58 (1860).
Selaginella lychnuchus Spring ex Klotzsch, Linnaea 20: 435 (1847), nomen, ex Spring, Monogr. Lyc. 2: 247, no. 186 (1849).
Selaginella macrostachya (Spring) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 144 (1843).
Selaginella magnafornensis Valdespino & C. López, PeerJ 6(e4708): 11 (2018).
Selaginella marahuacae A.R.Sm., Ann. Mo. Bot. Gard. 77: 268, f. 7F–I (1990).
Selaginella martensii Spring, Mém. Acad. Roy. Sci. Belg. 24: 129 (1849).
Selaginella mazaruniensis Jenman, Gard. Chron. 22: 210 (1897).
Selaginella mendoncae Hieron., Engl. & Prantl, Nat. Pflanzenf. 1(4): 693, no. 233. (1901 publ. 1902).
Selaginella meridensis Alston, Bull. Brit. Mus. (Nat. Hist.) Bot. 9(4): 267 (1981).
Selaginella mickelii Valdespino, Brittonia 44(3): 316 (1992).
Selaginella microdendron Baker, J. Bot. 23: 116 (1885).
Selaginella microdonta A.C. Sm., Bull. Torrey Bot. Club 58: 313 (1931).
Selaginella microphylla (Kunth) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 234 (1843).
Selaginella minima Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 139, no. 29 (1843).
Selaginella mixteca Mickel & Beitel, Pterid. Fl. Oaxaca (Mem. New York Bot. Gard. 46): 345 (1988).
Selaginella mollis A. Braun, Ann. Sci. Nat., Bot., sér. V, 5, 3: 276 (1865).
Selaginella monticola Valdespino, Phytotaxa 233(1): 154 (2015).
Selaginella moritziana Spring ex Klotzsch, Linnaea 20: 436 (1847).
Selaginella moritziana var. pearcei (Baker) Valdespino, Taxon 65(6): 1402 (2016).
Selaginella mortoniana Crabbe & Jermy, Am. Fern J. 63: 139 (1973).
Selaginella mosorongensis Hieron., Hedwigia 43: 4 (1904).
Selaginella mucronata G. Heringer, Salino & Valdespino, PhytoKeys 50: 78 (2015).
Selaginella mucugensis Valdespino, PhytoKeys 50: 82 (2015).
Selaginella muscosa Spring, Mart., Fl. Bras. 1(2): 120 (1840).
Selaginella myriostachya Valdespino, C. López & Góes-Neto, Phytotaxa 184(4): 235–244 (2014).
Selaginella nanophylla Valdespino, C. López & Góes-Neto, Phytotaxa 184(4): 235–244 (2014).
Selaginella nanuzae Valdespino, PhytoKeys 57: 98 (2015).
Selaginella neblinae A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 268 (1990).
Selaginella neospringiana Valdespino, PhytoKeys 57: 103 (2015).
Selaginella nothohybrida Valdespino, Brittonia 44(3): 319 (1992).
Selaginella novae-hollandiae (Sw.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 234, no. 161 (1843).
Selaginella oaxacana Spring, Mém. Acad. Roy. Sci. Belg. 24: 177 (1849).
Selaginella ophioderma Valdespino, Nordic J. Bot. 2022(3)-e03411: 2 (2022).
Selaginella orbiculifolia Shelton & Caluff, Willdenowia 33(2): 430 (2003).
Selaginella orizabensis Hieron., Hedwigia 41: 193, no. 16 (1902).
Selaginella osaensis A. Rojas, Revista Biol. Trop. 49(2): 446 (2001).
Selaginella ovifolia Baker, J. Bot. 22: 90 (1884).
Selaginella pallescens (C. Presl) Spring, Mart., Fl. Bras. 1(2): 132 (1840).
Selaginella palmiformis Alston ex Crabbe & Jermy, Am. Fern J. 63: 141 (1973).
Selaginella panurensis Baker, J. Bot. 21: 97 (1883).
Selaginella papillosa Valdespino, PhytoKeys 159: 81 (2020).
Selaginella pellucidopunctata Valdespino, PhytoKeys 57: 107 (2015).
Selaginella phiara Valdespino, C. López & L.A. Góes, Phytotaxa 184: 242 (2014).
Selaginella philipsonii (Jermy & J.M. Rankin) Valdespino, PhytoKeys 91: 32 (2017).
Selaginella plagiochila Baker, J. Bot. 21: 212, no. 67 (1883).
Selaginella plumieri Hieron., Urban, Symb. Ant. 7: 488 (1913).
Selaginella polyptera Valdespino, Mem. New York Bot. Gard. 88 (Pterid. Mexico): 588 (2004).
Selaginella popayanensis Hieron., Hedwigia 43: 9 (1904).
Selaginella porelloides (Lam.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10(1): 141 (1843).
Selaginella porphyrospora A. Braun, Ann. Sci. Nat., Bot., sér. V, 5, 3: 286 (1865).
Selaginella potaroensis Jenman, Gard. Chron. 3, 2: 154 (1887).
Selaginella praestans Alston, Bull. Brit. Mus. (Nat. Hist.) Bot. 9: 260 (1981).
Selaginella prasina Baker, J. Bot. 22: 113, no. 134 (1884).
Selaginella producta Baker, J. Bot. 21(8): 243 (1883).
Selaginella prolifera Valdespino, Mem. New York Bot. Gard. 88 (Pterid. Mexico): 590 (2004).
Selaginella pruskiana Valdespino, Brittonia 44(2): 202 (1992).
Selaginella psittacorrhyncha Valdespino, Phytoneuron 63: 2 (2017).
Selaginella pubens A.R. Sm., Ann. Mo. Bot. Gard. 77(2): 269 (1990).
Selaginella pubimarginata Valdespino, PhytoKeys 159: 87, 9–12 (2020).
Selaginella pulcherrima Liebm. ex E. Fourn., Mexic. Pl.1: 147 (1872).
Selaginella quadrifaria Alston, Jermy & Rankin, Bull. Brit. Mus. (Nat. Hist.) Bot. 9(4): 261 (1981).
Selaginella rachipterygia Valdespino, Cornejo & C. López, Novon 30(1): 70 (2022).
Selaginella radiata (Aubl.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 143, no. 54 (1843).
Selaginella ramosissima Baker, J. Bot. 23: 295 (1885).
Selaginella raynaliana Tardieu, Adansonia 3(3): 352–353, t. 1, f. 1–9 (1963).
Selaginella reflexa Underw., Bull. Torrey Bot. Club 21: 268 (1894).
Selaginella revoluta Baker, J. Bot. 21(5): 141 (1883).
Selaginella rhodostachya Baker, Timehri 5: 221 (1886), also Trans. Linn. Soc., Bot. 2: 296 (1887).
Selaginella roraimensis Baker, Timehri 5: 221 (1886), also Trans. Linn. Soc., Bot. 2: 295 (1887)
Selaginella rosea Alston, J. Bot. 70: 281–282 (1932).
Selaginella rostrata Valdespino, PhytoKeys 159: 93, 9 and 13 (2020).
Selaginella rotundifolia Spring, Bull. Acad. Roy. Soc. Bruxelles 10(1): 139 (1843).
Selaginella rzedowskii Lorea-Hem., Bol. Soc. Bot. Mexico 44: 24 (1983).
Selaginella salazariae Valdespino, Brittonia 45(4): 319 (1993).
Selaginella salinoi L.A. Góes & G. Heringer, Phytotaxa 224(3): 292 (2015).
Selaginella saltuicola Valdespino, PhytoKeys 50: 85 (2015).
Selaginella sandwithii Alston, Jermy & J.M. Rankin, Bull. Brit. Mus. (Nat. Hist.) Bot. 9(4): 289 (1981).
Selaginella scalariformis A.C. Sm., Bull. Torrey Bot. Club 58: 314 (1931).
Selaginella schultesii Alston ex Crabbe & Jermy, Am. Fern J. 63: 141 (1973).
Selaginella seemannii Baker, J. Bot. 21: 244 (1883).
Selaginella sematophylla Valdespino, G. Heringer & Salino, PhytoKeys 50: 89 (2015).
Selaginella serpens (Desv. ex Poir.) Spring, Bull. Acad. Roy. Soc. Bruxelles 10(1): 228 (1843).
Selaginella simplex Baker, J. Bot. 23: 293 (1885).
Selaginella smithiorum Valdespino, Brittonia 45(4): 322 (1993).
Selaginella sobolifera A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 269 (1990).
Selaginella solomonii Valdespino, Novon 24(1): 100 (2015).
Selaginella speciosa A. Braun, Ann. Sci. Nat., Bot., sér. V, 3: 274–275, no. 7 (1865).
Selaginella squamulosa Valdespino, PhytoKeys 91: 24 (2017).
Selaginella stenophylla A. Braun, App. Ind. Sem. Hort. Berol. 22 (1857).
Selaginella stomatoloma Valdespino, PhytoKeys 57: 113 (2015).
Selaginella striata Caluff & Shelton, Willdenowia 44(3): 313 (2014).
Selaginella subrugosa Mickel & Beitel, Pterid. Fl. Oaxaca (Mem. New York Bot. Gard. 46: 352 (1988).
Selaginella substipitata Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 227, no. 110 (1843).
Selaginella surucucusensis L.A. Goés & E.L.M. Assis, Kew Bull. 72(40): 1 (2017).
Selaginella tanyclada Alston ex Crabbe & Jermy, Am. Fern J. 63: 143 (1973).
Selaginella taylorii Valdespino, Brittonia 45(4): 320 (1993).
Selaginella terezoana Bautista, Bol. Mus. Paraense Emilio Goeldi, Bot., 45: 1 (1974).
Selaginella thysanophylla A.R.Sm., Ann. Mo. Bot. Gard. 77(2): 270 (1990).
Selaginella truncata H. Karst. ex A. Braun, Index Sem. Hort. Berol. 1857: 15 (1857).
Selaginella trygonoides Valdespino, PhytoKeys 57: 116 (2015).
Selaginella tuberculata Spruce ex Baker, J. Bot. 21(3): 83 (1883).
Selaginella tuberosa B. McAlpin & Lellinger, Brenesia 24: 409 (1986).
Selaginella tyleri A.C. Sm., Bull. Torrey Bot. Club 58: 311 (1931).
Selaginella umbrosa Lem. ex Hieron., Engl. & Prantl, Nat. Pflanzenf. 1(4): 683, f. 404 (1901).
Selaginella undata Shelton & Caluff, Willdenowia 33(2): 427 (2003).
Selaginella urquiolae Caluff & Shelton, Willdenowia 39(1): 112 (2009).
Selaginella valdepilosa Baker, J. Bot. 21: 82 (1883).
Selaginella ventricosa Valdespino & C. López, PeerJ 6(e4708): 15 (2018).
Selaginella vernicosa Baker, Timehri 5: 220 (1886), also Trans. Linn. Soc., Bot. 2: 295 (1887).
Selaginella vestiens Baker, J. Bot. 21: 97, no. 34 (1883).
Selaginella viticulosa Klotzsch, Linnaea 18: 524 (1844).
Selaginella wolffii Sodiro, Crypt. Vasc. Quit. 620 (1893), also Anales Univ. Centr. Ecuador 12(83): 490 (1895).
Selaginella wurdackii Alston, Bull. Brit. Mus. (Nat. Hist.) Bot. 9: 280 (1981).
Selaginella xanthoneura Valdespino, PhytoKeys 159: 98, figs. 5, 14–18 (2020).
Selaginella xiphophylla Baker, J. Bot. 22(10): 296 (1884).
Selaginella zartmanii Valdespino, C. López & A.M. Sierra, PeerJ 6(e4708): 21 (2018).
5.9. Incertae sedis
Selaginella birarensis Kuhn, Forschungsr. Gazelle, Bot. Alg. 19 (1889).
Selaginella boschai Hieron., Hedwigia 51: 243, no. 2 (1912).
Selaginella dahlii Hieron., Engl. Bot. Jahrb. 50: 2, 10, no. 2 (1913).
Selaginella firmuloides Warb., Monsunia 1: 105, 118, no. 77 (1900).
Selaginella ketra-ayam Alderw., Bull. Jard. Bot. Buitenz. II, 1: 24–25 (1911).
Selaginella ledermanni Hieron., Engl. Bot. Jahrb. 56: 224, no. 6 (1920).
Selaginella pilosula Alderw., Bull. Jard. Bot. Buitenzorg, ser. 3, 5: 233 (1922).
Selaginella polita Ridl., J. Fed. Mal. States Mus. 6: 202, no. 348 (1915).
Selaginella presliana Spring, Bull. Acad. Roy. Soc. Bruxelles 10: 137, no. 16 (1843).
Selaginella propinqua Alderw., Bull. Jard. Bot. Buitenz. III, 5: 233 (1922).
Selaginella pseudovolkensii Hosok., Trans. Nat. Hist. Soc. Taiwan 31: 471 (1941).
Selaginella ridleyi Baker, Ann. Bot. 8: 131, no. 58 (1894).
Selaginella schaffneri Hieron., Nat. Pflanzenf. 1: 674 (1902).
Selaginella sonneratii Hieron., Engl. Bot. Jahrb. 50: 2, 7, no. 1 (1913) [_soneratii_].
Declaration of competing interest
The authors declare no conflict of interest.
Author contributions
Conceptualization: LBZ; Investigation: XMZ; Writing, Reviewing, and Editing: LBZ and XMZ; Visualization: XMZ and LBZ; Supervision: LBZ.
Conflicts of Interest
We declare no conflicts of interest.
Acknowledgments
The research was partially supported by the Natural Science Foundation of China (#31900186, #32260050), Yunnan Fundamental Research Projects (Grant NO. 202301BF07001-016), and the Glory Light International Fellowship for Chinese Botanists at Missouri Botanical Garden (MO) to X.M. Zhou. We thank Arthur Haines, Michael Hassler, Petra Korall, Carl Rothfels, Alan Smith, and Alan Weakley for helpful email discussion in Feb. 2021, Carl Rothfels, Timothée Le Péchon and other collaborators for sharing materials for our earlier studies, Atsushi Ebihara and Noriaki Murakami for sharing specimen images, and Jing Zhao, Jian-Jun Yang, Shao-Li Fang, and Yu-Xin Li for helping with the figures, Robbin Moran and the owner of plants-of-styria.uni-graz.at/images/selaginella-selaginoides.html for allowing us to use their online images, and Alan Weakley and three anonymous reviewers for helpful comments.
Footnotes
Peer review under responsibility of Editorial Office of Plant Diversity.
Appendix A. Supplementary data
The following are the Supplementary data to this article.
Fig. S1
Maximum likelihood phylogeny of Selaginellaceae based on plastid rbcL. Maximum likelihood bootstrap support values are shown along the branches. Red and green names indicate the 7 subfamilies and 19 genera, respectively, recognized in current classification. Blue and black names indicate the subgenera and sections recognized by Zhou and Zhang (2015). Purple names indicate the 6 sections recognized by Zhou and Zhang (2015) and this study at the same rank.
Fig. S2
Maximum likelihood phylogeny of Selaginellaceae based on nuclear (5.8S, 18S, 26S, SQD1, and pgiC) sequences. Maximum likelihood bootstrap support values are shown along the branches. Red and green names indicate the 7 subfamilies and 19 genera, respectively, recognized in current classification. Blue and black names indicate the subgenera and sections recognized by Zhou and Zhang (2015). Purple names indicate the 6 sections recognized by Zhou and Zhang (2015) and this study at the same rank.
Fig. S3
Maximum likelihood phylogeny of Selaginellaceae based on plastid rbcL and nuclear (5.8S, 18S, 26S, SQD1, and pgiC) sequences. Maximum likelihood bootstrap support valuesare shown along the branches. Red and green names indicate the 7 subfamilies and 19 genera, respectively, recognized in current classification. Blue and black names indicate the subgenera and sections recognized by Zhou and Zhang (2015). Purple names indicate the 6 sections recognized by Zhou and Zhang (2015) and this study at the same rank.
Multimedia component 4
References
- Akaike H. A new look at the statistical model identification. Autom. Cont. IEEE Transact. 1974;19:716–723. [Google Scholar]
- Arrigo N., Therrien J., Anderson C.L., et al. A total evidence approach to understanding phylogenetic relationships and ecological diversity in Selaginella subg. Tetragonostachys. Am. J. Bot. 2013;100:1672–1682. doi: 10.3732/ajb.1200426. [DOI] [PubMed] [Google Scholar]
- Badré F. Institut de Recherche pour le Développement; Paris: 2008. Ptéridophytes. 1. Psilotacées à 26 Marsiléacées. Flore des Mascareignes-La Réunion, Maurice, Rodrigues. [Google Scholar]
- Baker J.G. A synopsis of the genus Selaginella. J. Bot., Le. 1883;21(1–5) 42–46, 80–84, 97–100, 141–145, 210–213, 240–244. [Google Scholar]
- Baker J.G. Bell; London: 1887. Handbook of the Fern-Allies: A Synopsis of the Genera and Species of the Natural Orders Equisetaceae, Lycopodiaceae, Selaginellaceae, Rhizocarpeae. [Google Scholar]
- Banks J.A. Selaginella and 400 million years of separation. Annu. Rev. Plant Biol. 2009;60:223–238. doi: 10.1146/annurev.arplant.59.032607.092851. [DOI] [PubMed] [Google Scholar]
- Bauer D.S., Prado J., Trovó M., et al. Megaspore investigations of Selaginella species from Sầo Paulo, Brazil. Am. Fern J. 2016;106:55–86. doi: 10.1640/0002-8444-106.2.55.1. [DOI] [Google Scholar]
- Börner C. Verlag; Leipzig: 1912. Volksflora, Eine Flora für das deutsche Volk. [Google Scholar]
- Chen C.-W., Perrie L., Glenny D., et al. National Museum of Natural Science; Taichung: 2017. Sol Amazing: Lycophytes and Ferns of the Solomon Slands; pp. 1–550. [Google Scholar]
- Chen D.-K., Zhou X.-M., Rothfels C.J., et al. A global phylogeny of Lycopodiaceae (Lycopodiales; lycophytes) with the description of a new genus, Brownseya, from Oceania. Taxon. 2022;71:25–51. doi: 10.1002/tax.12597. [DOI] [Google Scholar]
- Christenhusz J.M., Fay M., Bynd J.W. GLOVAP nomenclature Part 1 (the global flora) The Global Flora, Special Edition. 2018;4:1–155. [Google Scholar]
- Chu W.-M. In: Wu C.Y., editor. Vol. 20. Science Press; Beijing: 2006. Selaginellaceae; pp. 35–93. (Flora Yunnanica). [Google Scholar]
- Darriba D., Taboada G.L., Doallo R., et al. JModelTest 2: more models, new heuristics and parallel computing. Nat. Methods. 2012;9:772. doi: 10.1038/nmeth.2109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Gasper A.L., Dittrich V.A.O., Smith A.R., et al. A classification for Blechnaceae (Polypodiales: Polypodiopsida): new genera, resurrected names, and combinations. Phytotaxa. 2016;275:191–227. doi: 10.11646/phytotaxa.275.3.1. [DOI] [Google Scholar]
- Du X.-Y., Lu J.-M., Zhang L.-B., et al. Simultaneous diversification of polypodiales and angiosperms in the mesozoic. Cladistics. 2021;37:518–539. doi: 10.1111/cla.12457. [DOI] [PubMed] [Google Scholar]
- Farris J.S., Albert V.A., Källersjö M., et al. Parsimony jackknifing outperforms neighbor-joining. Cladistics. 1996;12:99–124. doi: 10.1111/j.1096-0031.1996.tb00196.x. [DOI] [PubMed] [Google Scholar]
- Fawcett S., Smith A.R. BRIT Press, Fort Worth Botanic Garden, Botanical Research Institute of Texas; U.S.A: 2021. A Generic Classifcation of the Thelypteridaceae. Sida, Bot. Misc. 59. [Google Scholar]
- Fawcett S., Smith A.R., Sundue M., et al. A global phylogenomic study of the Thelypteridaceae. Sys. Bot. 2021;46:891–915. doi: 10.1600/036364421X16370109698650. [DOI] [Google Scholar]
- Felsenstein J. Maximum likelihood and minimum-steps methods for estimating evolutionary trees from data on discrete characters. Syst. Biol. 1973;22:240–249. doi: 10.1093/SYSBIO/22.3.240. [DOI] [Google Scholar]
- Fraser-Jenkins C.R., Kandel D.R., Pariyar S. Vol. 1. National Herbarium and Plant Laboratories, Department of Plant Resources, Ministry of Forests and Soil Conservation; Kathmandu: 2015. (Ferns and Fern-Allies of Nepal). [Google Scholar]
- Fraser-Jenkins C.R., Gandhi K.N., Kholia B.S., et al. Bishen Singh Mahendra Pal Singh; Dehra Dun: 2017. An Annotated Checklist of Indian Pteridophytes Part-1 (Lycopodiaceae to Thelypteridaceae) [Google Scholar]
- Giorgi A.F.D., Holderegger R., Schneller J.J. Aspects of spore dispersal in Selaginella. Am. Fern J. 1997;87:93–103. [Google Scholar]
- Graustein J.E. Evidences of hybridism in Selaginella. Bot. Gaz. 1930;90:46–74. [Google Scholar]
- González A., Dittrich V.A.O., Arana M.D. Telmatoblechnum (Blechnaceae): a new genus for the Uruguayan flora. Darwiniana. 2020;8:525–529. doi: 10.14522/darwiniana.2020.82.920. [DOI] [Google Scholar]
- Hall T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999;41:95–98. [Google Scholar]
- Hassler M. 2022. World Ferns. Checklist of Ferns and Lycophytes of the World. Selaginella. [Google Scholar]
- Hellwig R.L. Spores of the heterophyllous Selaginellae of Mexico and Central America. Ann. Mo. Bot. Gard. 1969;56:444–464. [Google Scholar]
- Hieronymus G., Sadebeck R. In: Engler A., Prantl K., editors. Vol. 1. Engelmann; Leipzig: 1901. Selaginellaceae; pp. 621–716. (Die Natürlichen Pflanzenfamilien). pt 4. [Google Scholar]
- Jansen R.K., Ruhlman T.A. In: Genomics of Chloroplasts and Mitochondria. Bock R., Knoop V., editors. Springer; Dordrecht (The Netherlands): 2012. Plastid genomes of seed plants; pp. 103–126. [DOI] [Google Scholar]
- Jermy A.C. Subgeneric names in Selaginella. Fern Gaz. 1986;13:117–118. [Google Scholar]
- Jermy A.C. In: The Families and Genera of Vascular Plants. Kramer K.U., Green P.S., editors. Vol. 1. Springer; Berlin: Heidelberg and New York: 1990. Selaginellaceae; pp. 39–45. (Pteridophytes and Gymnosperms). [Google Scholar]
- Jermy A.C., Holmes J.S. ABRS/CSIRO Australia; Melbourne: 1998. Selaginellaceae in Flora of Australia Volume 48, Ferns, Gymnosperms and Allied Groups. [Google Scholar]
- Jermy A.C., Jones K., Colden C. Cytomorphological variation in Selaginella. J. Linn. Soc., Bot. 1967;60:147–158. [Google Scholar]
- Kang J.S., Zhang H.-R., Wang Y.-R., et al. Distinctive evolutionary pattern of organelle genomes linked to the nuclear genome in Selaginellaceae. Plant J. 2020;104:1657–1672. doi: 10.1111/tpj.15028. [DOI] [PubMed] [Google Scholar]
- Kato M. Tokai University Press; Tokyo: 2008. Illustrated Flora of Ferns and Fern Allies of South Pacific Islands. [in Japanese] [Google Scholar]
- Katoh K., Standley D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 2013;30:772–780. doi: 10.1093/molbev/mst010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klaus K.V., Schulz C., Bauer D.S., et al. Historical biogeography of the ancient lycophyte genus Selaginella: early adaptation to xeric habitats on Pangea. Cladistics. 2017;33:469–480. doi: 10.1111/cla.12184. [DOI] [PubMed] [Google Scholar]
- Korall P., Kenrick P. Phylogenetic relationships in Selaginellaceae based on rbcL sequences. Am. J. Bot. 2002;89:506–517. doi: 10.3732/ajb.89.3.506. [DOI] [PubMed] [Google Scholar]
- Korall P., Kenrick P. The phylogenetic history of Selaginellaceae based on DNA sequences from the plastid and nucleus: extreme substitution rates and rate heterogeneity. Mol. Phylogenet. Evol. 2004;31:852–864. doi: 10.1016/j.ympev.2003.10.014. [DOI] [PubMed] [Google Scholar]
- Korall P., Taylor W.A. Megaspore morphology in the Selaginellaceae in a phylogenetic context: a study of the megaspore surface and wall structure using scanning electron microscopy. Grana. 2006;45:22–60. doi: 10.1080/00173130500520453. [DOI] [Google Scholar]
- Korall P., Kenrick P., Therrien J.P. Phylogeny of Selaginellaceae: evaluation of generic/subgeneric relationships based on rbcL gene sequences. Int. J. Plant Sci. 1999;160:585–594. doi: 10.1086/314137. [DOI] [Google Scholar]
- Kung H.-S. Contributions to the genus Selaginella Beauv. from Sichuan. Acta Bot. Yunnan. 1981;3:251–256. [Google Scholar]
- Kung H.-S. Vol. 6. Science and Technology Press; Chengdu, Sichuan: 1988. (Flora Sichuanica). [Google Scholar]
- Kuntze O. Vol. 2. A. Felix; Leipzig: 1891. (Revisio Generarum Plantarum). [Google Scholar]
- Lamark J.-B., Mirbel B. Paris; Raynal: 1803. Histoire Naturelle des Végétaux, Classés par Familles. Vol. 3. [Google Scholar]
- Lopes L.K.C., Góes-Neto L.A.A., Feio A.C. Stem anatomy and its relevance for the taxonomic survey of Selaginella subg. Gymnogynum (Selaginellaceae) Plant Syst. Evol. 2020;306:1–11. doi: 10.1007/s00606-020-01655-x. [DOI] [Google Scholar]
- Love A., Love D. In IOPB chromosome number reports LIII. Taxon. 1976;25:483–500. [Google Scholar]
- Loyal D.S., Kumar V. Cytotaxonomic observations on three Selaginella species from north-western Himalayas. Indian Fern J. 1984;1:59–62. [Google Scholar]
- Liu B.-D., Bao W.-M., Aur C.-W. Studies on the spores of morphology of the family Selaginellaceae from China. Bull. Bot. Res. 1989;9:113–131. [Google Scholar]
- Liu J.-X., Zhang X.-C., Sun X.-H., et al. New information about the palynology of Selaginellaceae from China. J. Chin. Electr. Microsc. Soc. 2001;20:511–512. [Google Scholar]
- Lu T.N., Zhou X.-M., Zhang L., et al. A global plastid phylogeny of the cliff fern family Woodsiaceae and a two-genus classification of Woodsiaceae with the description of ×Woodsimatium nothogen. nov. Taxon. 2020;68:1149–1172. doi: 10.1002/tax.12180. [DOI] [Google Scholar]
- Maddison W.P., Maddison D.R. Mesquite, a modular system for evolutionary analysis. Version 2.75. 2011 https://www. mesquiteproject.org [Google Scholar]
- Marcon A.B., Barros I.C.L., Guerra M. Variation in chromosome numbers, CMA bands and 45S rDNA sites in species of Selaginella (Pteridophyta) Ann. Bot. 2005;95:271–276. doi: 10.1093/aob/mci022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason-Gamer R.J., Kellogg E.A. Testing for phylogenetic conflict among molecular data sets in the tribe Triticeae (Gramineae) Syst. Biol. 1996;45:524–545. doi: 10.1093/sysbio/45.4.524. [DOI] [Google Scholar]
- Marcon A.B. Karyology of some pteridophytes species occurring in the north-east of Brazil. Acta Bot. Brasil. 2003;17:19–26. doi: 10.1590/S0102-33062003000100002. [DOI] [Google Scholar]
- Mickel J.T., Hellwig R.L. Actino-plectostely, a complex new stelar pattern in Selaginella. Am. Fern J. 1969;59:123–134. doi: 10.2307/1545992. [DOI] [Google Scholar]
- Mickel J.T., Smith A.R. The pteridophytes of Mexico. Memoir. N. Y. Bot. Gard. 2004;88:1–1055. [Google Scholar]
- Miller H.A. The “moss” Hypopterygiopsis is Selaginella. Taxon. 1967;16:70–71. [Google Scholar]
- Miller M.A., Pfeiffer W., Schwartz T. Proceedings of the Gateway Computing Environments Workshop (GCE), 14 November 2010. 2010. Creating the CIPRES Science Gateway for inference of large phylogenetic trees; pp. 1–8. New Orleans, LA. [Google Scholar]
- Mower J.P., Ma P.-F., Grewe F., et al. Lycophyte plastid genomics: extreme variation in GC, gene and intron content and multiple inversions between a direct and inverted orientation of the rRNA repeat. New Phytol. 2019;222:1061–1075. doi: 10.1111/nph.15650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukhopadhyay R., Goswami N. Cytotaxonomic observations on two species of Selaginella P. Beauv. from India. Indian Fern J. 1996;13:22–29. doi: 10.1007/BF02344419. [DOI] [Google Scholar]
- Øllgaard B. Nota cientffica/short communication: nomenclatural changes in Brazilian lycopodiaceae. Rodriguesia. 2012;63:479–482. doi: 10.1590/S2175-78602012000200020. [DOI] [Google Scholar]
- Palisot de Beauvois A.M.F.J. Suite de l'Aethéogamie. Mag. Encycl. 1804;9:472–483. [Google Scholar]
- Pol D. Empirical problems of the hierarchical likelihood ratio test for model selection. Syst. Biol. 2004;53:949–962. doi: 10.1080/10635150490888868. [DOI] [PubMed] [Google Scholar]
- Posada D., Buckley T.R. Model selection and model averaging in phylogenetics: advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004;53:793–808. doi: 10.1080/10635150490522304. [DOI] [PubMed] [Google Scholar]
- PPG (The Pteridophyte Phylogeny Group) I A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016;54:563–603. doi: 10.1111/jse.12229. [DOI] [Google Scholar]
- Quanansah N. Sporangial distribution patterns in the strobili of African and Madagascan Selaginella. Ann. Bot. 1988;61:243–247. [Google Scholar]
- Quansah N., Thomas B.A. Sporophyll-pteryx in african and American Selaginella. Fern Gaz. 1985;13:49–52. [Google Scholar]
- Rambaut A., Drummond A.J. 2007. Tracer 1.4.http://beast.bio.ed.ac.uk/Tracer Available at: [Google Scholar]
- Reeve R.M. The spores of the genus Selaginella in north central and north eastern United States. Rhodora. 1935;37:342–345. [Google Scholar]
- Ronquist F., Huelsenbeck J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–1574. doi: 10.1093/bioinformatics/btg180. [DOI] [PubMed] [Google Scholar]
- Roux J.P. Pretoria; South Africa: 2009. Synopsis of the Lycopodiophyta and Pteridophyta of Africa, Madagascar and neighbouring islands. South African National Biodiversity Institute (SANBI) [Google Scholar]
- Rothmaler W. Pteridophyten-studien I. Feddes Repert. Spec. Nov. Regni Veg. 1944;54:55–82. [Google Scholar]
- Ruhlman T.A., Jansen R.K. In: Advances in Botanical Research. Chaw S.-M., Jansen R.K., editors. Elsevier; London: 2018. Aberration or analogy? The atypical plastomes of geraniaceae; pp. 223–262. [Google Scholar]
- Sakurai K. Bryoflora von Micronesia.I–II. Bot. Mag. Tokyo. 1943;57:249–257. [Google Scholar]
- Satou Z. Miscellaneous notes on ferns and fern allies. Hikobia. 1997;12:267–270. [Google Scholar]
- Schuettpelz E., Pryer K.M. Evidence for a Cenozoic radiation of ferns in an angiosperm-dominated canopy. Proc. Natl. Acad. Sci. U.S.A. 2009;106:11200–11205. doi: 10.1073/pnas.0811136106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuettpelz E., Rouhan G., Pryer K.M., et al. Are there too many fern genera? Taxon. 2018;67:473–480. doi: 10.12705/673.1. [DOI] [Google Scholar]
- Schulz C., Homberg J., Stützel T. Taxonomic revision of Selaginella subg. Ericetorum. Syst. Bot. 2013;38:5–14. [Google Scholar]
- Smith D.R. Unparalleled GC content in the plastid DNA of Selaginella. Plant Mol. Biol. 2009;71:627–639. doi: 10.1007/s11103-009-9545-3. [DOI] [PubMed] [Google Scholar]
- Smith A.R., Weststrand S., Korall P. Selaginella pectinata resurrected — The correct name for an unusual endemic spikemoss from Madagascar. Am. Fern J. 2016;106:131–134. [Google Scholar]
- Soják J. Generische problematik der Selaginellaceae. Preslia. 1992;64:151–158. [Google Scholar]
- Stamatakis A., Hoover P., Rougemont J. A rapid bootstrap algorithm for the RAxML Web servers. Syst. Biol. 2008;57:758–771. doi: 10.1080/10635150802429642. [DOI] [PubMed] [Google Scholar]
- Stefanović S., Rakotondrainibe F., Badre F. In: Flore de Madagascar et des Comores, Fam. 14. Humbert H., Leroy J.-F., editors. Museum national d'Histoire naturelle; Paris: 1997. Selaginellacees; pp. 1–68. [Google Scholar]
- Steyermark J.A. A remarkable new Selaginella from Venezuela. Ann. Mo. Bot. Gard. 1986;73:209–215. [Google Scholar]
- Sundue M.A., Parris B.S., Ranker T.A., et al. Global phylogeny and biogeography of grammitid ferns (Polypodiaceae) Mol. Phylogenet. Evol. 2014;81:195–206. doi: 10.1016/j.ympev.2014.08.017. [DOI] [PubMed] [Google Scholar]
- Swofford D. Sinauer Associates; Sunderland, MA: 2002. PAUP 4.0b10: Phylogenetic Analysis Using Parsimony. [DOI] [Google Scholar]
- Takamiya M. Comparative karyomorphology and interrelationships of Selaginella in Japan. J. Plant Res. 1993;106:149–166. [Google Scholar]
- Tryon R.M. Selaginella rupestris and its allies. Ann. Mo. Bot. Gard. 1955;42:1–99. [Google Scholar]
- Tryon A.F., Lugardon B. Springer; New York: 1991. Spores of the Pteridophyta: Surface, Wall Structure, and Diversity Based on Electron Microscope Studies. [Google Scholar]
- Tschermak-Woess E., Dolezal-Janish R. Über die karyologische Anatomie einiger Pteridophyten sowie auffallende Unterschiede im Kernvolumen bei Cyrtomium falcatum. Österr. Bot. Zeitsch. 1959;106:315–324. [Google Scholar]
- Tsuji S., Ueda K., Nishiyama T., et al. The chloroplast genome from a lycophyte (microphyllophyte), Selaginella uncinata, has a unique inversion, transpositions and many gene losses. J. Plant Res. 2007;120:281–290. doi: 10.1007/s10265-006-0055-y. [DOI] [PubMed] [Google Scholar]
- Turland N.J., Wiersema J.H., Barrie F.R., et al., editors. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Koeltz Botanical Books; Glashütten: 2018. (Regnum Vegetabile 159). [Google Scholar]
- Tzvelev N. De genere Selaginella P. Beauv. (Selaginellaceae) in rossia. Nov. Sist. Vysshikh Rastenii. 2004;36:22–27. [Google Scholar]
- Valdespino I.A. In: Flora of North America Editorial Committee. Flora of North America North of Mexico, editor. Vol. 2. Oxford University Press; New York: 1993. Selaginellaceae; pp. 38–63. Pteridophytes and Gymnosperms. [Google Scholar]
- Valdespino I.A. Notes on neotropical Selaginella (Selaginellaceae), including new species from Panama. Brittonia. 1993;45:315–327. doi: 10.2307/2807605. [DOI] [Google Scholar]
- Valdespino I.A. Novelties in Selaginella (Selaginellaceae – lycopodiophyta), with emphasis on Brazilian species. PhytoKeys. 2015;57:93–133. doi: 10.3897/phytokeys.57.6489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valdespino I.A. Selaginella ayitiensis and S. brigitteana (Selaginellaceae): two new species from Hispaniola Island. Willdenowia. 2019;49:71–80. doi: 10.3372/wi.49.49109. [DOI] [Google Scholar]
- Valdespino I.A. Selaginella aculeatifolia sp. nov. (Selaginellaceae– lycopodiophyta) from the foothills of Cerro Neblina, Venezuela. Nord. J. Bot., Le. 2016;34:370–375. doi: 10.1111/njb.00954. [DOI] [Google Scholar]
- Valdespino I.A. Selaginella hyalogramma (Selaginellaceae – lycopodiophyta): a new species from Venezuela, South America. Am. Fern J. 2017;107:72–83. doi: 10.1640/0002-8444-107.2.72. [DOI] [Google Scholar]
- Valdespino I.A. Validation of Selaginella psittacorrhyncha (Selaginellaceae), a new species from the Guiana highlands of Venezuela and Brazil. Phyton. 2017;63:1–8. [Google Scholar]
- Valdespino I.A. Novel fern- and centipede-like Selaginella (Selaginellaceae) species and a new combination from South America. PhytoKeys. 2017;91:13–38. doi: 10.3897/phytokeys.91.21417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valdespino I.A. Taxonomic innovations in South American Selaginella (Selaginellaceae, Lycopodiophyta): description of five new species and an additional range extension. PhytoKeys. 2020;159:71–113. doi: 10.3897/phytokeys.159.55330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valdespino I.A., López C.A. Selaginella moraniana (Selaginellaceae – lycopodiophyta): a new articulate species with puberulent lateral leaves from northwestern South America. Brittonia. 2020;72:23–34. doi: 10.1007/s12228-019-09596-7. [DOI] [Google Scholar]
- Valdespino I.A., Zimmer B. Typification of selected Neotropical Selaginella (Lycopodiophyta: Selaginellaceae) taxon names and some nomenclatural innovations. Taxon. 2016;65:1391–1408. doi: 10.12705/656.10. [DOI] [Google Scholar]
- Valdespino I.A., Heringer G., Salino A., et al. Seven new species of Selaginella subg. Stachygynandrum from Brazil and new synonyms for the genus. PhytoKeys. 2015;50:61–99. doi: 10.3897/phytokeys.50.4873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valdespino I.A., López C.A., Ceballos J. Selaginella germinans (Selaginellaceae), a new articulate species from Chapada dos Veadeiros region in the state of Goiás. Brazil. Bot. Lett. 2018;165:487–493. doi: 10.1080/23818107.2018.1496849. [DOI] [Google Scholar]
- Valdespino I.A., López C.A., Sierra A.M., et al. From the guiana highlands to the Brazilian atlantic rain forest: four new species of Selaginella (Selaginellaceae – lycopodiophyta: S. agioneuma, S. magnafornensis, S. ventricosa, and S. zartmanii) PeerJ. 2018;6 doi: 10.7717/peerj.4708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walton J., Alston A.H.G. In: Manual of Pteridology. Verdoorn F., editor. Nijhoff; The Hague: 1938. Lycopodiinae; pp. 500–506. [Google Scholar]
- Wan X., Zhou X.-M., Zhang L., et al. Proposal to conserve Selaginella, nom. cons., (Selaginellaceae) with a conserved type. Taxon. 2023;72:429–430. doi: 10.1002/tax.12918. [DOI] [Google Scholar]
- Wang L.-J., Zhang X.-C., Liu J.-X. Studies on the complementary relationship of surface ornamentations between megaspores and microspores. Microsc. Res. Tech. 2018;81:1474–1488. doi: 10.1002/jemt.23148. [DOI] [PubMed] [Google Scholar]
- Warburg O. Vol. 1. Engelmann; Leipzig: 1900. (Monsunia). [Google Scholar]
- Weakley A.S. The University of North Carolina Herbarium; Chapel Hill: 2012. Flora of the Southern and Mid-Atlantic States. [Google Scholar]
- Weakley A.S. In: Weakley A.S., Poindexter D.B., Sorrie B.A., et al., editors. Vol. 16. J. Bot. Res. Inst. Texas; 2022. Recognition of segregate genera in Selaginella s.l. for the Flora of the Southeastern United States, with four new combinations needed; pp. 403–405. (Studies in the Vascular Flora of the Southeastern United States. VIII). 377–418. [Google Scholar]
- Weakley, A.S., Southeastern Flora Team . Chapel Hill: University of North Carolina Herbarium; North Carolina Botanical Garden: 2022. Flora of the Southeastern United States. [Google Scholar]
- Weststrand S., Korall P. Phylogeny of Selaginellaceae: there is value in morphology after all. Am. J. Bot. 2016;103:2136–2159. doi: 10.3732/ajb.1600156. [DOI] [PubMed] [Google Scholar]
- Weststrand S., Korall P. Subgeneric classification of Selaginella (Selaginellaceae) Am. J. Bot. 2016;103:2160–2169. doi: 10.3732/ajb.1600288. [DOI] [PubMed] [Google Scholar]
- Wicke S., Schneeweiss G.M., de Pamphilis C.W., et al. The evolution of the plastid chromosome in land plants: gene content, gene order, gene function. Plant Mol. Biol. 2011;76:273–297. doi: 10.1007/s11103-011-9762-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiang Q.-P., Tang J.-Y., Yu J.-G., et al. The evolution of extremely diverged plastomes in Selaginellaceae (lycophyte) is driven by repeat patterns and the underlying DNA maintenance machinery. Plant J. 2022;111:768–784. doi: 10.1111/tpj.15851. [DOI] [PubMed] [Google Scholar]
- Xu Z.-C., Xin T.-Y., Bartels D., et al. Genome analysis of the ancient Tracheophyte Selaginella tamariscina reveals evolutionary features relevant to the acquisition of desiccation tolerance. Mol. Plant. 2018;11:983–994. doi: 10.1016/j.molp.2018.05.003. [DOI] [PubMed] [Google Scholar]
- Yang J., Xiang Q.-P., Zhang X.-C. Uncovering the hidden diversity of the rosette-forming Selaginella tamariscina group based on morphological and molecular data. Taxon. 2023;72:8–19. doi: 10.1002/tax.12817. [DOI] [Google Scholar]
- Yang J.-J., Parris B., Knapp R., et al. Circumscription of the grammitid fern genus Oreogrammitis (Polypodiaceae) with the description of three new genera: Calligrammitis, Devolia, and Glabrigrammitis. Phytotaxa. 2023;597:28–36. [Google Scholar]
- Zhang L.-B., Simmons M.P. Phylogeny and delimitation of the Celastrales inferred from nuclear and chloroplast genes. Syst. Bot. 2006;31:107–121. [Google Scholar]
- Zhang L., Zhang L.-B. Phylogeny, character evolution, and classification of the fern family Ophioglossaceae based on Sanger sequence data, plastomes, and morphology. Mol. Phylogenet. Evol. 2022;173:107512. doi: 10.1016/j.ympev.2022.107512. [DOI] [PubMed] [Google Scholar]
- Zhang L.-B., Zhou X.-M. A new classification of Lycopodiales based on molecular phylogenetics, morphology, and spore ornamentation. Indian Fern J. 2022;38:125–136. [Google Scholar]
- Zhang X.-C., Kato M., Nooteboom H.P. In: Wu Z.-Y., Raven P.H., Hong D.-Y., editors. 2–3. Science Press; Beijing: 2013. Selaginellaceae; pp. 37–66. (Flora of China). (St. Louis: Missouri Botanical Garden Press) [Google Scholar]
- Zhang H.-R., Zhang X.-C., Xiang Q.-P. Directed repeats co-occur with few short-dispersed repeats in plastid genome of a Spikemoss, Selaginella vardei (Selaginellaceae, Lycopodiopsida) BMC Genomics. 2019;20:1–11. doi: 10.1186/s12864-019-5843-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H.-R., Xiang Q.-P., Zhang X.-C. The unique evolutionary trajectory and dynamic conformations of DR and IR/DR-coexisting plastomes of the early vascular plant Selaginellaceae (Lycophyte) Genome Biol. Evol. 2019;11:1258–1274. doi: 10.1093/gbe/evz073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H.-R., Wei R., Xiang Q.-P., et al. Plastome-based phylogenomics resolves the placement of the Sanguinolenta group in the spikemoss of lycophyte (Selaginellaceae) Mol. Phylogenet. Evol. 2020;147:106788. doi: 10.1016/j.ympev.2020.106788. [DOI] [PubMed] [Google Scholar]
- Zhang L., Zhou X.-M., Liang Z.-L., et al. Phylogeny and classification of the tribe Lepisoreae (Polypodiaceae; pteridophyta) with the description of a new genus, Ellipinema gen. nov., segregated from Lepisorus. Mol. Phylogenet. Evol. 2020;148:106803. doi: 10.1016/j.ympev.2020.106803. [DOI] [PubMed] [Google Scholar]
- Zhang M.-H., Wei R., Xiang Q.-P., et al. Integrative taxonomy of the Selaginella helvetica group based on morphological, molecular and ecological data. Taxon. 2021;70:1163–1187. doi: 10.1002/tax.12565. [DOI] [Google Scholar]
- Zhang M.-H., Xiang Q.-P., Zhang X.-C. Plastid phylogenomic analyses of the Selaginella sanguinolenta group (Selaginellaceae) reveal conflict signatures resulting from sequence types, outlier genes, and pervasive RNA editing. Mol. Phylogenet. Evol. 2022;173:107507. doi: 10.1016/j.ympev.2022.107507. [DOI] [PubMed] [Google Scholar]
- Zhao Y.-C., Liu Q., Zhang X.-C. Studies on the Chinese species of Selaginella (II): a preliminary observation on the strobilar organization in Selaginella. J. Beijing Normal Univ. (Nat. Sci.) 2001;37:106–110. [Google Scholar]
- Zhao Y.-Y., Zhang Q.-Y., Jin Y., et al. Spore morphology of five species of Selaginellaceae from Guangxi. Biotechnol. Bull. 2006;18:504–509. [Google Scholar]
- Zhou X.-M., Zhang L.-B. A classification of Selaginella (Selaginellaceae) based on molecular (chloroplast and nuclear), macromorphological, and spore features. Taxon. 2015;64:1117–1140. doi: 10.12705/646.2. [DOI] [Google Scholar]
- Zhou X.-M., He Z.-R., Zhang L., et al. Selaginella chuweimingii (Selaginellaceae) sp. nov. from Yunnan, China. Phytotaxa. 2015;231:283–288. doi: 10.11646/phytotaxa.231.3.6. [DOI] [Google Scholar]
- Zhou X.-M., Jiang L.-J., Zhang L., et al. Spore morphology of Selaginella (Selaginellaceae) from China and its systematic significance. Phytotaxa. 2015;237:1–67. doi: 10.11646/phytotaxa.237.1.1. [DOI] [Google Scholar]
- Zhou X.-M., Rothfels C.J., Zhang L., et al. A large-scale phylogeny of the lycophyte genus Selaginella (Selaginellaceae: lycopodiopsida) based on plastid and nuclear loci. Cladistics. 2016;32:360–389. doi: 10.1111/cla.12136. [DOI] [PubMed] [Google Scholar]
- Zhou X.-M., Zhao J., Yang J.-J., et al. Plastome structure, evolution, and phylogeny of Selaginella. Mol. Phylogenet. Evol. 2022;169:107410. doi: 10.1016/j.ympev.2022.107410. [DOI] [PubMed] [Google Scholar]
- Zhou X.-M., Yang J.-J., Pollawatn R., et al. A global phylogeny of grammitid ferns (Polypodiaceae) and its systematic implications. Taxon. 2023;72 doi: 10.1002/tax.12992. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1
Maximum likelihood phylogeny of Selaginellaceae based on plastid rbcL. Maximum likelihood bootstrap support values are shown along the branches. Red and green names indicate the 7 subfamilies and 19 genera, respectively, recognized in current classification. Blue and black names indicate the subgenera and sections recognized by Zhou and Zhang (2015). Purple names indicate the 6 sections recognized by Zhou and Zhang (2015) and this study at the same rank.
Fig. S2
Maximum likelihood phylogeny of Selaginellaceae based on nuclear (5.8S, 18S, 26S, SQD1, and pgiC) sequences. Maximum likelihood bootstrap support values are shown along the branches. Red and green names indicate the 7 subfamilies and 19 genera, respectively, recognized in current classification. Blue and black names indicate the subgenera and sections recognized by Zhou and Zhang (2015). Purple names indicate the 6 sections recognized by Zhou and Zhang (2015) and this study at the same rank.
Fig. S3
Maximum likelihood phylogeny of Selaginellaceae based on plastid rbcL and nuclear (5.8S, 18S, 26S, SQD1, and pgiC) sequences. Maximum likelihood bootstrap support valuesare shown along the branches. Red and green names indicate the 7 subfamilies and 19 genera, respectively, recognized in current classification. Blue and black names indicate the subgenera and sections recognized by Zhou and Zhang (2015). Purple names indicate the 6 sections recognized by Zhou and Zhang (2015) and this study at the same rank.
Multimedia component 4