Structural Basis of BACH1 Phosphopeptide Recognition by BRCA1 Tandem BRCT Domains (original) (raw)
. Author manuscript; available in PMC: 2013 May 13.
Published in final edited form as: Structure. 2004 Jul;12(7):1137–1146. doi: 10.1016/j.str.2004.06.002
Summary
BRCT tandem domains, found in many proteins involved in DNA damage checkpoint and DNA repair pathways, were recently shown to be phosphopeptide binding motifs. Using solution nuclear magnetic resonance (NMR) spectroscopy and mutational analysis, we have characterized the interaction of BRCA1-BRCT domains with a phosphoserine-containing peptide derived from the DNA repair helicase BACH1. We show that a phenylalanine in the +3 position from the phosphoserine of BACH1 is bound to a conserved hydrophobic pocket formed between the two BRCT domains and that recognition of the phosphate group is mediated by lysine and serine side chains from the amino-terminal BRCT domain. Mutations that prevent phosphopeptide binding abolish BRCA1 function in DNA damage-induced checkpoint control. Our NMR data also reveal a dynamic interaction between BRCA1-BRCT and BACH1, where the bound phosphopeptide exists as an equilibrium of two conformations and where BRCA1-BRCT undergoes a transition to a more rigid conformation upon peptide binding.
Introduction
The BRCT (BRCA1 carboxyl-terminal) motif is an autonomously folded domain of ~100 amino acids first identified as a tandem repeat at the carboxyl terminus of the breast cancer susceptibility protein BRCA1 and subsequently found in a wide array of proteins involved in DNA repair, DNA recombination, and cell cycle control (Bork et al., 1997; Callebaut and Mornon, 1997; Koonin et al., 1996; Miki et al., 1994). In the human genome alone, more than 30 BRCT domain-containing proteins have been identified.
The BRCT domains are essential for the tumor suppressor function of BRCA1. Eighty percent of BRCA1 mutations resulting in a truncated form of this protein (one or both BRCT domains lacking) lead to increased lifetime risk of breast and ovarian cancers (Futreal et al., 1994). Clinically relevant mutations are found within each BRCT motif of BRCA1 and within the BRCA1-interacting domains of some proteins, like the DNA helicase BACH1, involved in DNA repair (Cantor et al., 2001, 2004; Menichini and Linial, 2001).
Recently, we showed that the tandem BRCT domains of BRCA1 bind a specific phosphoserine residue (pSer990) of the DNA repair helicase BACH1 in vitro and in vivo (Yu et al., 2003). We also showed that phospho-dependent BACH1 binding to BRCA1 is cell cycle regulated, with phosphorylated BACH1 being present only in S-G2-M cells, as is the BRCA1/BACH1 complex (Yu et al., 2003). In addition, studies using phosphopeptide libraries revealed that a phenylalanine in the +3 position from the phosphoserine was necessary for the interaction of BRCA1-BRCT with BACH1 (Manke et al., 2003; Rodriguez et al., 2003). Recognition of phosphoserine-as well as phosphothreonine-containing peptides, with a strong preference for aromatic or aliphatic residues in the +3 position, appears to be a general property of phosphopeptide sequences recognized by several other BRCT domains (Manke et al., 2003; Rodriguez et al., 2003; Yu et al., 2003).
Our present study, using NMR spectroscopy and mutational analysis, reveals that the phenylalanine in the +3 position from the phosphoserine of BACH1 binds a hydrophobic pocket at the interface between the two BRCT domains and that the BACH1 phosphate group is recognized by a lysine and a serine residue from the N-terminal BRCT domain. We show that these two BRCA1 residues are essential for the normal functioning of BRCA1 in vivo, particularly in the G2/M checkpoint control of DNA damage. Our work also provides a structural basis for the well-characterized M1775R mutation found in breast and ovarian cancers (Ekblad et al., 2002; Futreal et al., 1994; Williams and Glover, 2003). Importantly, our NMR data reveal a dynamic interaction between BRCA1-BRCT and BACH1, where the bound phosphopeptide exists as an equilibrium of two conformations and where BRCA1-BRCT undergoes a transition to a more rigid conformation upon peptide binding. Motions on the microsecond to millisecond time scale detected in free BRCA1-BRCT are strongly reduced when BACH1 binds.
Results and Discussion
Identification of the Phosphopeptide Binding Site of BRCA1-BRCT
We used solution NMR spectroscopy to characterize the interaction of human BRCA1-BRCT tandem domains (residues 1646–1859) with a phosphorylated peptide derived from the human DNA repair helicase BACH1 (BACH1p; residues 985–997) (Figure 1). For the rest of the text, residues of BACH1p will be numbered from 1 to 13, with the phosphoserine at position 6 indicated by pSer6.
Figure 1. Amino Acid Sequence of BACH1 Phosphopeptide.

The numbering scheme adopted for BACH1 phosphopeptide (BACH1p) is 1 to 13. The sequence corresponds to residues 985–998 of BACH1 protein. The phosphorylated Ser is indicated by pSer6. Relative positions of other residues from pSer6 are also indicated.
BRCA1-BRCT/BACH1p complex has unfavorable NMR relaxation properties that are most likely a consequence of the combined large size (227 amino acids) and prolate ellipsoidal shape of BRCA1-BRCT, leading to slow tumbling in solution. In order to get the data needed for our study, it was necessary to use deuteration and multiple selective labeling techniques (Figure 2).
Figure 2. Variation in BRCA1-BRCT Chemical Shifts upon BACH1p Binding.
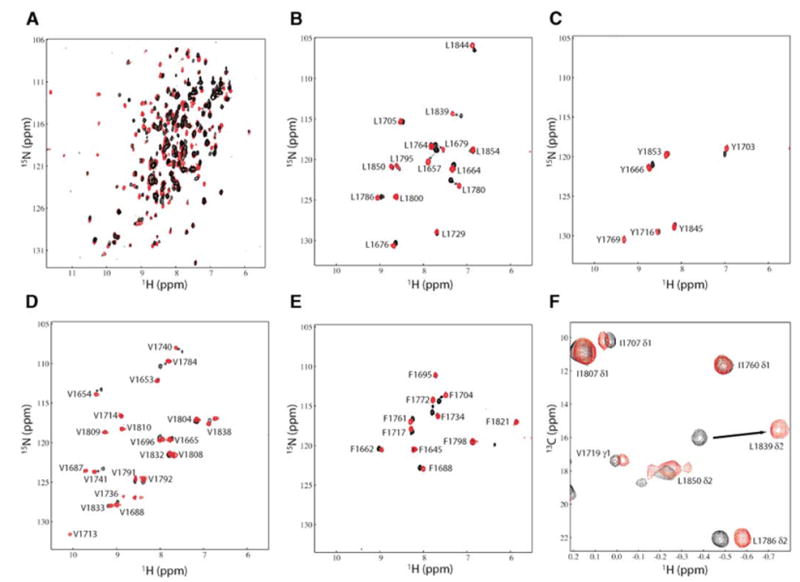
(A) Overlay of 15N-HSQC spectra of 15N/[50%2H]-labeled BRCA1-BRCT, free (black) and in a 1:1 complex with nonlabeled BACH1p (red), illustrating the extent of chemical shift perturbations upon peptide binding.
(B–E) Overlay of 15N-HSQC spectra of BRCA1-BRCT, free (black) and bound to nonlabeled BACH1p (red), selectively 15N-labeled for Leu, Tyr, Val, and Phe, respectively.
(F) Overlay of a selected region of the 13C-HSQC spectrum of 15N/13C/2H-labeled BRCA1-BRCT, but protonated at the methyl groups of Val, Leu, and Ile (δ1), in the absence (black) and presence (red) of nonlabeled BACH1p.
We prepared several BRCA1-BRCT protein samples, namely: (1) uniformly 15N-, 15N/13C-, 15N/[50% 2H]-, 15N/13C/[60% 2H]-, and 15N/13C/2H-labeled; (2) specifically 15N-and 15N/13C-labeled in one amino acid (Val, Leu, Phe, Tyr, Gly, or Ser) at a time; and (3) 15N/13C/2H-labeled in all amino acids except Val, Leu, and Ile (δ1) whose methyl groups are selectively protonated (13C/1H) (Goto et al., 1999; Gross et al., 2003; Hilty et al., 2002).
We were able to assign 91% of backbone HN, N, CO, Cα, and Cβ resonances, 94% of side chain methyls of Val, Leu, and Ile, 47% of aromatic protons of Phe and Tyr, 80% of Trp side chain amide, and a few Thr methyl groups and His ring protons of BRCA1-BRCT in complex with BACH1p. Assignments of methyl groups proved to be very useful in locating the phosphopeptide binding site, as we were able to identify several intermolecular NOEs involving four methyl-containing amino acids of the protein (see below). A nonlabeled BACH1p peptide was used for 1H resonance assignments of its free and bound forms.
We have identified a total of 691 protein intramolecular NOEs of which 453 are medium-range (|i – j| = 1 to 4) and 238 are long-range (|i – j| ≥ 5). The limited number of NOEs observed is due to the relatively small number of protons in the deuterated protein samples used. From NOEs, chemical shift index (CSI) (Wishart and Sykes, 1994), and values of Phi and Psi angles predicted by TALOS (Cornilescu et al., 1999), we were able to identify the secondary structure elements of BRCA1-BRCT in complex with BACH1p. A summary is provided in Figure 3A. Ninety-eight Phi and ninety-eight Psi backbone torsion angles have been derived according to these secondary structure elements. Consistent with the above, we observed 59 slowly exchanging amide protons, the majority of which are involved in hydrogen bonds. The number of NOEs and torsion angles obtained would be insufficient to determine an NMR structure per se. However, with the availability of a 2.5 Å resolution crystal structure of free BRCA1-BRCT (Williams et al., 2001), it was possible to analyze our NMR data. As shown in Figure 3A, the secondary structure elements found for bound BRCA1-BRCT closely match those in the crystal structure of free BRCA-BRCT. In addition, all medium and long-range NOEs of bound BRCA1-BRCT are consistent with the tertiary structure of free BRCA1-BRCT (Figure 3B). Thus, the use of the crystal structure to interpret our NMR data is valid.
Figure 3. Summary of NMR Data for BRCA1-BRCT in Complex with BACH1p.
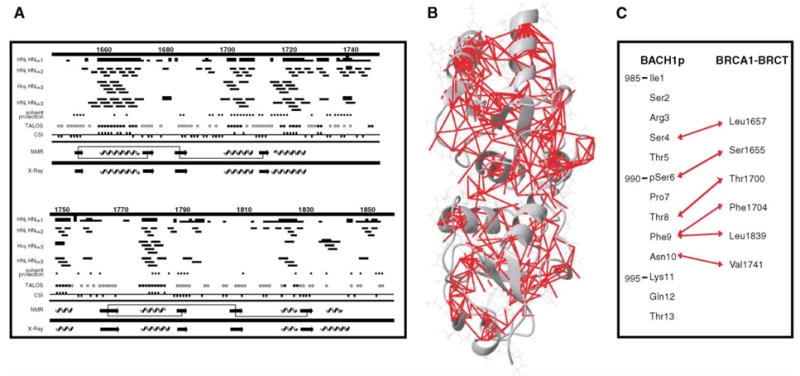
(A) Sequence of BRCA1-BRCT (1646–1859) with a summary of secondary structure elements derived from NMR data of the BACH1p-bound protein: interproton NOEs, solvent protection, TALOS, and chemical shift index (CSI) results. NOEs were measured from 100, 200, and 300 ms mixing times 3D 15N-edited and 13C-edited NOESY data recorded on 50%, 60%, and 100% deuterated 15N- and 15N/13C-labeled BRCA1-BRCT samples, as well as on a 15N/13C-labeled and deuterated (but protonated at Val, Leu, and Ile δ1 methyls) protein. An NOE is indicated by a bar; the thickness is proportional to intensity. A solid circle represents a residue significantly protected from 2H exchange (i.e., 1H signal is still visible in the HSQC spectrum 4 hr after exchange in D2O). Solid and empty squares indicate that Phi and Psi angles predicted by TALOS are compatible with those expected for α helices and β strands, respectively. Negative and positive CSI are designated by a square below and above the line, respectively. β strands (arrows) and α helices (coils) deduced from all the NMR data are summarized (NMR). Thin lines connecting the β strands indicate that long-range HNi-HNj and Hαi-HNj NOEs have been observed. The NMR data are in very good agreement with the secondary structures found in the crystal structure of free BRCA1-BRCT (X-ray).
(B) Mapping of interresidue NOEs on the crystal structure of free BRCA1-BRCT (PDB entry 1JNX). Protons involved in NOEs are connected by red lines; 453 medium-range and 238 long-range NOEs are shown.
(C) Residues involved in intermolecular NOEs are connected by red double arrows.
An attempt to identify the region of BRCA1-BRCT that contacts BACH1p was first made by monitoring the changes in 15N, 13C, and 1H chemical shifts in the 15N-and 13C-heteronuclear single-quantum coherence (HSQC) spectra of BRCA1-BRCT in the absence and in the presence of BACH1p (Figure 2). The changes in the 15N- HSQC spectra revealed that the exchange between the free and bound states of BRCA1-BRCT is slow on the NMR chemical shift time scale, as two signals corresponding to free and bound states were observed during titration. This is consistent with a dissociation constant of 0.7 ± 0.1 μM that we determined by isothermal titration calorimetry (ITC) at a temperature, pH, and ionic strength identical to those used in our NMR experiments. A surprisingly large number of BRCA1-BRCT resonances (at least 49) are affected by the interaction with BACH1p. These chemical shift perturbations occur not only for residues at the surface, but for those in the interior of BRCA1-BRCT as well, suggesting that there is a change in conformation of the protein upon peptide binding. One should note, however, that even if a large number of resonances are altered after interaction with BACH1p, residues that are away from the binding site exhibit small variations in chemical shifts (chemical shifts change by less than 0.05 and 0.25 ppm in the proton and nitrogen dimensions, respectively), indicating that any change in conformation would be of small amplitude.
A molecular representation of BRCA1-BRCT highlighting surface residues with changes in chemical shifts due to BACH1p binding is shown in Figures 4A and 4B. The affected residues, the majority of which belong to the first BRCT domain, the BRCT-BRCT interface, and the N-terminal half of the inter-BRCT linker region (residues 1737–1759), localize mainly on one side of the protein.
Figure 4. Identification of the BACH1p Binding Site on BRCA1-BRCT.
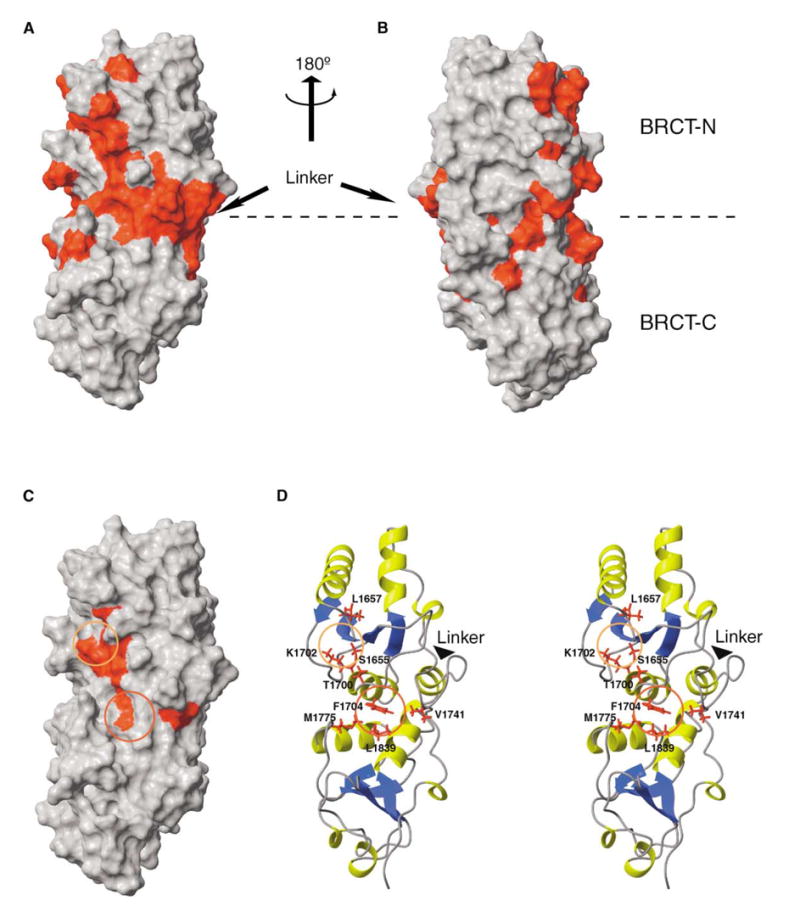
(A) Changes in chemical shifts mapped on the crystal structure of free BRCA-BRCT. Shown in red are residues that are affected upon BACH1p binding.
(B) Same as (A) after 180° rotation about the vertical axis.
(C) Residues involved in intermolecular NOEs with BACH1p mapped on the crystal structure of free BRCA-BRCT. The region involved in phosphoserine binding is circled in yellow, while the hydrophobic pocket is indicated by a red circle.
(D) Stereo ribbon representation of BRCA1-BRCT showing the amino acids involved in intermolecular NOEs, as well as Met1775 and Lys1702. Red and yellow circles are defined as in (C).
To more precisely locate the peptide binding site, we analyzed the intermolecular NOEs found between BRCA1-BRCT and BACH1p. We have identified 25 intermolecular NOEs involving Ser4, pSer6, Thr8, Phe9, and Asn10 of BACH1p and several residues of BRCA1-BRCT. The pairs of protein-peptide residues for which intermolecular NOEs have been observed are shown in Figure 3C. Some of these NOEs are illustrated in Figures 5C and 5D. The aromatic ring and β protons of Phe9 from BACH1p have NOEs with the methyl groups of Leu1839 and the aromatic ring protons of Phe1704 from BRCA1-BRCT. These contacts position Phe9 very well to a predominantly hydrophobic pocket formed by Phe1704 and Leu1839 near the inter-BRCT domain inter- face (Figures 4C and 4D). Consistent with this, the HN signals of Phe1704 and L1839 undergo large chemical shift changes from the free to the bound state (Figure 2). Furthermore, as shown in Figure 2F, the δ2 methyl group of L1839 has a very upfield shifted chemical shift in the complex, probably due to the ring current effects of nearby Phe1704 and Phe9. The favorable hydrophobic interaction described above provides a rationale for why an aromatic residue in the +3 position (Phe9) results in tight interaction of BACH1p with BRCA1-BRCT. Other intermolecular NOEs were identified that involve Ser4, pSer6, Thr8, and Asn10 of BACH1p and Leu1657, Ser1655, Thr1700, and Val1741 of BRCA1-BRCT, respectively (Figure 3C). The close contact between Thr8 and Thr1700 could explain the exclusive selection of a methyl group-containing amino acid in the +2 position from the phosphoserine as determined by phosphopeptide library screens (Rodriguez et al., 2003) (Figure 5D). Taken together, our NOE data place the phosphate group of BACH1p (pSer6) in close proximity to the hydroxyl group of Ser1655 and side chain amino group of Lys1702 (Figure 4D). The hydroxyl group of Ser1655 and side chain amino group of Lys1702 are very likely to be involved in hydrogen bonds and/or charge/charge interaction with the phosphate. Coincidentally, Lys1702 is the only charged amino acid in this region of BRCA1. Mutation of either Ser1655 or Lys1702 to an alanine results in a loss of interaction with BACH1p and a loss of function of BRCA1 in DNA damage-induced checkpoint control (see mutational studies below).
Figure 5. Evidence of Conformational Equilibrium in Bound BACH1p.
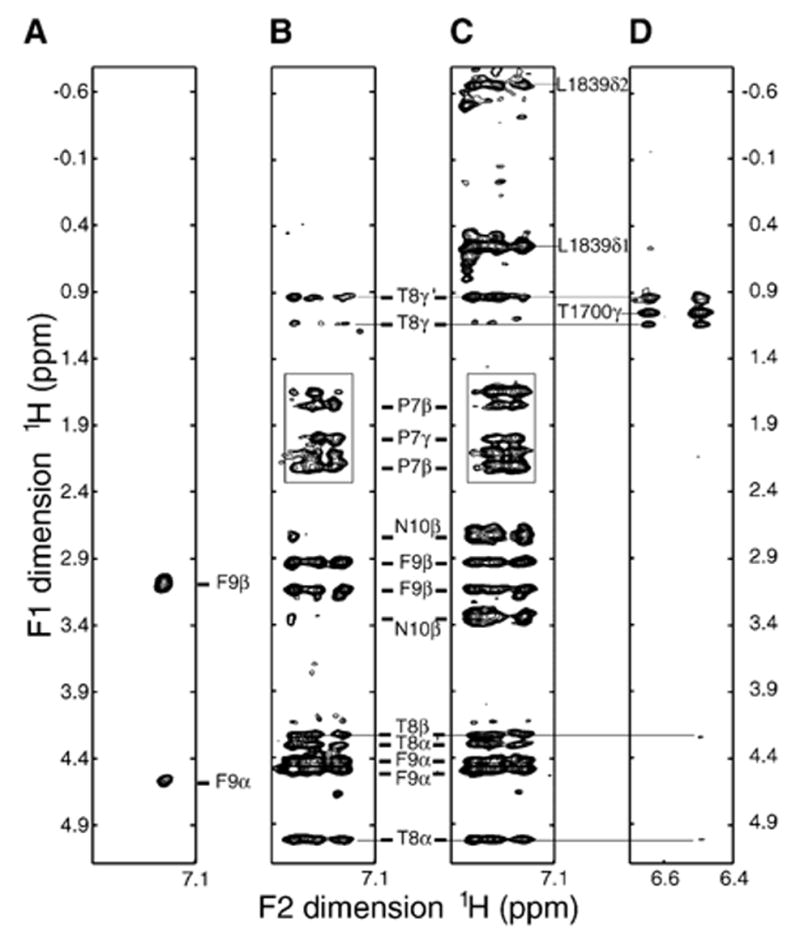
Regions from the 2D 1H-1H NOESY (A), F1/F2 double-filtered NOESY (B), F2-filtered NOESY (C), and F2 15N-edited NOESY (D) spectra of the free BACH1p peptide (A) and BRCA1-BRCT/BACH1p complex (B–D). BACH1p peptide was nonlabeled in (A) to (D). BRCA1-BRCT was 15N/13C/2H-labeled with protonated methyls (Val, Leu, and Ile δ1) in (B) and (C) and 15N/13C/[60%-2H]-labeled in (D). The F2 dimension in (A)–(C) corresponds to the aromatic region of BACH1p Phe9. The F2 dimension in (D) corresponds to the HN of Thr1700 without 15N decoupling. Experiments were done in D2O at 30°C and pH 7.5. Thr1700 HN is protected from exchange with D2O as seen in (D). (A) and (B) show intrapeptide NOEs. (C) displays both intrapeptide and peptide/protein NOEs. (D) contains intraprotein and peptide/protein NOEs. Note two sets of signals for Phe9 Hα, Thr8 Hβ, and Thr8 Hγ indicating two conformations of bound BACH1p in (B)–(D). In one of these conformations, Phe 9 is close in space to Pro7, as indicated by a box in (B) and (C).
Where the phosphopeptide binds explains the effect of missense mutation M1775R, which prevents BRCA1/BACH1 interaction and which is linked to breast and ovarian cancers (Cantor et al., 2001; Manke et al., 2003). Met1775 is close to Phe1704 and Leu1839, for which intermolecular NOEs with Phe9 of BACH1p have been identified (Figures 4D and 5C). This M1775R mutation very likely prevents Phe9 binding to the hydrophobic pocket of BRCA1 because of steric hindrance by the bulky guanidinium group of arginine. This can be seen from the spatial orientation of the arginine side chain in the crystal structure of this BRCA1-BRCT mutant (Williams and Glover, 2003). The guanidinium protons of Arg1775 are positioned 3–6 Å away from the closest protons of Thr1700, Phe1704, and Leu1839. These three residues have intermolecular NOEs with BACH1p (Figure 3C).
Conformational Equilibrium in BACH1 Phosphopeptide
The pattern of intermolecular NOEs observed for BACH1p in the presence of BRCA1-BRCT is consistent with the peptide having a predominantly extended structure in the bound state (Figure 3C). The downfield shifted Hα chemical shifts of several amino acids of BACH1p are also indicative of this conformation. However, careful examination of intrapeptide and protein-peptide NOEs clearly shows that bound BACH1p exists in equilibrium between two conformations (Figure 5). While the signals could not be distinguished as belonging to one conformer or the other, it is clear that there are two sets of 1H chemical shifts for the following BACH1p residues: Pro7, Thr8, Phe9, Asn10, Lys11, and Gln12. Similar results were obtained for a shorter version of BACH1p (residues 3–11) when titrated with BRCA1-BRCT. An example of intermolecular NOEs involving Thr1700 of BRCA1-BRCT and the Thr8 methyl groups of the two BACH1p conformations is shown in Figure 5D.
The origin of this conformational equilibrium is probably the result of cis/trans isomerization of the pSer6-Pro7 peptidyl bond in BACH1p upon binding BRCA1-BRCT. In free BACH1p, only 10% of the pSer6-Pro7 peptidyl bond is in a cis conformation, while in the complex two BACH1p conformations coexist in an approximate ratio of 1:1. In only one of the two bound conformations, the phenyl ring of Phe9 is close in space to Pro7, as shown by multiple NOEs (Figures 5B and 5C). The observation that two conformations coexist in bound BACH1p is intriguing. Proline-containing motifs are important ligands in several other signaling domains, like SH3 and WW domains (Koch et al., 1991; Lu, 2004; Reimer and Fischer, 2002; Staub and Rotin, 1996). In those cases, however, only one isomer binds the protein.
Dynamics of BRCA1/BACH1 Interaction
A very interesting aspect of BRCA1/BACH1 interaction is the altered BRCA1 dynamics upon BACH1 peptide binding, as evidenced by an approximately 25% increase in integral intensity of the 15N-HSQC spectrum of the complex. The smaller integral intensity for free BRCA1-BRCT reflects a less efficient 15N-1H magnetization transfer due to faster transverse proton relaxation (broader lines). It is known that the efficacy of magnetization transfer in heteronuclear experiments strongly depends on the linewidth of proton resonances (Macura et al., 1985).
The broad proton resonances in the free protein are indicative of a conformational exchange process by slow motions on the microsecond to millisecond time scale (Botuyan et al., 2001; Kern and Zuiderweg, 2003). We ruled out the possibility of aggregation that would contribute to faster relaxation in the free state, as both free and complexed BRCA1-BRCT have similar elution profiles of a monomeric species by gel filtration chromatography. Broadening is particularly evident in the inter-BRCT linker region, which is consistent with this region having poorly defined electron density in the crystal structure of free BRCA1-BRCT (Williams et al., 2001). Virtually all HN signals for which a change in chemical shift is observed upon peptide binding display exchange broadening in free BRCA1-BRCT. The sharpening of resonance lines after addition of BACH1p strongly suggests that the protein becomes more rigid in the bound state.
The change in linewidth is particularly striking for the hydroxyl proton resonance of Tyr1703, which is extremely broad in free BRCA1-BRCT but becomes a sharp signal at 12.3 ppm in the complex (Figure 6A). Narrowing of this Tyr1703 hydroxyl signal constitutes strong evidence of its participation in hydrogen bonding. The aromatic ring of Tyr1703 is buried at the interface of the two BRCT domains, with the hydroxyl group well positioned to form a hydrogen bond with the side chain carboxylate of Asp1739 from the inter-BRCT linker region (Figure 6B). Since Tyr1703 is located next to Lys1702 and Phe1704, which we determined to be two key residues for phosphopeptide binding, the hydrogen bond between Tyr1703 and Asp 1739 would bridge the phosphopeptide binding site to the linker region located on the opposite side of the inter-BRCT interface (Figure 6B). We propose that the network of hydrogen bonds (Tyr1703-Asp1739, Ser1655/Lys1702-pSer6), established upon complex formation, triggers an allosteric change in conformation, or ordering, of the linker region. BRCA1 is known to participate in large molecular assemblies at the site of DNA damage. Such a conformational switch could be a mechanism for coupling phosphoprotein binding to other BRCA1-mediated interactions. It is noteworthy that 53BP1-BRCT, which is structurally very similar to BRCA1-BRCT and which is thought to bind phosphopeptides (Yu et al., 2003), uses its inter-BRCT linker region to interact with p53 (Ekblad et al., 2004; Joo et al., 2002).
Figure 6. Tyr1703 Connnects the Phophopeptide Binding Site to the Inter-BRCT Linker.
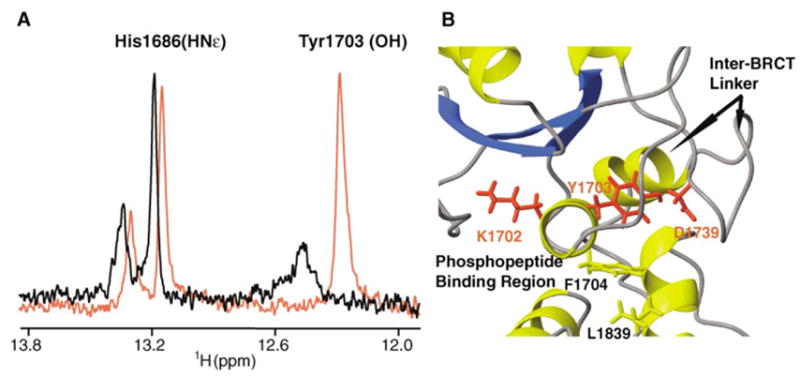
(A) Superposition of a selected region from the 1H spectra of 15N/13C/2H-labeled BRCA1-BRCT, free (black) and bound to nonlabeled BACH1p (red), showing the hydroxyl proton of Tyr1703. Note that other 1H signals in the spectra are 15N-coupled. Experiments were recorded at 30°C and pH 7.5.
(B) Location of Tyr1703 in the 3D structure of BRCA1-BRCT. Tyr1703 bridges the phosphopeptide binding site to the inter-BRCT linker region.
Effects of Mutations of Key Residues for Phosphopeptide Binding
To verify that key phosphate-interacting residues identified by NMR spectroscopy are required for mediating phospho-dependent BRCA1/BACH1 interaction, Ser1655 and Lys1702 were changed to alanine. Judging from the corresponding 15N-HSQC spectra and gel filtration chromatograms, these two mutations do not affect the integrity of the 3D structure (data not shown). An in vitro GST pull-down assay showed that only wild-type BRCA1-BRCT domain, but neither S1655A nor K1702A mutant, interacted with BACH1 (Figure 7A). We also expressed wild-type and mutants of BRCA1 in 293T cells and showed that only wild-type BRCA1 coimmunoprecipitated with BACH1 (Figure 7B). These data demonstrate that Ser1655 and Lys1702 are essential for the phospho-dependent BRCA1/BACH1 interaction, both in vitro and in vivo.
Figure 7. S1655A and K1702A Mutants of BRCA1 Do Not Interact with BACH1 In Vitro and In Vivo.
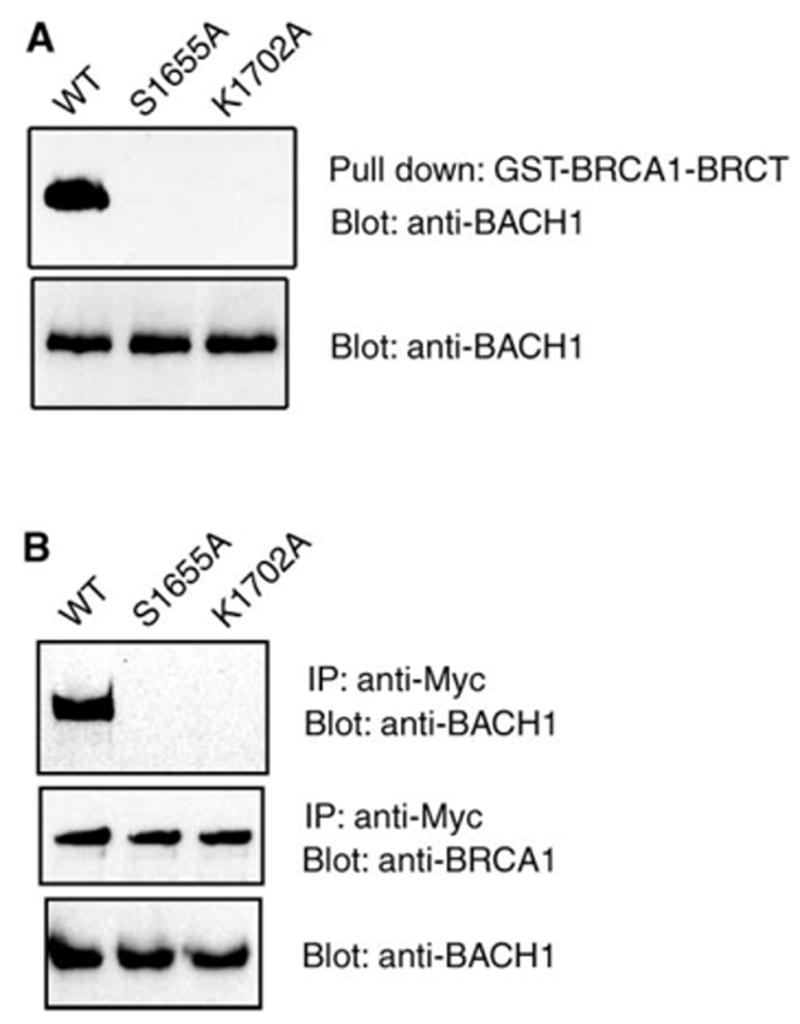
(A) 293T cell lysates were incubated with wild-type or mutant GST-BRCA1-BRCT proteins immobilized on Sepharose beads. Proteins bound to beads were eluted and separated by SDS-PAGE. Western blots were performed with anti-BACH1 antibody (upper panel). Western blots of whole-cell lysates were also included to demonstrate equal amounts of BACH1 in these samples (lower panel).
(B) Myc epitope-tagged wild-type and mutant BRCA1 were transiently expressed in 293T cells. Immunoprecipitation and immunoblotting experiments were performed with anti-myc or anti-BACH1 antibodies as indicated.
Next we examined whether the S1655A and K1702A mutants would affect BRCA1 function. Normal cells treated with ionizing radiation arrest in the G2 phase of the cell cycle and do not enter mitosis until DNA repair is completed. It has been shown that BRCA1/BACH1 interaction is required for this DNA damage-induced checkpoint control (Yu et al., 2003). To test if S1655A or K1702A mutation would affect BRCA1 checkpoint function, we used BRCA1-deficient HCC1937 cells that have G2/M checkpoint defects (Xu et al., 2001; Yarden et al., 2002; Yu et al., 2003). Expression of exogenous wild-type BRCA1 restored this G2/M checkpoint in HCC1937 cells, but S1655A and Lys1702A mutant proteins did not (Figure 8A).
Figure 8. Phosphoprotein Binding Activity of BRCA1 Is Required for DNA Damage-Induced G2/M Checkpoint Control and BRCA1 Nuclear Foci Formation.
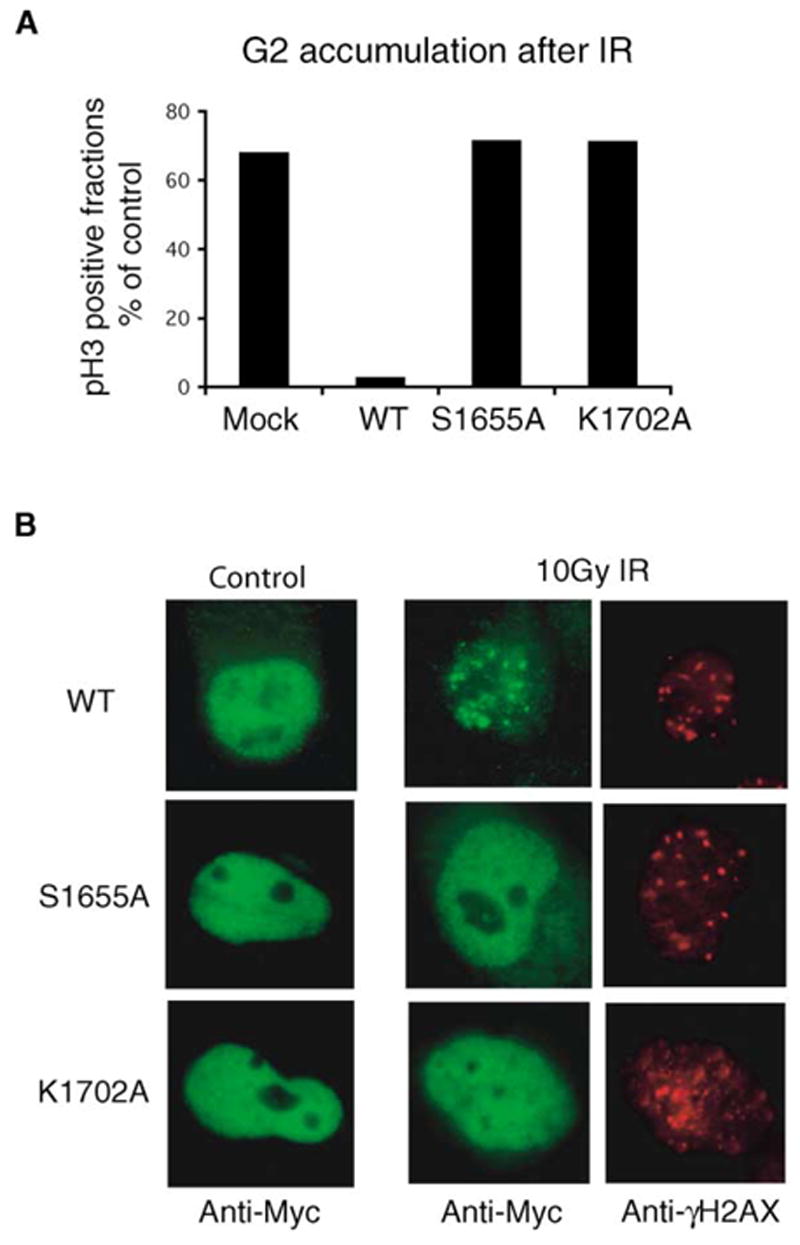
Wild-type or mutant BRCA1 was transiently expressed in HCC1937 cells.
(A) G2/M checkpoint assays were conducted as described in the Experimental Procedures. The fraction of cells in M phase is expressed as a percentage of that measured in unirradiated control cells.
(B) Three hours after γ-irradiation, immunostaining experiments were performed with the indicated antibodies.
The BRCA1-BRCT domains are necessary for the relocalization of BRCA1 to the sites of DNA damage following γ irradiation (Manke et al., 2003). However, only wild-type BRCA1, but neither S1655A nor K1702A mutant, translocated to nuclear foci following radiation and colocalized with γhiston2AX, a marker for the sites of DNA breaks (Figure 8B). These results confirm that Ser1655 and Lys1702, both involved in phosphate group recognition, are essential for BRCA1/BACH1 interaction and are equally required for the in vivo function of BRCA1.
Conclusion
Following up on our recent finding that phosphorylated DNA repair helicase BACH1 forms a complex with BRCA1-BRCT (Yu et al., 2003), we have characterized this interaction using NMR spectroscopy in combination with biochemical and cell biology assays. Our study explains why two BRCT domains are required for binding BACH1 phosphopeptide, as residues from both BRCT domains are needed to form a hydrophobic pocket that accommodates a key phenylalanine (Phe9) of BACH1p. We show that two residues from the first BRCT domain (Ser1655 and Lys1702) mediate phosphate recognition. Their mutation to alanine, while not affecting the overall structure of the protein, disrupts the ability of BRCA1 to bind BACH1 phosphopeptide and inhibits BRCA1 nuclear foci formation and DNA damage-induced G2/M checkpoint control. This work also reveals that a cancer-causing missense mutation alters the phosphopeptide binding site of BRCA1.
The availability of NMR spectral data on BRCA1-BRCT in complex with a phosphopeptide could be useful in high-throughput screening for inhibitors of BRCA1/phosphoprotein interaction using the structure-activity relationship (SAR) NMR method (Hajduk et al., 1999). Because BRCA1 plays an important role in DNA repair, inhibitors of BRCA1 interactions could enhance the effect of chemotherapy and radiation in killing tumor cells.
Our study of BRCA1-BRCT/BACH1p complex will be relevant in understanding the mode of action of other proteins, such as MDC1, BARD1, PTIP, Rad9, and 53BP1, all of which contain tandem BRCT domains and all of which bind phosphopeptides (Manke et al., 2003; Rodriguez et al., 2003; Yu et al., 2003). Our previous work suggested that single BRCT-containing proteins, like FCP1, could also bind phosphopeptides (Yu et al., 2003). Sequence alignment with the first BRCT domain of BRCA1 indicates that Ser1655, Thr1700, and Lys1702, which are key residues for phosphopeptide recognition by BRCA1, are conserved in FCP1. A number of studies have shown that isolated BRCT domains often form homodimers (Caldecott, 2003; Krishnan et al., 2001; Zhang et al., 1998). It would not be surprising if FCP1 would form a homodimer that could bind a phosphopeptide in the same way as the tandem BRCT domains of BRCA1.
Experimental Procedures
Cloning
The cDNA fragments encoding wild-type and various mutant (Y1703F, S1655A, K1702A) BRCA1-BRCT (residues 1646–1859) proteins were incorporated into a modified version of pET43.1a (Novagen) vector, resulting in proteins that have NusA-polyhistidetag at the N terminus cleavable by TEV protease.
Protein Labeling
Uniformly labeled BRCA1 proteins were overexpressed in E. coli BL21(DE3) host grown in M9 minimal media. Depending on the type of labeling (15N, 15N/2H, 15N/13C, 15N/13C/2H), the M9 media was supplemented with various isotopes at concentrations of 1 g/l 15NH4Cl, 3 g/l 13C-glucose, and 3 g/l 13C/2H-glucose. Isotopically 15N-, 15N/13C-, or 15N/13C/2H-enriched media was also added at 10% (v/v) (Silantes-OD media) or 0.1% (w/v) (Isogro media) in the large-scale 1 liter protein production. The amount of D2O added to the M9 media was varied depending on the level of deuteration desired (Gardner and Kay, 1998; Goto et al., 1999). Isotopes were purchased from Isotec, Cambridge Isotope Laboratories, and Silantes.
In a typical 15N and/or 13C labeling, freshly transformed BL21(DE3) cells were first grown in 3 ml LB media at 37°C. Once an OD600 of about 0.5 was reached, the cells were centrifuged and the bacterial pellet resuspended in 25 ml of fresh M9 media. The culture was incubated until an OD600 of 0.5 and then transferred to a 1 liter M9 media. Cell growth was continued up to an OD600 of 0.6–0.9, after which the culture was transferred to a 15°C shaker incubator and incubated for an additional hour before 0.2 mM IPTG (final concentration) was added. The cells were harvested 16 hr post induction by centrifugation at 4000 × g for 20 min. When deuteration was performed in addition to 15N and/or 13C labeling, 15N/13C/2H or 15N/2H isotopically enriched media was substituted for the LB starter media utilized above. The use of this media at the outset greatly facilitates bacterial adjustment to D2O, avoiding additional and longer M9 rein-oculations described in the literature. Protein induction was the same as above.
For selective labeling of individual amino acids, 1 liter of M9 media was prepared containing 1 g each of adenine, guanosine, and uracil; 0.4 g of thymine and cytosine; 50–100 mg of the 15N and/or 13C-labeled amino acid; and 100 mg of all other amino acids (nonlabeled) (Vadrevu et al., 2003). The auxotrophic bacterial strain CT19 was used to selectively label valine, leucine, tyrosine, and phenylalanine one at a time (Waugh, 1996). Selective labeling of glycine and serine was done with BL21(DE3).
Finally, we also prepared 15N/13C/2H-labeled protein selectively protonated at the two methyl groups of valine and leucine, and the δ1 methyl group of isoleucine. The procedure is similar to uniform labeling except that inclusion of 100 mg/l each of [3-2H]/13C α-ketoisovalerate (to label leucine and valine methyls) and/or [3,3-2H2]/13C α-ketobutyrate (to label isoleucine δ1 methyl) was done just prior to induction (Goto et al., 1999; Hilty et al., 2002).
Protein Purification
The cells were resuspended in 50 mM sodium phosphate buffer (pH 7.5) containing 300 mM NaCl (bind buffer), lysed with a high-pressure microfluidizer Emulsiflex C-5 (Avestin), and then centrifuged at 20,000 × g for 30 min to isolate the supernatant fraction. The cell lysate was next loaded onto a nickel-NTA column (Qiagen), washed, and then eluted with bind buffer containing 20 mM and 250 mM imidazole, respectively. The NusA-polyhistidine tag was then cleaved with TEV protease overnight at room temperature. Using the original bind buffer without any imidazole for size exclusion chromatography, the protein was further purified by passing through a preparative Superdex 75 column (Amersham Biosciences). Deuterated DTT was added to the protein sample to a final concentration of 1 mM.
NMR Samples
The purified BRCA1-BRCT protein was then concentrated using 10 kDa MW cut-off centriplus and centricon (Millipore) until the concentration was ~0.5–0.7 mM. The final NMR buffer for BRCA1-BRCT protein was 50 mM sodium phosphate (pH 7.5), 300 mM NaCl, 1 mM DTT, 7% D2O, and 93% H2O. In some cases where the protein (free or bound) needed to be in 100% D2O, the sample was lyophilized and exchanged in pure D2O. The various protein-peptide complexes were prepared by titrating labeled BRCA1-BRCT with a solution (same buffer) of nonlabeled BACH1p and monitoring the 15N-HSQC spectra of the protein. All the complexes used for the NMR experiments approximate a 1:1 protein:peptide ratio.
NMR Spectroscopy
NMR data were acquired on a 600 MHz Bruker Avance spectrometer equipped with a cryoprobe. All experiments were carried out at 30°C. To assign protein resonances, the following experiments were acquired: HNCO, HN(CA)CO, HNCA, HN(CO)CA, HNCACB, HN(CA)CB, HN(CO)CACB, HN(COCA)CB, and HCCH-TOCSY (Cavanagh et al., 1996). An HC(CO)NH-TOCSY optimized for assignment of methyl groups (Val, Leu, and Ile) that are protonated in an otherwise deuterated 15N-, 13C-labeled protein was recorded (Gardner et al., 1996; Hilty et al., 2002). Three-dimensional 15N- and 13C-edited and filter-edited NOESY-HSQC and 15N-edited NOESY-TROSY (Pervushin et al., 1997) experiments with 100, 200, and 300 ms mixing times were also performed to aid in the assignment of side chains and generate NOEs. High-resolution 2D 1H-1H NOESY (100 and 200 ms mixing times) and TOCSY (60 ms mixing time) experiments were collected especially to identify intrapeptide and gain additional protein-peptide intermolecular NOEs (Ikura and Bax, 1992; Zwahlen et al., 1997). A 3D F1-edited 13C F2-filtered HMQC-NOESY was also recorded (Lee et al., 1994). To confirm assignments for valine, leucine, phenylalanine, tyrosine, glycine, and serine and to facilitate the monitoring of chemical shift changes, HSQCs were performed on protein samples 15N- and/or 13C-labeled for these residues, in the absence and in the presence of BACH1p. Data were processed with the NMRPipe/NMRDraw (Delaglio et al., 1995) software and analyzed with the PIPP program (Garrett et al., 1991) and NMRView (Johnson and Blevins, 1994) analysis package on PC Linux and SGI workstations. A software extension of NMRview, NVAsn, was utilized to facilitate NOE assignments. TALOS was run using Cα and CO chemical shifts to predict Phi and Psi angles (Cornilescu et al., 1999). Cα and CO chemical shifts were used to calculate the chemical shift index (CSI) (Wishart and Sykes, 1994). A low-definition structure was calculated using CYANA (Guntert et al., 1997) with 691 NOEs, excluding intra-residue NOEs, and 196 torsion angle constraints deduced from secondary structure analysis, only for the purpose of showing that secondary and tertiary structures of the bound protein are similar to the crystal structure of free BRCA1-BRCT. All covalent parameters in CYANA are fixed at their ideal values throughout the calculation in torsion angle space. For the ten best structures calculated, there were no distance and angle violations greater than 0.40 ± 0.05 Å and 4.5 ± 0.5°, respectively. All molecular representations were generated using MOLMOL (Koradi et al., 1996).
Isothermal Titration Calorimetry
All ITC measurements were recorded using a VP-ITC instrument (MicroCal) and acquired at 30°C on samples containing 20 μM BRCA1-BRCT (starting concentration), 50 mM sodium phosphate buffer (pH 7.5), and 300 mM NaCl. Incremental amounts of an 800 μM solution of BACH1 phosphopeptide were added. All data analyses were performed using the ORIGIN software (version 7) from MicroCal.
Cell Culture, Antibodies, and Plasmids
All cell lines were maintained in RPMI 1640 medium with 10% fetal calf serum at 37°C with 5% CO2 (v/v). Rabbit anti-BACH1 and γH2AX antibodies were generated as described previously (Rogakou et al., 1999; Yu et al., 2003). Constructs of GST-BRCA1-BRCT and pβmyc-BRCA1 were generated as described previously (Yu et al., 2003). All BRCA1 mutants were made using the Quickchange site-directed mutangenesis kit (Invitrogen).
GST Pull-Down Assay, Immunoprecipitation, and Immunoblotting
5 μg plasmid with 15 μl FuGENE 6 was used for transfection according to the protocol provided by the manufacturer (Roche Diagnostics). GST pull-down assay, immunoprecipitation, and immunoblotting were performed as described previously (Yu et al., 2003).
G2/M Accumulation Assay and Study of Damage-Induced BRCA1 Localization
HCC1937 cells were transiently transfected with plasmids encoding wild-type or mutant BRCA1. The radiation-induced G2/M accumulation assay was performed as described previously (Yu et al., 2003). For the study of BRCA1 relocalization following DNA damage, transfected HCC1937 cells were mock treated or exposed to 10 Gy γ irradiation. Cells were fixed 3 hr later with 3% paraformaldehyde and costained with anti-Myc (9E10) and anti-γH2AX antibodies, followed by incubation with fluorescence-conjugated secondary antibodies.
Acknowledgments
We gratefully acknowledge Drs. Gerard Kroon, Micah Gearhart, and Dan Garrett for help with NMR analysis software, Drs. Walter Chazin, Shibani Bhattacharya, and Jaison Jacobs for pulse programs, Drs. David Waugh and David LeMaster for providing auxotrophic bacterial strains, and Dr. Ramakrishna Vadrevu for advice on selective labeling. This work is supported by grants from the National Institutes of Health to J.C. (RO1 CA89239) and to G.M. (RO1 CA109449). J.C. is a recipient of a DOD breast cancer career development award (DAMD17-02-1-0472).
Footnotes
Note Added in Proof
While our manuscript was under review, three reports on crystal structures of BRCA1-BRCT/phosphopeptide complexes were published. Two of these structures are in complex with a BACH1 phosphopeptide similar to what is presented in our publication (Clapperton et al., 2004; Shiozaki et al., 2004), while the third structure is BRCA1-BRCT bound to a peptide derived from a phosphopeptide library screen (Williams et al., 2004). It is noteworthy that all the intermolecular contacts identified using NMR spectroscopy are consistent with the two crystal structures of BRCA1-BRCT/BACH1 phosphopeptide complex. Our study is a good illustration of the complementarity between NMR spectroscopy and X-ray crystallography. While NMR shows that the bound BACH1 peptide is in equilibrium between two conformations, only one peptide conformation is observed in all three crystal structures.
References
- Bork P, Hofmann K, Bucher P, Neuwald AF, Altschul SF, Koonin EV. A superfamily of conserved domains in DNA damage-responsive cell cycle checkpoint proteins. FASEB J. 1997;11:68–76. [PubMed] [Google Scholar]
- Botuyan MVE, Mer G, Yi GS, Koth CM, Case DA, Edwards AM, Chazin WJ, Arrowsmith CH. Solution structure and dynamics of yeast elongin C in complex with a von Hippel-Lindau peptide. J Mol Biol. 2001;312:177–186. doi: 10.1006/jmbi.2001.4938. [DOI] [PubMed] [Google Scholar]
- Caldecott KW. The BRCT domain: signaling with friends? Science. 2003;302:579–580. doi: 10.1126/science.1091463. [DOI] [PubMed] [Google Scholar]
- Callebaut I, Mornon JP. From BRCA1 to RAP1: a widespread BRCT module closely associated with DNA repair. FEBS Lett. 1997;400:25–30. doi: 10.1016/s0014-5793(96)01312-9. [DOI] [PubMed] [Google Scholar]
- Cantor SB, Bell DW, Ganesan S, Kass EM, Drapkin R, Grossman S, Wahrer DC, Sgroi DC, Lane WS, Haber DA, Livingston DM. BACH1, a novel helicase-like protein, interacts directly with BRCA1 and contributes to its DNA repair function. Cell. 2001;105:149–160. doi: 10.1016/s0092-8674(01)00304-x. [DOI] [PubMed] [Google Scholar]
- Cantor S, Drapkin R, Zhang F, Lin Y, Han J, Pamidi S, Livingston DM. The BRCA1-associated protein BACH1 is a DNA helicase targeted by clinically relevant inactivating mutations. Proc Natl Acad Sci USA. 2004;101:2357–2362. doi: 10.1073/pnas.0308717101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanagh J, Fairbrother WJ, Palmer AG, Skelton NJ. Protein NMR Spectroscopy: Principles and Practice. San Diego, CA: Academic Press; 1996. [Google Scholar]
- Clapperton JA, Manke IA, Lowery DM, Ho T, Haire LF, Yaffe MB, Smerdon SJ. Structure and mechanism of BRCA1 BRCT domain recognition of phosphorylated BACH1 with implications for cancer. Nat Struct Mol Biol. 2004;11:512–518. doi: 10.1038/nsmb775. [DOI] [PubMed] [Google Scholar]
- Cornilescu G, Delaglio F, Bax A. Protein backbone angle restraints from searching a database for chemical shift and sequence homology. J Biomol NMR. 1999;13:289–302. doi: 10.1023/a:1008392405740. [DOI] [PubMed] [Google Scholar]
- Delaglio F, Grzesiek S, Vuister GW, Zhu G, Pfeifer J, Bax A. NMRPipe: a multidimensional spectral processing system based on UNIX pipes. J Biomol NMR. 1995;6:277–293. doi: 10.1007/BF00197809. [DOI] [PubMed] [Google Scholar]
- Ekblad CM, Wilkinson HR, Schymkowitz JW, Rousseau F, Freund SM, Itzhaki LS. Characterisation of the BRCT domains of the breast cancer susceptibility gene product BRCA1. J Mol Biol. 2002;320:431–442. doi: 10.1016/s0022-2836(02)00478-3. [DOI] [PubMed] [Google Scholar]
- Ekblad CM, Friedler A, Veprintsev D, Weinberg RL, Itzhaki LS. Comparison of BRCT domains of BRCA1 and 53BP1: a biophysical analysis. Protein Sci. 2004;13:617–625. doi: 10.1110/ps.03461404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Futreal PA, Liu Q, Shattuck-Eidens D, Cochran C, Harshman K, Tavtigian S, Bennett LM, Haugen-Strano A, Swensen J, Miki Y, et al. BRCA1 mutations in primary breast and ovarian carcinomas. Science. 1994;266:120–122. doi: 10.1126/science.7939630. [DOI] [PubMed] [Google Scholar]
- Gardner KH, Kay LE. The use of 2H, 13C, 15N multidimensional NMR to study the structure and dynamics of proteins. Annu Rev Biophys Biomol Struct. 1998;27:357–406. doi: 10.1146/annurev.biophys.27.1.357. [DOI] [PubMed] [Google Scholar]
- Gardner KH, Konrat R, Rosen MK, Kay LE. A (H)C(CO)NH-TOCSY pulse scheme for sequential assignment of protonated methyl groups in otherwise deuterated 15N, 13C-labeled proteins. J Biomol NMR. 1996;8:351–356. doi: 10.1007/BF00410333. [DOI] [PubMed] [Google Scholar]
- Garrett DS, Powers R, Gronenborn AM, Clore GM. A common sense approach to peak picking in two-, three- and four-dimensional spectra using automatic computer analysis of contour diagrams. J Magn Reson. 1991;95:214–220. doi: 10.1016/j.jmr.2011.09.007. [DOI] [PubMed] [Google Scholar]
- Goto NK, Gardner KH, Mueller GA, Willis RC, Kay LE. A robust and cost-effective method for the production of Val, Leu, Ile (delta 1) methyl-protonated 15N-, 13C-, 2H-labeled proteins. J Biomol NMR. 1999;13:369–374. doi: 10.1023/a:1008393201236. [DOI] [PubMed] [Google Scholar]
- Gross JD, Gelev VM, Wagner G. A sensitive and robust method for obtaining intermolecular NOEs between side chains in large protein complexes. J Biomol NMR. 2003;25:235–242. doi: 10.1023/a:1022890112109. [DOI] [PubMed] [Google Scholar]
- Guntert P, Mumenthaler C, Wüthrich K. Torsion angle dynamics for NMR structure calculation with the new program DY-ANA. J Mol Biol. 1997;273:283–298. doi: 10.1006/jmbi.1997.1284. [DOI] [PubMed] [Google Scholar]
- Hajduk PJ, Meadows RP, Fesik SW. NMR-based screening in drug discovery. Q Rev Biophys. 1999;32:211–240. doi: 10.1017/s0033583500003528. [DOI] [PubMed] [Google Scholar]
- Hilty C, Fernandez C, Wider G, Wüthrich K. Side chain NMR assignments in the membrane protein OmpX reconstituted in DHPC micelles. J Biomol NMR. 2002;23:289–301. doi: 10.1023/a:1020218419190. [DOI] [PubMed] [Google Scholar]
- Ikura I, Bax A. Isotope filtered 2D NMR of a protein-peptide complex: study of a skeletal muscle myosin light chain kinase fragment bound to calmodulin. J Am Chem Soc. 1992;114:2433–2440. [Google Scholar]
- Johnson BA, Blevins RA. NMRView: a computer program for visualization and analysis of NMR data. J Biomol NMR. 1994;4:603–614. doi: 10.1007/BF00404272. [DOI] [PubMed] [Google Scholar]
- Joo WS, Jeffrey PD, Cantor SB, Finnin MS, Livingston DM, Pavletich NP. Structure of the 53BP1 BRCT region bound to p53 and its comparison to the BRCA1-BRCT structure. Genes Dev. 2002;16:583–593. doi: 10.1101/gad.959202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kern D, Zuiderweg ER. The role of dynamics in allosteric regulation. Curr Opin Struct Biol. 2003;13:748–757. doi: 10.1016/j.sbi.2003.10.008. [DOI] [PubMed] [Google Scholar]
- Koch CA, Anderson D, Moran MF, Ellis C, Pawson T. SH2 and SH3 domains: elements that control interactions of cytoplasmic signaling proteins. Science. 1991;252:668–674. doi: 10.1126/science.1708916. [DOI] [PubMed] [Google Scholar]
- Koonin EV, Altschul SF, Bork P. BRCA1 protein products. Functional motifs Nat Genet. 1996;13:266–268. doi: 10.1038/ng0796-266. [DOI] [PubMed] [Google Scholar]
- Koradi R, Billeter M, Wüthrich K. MOLMOL: a program for display and analysis of macromolecular structures. J Mol Graph. 1996;14:51–55. doi: 10.1016/0263-7855(96)00009-4. [DOI] [PubMed] [Google Scholar]
- Krishnan VV, Thornton KH, Thelen MP, Cosman M. Solution structure and backbone dynamics of the human DNA ligase III alpha BRCT domain. Biochemistry. 2001;40:13158–13166. doi: 10.1021/bi010979g. [DOI] [PubMed] [Google Scholar]
- Lee W, Revington MJ, Arrowsmith CH, Kay LE. A pulsed field gradient isotope-filtered 3D 13C HMQC-NOESY experiment for extracting intermolecular NOE contacts in molecular complexes. FEBS Lett. 1994;350:87–90. doi: 10.1016/0014-5793(94)00740-3. [DOI] [PubMed] [Google Scholar]
- Lu KP. Pinning down cell signalling, cancer and Alzheimer’s disease. Trends Biochem Sci. 2004;29:200–209. doi: 10.1016/j.tibs.2004.02.002. [DOI] [PubMed] [Google Scholar]
- Macura S, Bremer J, Brown LR. Heteronuclear correlation 2D NMR spectroscopy of macromolecules. J Magn Reson. 1985;63:484–493. [Google Scholar]
- Manke IA, Lowery DM, Nguyen A, Yaffe MB. BRCT repeats as phosphopeptide-binding modules involved in protein targeting. Science. 2003;302:636–639. doi: 10.1126/science.1088877. [DOI] [PubMed] [Google Scholar]
- Menichini P, Linial M. SUVi and BACH1: a new subfamily of mammalian helicases? Mutat Res. 2001;487:67–71. doi: 10.1016/s0921-8777(01)00104-5. [DOI] [PubMed] [Google Scholar]
- Miki Y, Swensen J, Shattuck-Eidens D, Futreal PA, Harshman K, Tavtigian S, Liu Q, Cochran C, Bennett LM, Ding W, et al. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1. Science. 1994;266:66–71. doi: 10.1126/science.7545954. [DOI] [PubMed] [Google Scholar]
- Pervushin K, Riek R, Wider G, Wüthrich K. Attenuated T2 relaxation by mutual cancellation of dipole-dipole coupling and chemical shift anisotropy indicates an avenue to NMR structures of very large biological macromolecules in solution. Proc Natl Acad Sci USA. 1997;94:12366–12371. doi: 10.1073/pnas.94.23.12366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reimer U, Fischer G. Local structural changes caused by peptidyl-prolyl cis/trans isomerization in the native state of proteins. Biophys Chem. 2002;96:203–212. doi: 10.1016/s0301-4622(02)00013-3. [DOI] [PubMed] [Google Scholar]
- Rodriguez M, Yu X, Chen J, Songyang Z. Phospho-peptide binding specificities of BRCA1 COOH-terminal (BRCT) domains. J Biol Chem. 2003;278:52914–52918. doi: 10.1074/jbc.C300407200. [DOI] [PubMed] [Google Scholar]
- Rogakou EP, Boon C, Redon C, Bonner WM. Mega-base chromatin domains involved in DNA double-strand breaks in vivo. J Cell Biol. 1999;146:905–916. doi: 10.1083/jcb.146.5.905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiozaki EN, Gu L, Yan N, Shi Y. Structure of the BRCT repeats of BRCA1 bound to a BACH1 phosphopeptide: implications for signaling. Mol Cell. 2004;14:405–412. doi: 10.1016/s1097-2765(04)00238-2. [DOI] [PubMed] [Google Scholar]
- Staub O, Rotin D. WW domains. Structure. 1996;4:495–499. doi: 10.1016/s0969-2126(96)00054-8. [DOI] [PubMed] [Google Scholar]
- Vadrevu R, Falzone CJ, Matthews CR. Partial NMR assignments and secondary structure mapping of the isolated alpha subunit of Escherichia coli tryptophan synthase, a 29-kD TIM barrel protein. Protein Sci. 2003;12:185–191. doi: 10.1110/ps.0221103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waugh DS. Genetic tools for selective labeling of proteins with alpha-15N-amino acids. J Biomol NMR. 1996;8:184–192. doi: 10.1007/BF00211164. [DOI] [PubMed] [Google Scholar]
- Williams RS, Glover JN. Structural consequences of a cancer-causing BRCA1-BRCT missense mutation. J Biol Chem. 2003;278:2630–2635. doi: 10.1074/jbc.M210019200. [DOI] [PubMed] [Google Scholar]
- Williams RS, Green R, Glover JN. Crystal structure of the BRCT repeat region from the breast cancer-associated protein BRCA1. Nat Struct Biol. 2001;8:838–842. doi: 10.1038/nsb1001-838. [DOI] [PubMed] [Google Scholar]
- Williams RS, Lee MS, Hau DD, Glover JN. Structural basis of phosphopeptide recognition by the BRCT domain of BRCA1. Nat Struct Mol Biol. 2004;11:519–525. doi: 10.1038/nsmb776. [DOI] [PubMed] [Google Scholar]
- Wishart DS, Sykes BD. The 13C chemical shift index. A simple method for the identification of protein secondary structure using 13C chemical shift data. J Biomol NMR. 1994;4:171–180. doi: 10.1007/BF00175245. [DOI] [PubMed] [Google Scholar]
- Xu B, Kim S, Kastan MB. Involvement of BRCA1 in S-phase and G(2)-phase checkpoints after ionizing irradiation. Mol Cell Biol. 2001;21:3445–3450. doi: 10.1128/MCB.21.10.3445-3450.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yarden RI, Pardo-Reoyo S, Sgagias M, Cowan KH, Brody LC. BRCA1 regulates the G2/M checkpoint by activating Chk1 kinase upon DNA damage. Nat Genet. 2002;30:285–289. doi: 10.1038/ng837. [DOI] [PubMed] [Google Scholar]
- Yu X, Chini CC, He M, Mer G, Chen J. The BRCT domain is a phospho-protein binding domain. Science . 2003;302:639–642. doi: 10.1126/science.1088753. [DOI] [PubMed] [Google Scholar]
- Zhang X, Morera S, Bates PA, Whitehead PC, Coffer AI, Hainbucher K, Nash RA, Sternberg MJ, Lindahl T, Freemont PS. Structure of XRCC1 BRCT domain: a new protein-protein interaction module. EMBO J. 1998;17:6404–6411. doi: 10.1093/emboj/17.21.6404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zwahlen C, Legault P, Vincent SJF, Greenblat J, Konrat R, Kay LE. Methods for measurements of intermolecular NOEs by multinuclear NMR spectroscopy: application to a bacteriophage N-peptide/box B RNA complex. J Am Chem Soc. 1997;119:6711–6721. [Google Scholar]