The Bacillus subtilis endospore: assembly and functions of the multilayered coat (original) (raw)
. Author manuscript; available in PMC: 2023 Feb 9.
Published in final edited form as: Nat Rev Microbiol. 2012 Dec 3;11(1):33–44. doi: 10.1038/nrmicro2921
Abstract
Sporulation in Bacillus subtilis involves an asymmetric cell division followed by differentiation into two cell types, the endospore and the mother cell. The endospore coat is a multilayered shell that protects the bacterial genome during stress conditions and is composed of dozens of proteins. Recently, fluorescence microscopy coupled with high-resolution image analysis has been applied to the dynamic process of coat assembly and has shown that the coat is organized into at least four distinct layers. In this Review, we provide a brief summary of B. subtilis sporulation, describe the function of the spore surface layers and discuss the recent progress that has improved our understanding of the structure of the endospore coat and the mechanisms of coat assembly.
Bacteria have many strategies for surviving environmental challenges. These strategies frequently involve rapid changes in gene expression that temporarily alter the phenotype of a cell and allow it to survive. A more sophisticated and prolonged example of stress response is spore formation, or sporulation, wherein the bacterial genome is sequestered in a safe place (the spore) until environmental conditions improve, upon which the spore quickly germinates and returns to the vegetative state. Endospores are formed and nurtured completely within a mother cell, which must lyse to release the spore into the environment. Unrelated mechanisms of bacterial sporulation1 include the formation of exospores, myxospores and akinetes (BOX 1).
Box 1 |. Resting cells: endospores, exospores, myxospores and akinetes.
A variety of metabolically dormant bacterial cells, called resting cells, have been found in nature1. Endospores form inside a mother cell after the separation of the sporangium into distinct forespore and mother cell compartments; however, endospore formation is not limited to the production of a single spore per mother cell. Metabacterium polyspora, which is found in the gastrointestinal microbiota of guinea pigs, forms endospores at both poles of the mother cell_. M. polyspora_ endospores also appear to have the capacity to divide further by binary fission, enabling the production of more than three spores per cell108. Genetic manipulation of Bacillus subtilis can also result in the formation of two viable endospores in a single mother cell109.
Various other resting cell types that form ‘outside’ the original vegetative cell have also been described. _Streptomyces_species sporulate by forming a multinucleate sporogenic cell at the leading tip of an aerial hypha filament. Divisomes are assembled along the length of the sporogenic cell to partition each chromosome into an individual compartment that will eventually become an exospore110. During myxospore formation in Myxococcus species and akinete formation in heterocyst-forming species of Cyanobacteria, an entire individual cell transforms its morphology to form a resting cell in the absence of division1. To our knowledge, there has been no comprehensive parallel study of the relative resistance properties of the known types of resting cells; however, meta-analysis suggests that all resting cell types appear to be resistant to desiccation, whereas resistance to other types of stress such as extreme heat and predation by protozoa is variable111. The spore coat of myxospores has been investigated to some extent112, but it appears to be unrelated to the spore coat of endospores, both in terms of composition (it is made essentially of exopolysaccharides) and mechanism of assembly, which requires a dedicated protein machinery.
Endospores (hereafter referred to as spores) exhibit extraordinary resistance properties and have served as textbook examples of long-term cell survival2–4. The exact mechanism of spore persistence is unknown, but in addition to being resistant to ultraviolet (UV) radiation, chemicals (such as peroxide and hypochlorite), extreme heat and other stresses, spores are metabolically dormant and partially dehydrated, which is likely to allow their survival in nutrient-free and harsh environments. Although the upper limit of viability is unknown, some researchers have reported the revival of spores from samples ranging in age from decades to several thousands of years5, a claim that is difficult to prove unambiguously. Nevertheless, viable spores have been isolated from specimens such as dried plant samples dating from 1640 onwards6 and a flask of Bacillus anthracis spores sealed in Louis Pasteur’s laboratory for 60 years7
Two taxa within the phylum Firmicutes — the aerobic Bacillaceae and the anaerobic Clostridia — form spores via an evolutionarily conserved mechanism8,9 (see below). These bacteria include the pathogens_B. anthracis_ and Clostridium difficile, and the model organism Bacillus subtilis. Sporulation is a simple example of differentiation: a cell replicates its DNA, divides asymmetrically and places copies of its genome in both compartments. Over the next 8 to 10 hours, the smaller of the two compartments, the forespore (or prespore), develops into a mature spore capable of protecting the genome. When the spore is released into the surrounding environment it can persist or germinate to re-enter the vegetative cycle. A variety of spore-forming species are routinely found in soil samples10, but they have also been isolated from ecosystems as diverse as hot springs11, arctic sediments12 and the mammalian gastrointestinal tract13.
Our focus here will be the spore coat, a structure present on the exterior of all spores that is crucial for conferring resistance to environmental stress. The paradigm for coat architecture is provided by B_. subtilis_, which uses at least 70 different proteins to build this multilayered structure. In this Review, we provide a brief overview of sporulation in B. subtilis with a focus on the assembly and function of the coat. We discuss recent investigations combining genetic, biochemical and cytological approaches that have led to a better understanding of coat morphogenesis. Knowledge about the molecular mechanisms behind the formation of this complex structure should reveal novel principles underlying the assembly of multiprotein structures in various organisms.
Sporulation in Bacillus subtilis
In laboratory cultures, sporulation begins at the onset of stationary phase when nutrients are depleted; however, the ultimate ‘molecular signal’ of sporulation, if it exists, remains elusive. Sporulation is triggered by the activation of histidine sensor kinases (including KinA, KinB and KinC), which shuttle phosphate through an extended phosphorelay, resulting in phosphorylation of the master regulator of sporulation, the transcription factor Spo0A. Simply upregulating the expression of KinA appears to be sufficient to trigger sporulation, regardless of nutrient status14. Phosphorylated Spo0A controls a large regulon of genes, including those involved in asymmetric cell division and those involved in activation of the sporulation-specific sigma factors15. An asymmetric division of the sporulating cell creates a sporangium composed of two compartments, the larger mother cell and the smaller forespore, which ultimately becomes the spore (FIG. 1). The next stage is the engulfment of the forespore by the mother cell in a process resembling phagocytosis. After the completion of engulfment, the forespore is a double membrane-bound cell within the mother cell. Engulfment is coordinated with the building of two external protective structures: the cortex is composed of peptidoglycan16 and is assembled between the inner and outer forespore membranes, and the proteinaceous coat comprises the outermost layer of the spore17,18. Spore peptidoglycan precursors are synthesized in the mother cell19 and are flipped across the outer forespore membrane into the intermembrane space by an as yet incompletely understood mechanism20. The coat is composed of at least 70 individual proteins that are produced by the mother cell and begin to localize to the spore surface during engulfment21.
Figure 1 |. The sporulation and germination cycle in Bacillus subtilis.
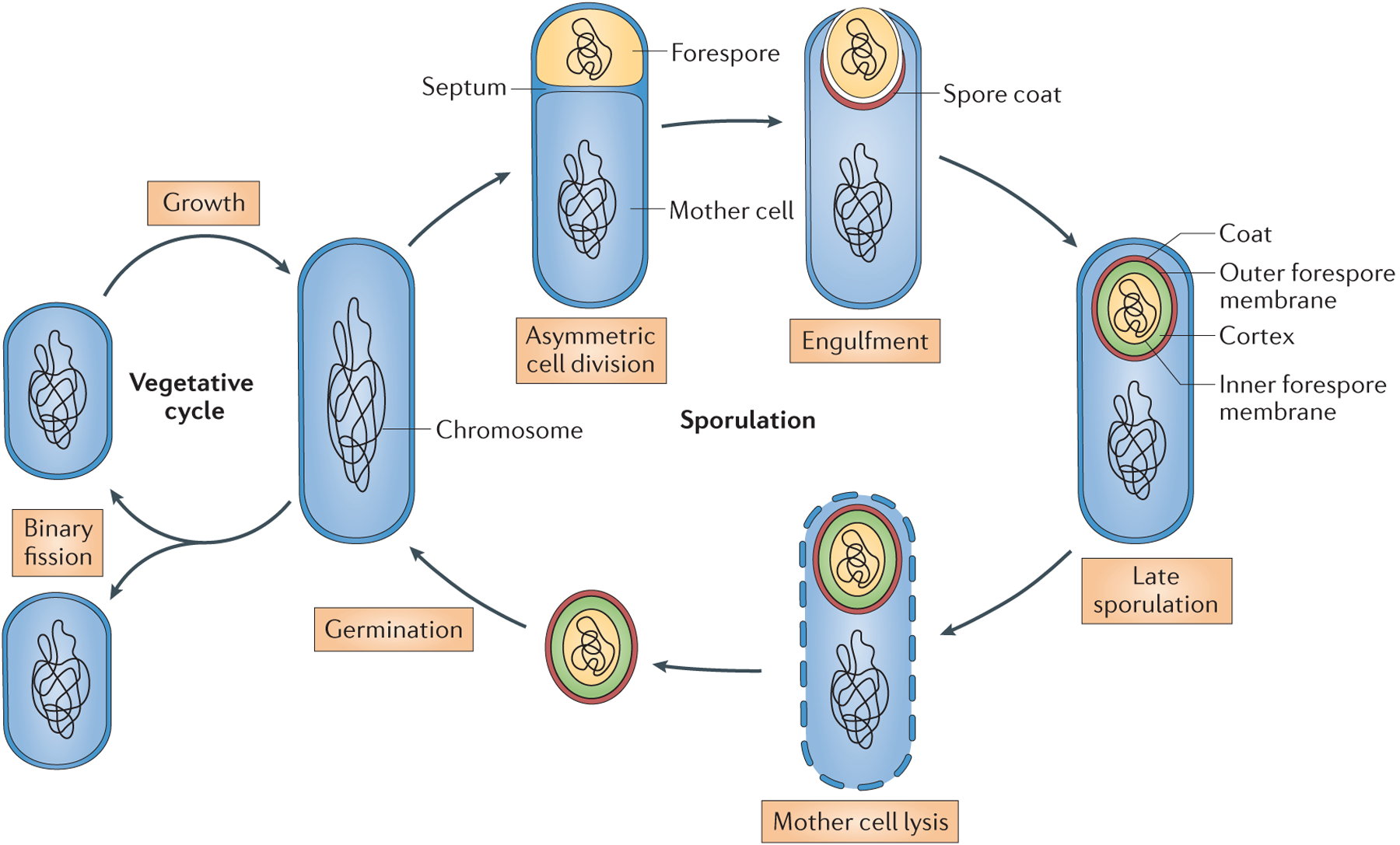
Sporulation begins when a sporangium divides asymmetrically to produce two compartments: the mother cell and the forespore, which are separated by a septum. Next, the mother cell engulfs the forespore, and following membrane fission at the opposite pole of the sporangium, a double-membrane bound forespore is formed. Coat assembly begins just after the initiation of engulfment and continues throughout sporulation. The peptidoglycan cortex between the inner and outer forespore membranes is assembled during late sporulation. In the final step, the mother cell lyses to release a mature spore into the environment. Spores are capable of quickly germinating and resuming vegetative growth in response to nutrients.
Structure of the spore
Since the initial application of electron microscopy to sporulating bacteria, researchers have been captivated by the concentrically arranged layers making up the spore external structure22,23 (FIG. 2). The genome is contained in the partially dehydrated central core, in which most of the water has been replaced with Ca2+-dipicolinic acid. The core is surrounded, from innermost to outermost, by the inner spore membrane, germ cell wall (a thin layer of vegetative cell wall-like peptidoglycan that is not depicted in the figures), cortex, outer spore membrane and coat. Most of these structures are rapidly lost upon germination: the coat is shed and the cortex is degraded by cell wall hydrolases. The germ cell wall is retained to serve as a primer for vegetative cell wall assembly. The inner spore membrane becomes the plasma membrane, whereas the fate of the outer spore membrane is less clear. It may be lost during spore maturation, but there is at least one organism (Acetonema longum) in which it persists after germination, thus generating a double membrane vegetative cell akin to Gram-negative bacteria24.
Figure 2 |. Spore ultrastructure.
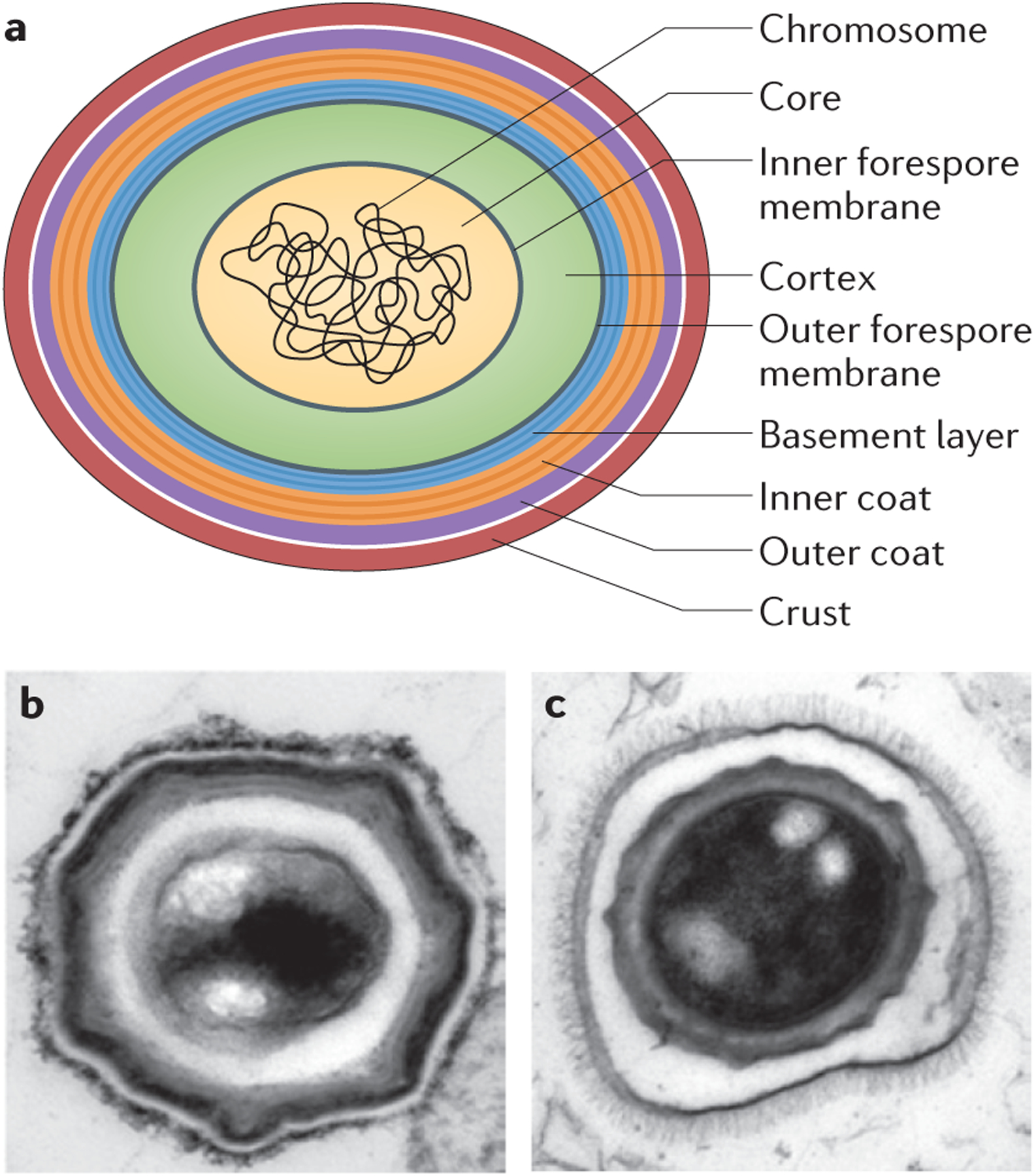
a | Cartoon of a typical Bacillus subtilis_spore. The multiple layers of the spore serve to protect the genome, which is housed in the partially dehydrated central core. The inner forespore membrane (grey) is visible in electron micrographs of sectioned spores, but the outer forespore membrane is not usually visible. The core is protected by the cortex (green) and the spore coat, which consists of four layers: the basement layer (blue), inner coat (orange), outer coat (purple) and crust (red), which is only visible after ruthenium red staining. The concentric rings of the basement layer and inner coat reflect the lamellar appearance of the inner coat in electron micrographs. It is unknown how these lamellae form or whether they are composed of specific proteins. b | Thin section transmission electron micrograph of a B. subtilis spore stained with ruthenium red.c | Thin section transmission electron micrograph of a_Bacillus anthracis spore stained with osmium tetroxide. The exosporium is the outermost protective layer of the spore and it contains hair-like projections. It is separated from the rest of the spore by a large interspace. Part b is reproduced, with permission, from REF. 25 © (2010) Elsevier.
Three layers of the B. subtilis spore coat are observed in thin-section electron microscopy: a lamellar inner coat, a more coarsely layered outer coat and a recently identified layer named the crust23,25(FIG. 2a,b). In some species, such as B. anthracis, a different type of outermost layer exists, called the exosporium22. The exosporium is a distinct glycoprotein layer that surrounds the spore and is separated by a gap called the interspace26–29 (FIG. 2c). In Bacillus cereus and its close relatives_B. anthracis_ and Bacillus thuringensis, the exosporium consists of a basal layer adorned with hair-like projections. Recent structural studies have documented the crystalline two-dimensional architecture of the basal layer30. In isolation, or in combination with the exosporium, the coat fulfils several functions other than protection of the genome (see below).
Between species, the spore surface differs most obviously by the presence or absence of an exosporium, but there is also considerable diversity in the individual coat proteins. Analysis of existing genome sequences suggests that about half of the known B. subtilis coat protein genes have recognizable orthologues in other Bacillus species, whereas the other half do not appear to be conserved at all. In the genomes of Clostridium spp., conservation of B. subtilis coat genes is even more limited8,9,17,31. Considering that the spore surface represents the interface between spores and the environment, diversity in coat composition may have a crucial role in defining the ecological niches of spore-forming bacteria (see below).
Functions of the spore coat
Protection from environmental stress.
The resistance of spores to extreme environmental stresses is likely to be an emergent property of spore architecture, making it difficult to describe causal connections between individual stresses and resistance mechanisms. For example, resistance to extreme heat requires partial dehydration of the spore core, a functional cortex, RecA-dependent DNA repair machinery and an array of small acid-soluble proteins, which bind to the genome in the spore core32. Similarly, spores may be resistant to desiccation because the spore core itself is partially dehydrated. Late in sporulation, most of the water in the spore core is replaced with Ca2+-dipicolinic acid, and mutants deficient in Ca2+-dipicolinic acid production do not survive desiccation.
Spores of B. subtilis are also more resistant to UV irradiation and peroxide than vegetative cells32. CotA, a copper-dependent laccase found in the outer coat, contributes to protection against UV radiation and peroxide by generating a pigment that is structurally similar to melanin33. Melanin confers resistance to reactive oxygen species in a number of different microorganisms34, and deactivation of reactive oxygen species may be a common mediator of resistance to both UV light and peroxide. Melanin has also been shown to interfere with phagocytosis of fungal pathogens such as Cryptococcus neoformans by human macrophages, and it is necessary for yeast survival within macrophages35. It is currently unknown whether CotA has a similar effect on the phagocytosis and survival of spores.
The major known function of the coat is spore protection and, indirectly, preservation of the spore genome. Remarkably, B. subtilis spores can survive ingestion by bacteriovores as different as the protozoan Tetrahymena thermophila and the nematode_Caenorhabditis elegans_36–38. After being fed wild-type spores of B. subtilis, electron microscopy revealed that T. thermophila phagosomes were full of intact, undigested spores; however, mutations that disrupt the coat result in rapid digestion of the spore cortex and core. Thus, in the natural environment, the coat is likely to provide physical protection from predators. Although the exact molecular mechanism of this protection is unknown, the genomes of both_T. thermophila_ and C. elegans contain multiple homologues of putative peptidoglycan hydrolases, suggesting that the coat may have an important role in limiting access of these enzymes to the peptidoglycan-containing cortex.
Regulation of germination.
For dormancy to be a viable survival strategy, spores must be able to germinate rapidly when nutrients become available again. Spores continually monitor their surroundings using an array of germination receptors embedded in the inner spore membrane. Germinants of B. subtilis include sugars, amino acids, peptidoglycan fragments and ions39,40. As germinants will reach their receptors by traversing the spore coat and cortex, the coat must ultimately be both protective and permeable. The coat can be conceptualized as a molecular sieve that excludes large molecules, such as lysozyme, while allowing the passage of small-molecule germinants22,41.
Although the effect of the coat on the initiation of germination may be strictly passive, the coat and the exosporium also have the potential to enzymatically affect germination. An intrinsic property of spore surfaces is the ability to convert l-alanine to d-alanine using an alanine racemase42. Becausel-alanine is a potent germinant, conversion of l-alanine to d-alanine by the coat could suppress germination. Recently, an alanine racemase enzyme (Alr) from the exosporium of B. anthracis spores was characterized43,44. During sporulation of B. anthracis cells lacking Alr, forespores germinate prematurely within the mother cell, suggesting that Alr insulates the developing forespore from free l-alanine in the mother cell cytoplasm45. An orthologue of alr in B. subtilis(yncD) encodes an alanine racemase that localizes to the outer coat layer25,46; however, deletion of_yncD_ does not result in premature forespore germination47.
The coat also influences germination by sequestering enzymes required for degradation of the cortex peptidoglycan. In particular, the B. subtilis inner coat contains at least two peptidoglycan hydrolases, CwlJ and YaaH (SleL)21,48–51. Furthermore, peptidoglycan-binding domains, such as the LysM domain52, are present in several coat proteins (including YaaH), but their exact functions have not yet been determined in the context of the sporulation–germination cycle. It is possible that the impact of the coat on peptidoglycan hydrolysis is more complex than currently suspected. The functional interdependency between the coat and cortex layers is emphasized further by the recent discovery of CmpA, a mother cell-expressed protein that appears to be involved in a checkpoint ensuring that cortex synthesis is delayed until coat assembly successfully initiates53.
After the initiation of germination, the coat must be opened and ultimately shed to allow the vegetative cell to grow out54. In B. subtilis, it is unclear whether outgrowth is a polarized process, wherein one pole of the spore is favoured over another. In B. anthracis, outgrowth appears to be biased towards the mother cell proximal (MCP) pole, which is the initial localization site of the spore coat and exosporium proteins55.
Defining an ecological niche?
For all bacteria, the properties of the cell surface define the range of interactions that the bacterium may establish with other organisms and surfaces. The diversity of spore surface structures (see, for example, REFS 22,27,56–58) and the conservation of only a subset of coat proteins8,9,17, which includes the coat morphogenetic proteins17, suggests that spore surface properties are diverse and may contribute to the wide dispersal of spore-forming bacteria. The available data suggest that, between species, spore surfaces can display varying degrees of hydrophobic or hydrophilic characteristics29,59,60.
If the function of the coat is to resist ubiquitous environmental stresses, then why is coat structure so variable? In contrast to the coat, the core and cortex do not usually display interspecies differences that are distinguishable by electron microscopy. Given the diversity in coat structure among species, it seems reasonable to speculate that this diversity is driven by adaptation to a variety of niches. However, as any specific niche harbouring bacterial spores typically possesses diverse species (see, for example, REF.61), addressing this question is extremely complex62. The functions of the coat and the exosporium might have evolved in an exquisitely fine-tuned manner to allow spores to thrive in diverse habitats, both natural and man-made. For example, to cause disease in a hospital setting, a C. difficile spore needs to interact with abiotic surfaces, such as stainless steel, as well as the varied microenvironments within a host. During their transit between niches, it is likely that the level and type of stresses vary considerably, and the surfaces to which spores need to adhere represent another important source of variation. Thus, spore transmission in complex environments, where the ability to adhere to and survive on variable surface structures is crucial, could be a key factor that promotes diversity in coat structure and composition. Moreover, the ability of a spore to attach to or avoid specific surfaces could facilitate future growth after germination. Elucidation of the physical and chemical properties of spores, in conjunction with their variability in different ecological conditions, is an important goal for future research and should help to identify specific environmental features driving coat variation.
Spore coat assembly
The first coat proteins were identified by reverse genetic approaches. Specifically, extracts of coat material were subjected to peptide sequencing, the resulting sequences were cloned and mutants were constructed to determine the role of each protein in coat assembly, which was examined by electron microscopy63–65 (TABLE 1). Although electron microscopy offers unparalleled resolution of subcellular structures, the lack of systems for high-sensitivity labelling of individual proteins has limited its application. More recently, proteomics approaches and global transcriptional profiling of the mother cell compartment led to the discovery of several novel spore proteins66–70. Candidate coat proteins can then be verified by fluorescence microscopy, which involves examining the localization of green fluorescent protein (GFP)-tagged candidate coat proteins in mutants with disrupted coat assembly71,72. Moreover, the combination of a GFP fusion protein with a fluorescent membrane stain allows the kinetics of coat protein localization relative to the membrane dynamics of engulfment to be observed21.
Table 1 |.
Table of representative coat proteins from Bacillus subtilis
| Coat protein | EM phenotype of null mutant | Dependent coat proteins | Domains* | Conservation‡ | Refs |
|---|---|---|---|---|---|
| Basement layer proteins (SpoIVA-dependent) | |||||
| SpoIVA | No coat attached, no cortex | All tested | Walker A ATPase | All endospore formers | 73,75,86,93 |
| SpoVM | No cortex, encasement blocked | All tested | Amphipathic α-helix | All members of the Bacillaceae and most_Clostridium_ spp. | 74,82,83,90,118 |
| SpoVID | Encasement blocked | Most tested proteins | LysM | Most members of the Bacillaceae and no Clostridium spp. | 64,78,83,91,92 |
| LipC (also known as YcsK) | ND | ND | Lipase | All members of the Bacillaceae and most_Clostridium_ spp. | 122 |
| YhaX | ND | ND | HAD (haloacid dehydrogenase) family | All members of the Bacillaceae and most_Clostridium_ spp. | 25,72 |
| YheD | ND | ND | – | Some members of the Bacillaceae and no_Clostridium_ spp. | 88 |
| Inner coat proteins (SafA-dependent) | |||||
| SafA | No inner coat | 17 | LysM | Most members of the Bacillaceae and no_Clostridium_ spp. | 76,77,85 |
| CotD | ND | ND | – | Most members of the Bacillaceae and no_Clostridium_ spp. | 25,51,63 |
| CotT | Thinner inner coat | ND | – | Only found in Bacillus subtilis | 25,51,123 |
| CwlJ | ND | ND | Peptidoglycan hydrolase | All endospore formers | 21,48,49 |
| OxdD (also known as YoaN) | ND | ND | Oxalate decarboxylase | Most members of the Bacillaceae and some_Clostridium_ spp. | 124 |
| YaaH (also known as SleL) | ND | ND | LysM (x2)§,_N_-acetyl-glucosaminidase | All members of the Bacillaceae and most_Clostridium_ spp. | 25,48,50,51 |
| YuzC | ND | ND | – | Some members of the Bacillaceae and no_Clostridium_ spp. | 25,72 |
| Outer coat proteins (CotE-dependent) | |||||
| CotE | No outer coat | 16 | – | All members of the Bacillaceae and no_Clostridium_ spp. | 25,65,71,78,94 |
| CotA | No phenotype | ND | Multi-copper oxidase | Most members of the Bacillaceae and some_Clostridium_ spp. | 25,33,51,63 |
| CotM | Modest outer coat defect | ND | α-crystallin family of stress proteins | Most members of the Bacillaceae and some_Clostridium_ spp. | 25,125 |
| CotO (also known as YjbX) | Outer coat defect | ND | – | Most members of the Bacillaceae and no_Clostridium_ spp. | 25,72,102 |
| CotS | No phenotype | ND | Bacterial spore kinase | Most members of the Bacillaceae and some_Clostridium_ spp. | 25,69,126,127 |
| YncD | ND | ND | Alanine racemase | All endospore formers | 25,46 |
| YtxO | ND | ND | – | Most members of the Bacillaceae and no_Clostridium_ spp. | 25,51,69 |
| Crust proteins (CotX/Y/Z-dependent) | |||||
| CotX/CotY/CotZ∥ | No crust attached | 1 | – | Most members of the Bacillaceae and no_Clostridium_ spp. | 25,79,97 |
| CotW | ND | ND | Coiled coil | Some members of the Bacillaceae and no_Clostridium_ spp. | 25,97 |
Coat morphogenetic proteins.
The first description of spore coat structure came from thin-section electron microscopy23 (FIG. 2). Distinct inner and outer coat layers were made visible using osmium tetroxide staining: the outer coat was darkly stained, whereas the inner coat was stained more lightly and contained multiple concentric dark lamellae. Initial forward genetic screens aimed at identifying the genes required for sporulation in B. subtilis revealed two loci that have a major effect on coat structure:spoIVA73 and spoVM74. Electron microscopy of_spoIVA_ mutants showed that coat material was present, but instead of encircling the spore, the coat assembled in aggregates floating in the mother cell cytoplasm73,75. Interestingly, the darkly and lightly stained layers of the coat were still present, suggesting that even in the absence of spore-surface localization, some self-assembly was preserved. These data are consistent with a role for SpoIVA in anchoring the coat to the spore surface. By contrast, a transposon insertion in _spoVM_resulted in a spore coat that was localized to the spore surface but was only partially attached and significantly disorganized74.
Among the genes identified using reverse genetics approaches were_safA_, cotE and_spoVID_64,65,76,77. Electron microscopy of mutant strains revealed that_safA_ mutant spores lack an inner coat but retain a seemingly unaffected outer coat76, whereas cotE mutant spores retain an inner coat but lack an outer coat65. By contrast, the phenotype of _spoVID_mutant spores resembled that of the spoVM mutant spores64; in both cases the two coat layers could be distinguished, but the coat did not form a shell of protein around the forespore and was often found detached from the forespore surface. Subsequent work showed that spoVID, along with spoVM, is required for a later stage in coat assembly, spore encasement (see below). Collectively, the electron microscopy data suggested that assembly of the two layers occurs independently: SafA is necessary for assembly of the inner coat, whereas CotE is necessary for assembly of the outer coat. We now know that these two proteins, together with SpoIVA, form the core of a modular network responsible for assembly of a scaffold cap of the spore coat on the MCP pole of the forespore25 (FIG. 3). By contrast, SpoVM and SpoVID are crucial for the transition from a single cap to a full spherical shell that encases the spore. These key proteins (SpoIVA, SpoVM, SpoVID, SafA and CotE) were named morphogenetic proteins as they have a major role in coat morphogenesis but do not affect gene expression in the mother cell.
Figure 3 |. The coat genetic interaction network.
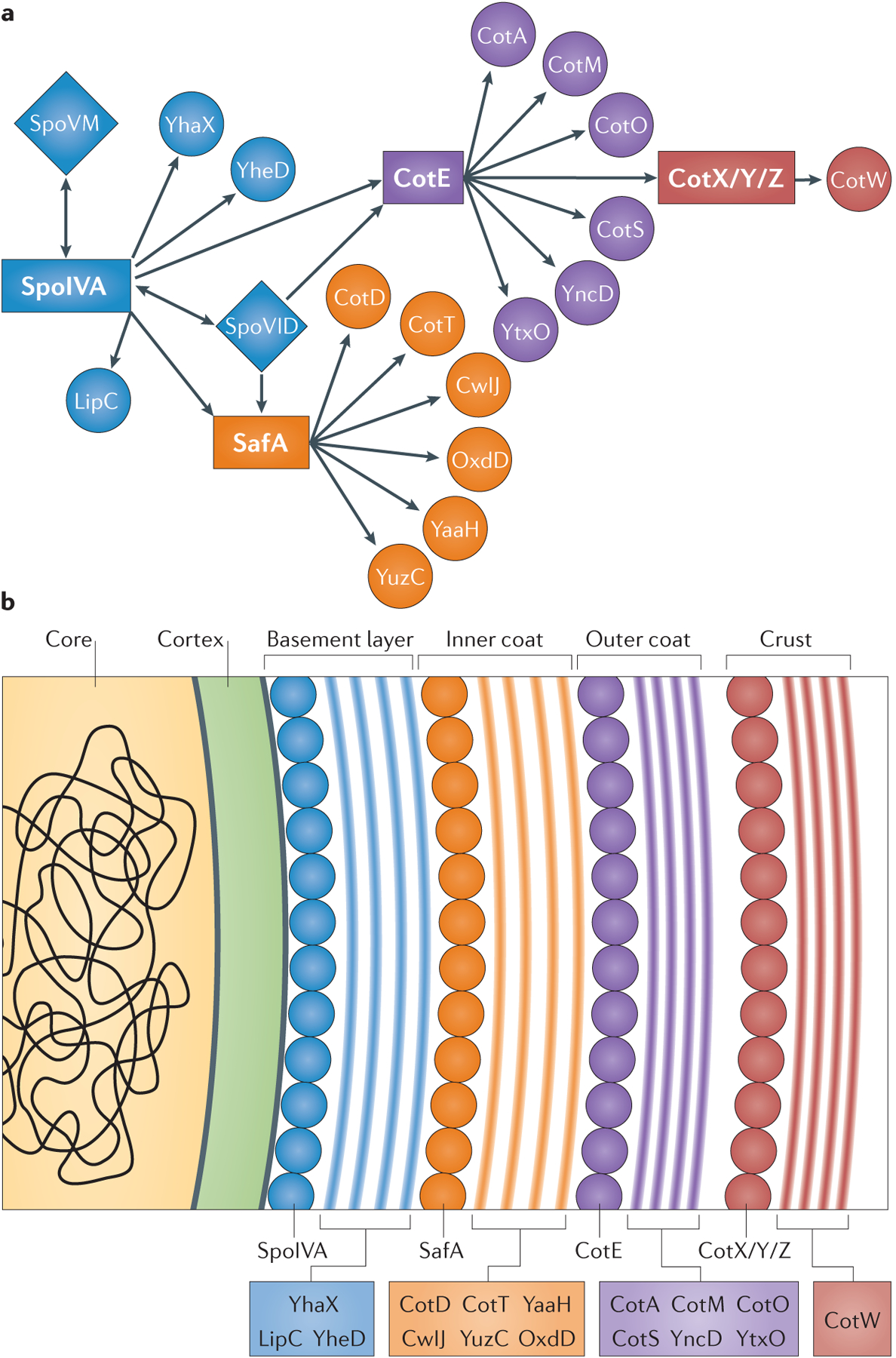
a | Recognition of the spatially scaled genetic interaction network led to the proposal of a coat made up of four layers (the basement layer (blue), inner coat (orange), outer coat (purple) and crust (red)), each with its own independent morphogenetic protein (SpoIVA, SafA, CotE and CotX/CotY/CotZ, respectively). The morphogenetic proteins SpoVM and SpoVID are required for spore encasement (FIG. 4). b | Assembly of each layer may be driven by the multimerization of the underlying morphogenetic protein. Polymerization of a morphogenetic protein would, presumably, create the necessary binding sites for each of the individual coat proteins that make up each layer. It remains to be determined whether and how the proteins of the individual layers interact with adjacent layers and how adjacent layers interact to form the mature spore coat.
The spore coat genetic interaction network.
The application of immuno-electron microscopy to B. subtilis spores revealed the subcellular localization of the morphogenetic proteins and provided initial clues as to how these proteins direct spore coat protein assembly: SpoIVA was detected in close proximity to the membrane, SafA in the inner coat77 and CotE at the interface of the inner and outer coats78. By using the localization data of these three proteins and the phenotypes of the corresponding mutants, it was possible to draw the skeleton of an interaction network (FIG. 3a). Our laboratories and others took a genetic approach to further characterize this network25,69,79. Spore coat proteins were defined by a failure to localize to the spore surface in the absence of SpoIVA, and catalogued as inner or outer coat proteins based on their inability to localize to the spore surface in mutants of safA_and cotE, respectively (TABLE 1). To date, 41 GFP-tagged spore coat proteins have been examined for genetic interactions with spoIVA, cotE and_safA25,69.
These investigations revealed a network of genetic interactions with three nearly independent modules: a SafA-dependent subnetwork, a CotE-dependent subnetwork and a third module consisting of proteins that are independent of both SafA and CotE, and primarily dependent on SpoIVA (FIG. 3a; TABLE 1). Spatial information was inferred from the genetic dependencies. For example, because SpoIVA is necessary to anchor the coat to the spore surface, the subnetwork around SpoIVA should localize closest to the spore membrane.
To test whether the genetic interaction network accurately predicted the localization of individual coat proteins, high-resolution image analysis was used to circumvent the theoretical limit of light microscopy, which overlaps with the maximal width of the spore coat (~200 nm). The distances between the spore membrane and 17 individual coat protein fusions were measured. When these data were then sorted by the mean distance between the spore membrane and each fusion protein, coat proteins grouped according to the interactions predicted by the genetic interaction network: CotE-independent proteins were closest to the membrane and CotE-dependent proteins were furthest from the membrane25. Unexpectedly, a group of proteins that are significantly separated from CotE was also identified, suggesting the possibility of an additional layer and perhaps a fourth genetic interaction module controlled by cotX, cotY and_cotZ_25. Notably, a homologue of CotY had been identified in extracts of the exosporium of B. anthracis80, but as mentioned above, B. subtilis_spores do not have an exosporium. Electron microscopy in combination with conventional staining indicated that the outer coat is the outermost layer in_B. subtilis. However, more recent analysis using staining with ruthenium red showed an additional electron-dense layer assembled over the outer coat81, which we have named the crust25. Because the contours of the outer coat and crust follow each other very closely, we propose that these layers are in direct contact, in spite of the electron-translucent region seen by transmission electron microscopy. The crust was absent in Δ_cotXYZ_ mutant spores, arguing that CotX, CotY and CotZ, individually or in combination, are morphogenetic proteins involved in crust assembly25,79.
Encasement: the second step in coat assembly.
The morphogenetic proteins discussed thus far have similar mutant phenotypes in that they appear to be involved in localizing spore coat proteins to the spore surface. Another distinct phenotypic class also exists: in these mutants, coat proteins localize to the spore surface, but the coat fails to form a full shell around the spore. Two examples are mutants of_spoVID_ and spoVM64,82,83 (TABLE 1). In these two mutants, most of the 41 GFP-tagged coat proteins localize to the MCP pole of the forespore but fail to transition to complete concentric rings of fluorescence78,83–87. This phenomenon was named ‘spore encasement’ to distinguish it from the localization of coat proteins to the spore surface, which is controlled by SpoIVA, SafA, CotE, CotX, CotY and CotZ.
Morphogenesis of the spore coat in real time
The genetic interaction network was assembled using data from mature spores; however, it has been clear from the first observations of the localization of individual coat proteins that coat morphogenesis is dynamic and coordinated with spore development78,88 (FIG. 4). For example, SpoIVA and SpoVM were observed tracking along with the engulfing forespore membrane78,86,89,90, whereas CotE appears to localize in two distinct steps: first forming a cap on the MCP pole of the spore and later forming a second cap on the mother cell distal (MCD) pole of the spore after the completion of engulfment71,83,89.
Figure 4 |. A model for coat morphogenesis: successive waves of spore encasement.
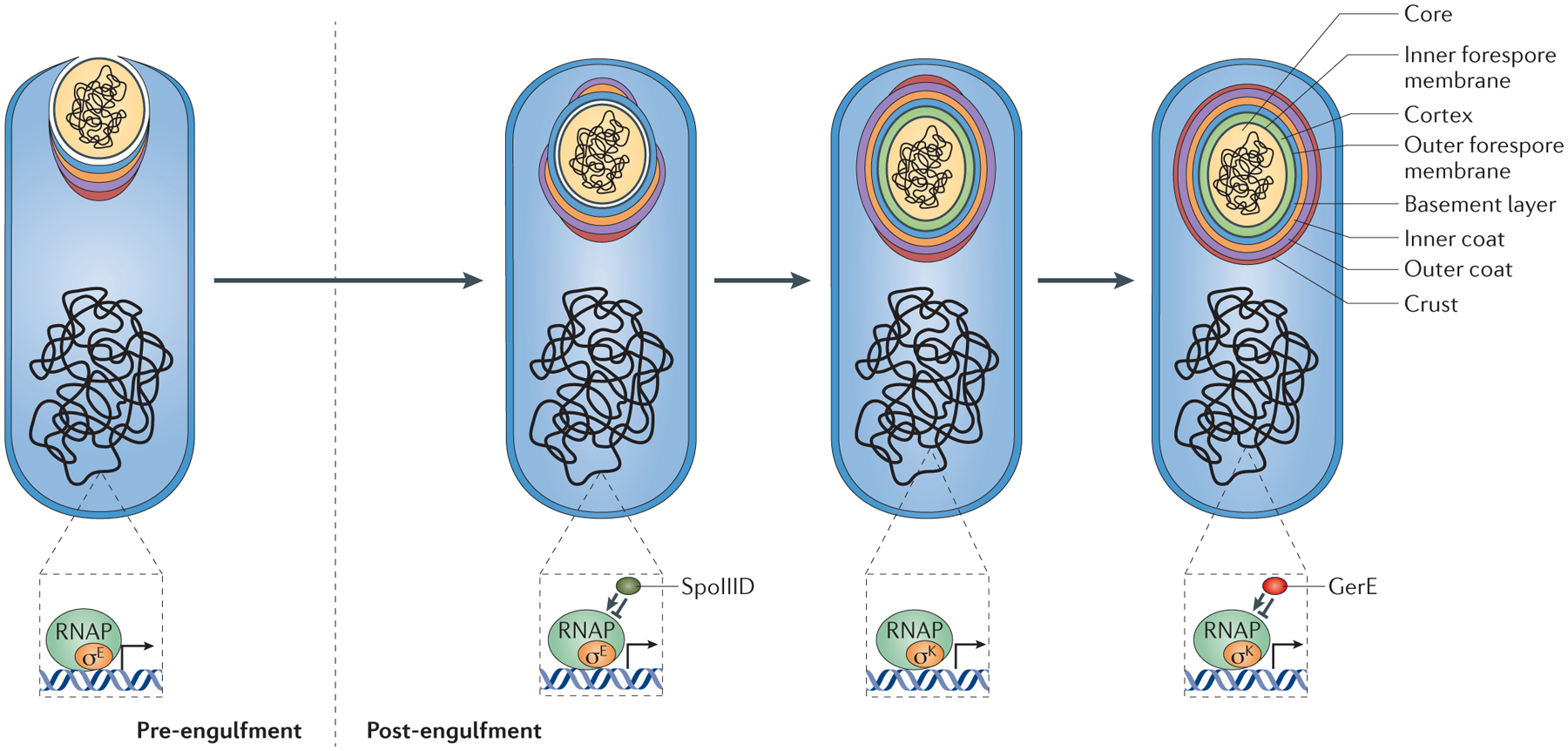
Coat morphogenesis begins with the assembly of a scaffold containing half of all coat proteins on the mother cell proximal (MCP) pole of the forespore. Assembly of the basement layer (blue), which consists of proteins that are solely under the control of the sigma factor σE for transcription, is dependent on SpoIVA. Encasement by SpoIVA is likely to be driven by multimerization and happens concomitantly with engulfment. Assembly of the inner (orange) and outer (purple) coat layers is dependent on SafA and CotE, respectively. Proteins of the inner and outer coat appear to have delayed encasement kinetics in comparison to the basement layer, and although they begin to assemble on the MCP pole during engulfment, they only form a cap on the mother cell distal (MCD) pole of the forespore after engulfment is complete. Most proteins in this class show dependency on combinations of σE and σK, as well as the transcription factor SpoIIID for expression. CotZ, a key component of the crust (red) belongs to the last encasement class and is dependent on σE, σK and the transcription factor GerE for expression. All four layers contain late-expressed σK-dependent proteins that are not part of the initial scaffold. These proteins presumably diffuse through the permeable coat matrix to reach their final location within the coat. Transcriptional regulation in the mother cell controls the kinetics of spore encasement, in particular the two sigma factors — σE(for the control of early gene expression during engulfment) and σK (for the control of late gene expression post-engulfment) — and two mother cell-specific transcription factors — SpoIIID (which is turned on by σE and modulates the σE regulon) and GerE (which is turned on by σK and modulates the σK regulon). RNAP, RNA polymerase.
Recently, the dynamics of spore coat protein localization over the course of sporulation were examined by fluorescence microscopy of the library of 41 GFP-tagged coat proteins21. The proteins could be divided into two roughly equal groups based on whether initial localization on the forespore surface occurred during or after engulfment. The proteins that localized during engulfment did so simultaneously to the forespore MCP pole and included the morphogenetic proteins SpoIVA, SpoVM and SpoVID. These three proteins tracked along the membrane, forming a full shell of fluorescence upon the completion of engulfment (FIG. 4). Thus, their presence in the earliest kinetic class links the genetically uncoupled phenomena of coat protein localization and encasement. Consistent with these observations, these proteins interact physically in biochemical assays83,87,91. Surprisingly, additional waves of encasement could be readily distinguished. Among the early-localizing coat proteins, another wave of encasement occurred only after completion of engulfment and the establishment of the MCD pole of the forespore, arguing that proteins in this class, including CotE, require the MCD pole for encasement (FIG. 4). Importantly, this later wave is dependent on SpoVM and SpoVID, as highlighted by the observation that a direct interaction between CotE and the amino-terminal domain of SpoVID is crucial for encasement of the outer coat92. The final wave of encasement among the early-localizing coat proteins, which includes the crust protein CotZ, occurs even later (FIG. 4). In summary, encasement by the morphogenetic proteins proceeds in a distinct order from basement layer to outer coat to crust. As such, the timing of encasement (which is subject to transcriptional control, see below) may represent the primary organizational phenomenon of coat morphogenesis21.
Encasement.
The molecular mechanism driving encasement is unknown; however, the sequential encasement of the spore by the morphogenetic proteins may offer some clues. For example, multimerization of all or some of the morphogenetic proteins is a plausible model of coat assembly (FIG. 3b). Purified SpoIVA polymerizes in an ATP-dependent manner and forms cables of varying diameter93, and the morphogenetic proteins SafA, CotE and CotZ have all been shown to self-interact biochemically84,94,95.
Because polymerization is limited by the availability of polymer subunits, one way to control the timing of encasement would be to limit the availability of coat protein monomers so that they are only expressed when they are needed. Gene expression in the mother cell is organized into a series of feedforward loops that result in successive waves of gene transcription and protein synthesis68,96. This hierarchical organization of gene expression is the result of transcriptional control of promoters by compartment-specific sigma factors and transcription factors (FIG. 4). The sigma factor σE is activated shortly after the completion of asymmetric division and regulates early gene expression in the mother cell. After the completion of engulfment, a second sigma factor, σK, replaces σE and regulates late gene expression in the mother cell. Among the >250 genes in the σE regulon is the gene encoding the transcription factor SpoIIID68,96. SpoIIID regulates subsets of the σE regulon, both positively and negatively. This results in three waves of gene expression: an initial wave that is σE-dependent and later repressed by SpoIIID, a second wave under the control of σE alone and unaffected by SpoIIID, and a third wave that is σE-dependent and further upregulated by SpoIIID68. Similarly, a late mother cell-specific transcription factor, GerE, positively and negatively regulates subsets of the σK regulon, also subdividing it into three waves68.
The genes encoding the coat morphogenetic proteins belong to four temporally distinct classes of promoter regulation. Although clusters of sporulation genes can be found in the genome, genes encoding coat proteins appear to be randomly distributed around the chromosome and are rarely organized into operons (one exception being the cotXYZ cluster of genes that is required for crust formation97,98). SpoIVA is expressed in a short pulse under the control of σE, and its synthesis is later repressed by SpoIIID68,75. SafA is upregulated by σE alone and is unaffected by SpoIIID76, whereas CotE is expressed in two pulses, first by σE and then by σK, before being repressed by GerE68,99. Expression of the crust protein operon, cotYZ, is activated by σE, σK and GerE68,98. The transcriptional data correspond well to the localization kinetics of these proteins (as discussed earlier) and support the notion that the major coat morphogenetic proteins are expressed in ordered waves. SpoIVA is a member of the first kinetic class of fusion proteins that finish encasement at the end of engulfment86. The localization kinetics of SafA are unknown; however, it is either a member of the first kinetic class (considering that it interacts directly with SpoVID85) or, along with CotE, it could belong to a second kinetic class. Finally, encasement by CotZ begins only after the spore becomes visible by phase contrast microscopy as a dark sphere in the mother cell cytoplasm. Although it is initially transcribed at a modest level under the control of σE, cotYZ is only transcribed at a high level during late sporulation.
Summary of coat assembly.
The large number of proteins that localize simultaneously to the MCP pole indicate that the coat may be spatially organized into layers from the beginning of the assembly process, as previously suggested78 (FIG. 4). Time-resolved measurements of individual protein fusions during engulfment confirmed that proteins are organized into four distinct layers early in morphogenesis and may form an organized scaffold on the MCP pole21. Encasement then occurs in successive waves, beginning with proteins of the basement layer, followed by the outer coat and finally the crust. This establishes each layer of the scaffold on both poles and around the circumference of the spore. Intuitively, the easiest way to continue coat assembly for the late-expressed coat proteins would be to build additional layers on top of the pre-existing scaffold. The presence of some inner coat proteins among the latest-expressed coat gene promoters suggests that the coat remains permeable late in morphogenesis, even to protein fusions as large as 70 kDa21. Thus, with all coat layers present at the beginning of coat morphogenesis, we expect that the major mechanism of addition of late-expressed coat proteins is by diffusion through the permeable coat. Crosslinking among proteins may limit this permeability. The one example of crosslinking among coat proteins that has been described in detail, the Tgl-mediated crosslinking of GerQ, appears to occur only after mother cell lysis and the release of spores into the environment100.
Outstanding questions and concluding remarks
Many important questions regarding the mechanism of spore coat assembly have not yet been addressed. In addition to CotE, other coat morphogenetic proteins such as CotO and CotH appear to be necessary for the recruitment of proteins to the outer coat101–103. This raises the possibility of hierarchical interactions among individual proteins within a single coat layer. It also remains to be determined whether the current genetic interaction network accurately predicts physical interactions among proteins. For example, does CotE exclusively interact with predicted outer coat proteins and not with inner coat proteins? Various physical interactions between coat proteins have been described77,83–85,87,91,92,95,104, but this has not yet been comprehensively investigated with, for example, a high-throughput two-hybrid technique. A systematic test of all possible physical interactions among coat proteins would also address the extent of hierarchical organization within coat layers.
Another potentially interesting question is how the thickness of each layer is determined. A possible clue is provided by a study that analysed B. subtilis spores with mutations in the spore coat protein genes_cotH_ and cotO102. Individually, mutation in either gene results in a disorganized outer coat. The double mutation has an intriguing additional phenotype: outer coat proteins form aggregates that bulge out significantly from the spore surface, resulting in a thicker outer coat. It is possible that proteins such as CotH and CotO act synergistically to enforce a specific width on the outer coat, analogous to the ‘molecular rulers’ of flagella and injectisomes105.
The coat morphogenetic proteins may be important targets for evolutionary adaptation across species, providing a pathway for rapid modification of coat architecture. Thus, an especially interesting area of future research will be to compare the coat protein assembly network of B. subtilis with networks in other species. B. anthracis possesses orthologues of several B. subtilis coat morphogenetic proteins, so it is tempting to speculate that B. anthracis uses some of the same proteins as_B. subtilis_ to direct assembly of its coat27. For example, SpoIVA is necessary to anchor the coat to the spore surface in both species. By contrast, deletion of_cotE_ in B. anthracis has only a modest effect on coat assembly; however, it is necessary for proper assembly of the exosporium.
The dynamics of spore encasement may also be largely conserved among spore-forming bacteria. In early electron micrographs, assembly of the exosporium in_B. anthracis_ was observed to progress from an initial cap on the MCP pole of the spore to a full shell106. These electron microscopy observations appear to agree with the localization dynamics of individual exosporium proteins, which form an initial cap on the MCP pole before transitioning to a complete shell of protein that surrounds the spore107. In cells deleted of exsY, a homologue of B. subtilis cotZ, the exosporium assembles as a cap on the MCP pole of the forespore but fails to encase the spore26. This pheno type is reminiscent of the spore encasement phenotypes of the _spoVM_and spoVID mutants of B. subtilis83.
Through a combination of genetic, biochemical and microscopy studies, much progress has been made in understanding how the spore protective layers are assembled in B. subtilis. By examining how spores interact with various surfaces, biofilms and microbial consortia, it should be possible to connect what has been learned in the laboratory to what occurs in nature. Future studies that assess how well-conserved the proteins and assembly mechanisms are should help us to determine whether the phenomena observed in B. subtilis coat assembly are applicable to other spore formers. These future goals also bring to mind a key gap in our knowledge: we currently know very little about the diversity in spore coat structure and composition among species. Progress in addressing some of these outstanding questions could be substantially facilitated by analysis of species beyond the familiar models. Importantly, knowledge of the coat assembly mechanisms could also be exploited in various applications with public health implications, such as vaccine development (BOX 2).
Box 2 |. Applications of spore surfaces.
An understanding of spore coat organization may benefit the development of microbial cell surface display technologies, which have been used to produce biocatalysts, biosorbents and vaccines113. Bacillus subtilis is edible and has been used in Japan for centuries to ferment soy beans for the production of natto. Spores are also naturally heat-stable, so spore-based therapies may not require refrigeration, eliminating a major cost of vaccine distribution to developing nations114. Several groups have attempted to generate vaccines against Clostridium tetani by engineering the surface of B. subtilis_spores to display C. tetani antigens, and this has shown some promise115,116. However, the antigens were fused to the outer coat proteins CotB and CotC, which are covered by the crust layer in_B. subtilis. Antigen display on the crust or preparations of crust-free spores, such as the Δ_cotXYZ_ strain, may prove to be more successful strategies for vaccine development. In order to avoid using protein fusions to coat proteins, attempts have been made to use heterologous proteins that can be adsorbed to the spore surface. Nevertheless, this approach would also require a detailed characterization of the spore surface to determine how the different coat layers influence adsorption properties. For example, a recent study indicated that the efficiency of adsorption of β-galactosidase is improved in spores that lack the crust or the outer coat117.
Another avenue to explore in surface-display technologies is the generation of synthetic vesicles encased by selected coat proteins that could be tailor-made to display specific properties. Eukaryotic vesicles are readily produced with purified components in vitro, so it should also be possible to produce vesicles coated with spore coat proteins. A first step in that direction was accomplished with the coat morphogenetic protein SpoVM fused to green fluorescent protein (SpoVM–GFP), which appears to bind preferentially to sites of positive membrane curvature and displays affinity for vesicles close to the size of_B_. subtilis spores (<2 μm)118. By changing a proline to alanine, potentially straightening the SpoVM helix, it was possible to expand the size range of vesicles bound by SpoVM–GFP. Our understanding of the mechanisms of coat assembly in bacteria can be integrated with what is known about coated vesicle formation in eukaryotes. The first step in both events is the localization of proteins to a site of membrane curvature119,120. A common mediator of curvature recognition among the domains of life may be amphipathic α-helices. This feature is common to both SpoVM and the GTPases, along with BAR (Bin–Amphiphysin–Rvs) domain-containing proteins that are involved in the early steps of eukaryotic vesicle formation121. Second, a scaffold of coat material is assembled at the initial site of localization, and protein polymerization proceeds around the circumference of the developing vesicle. Ultimately, the process terminates with membrane fission at the opposite pole of the vesicle.
Acknowledgements
We apologize to colleagues whose work could not be cited in full owing to space limitations. Work in P.E.’s laboratory is supported by grant GM081571 from the US National Institutes of Health (NIH). Work in A.D.’s laboratory is supported by NIH grants R21AI097934 and R01AI093493, and HDTRA1-11-1-0051 from the US Department of Defense.
Glossary
Sporulation
The developmental process of spore formation.
Endospores
Metabolically dormant cells composed of a partially dehydrated central core (containing the genome) surrounded by several concentrically arranged protective layers. An endospore develops inside a mother cell.
Mother cell
The larger of the two compartments formed by asymmetric division of the sporulating cell, it synthesizes most of the building blocks required to assemble the endospore protective structures and lyses at the end of sporulation, releasing the spore into the environment.
Forespore
The smaller of the two compartments that are formed by asymmetric division of the sporulating cell. It matures into an endospore.
Coat
A spore protective structure, which is made up of dozens of proteins. It is usually multilayered, consisting of inner and outer layers.
Sporangium
A vessel in which spores are formed. In endospore formation it refers to a cell that has entered sporulation by dividing asymmetrically.
Engulfment
The morphological transition in sporulation during which the mother cell swallows the forespore in a phagocytosis-like process involving membrane migration. After engulfment is complete, the forespore becomes a cell within the mother cell cytoplasm.
Cortex
A spore protective structure composed of peptidoglycan. It is assembled between the inner and outer forespore membranes.
Crust
The outermost layer of the coat in Bacillus subtilis. It closely follows the contours of the outer coat.
Exosporium
The outermost structure of the spore in several species. It is a protein (and, in at least some cases, a glycoprotein) layer separated from the outer coat by a large gap of irregular width referred to as the interspace.
Bacteriovores
Free-living heterotrophs that feed on bacteria.
Lamellae
The characteristic alternating dark and light rings of the inner coat that are visible by electron microscopy.
Encasement
The morphological transition in spore coat assembly from a cap of coat proteins on the mother cell proximal pole of the forespore to a symmetric distribution around the circumference of the spore.
Injectisomes
In Gram-negative bacteria, a family of secretion systems that have a molecular architecture homologous to flagella.
Natto
A Japanese dish of cooked soy beans fermented by a strain of_Bacillus subtilis_. It has a pungent aroma and a unique texture.
Footnotes
Competing interests statement
The authors declare no competing financial interests.
References
- 1.Brun Y & Shimkets LJ Prokaryotic Development (ASM Press, 2000). [Google Scholar]
- 2.Cohn F Untersuchungen über Bacterien. IV. Beiträge zur Biologie der Bacillen. Beiträge Biol. Pflanzen 7, 249–276 (1877). [Google Scholar]; Along with Koch (below), this paper contains the first (and highly prescient) description of sporulation.
- 3.Koch R Untersuchungen über Bacterien V: Die Aetiologie der Milzbrand-Krankheit, begründet auf die Entwicklungsgeschichte der_Bacillus anthracis_. Beiträge Biol. Pflanzen 7, 277–308 (1877). [Google Scholar]
- 4.Nicholson WL, Munakata N, Horneck G, Melosh HJ & Setlow P Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiol. Mol. Biol. Rev 64, 548–572 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kennedy MJ, Reader SL & Swierczynski LM Preservation records of micro-organisms: evidence of the tenacity of life. Microbiology 140, 2513–2529 (1994). [DOI] [PubMed] [Google Scholar]
- 6.Sneath PH Longevity of micro-organisms. Nature 195, 643–646 (1962). [DOI] [PubMed] [Google Scholar]
- 7.Jacotot H & Virat B La longévité des spores de B. anthracis (premier vaccin de Pasteur). Ann. Inst. Pasteur 87, 215–217 (1954). [PubMed] [Google Scholar]
- 8.de Hoon MJ, Eichenberger P & Vitkup D Hierarchical evolution of the bacterial sporulation network. Curr. Biol 20, R735–R745 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Galperin MY et al. Genomic determinants of sporulation in Bacilli and Clostridia: towards the minimal set of sporulation-specific genes. Environ. Microbiol 14, 2870–2890 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]; A comprehensive study of sporulation gene conservation among endospore formers.
- 10.Earl AM, Losick R & Kolter R Ecology and genomics of Bacillus subtilis. Trends Microbiol. 16, 269–275 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wu M et al. Life in hot carbon monoxide: the complete genome sequence of_Carboxydothermus hydrogenoformans_Z-2901. PLoS Genet. 1, e65 (2005) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hubert C et al. A constant flux of diverse thermophilic bacteria into the cold Arctic seabed. Science 325, 1541–1544 (2009). [DOI] [PubMed] [Google Scholar]
- 13.Eckburg PB et al. Diversity of the human intestinal microbial flora. Science 308, 1635–1638 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fujita M & Losick R Evidence that entry into sporulation in Bacillus subtilis is governed by a gradual increase in the level and activity of the master regulator Spo0A. Genes Dev. 19, 2236–2244 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Molle V et al. The Spo0A regulon of Bacillus subtilis. Mol. Microbiol 50, 1683–1701 (2003). [DOI] [PubMed] [Google Scholar]
- 16.Popham DL Specialized peptidoglycan of the bacterial endospore: the inner wall of the lockbox. Cell. Mol. Life Sci 59, 426–433 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Henriques AO & Moran CP Jr. Structure, assembly, and function of the spore surface layers. Ann. Rev. Microbiol 61, 555–588 (2007). [DOI] [PubMed] [Google Scholar]; A thorough review that includes a comprehensive list of coat proteins in B. subtilis and B. anthracis.
- 18.Driks A The Bacillus anthracis spore. Mol. Aspects Med 30, 368–373 (2009). [DOI] [PubMed] [Google Scholar]
- 19.Vasudevan P, Weaver A, Reichert ED, Linnstaedt SD & Popham DL Spore cortex formation in Bacillus subtilis is regulated by accumulation of peptidoglycan precursors under the control of sigma K. Mol. Microbiol 65, 1582–1594 (2007). [DOI] [PubMed] [Google Scholar]
- 20.Fay A & Dworkin J_Bacillus subtilis_ homologs of MviN (MurJ), the putative Escherichia coli lipid II flippase, are not essential for growth. J. Bacteriol 191, 6020–6028 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.McKenney PT & Eichenberger P Dynamics of spore coat morphogenesis in Bacillus subtilis. Mol. Microbiol 83, 245–260 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study ties the phenomenon of spore encasement to the regulation of expression of individual spore coat genes.
- 22.Aronson AI & Fitz-James P Structure and morphogenesis of the bacterial spore coat. Bacteriol. Rev 40, 360–402 (1976). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Warth AD, Ohye DF & Murrell WG The composition and structure of bacterial spores. J. Cell Biol 16, 579–592 (1963). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tocheva EI et al. Peptidoglycan remodeling and conversion of an inner membrane into an outer membrane during sporulation. Cell 146, 799–812 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McKenney PT et al. A distance-weighted interaction map reveals a previously uncharacterized layer of the Bacillus subtilis spore coat. Curr. Biol 20, 934–938 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Boydston JA, Yue L, Kearney JF & Turnbough CL Jr. The ExsY protein is required for complete formation of the exosporium of Bacillus anthracis. J. Bacteriol 188, 7440–7448 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Giorno R et al. Morphogenesis of the _Bacillus anthracis_spore. J. Bacteriol 189,). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bozue J et al. Bacillus anthracis spores of the_bclA_ mutant exhibit increased adherence to epithelial cells, fibroblasts, and endothelial cells but not to macrophages. Infect. Immun 75, 4498–4505 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chen G, Driks A, Tawfiq K, Mallozzi M & Patil S Bacillus anthracis and Bacillus subtilis spore surface properties and transport. Colloids Surf. B Biointerfaces 76, 512–518 (2010). [DOI] [PubMed] [Google Scholar]
- 30.Kailas L et al. Surface architecture of endospores of the Bacillus cereus/anthracis/thuringiensis family at the subnanometer scale. Proc. Natl Acad. Sci. USA 108, 16014–16019 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study provides a high-resolution characterization of the exosporium structure, revealing a crystalline layer made of a honeycomb-like array of cups.
- 31.Permpoonpattana P et al. Surface layers of _Clostridium difficile_endospores. J. Bacteriol 193, 6461–6470 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Setlow P I will survive: DNA protection in bacterial spores. Trends Microbiol. 15, 172–180 (2007). [DOI] [PubMed] [Google Scholar]
- 33.Hullo MF, Moszer I, Danchin A & Martin-Verstraete I CotA of Bacillus subtilis is a copper-dependent laccase. J. Bacteriol 183, 5426–5430 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu GY & Nizet V Color me bad: microbial pigments as virulence factors. Trends Microbiol. 17, 406–413 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Eisenman HC & Casadevall A Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol 93, 931–940 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Klobutcher LA, Ragkousi K & Setlow P The Bacillus subtilis spore coat provides “eat resistance” during phagocytic predation by the protozoan_Tetrahymena thermophila_. Proc. Natl Acad. Sci. USA 103, 165–170 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Laaberki MH & Dworkin J Role of spore coat proteins in the resistance of Bacillus subtilis spores to _Caenorhabditis elegans_predation. J. Bacteriol 190, 6197–6203 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Carroll AM, Plomp M, Malkin AJ & Setlow P Protozoal digestion of coat-defective Bacillus subtilis spores produces “rinds” composed of insoluble coat protein. Appl. Environ. Microbiol 74, 5875–5881 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Paredes-Sabja D, Setlow P & Sarker MR Germination of spores of Bacillales and_Clostridiales_ species: mechanisms and proteins involved. Trends Microbiol. 19, 85–94 (2011). [DOI] [PubMed] [Google Scholar]
- 40.Shah IM, Laaberki MH, Popham DL & Dworkin J A eukaryotic-like Ser/Thr kinase signals bacteria to exit dormancy in response to peptidoglycan fragments. Cell 135, 486–496 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Driks A_Bacillus subtilis_ spore coat. Microbiol. Mol. Biol. Rev 63, 1–20 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Stewart BT & Halvorson HO Studies on the spores of aerobic bacteria. I. The occurrence of alanine racemase. J. Bacteriol 65, 160–166 (1953). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Steichen C, Chen P, Kearney JF & Turnbough CL Jr. Identification of the immunodominant protein and other proteins of the Bacillus anthracis exosporium. J. Bacteriol 185, 1903–1910 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Todd SJ, Moir AJ, Johnson MJ & Moir A Genes of Bacillus cereus and Bacillus anthracis encoding proteins of the exosporium. J. Bacteriol 185, 3373–3378 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chesnokova ON, McPherson SA, Steichen CT & Turnbough CL Jr. The spore-specific alanine racemase of Bacillus anthracis and its role in suppressing germination during spore development. J. Bacteriol 191, 1303–1310 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pierce KJ, Salifu SP & Tangney M Gene cloning and characterization of a second alanine racemase from Bacillus subtilis encoded by_yncD_. FEMS Microbiol. Lett 283, 69–74 (2008). [DOI] [PubMed] [Google Scholar]
- 47.Butzin XY et al. Analysis of the effects of a gerP mutation on the germination of spores of Bacillus subtilis. J. Bacteriol 194, 5749–5758 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chirakkal H, O’Rourke M, Atrih A, Foster SJ & Moir A Analysis of spore cortex lytic enzymes and related proteins in_Bacillus subtilis_ endospore germination. Microbiology 148, 2383–2392 (2002). [DOI] [PubMed] [Google Scholar]
- 49.Bagyan I & Setlow P Localization of the cortex lytic enzyme CwlJ in spores of_Bacillus subtilis_. J. Bacteriol 184, 1219–1224 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lambert EA & Popham DL The Bacillus anthracis SleL (YaaH) protein is an N-acetylglucosaminidase involved in spore cortex depolymerization. J. Bacteriol 190, 7601–7607 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Imamura D, Kuwana R, Takamatsu H & Watabe K Localization of proteins to different layers and regions of_Bacillus subtilis_ spore coats. J. Bacteriol 192, 518–524 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Buist G, Steen A, Kok J & Kuipers OP LysM, a widely distributed protein motif for binding to (peptido)glycans. Mol. Microbiol 68, 838–847 (2008). [DOI] [PubMed] [Google Scholar]
- 53.Ebmeier SE, Tan IS, Clapham KR & Ramamurthi KS Small proteins link coat and cortex assembly during sporulation in Bacillus subtilis. Mol. Microbiol 84, 682–696 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Santo LY & Doi RH Ultrastructural analysis during germination and outgrowth of_Bacillus subtilis_ spores. J. Bacteriol 120, 475–481 (1974). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Steichen CT, Kearney JF & Turnbough CL Jr. Non-uniform assembly of the _Bacillus anthracis_exosporium and a bottle cap model for spore germination and outgrowth. Mol. Microbiol 64, 359–367 (2007). [DOI] [PubMed] [Google Scholar]; A description of the polarity of the spore envelope and a model for spore germination and outgrowth.
- 56.Holt SC & Leadbetter ER Comparative ultrastructure of selected aerobic spore-forming bacteria: a freeze-etching study. Bacteriol. Rev 33, 346–378 (1969). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Traag BA et al. Do mycobacteria produce endospores? Proc. Natl Acad. Sci. USA 107, 878–881 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Walker JR et al. Clostridium taeniosporum spore ribbon-like appendage structure, composition and genes. Mol. Microbiol 63, 629–643 (2007). [DOI] [PubMed] [Google Scholar]
- 59.Lequette Y et al. Role played by exosporium glycoproteins in the surface properties of Bacillus cereus spores and in their adhesion to stainless steel. Appl. Environ. Microbiol 77, 4905–4911 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Buhr TL, Young AA, Minter ZA, Wells CM & Shegogue DA Decontamination of a hard surface contaminated with_Bacillus anthracis_ΔSterne and B. anthracis Ames spores using electrochemically generated liquid-phase chlorine dioxide (eClO2). J. Appl. Microbiol 111, 1057–1064 (2011). [DOI] [PubMed] [Google Scholar]
- 61.Siala A, Hill IR & Gray TRG Populations of spore-forming bacteria in an acid forest soil, with special reference to Bacillus subtilis. J. Gen. Microbiol 8, 183–190 (1974). [Google Scholar]
- 62.Nicholson WL Roles of Bacillus endospores in the environment. Cell. Mol. Life Sci 59, 410–416 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Donovan W, Zheng LB, Sandman K & Losick R Genes encoding spore coat polypeptides from Bacillus subtilis. J. Mol. Biol 196, 1–10 (1987). [DOI] [PubMed] [Google Scholar]
- 64.Beall B, Driks A, Losick R & Moran CP Jr. Cloning and characterization of a gene required for assembly of the Bacillus subtilis spore coat. J. Bacteriol 175, 1705–1716 (1993). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zheng LB, Donovan WP, Fitz-James PC & Losick R Gene encoding a morphogenic protein required in the assembly of the outer coat of the _Bacillus subtilis_endospore. Genes Dev. 2, 1047–1054 (1988). [DOI] [PubMed] [Google Scholar]
- 66.Lai EM et al. Proteomic analysis of the spore coats of Bacillus subtilis and Bacillus anthracis. J. Bacteriol 185, 1443–1454 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kuwana R et al. Proteomics characterization of novel spore proteins of_Bacillus subtilis_. Microbiology 148, 3971–3982 (2002). [DOI] [PubMed] [Google Scholar]
- 68.Eichenberger P et al. The program of gene transcription for a single differentiating cell type during sporulation in Bacillus subtilis. PLoS Biol. 2, e328 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kim H et al. The Bacillus subtilis spore coat protein interaction network. Mol. Microbiol 59, 487–502 (2006). [DOI] [PubMed] [Google Scholar]
- 70.Abhyankar W et al. Gel-free proteomic identification of the Bacillus subtilis insoluble spore coat protein fraction. Proteomics 11, 4541–4550 (2011). [DOI] [PubMed] [Google Scholar]
- 71.Webb CD, Decatur A, Teleman A & Losick R Use of green fluorescent protein for visualization of cell-specific gene expression and subcellular protein localization during sporulation in Bacillus subtilis. J. Bacteriol 177, 5906–5911 (1995). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Eichenberger P et al. The σE regulon and the identification of additional sporulation genes in Bacillus subtilis. J. Mol. Biol 327, 945–972 (2003). [DOI] [PubMed] [Google Scholar]
- 73.Piggot PJ & Coote JG Genetic aspects of bacterial endospore formation. Bacteriol. Rev 40, 908–962 (1976). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Levin PA et al. An unusually small gene required for sporulation by_Bacillus subtilis_. Mol. Microbiol 9, 761–771 (1993). [DOI] [PubMed] [Google Scholar]
- 75.Roels S, Driks A & Losick R Characterization of spoIVA, a sporulation gene involved in coat morphogenesis in Bacillus subtilis. J. Bacteriol 174, 575–585 (1992). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Takamatsu H, Kodama T, Nakayama T & Watabe K Characterization of the yrbA gene of_Bacillus subtilis_, involved in resistance and germination of spores. J. Bacteriol 181, 4986–4994 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ozin AJ, Henriques AO, Yi H & Moran CP Jr. Morphogenetic proteins SpoVID and SafA form a complex during assembly of the Bacillus subtilis spore coat. J. Bacteriol 182, 1828–1833 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Driks A, Roels S, Beall B, Moran CP Jr & Losick R Subcellular localization of proteins involved in the assembly of the spore coat of Bacillus subtilis. Genes Dev. 8, 234–244 (1994). [DOI] [PubMed] [Google Scholar]; This article proposes a seminal model for coat assembly.
- 79.Imamura D, Kuwana R, Takamatsu H & Watabe K Proteins involved in formation of the outermost layer of_Bacillus subtilis_ spores. J. Bacteriol 193, 4075–4080 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Redmond C, Baillie LW, Hibbs S, Moir AJ & Moir A Identification of proteins in the exosporium of Bacillus anthracis. Microbiology 150, 355–363 (2004). [DOI] [PubMed] [Google Scholar]
- 81.Waller LN, Fox N, Fox KF, Fox A & Price RL Ruthenium red staining for ultrastructural visualization of a glycoprotein layer surrounding the spore of Bacillus anthracis and Bacillus subtilis. J. Microbiol. Methods 58, 23–30 (2004). [DOI] [PubMed] [Google Scholar]; This study introduces the use of ruthenium red staining to analyse the outermost layer of the spore.
- 82.Cutting S et al. SpoVM, a small protein essential to development in_Bacillus subtilis_, interacts with the ATP-dependent protease FtsH. J. Bacteriol 179, 5534–5542 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Wang KH et al. The coat morphogenetic protein SpoVID is necessary for spore encasement in Bacillus subtilis. Mol. Microbiol 74, 634–649 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ozin AJ, Samford CS, Henriques AO & Moran CP Jr. SpoVID guides SafA to the spore coat in Bacillus subtilis. J. Bacteriol 183, 3041–3049 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Costa T, Isidro AL, Moran CP Jr & Henriques AO Interaction between coat morphogenetic proteins SafA and SpoVID. J. Bacteriol 188, 7731–7741 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Price KD & Losick R A four-dimensional view of assembly of a morphogenetic protein during sporulation in Bacillus subtilis. J. Bacteriol 181, 781–790 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ramamurthi KS, Clapham KR & Losick R Peptide anchoring spore coat assembly to the outer forespore membrane in Bacillus subtilis. Mol. Microbiol 62, 1547–1557 (2006). [DOI] [PubMed] [Google Scholar]
- 88.van Ooij C, Eichenberger P & Losick R Dynamic patterns of subcellular protein localization during spore coat morphogenesis in Bacillus subtilis. J. Bacteriol 186, 4441–4448 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Pogliano K, Harry E & Losick R Visualization of the subcellular location of sporulation proteins in Bacillus subtilis using immunofluorescence microscopy. Mol. Microbiol 18, 459–470 (1995). [DOI] [PubMed] [Google Scholar]
- 90.van Ooij C & Losick R Subcellular localization of a small sporulation protein in_Bacillus subtilis_. J. Bacteriol 185, 1391–1398 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Mullerova D, Krajcikova D & Barak I Interactions between Bacillus subtilis early spore coat morphogenetic proteins. FEMS Microbiol. Lett 299, 74–85 (2009). [DOI] [PubMed] [Google Scholar]
- 92.de Francesco M et al. Physical interaction between coat morphogenetic proteins SpoVID and CotE is necessary for spore encasement in Bacillus subtilis. J. Bacteriol 194, 4941–4950 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ramamurthi KS & Losick R ATP-driven self-assembly of a morphogenetic protein in_Bacillus subtilis_. Mol. Cell 31, 406–414 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Little S & Driks A Functional analysis of the _Bacillus subtilis_morphogenetic spore coat protein CotE. Mol. Microbiol 42, 1107–1120 (2001). [DOI] [PubMed] [Google Scholar]
- 95.Krajcikova D, Lukacova M, Mullerova D, Cutting SM & Barak I Searching for protein-protein interactions within the_Bacillus subtilis_ spore coat. J. Bacteriol 191, 3212–3219 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Kroos L, Kunkel B & Losick R Switch protein alters specificity of RNA polymerase containing a compartment-specific sigma factor. Science 243, 526–529 (1989). [DOI] [PubMed] [Google Scholar]
- 97.Zhang J, Fitz-James PC & Aronson AI Cloning and characterization of a cluster of genes encoding polypeptides present in the insoluble fraction of the spore coat of_Bacillus subtilis_. J. Bacteriol 175, 3757–3766 (1993). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Zhang J, Ichikawa H, Halberg R, Kroos L & Aronson AI Regulation of the transcription of a cluster of Bacillus subtilis spore coat genes. J. Mol. Biol 240, 405–415 (1994). [DOI] [PubMed] [Google Scholar]
- 99.Zheng LB & Losick R Cascade regulation of spore coat gene expression in_Bacillus subtilis_. J. Mol. Biol 212, 645–660 (1990). [DOI] [PubMed] [Google Scholar]
- 100.Ragkousi K & Setlow P Transglutaminase-mediated cross-linking of GerQ in the coats of_Bacillus subtilis_ spores. J. Bacteriol 186, 5567–5575 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Naclerio G, Baccigalupi L, Zilhao R, De Felice M & Ricca E_Bacillus subtilis_ spore coat assembly requires_cotH_ gene expression. J. Bacteriol 178, 4375–4380 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.McPherson DC et al. Characterization of the Bacillus subtilis spore morphogenetic coat protein CotO. J. Bacteriol 187, 8278–8290 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Giglio R et al. Organization and evolution of the cotG and_cotH_ genes of Bacillus subtilis. J. Bacteriol 193, 6664–6673 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Isticato R et al. CotC-CotU heterodimerization during assembly of the_Bacillus subtilis_ spore coat. J. Bacteriol 190, 1267–1275 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Erhardt M, Namba K & Hughes KT Bacterial nanomachines: the flagellum and type III injectisome. Cold Spring Harb. Perspect. Biol 2, a000299 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ohye DF & Murrell WG Exosporium and spore coat formation in Bacillus cereus T. J. Bacteriol 115, 1179–1190 (1973). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Thompson BM & Stewart GC Targeting of the BclA and BclB proteins to the Bacillus anthracis spore surface. Mol. Microbiol 70, 421–434 (2008). [DOI] [PubMed] [Google Scholar]
- 108.Angert ER Alternatives to binary fission in bacteria. Nature Rev. Microbiol 3, 214–224 (2005). [DOI] [PubMed] [Google Scholar]
- 109.Eldar A et al. Partial penetrance facilitates developmental evolution in bacteria. Nature 460, 510–514 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Flardh K & Buttner MJ_Streptomyces_ morphogenetics: dissecting differentiation in a filamentous bacterium. Nature Rev. Microbiol 7, 36–49 (2009). [DOI] [PubMed] [Google Scholar]
- 111.Sudo SZ & Dworkin M Comparative biology of prokaryotic resting cells. Adv. Microb. Physiol 9, 153–224 (1973). [DOI] [PubMed] [Google Scholar]
- 112.Muller FD, Schink CW, Hoiczyk E, Cserti E & Higgs PI Spore formation in Myxococcus xanthus is tied to cytoskeleton functions and polysaccharide spore coat deposition. Mol. Microbiol 83, 486–505 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Wu CH, Mulchandani A & Chen W Versatile microbial surface-display for environmental remediation and biofuels production. Trends Microbiol. 16, 181–188 (2008). [DOI] [PubMed] [Google Scholar]
- 114.Cutting SM, Hong HA, Baccigalupi L & Ricca E Oral vaccine delivery by recombinant spore probiotics. Int. Rev. Immunol 28, 487–505 (2009). [DOI] [PubMed] [Google Scholar]
- 115.Permpoonpattana P et al. Immunization with Bacillus spores expressing toxin A peptide repeats protects against infection with Clostridium difficile strains producing toxins A and B. Infect. Immun 79, 2295–2302 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Amuguni H et al. Sublingual immunization with an engineered Bacillus subtilis strain expressing tetanus toxin fragment C induces systemic and mucosal immune responses in piglets. Microbes Infect. 14, 447–456 (2012). [DOI] [PubMed] [Google Scholar]
- 117.Sirec T et al. Adsorption of beta-galactosidase of Alicyclobacillus acidocaldarius on wild type and mutants spores of_Bacillus subtilis_. Microb. Cell Fact 11, 100 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ramamurthi KS, Lecuyer S, Stone HA & Losick R Geometric cue for protein localization in a bacterium. Science 323, 1354–1357 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study reveals that the coat morphogenetic protein SpoVM has the ability to recognize positive membrane curvature.
- 119.Huang KC & Ramamurthi KS Macromolecules that prefer their membranes curvy. Mol. Microbiol 76, 822–832 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Pucadyil TJ & Schmid SL Conserved functions of membrane active GTPases in coated vesicle formation. Science 325, 1217–1220 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Bhatia VK, Hatzakis NS & Stamou D A unifying mechanism accounts for sensing of membrane curvature by BAR domains, amphipathic helices and membrane-anchored proteins. Semin. Cell Dev. Biol 21, 381–390 (2010). [DOI] [PubMed] [Google Scholar]
- 122.Masayama A et al. A novel lipolytic enzyme, YcsK (LipC), located in the spore coat of Bacillus subtilis, is involved in spore germination. J. Bacteriol 189, 2369–2375 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Bourne N, FitzJames PC & Aronson AI Structural and germination defects of Bacillus subtilis spores with altered contents of a spore coat protein. J. Bacteriol 173, 6618–6625 (1991). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Costa T, Steil L, Martins LO, Volker U & Henriques AO Assembly of an oxalate decarboxylase produced under σK control into the _Bacillus subtilis_spore coat. J. Bacteriol 186, 1462–1474 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Henriques AO, Beall BW & Moran CP Jr. CotM of Bacillus subtilis, a member of the α-crystallin family of stress proteins, is induced during development and participates in spore outer coat formation. J. Bacteriol 179, 1887–1897 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Takamatsu H et al. A spore coat protein, CotS, of _Bacillus subtilis_is synthesized under the regulation of σK and GerE during development and is located in the inner coat layer of spores. J. Bacteriol 180, 2968–2974 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Scheeff ED et al. Genomics, evolution, and crystal structure of a new family of bacterial spore kinases. Proteins 78, 1470–1482 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]