Transforming growth factor-β and breast cancer: Mammary gland development (original) (raw)
Abstract
Transforming growth factor (TGF)-β1 is a pluripotent cytokine that profoundly inhibits epithelial proliferation, induces apoptosis, and influences morphogenesis by mediating extracellular matrix deposition and remodeling. The physiologic roles of the action of TGF-β in mammary gland, indeed in most tissues, are poorly understood. In order to understand the actions of TGF-β, we need to take into account the complexity of its effects on different cell types and the influence of context on cellular responses. This task is further compounded by multiple mechanisms for regulating TGF-β transcription, translation, and activity. One of the most significant factors that obscures the action of TGF-β is that it is secreted as a stable latent complex, which consists of the 24-kDa cytokine and the 80-kDa dimer of its prepro region, called latency-associated peptide. Latency imposes a critical restraint on TGF-β activity that is often overlooked.The extracellular process known as activation, in which TGF-β is released from the latent complex, is emphasized in the present discussion of the role of TGF-β in mammary gland development. Definition of the spatial and temporal patterns of latent TGF-β activation in situ is essential for understanding the specific roles that TGF-β plays during mammary gland development, proliferation, and morphogenesis.
Keywords: transforming growth factor (TGF)-β1, activation, latent, mammary, proliferation
Introduction
This review describes briefly mammary gland development and discusses the postulated role of TGF-β1 in mammary gland development as evidenced by its effects in mammary epithelial cell culture and animal studies. TGF-β has been strongly implicated in the control of mammary epithelial growth [1] and in breast cancer cells [2,3]. Cell culture studies have also shown that the effects of TGF-β are pleiotropic, context-dependent, concentration-dependent, and can be indirect. As a consequence it is difficult to predict the endogenous action of TGF-β in mammary gland. However, mammary gland mouse models in which exogenous TGF-β or blocking antibodies are administered indicate that regulation of growth and morphogenesis is a potential function of TGF-β (Fig. 1). Transgenic models in which the activity of TGF-β is manipulated have confirmed this conclusion. Interpretation of these animal studies are confounded by the fact that the lack of knowledge about when and where TGF-β is active in situ, however. Preliminary studies from our laboratory have provided an indication of the complexity that awaits analysis of in situ activity.
Figure 1.
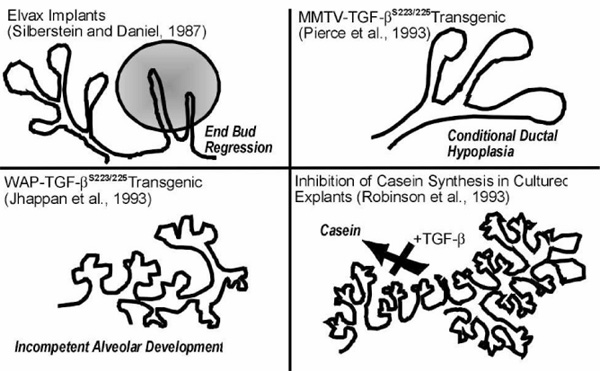
Effects of transforming growth factor (TGF)-β in mammary gland. A variety of studies suggest that TGF-β contributes to morphogenesis, growth, and function in mouse mammary gland.
Mammary gland development
The seminal work of Daniel and coworkers [4,5] focused attention on the role of TGF-β in mammary gland development soon after its characterization. The morphologic and functional development of the mammary gland is largely postnatal in both humans and mice (for review [6]). Under the effects of hormones of puberty, which begins just after weaning in mice and continues for several weeks (ie 3 weeks to 6-8 weeks of age), the mammary tree is established within an adipose stroma known as the fat pad. The epithelium, usually a simple epithelium consisting of one or two cell layers, is ensheathed in a fibrous stroma. This period of growth is characterized by a specialized morphologic unit called the end-bud, which is a multicellular, multilayered structure whose function is to extend the ductal epithelial tree to the boundaries of the fat pad. The pattern of the resulting tissue can be visualized in a whole-mount preparation and is often used as an end-point for evaluating the role of factors such as TGF-β in guiding morphology by measuring the extent of branching, the branching pattern by branch-point analysis, and the distribution of end-buds and their character, size, shape, and persistence. Once the ductal tree is established, repeated estrus cycles of ovarian hormones elicit further elaboration of the epithelium in some mouse strains, generating small lateral branch points. In a nulliparous animal approximately 10% of the tissue is epithelial.
Under the effects of hormones of pregnancy, a burst of growth and differentiation results in lobuloalveolar differentiation of the epithelium into functional secretory units that produce milk that is carried to the teat by large conducting ducts. The resulting fully functional gland is 90% epithelial. Upon weaning, a process of involution destroys the majority of secretory epithelium, leaving the ductal tree extant for repeated cycles of growth and differentiation.
The remarkable capacity of the mammary epithelium to undergo development and differentiation provides a research model in which the factors that influence growth, morphologic patterning, proliferation, and differentiation can readily be explored. Furthermore, in mice epithelium can be transplanted to new stroma that is usually cleared of endogenous epithelium, which offers an exceptional research tool for studying the contribution of the stroma to the control of epithelial function and remodeling. Recombinations of transgenic and wild-type epithelia and stromas, and the ability to transplant intact fat pads to new hosts, allows determination of whether the source of a particular factor (ie different tissue compartments) affects the maturation of the mammary gland.
Transforming growth factor-beta regulation and expression
There are three mammalian isoforms of TGF-β. All three isoforms bind to the same cell surface receptors and, in cell culture, often appear to elicit similar responses. For the purposes of this review we focus on the well-studied TGF-β1 isoform, its localization in the mammary gland, its effects in cell culture models, and the consequences of manipulating its activity in vivo by various means. The activity of all TGF-β isoforms is restrained by secretion as latent complexes [latent transforming growth factor (LTGF)-β] [7]. (Regarding nomenclature, TGF-β refers to the active cytokine or its effects, whereas LTGF-β designates the latent form.) LTGF-β consists of the 24-kDa cytokine and a 80-kDa dimer of its prepro region called latency-associated peptide (LAP), which contains the signal sequence for secretion (Fig. 2). To our knowledge, all cells secrete LTGF-β, which underscores the importance of activation events associated with the release of TGF-β from LAP.
Figure 2.
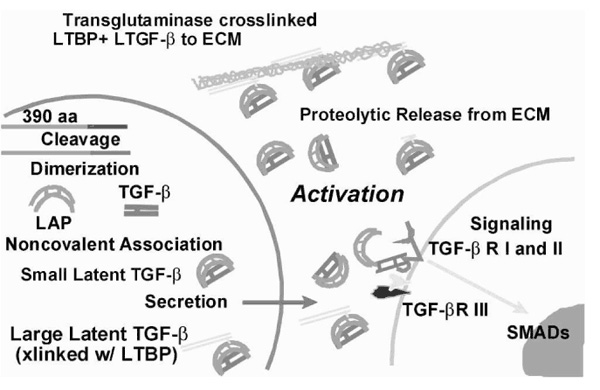
Transforming growth factor (TGF)-β production, secretion, and activation. Some elements of protein processing and post-translation modifications involved in the regulatory control of TGF-β are depicted. The TGF-β1 gene encodes a 390 amino acid polypeptide that is cleaved into two polypeptides that form homodimers during protein processing: latency-associated peptide (LAP) and TGF-β. These homodimers are noncovalently associated to form the small latent TGF-β complex, which is secreted. Alternatively, this complex can be covalently linked by disulfide bonds to a latent TGF-β binding protein (LTBP) before secretion. LTBP provides means of anchoring latent TGF (LTGF)-β in the extracellular matrix (ECM), which may involve cross-linking by transglutaminase and which requires proteolytic processing to release LTGF-β before activation. Activation occurs extracellularly to release TGF-β at or near the cell surface so that it immediately binds to its receptors. TGF-β receptors I and II form a heterocomplex that signals via the SMAD signal transduction protein family.TGF-β receptor III, also known as betaglycan, is nonsignaling but may be involved in presenting TGF-β to its signaling receptors.
TGF-β message levels do not usually reflect protein production, or, more importantly, protein activity. For example, tamoxifen increases both LTGF-β production and activation without any effect on mRNA levels [2]. Differences in mRNA translation are in part due to a 5'-untranslated region stem-loop that binds a regulatory protein, which can result in increased protein production by factors of 10 without changes in mRNA levels [8]. In addition, protein assays are complicated by the fact that tissue extraction protocols activate the endogenous LTGF-β [9]. Because the biologic activity of TGF-β is controlled by its release from the latent complex, elevated expression of latent complex is not likely to have biologic consequences, whereas increased activation, even without changes in synthesis, will profoundly affect physiologic events [10]. The importance of activation is supported by the lack of a phenotype in transgenic mice that overexpress LTGF-β in mammary gland [11]. A recent study in TGF-β-null heterozygotes, however, has shown there is a dramatic loss of circulating and tissue TGF-β levels as a result of the haploid genotype [12]. This surprising finding indicates that TGF-β plays a significant role in its own regulation.
A variety of immunostaining studies have been conducted in both murine and human tissues. These are difficult to summarize because of differences in tissue preparation, the antibodies employed, and their uncertain relevance to understanding the functional role of TGF-β in situ. Several studies localized TGF-β to the human breast epithelium and stroma [13,14], but at least one study [15] found it to be associated only with epithelial cells; this is probably due to either the antibody or fixative (see below). TGF-β is also found in the conditioned media of cultured normal human epithelial cells [16,17,18] and fibroblasts [18,19]. A recent study that immunolocalized TGF-β receptors I, II, and III in normal human breast [20] demonstrated that these are widely expressed in the breast epithelium and stroma.
Robinson et al [21] used Northern blot analyses, in situ hybridization, and immunocytochemistry to define TGF-β expression patterns during murine mammary gland development and differentiation to establish that all three mammalian TGF-β isoforms are expressed in the epithelium during all phases of mammary development. TGF-β2 was less abundant, however, whereas TGF-β3 was the only isoform found in myoepithelial cells. TGF-β1 transcripts and immunoreactivity decreased during pregnancy, but those of TGF-β2 and TGF-β3 increased until the onset of lactation [210]. Intraepithelial immunoreactivity in that study corresponded to transcript localization, indicating sites of synthesis. Immunolocalization using antibodies to mature TGF-β versus TGF-β1 LAP showed an interesting pattern of deposition that was suggested to indicate that TGF-β activity inhibits proliferation and its depletion is necessary for growth [22]. Although activation was implicit in the interpretation of the immunostaining, the model did not provide confirmation of activity, due in large part to the lack of reagents to differentially localize active TGF-β versus LTGF-β.
Studies in our laboratory demonstrate that under certain conditions some antibodies can discriminate between active and latent TGF-β [23]. We determined the specificity of TGF-β antibodies by using a immunodetection method that preserves endogenous LTGF-β in conjunction with engineered control tissues that specifically produce latent versus constituitively active TGF-β [24]. Our extensive screening of available TGF-β antibodies demonstrated that antibody source, fixation, and tissue preparation influences the ability to discriminate between active and latent forms [25]; thus, retrospective interpretation of other immunostaining protocols and antibodies combinations is not possible.
One of the most striking features of our immunolocalization in mammary gland using antibodies that discriminate between TGF-β and LTGF-β is the contrast between the highly restricted localization of TGF-β compared with the broad distribution of LTGF-β. Our initial studies concerned the effect of ionizing radiation on the mammary gland microenvironment. We observed that the immunoreactivity of activation-specific antibodies LC(1-30) and CC(1-30) is restricted to the normal mammary epithelium [26], whereas LAP immunoreactivity is widely distributed in the epithelium, fibrous stroma, and adipose stroma [24]. A remarkably rapid (within 1 h) shift from predominantly LAP to TGF-β immunoreactivity in all of these tissue compartments led us to conclude that ionizing radiation leads to LTGF-β activation [24]. Our conclusion is supported by additional studies that demonstrated that radiation induces stromal extracellular matrix (ECM) remodeling (eg collagen III), a known target of TGF-β action [26], and that administration of TGF-β-neutralizing antibodies blocks ECM remodeling in irradiated animals [27]. These studies indicated that, despite the apparent abundance of LTGF-β in normal, nulliparous mammary gland, active TGF-β is highly localized.
Transforming growth factor-beta responses in cultured cell models
Much of our understanding of the biology of TGF-β comes from studies of cultured cells exposed to recombinant or purified active TGF-β. Nonetheless, the specific physiologic roles of the action of TGF-β in mammary gland are not well understood, which is due in part to the complexity of its effects on different cell types that is compounded by regulation at multiple levels and modified by cellular context [23]. Examples of these effects are summarized below. TGF-β has been shown to inhibit profoundly proliferation of normal rodent and human mammary epithelial cells in cell culture, and may also modify the production of differentiated proteins (for review [28]). Both cell type and context influence TGF-β action. 3T3 cell preadipocytes spontaneously activate their own secreted LTGF-β, whereas mature adipocytes do not, which implicates TGF-β activation as a potent negative regulator of adipocyte differentiation [29]. Fetal versus adult fibroblasts [30], young versus old smooth muscle cells [31], and normal human mammary epithelial cells grown on ECM versus plastic [32] are differentially sensitive to TGF-β growth inhibition. In addition to reversible growth inhibition, TGF-β can also induce apoptosis. Hormone withdrawal in hormone-dependent tissues such as prostate [33] and uterus [34] is a potent inducer of TGF-β-mediated apoptosis. In mammary gland, TGF-β mRNA increases early in involution and its activity is thought to mediate this apoptotic remodeling process [35]. Notably, mammary secretions during involution contain high levels of active TGF-β and extremely high levels of LTGF-β that regulate the mononuclear phagocytes involved in apoptotic remodeling of the gland [36].
TGF-β-induced ECM contributes to growth control in cultured cells [37] and may be involved in the changes in ECM during mammary gland development [13,22] and the menstrual cycle [38]. The cessation of DNA synthesis and end-bud regression caused by exogenous TGF-β-induced ECM is postulated to involve the concomitant induction of periepithelial ECM around end-buds [39]. This regulation may be offset by feedback mechanisms as evidenced by suppression of TGF-β-induced ECM mRNA levels in murine mammary cells cultured within a basement membrane-type ECM [40]. Even cells that lack a proliferative response to TGF-β-induced ECM, such as breast cancer cells, may still respond with specialized ECM production [16], that in turn may modify the tissue microenvironment in such a way as to affect growth. For example, recent mammary cell culture studies [41] also point to a role of ECM in modulating hormone response. The antiestrogen tamoxifen affects stromal production and activity of TGF-β, which has been postulated to contribute to its therapeutic benefit [42]. An interesting model of hormone-mediated remodeling in uterine tissue indicates that stromal activation of TGF-β mediates epithelial production of the proteases that are necessary for degradation of basement membrane [43].
The addition of TGF-β to mammary cell cultures supports a role in regulation of morphogenesis and control of differentiation. TGF-β suppressed the ability of mammary explants cultured with lactogenic hormones to secrete casein [44]. It abolished ductal morphogenesis by human mammary epithelial cells cultured within a basement membrane-type ECM, whereas antibodies to TGF-β stimulated duct formation [45]. In another study [46], however, cultured mammary cell lines exhibited a biphasic response to TGF-β concentration; picomolar concentrations inhibited branching morphogenesis, whereas fentomolar concentrations stimulated it. Furthermore, chronic TGF-β exposure can elicit phenotypic transformation under certain conditions, resulting in the acquisition of mesenchymal-like transformation of mammary epithelial cells [47], or myofibroblast characteristics in stromal cells [48]. The role of such phenomena in vivo remains obscure, but may contribute to desmoplasia and invasive behavior in neoplasia.
Manipulating transforming growth factor-beta activity in the mammary gland
The consequences of endogenous TGF-β activity in the mammary gland have been evaluated by using exogenous delivery of TGF-β or neutralizing antibodies. The pivotal observations by Daniel and coworkers [4,5], who administered exogenous TGF-β via diffusion from miniature inorganic pellets, showed that end-buds undergo reversible regression during puberty, whereas alveolar buds in pregnancy, which are also actively proliferating, do not. During puberty, end-bud DNA synthesis was profoundly inhibited within 12 h of exposure to TGF-β-containing implants, and by 48 h this resulted in an 80-90% decrease in end-bud number [4]. In contrast, DNA synthesis in distal ductal epithelium and fibroblasts was unaffected, indicating an underlying difference in responses and/or access to these cells. End-bud regression was preceded by the deposition of a thick, collagen- and glycosoaminoglycan-rich ECM, which was postulated to force premature differentiation [49]. This selective regression of end-buds was reversible if the implant was removed, even after long-term exposures [5], indicating that the gland did not become refractory or lose its multipotent stem-cell population. In contrast, exogenous TGF-β did not inhibit lobuloalveolar growth, morphogenesis or differentiation. This stage specificity belies a simple role for TGF-β as a DNA synthesis inhibitor and points to the possible differences in how these two actively proliferating structures are regulated. It may also indicate that endogenous TGF-β activity is differentially regulated during these distinct phases of concomitant mammary gland growth and differentiation.
Transgenic manipulation of TGF-β in mammary gland underscores the importance of defining when and where the protein is active. As mentioned above, overexpression of LTGF-β does not produce a mammary phenotype. TGF-β can be produced in a constitutively active form by expression of a construct in which the cysteines at positions 223 and 225 are mutated to serines, however, preventing dimerization of LAP [50]. Two transgenic models have been created with dramatically different phenotypes, depending on the promoter that is used to drive expression of the transgene. Murine mammary tumor virus (MMTV)-long terminal repeat promoter is used to direct transgene expression to the mammary epithelium. When TGF-β223-225 is expressed on an MMTV promoter, the gland is transiently hypoplastic during ductal morphogenesis, but recovers and is able to lactate and support offspring without apparent difficulty [11]. It has been suggested by Smith [51] that additional expression of the MMTV-driven transgene in the salivary gland may influence this phenotype. If the mutated constitutively active TGF-β is driven by the whey acidic protein (WAP) promoter, however, a milk protein that is highly expressed during pregnancy and lactation, ductal morphogenesis is unaffected, but alveolar development is greatly compromised [52]
Subsequent experiments demonstrated that proliferation was uninhibited but that apoptosis was stimulated during both estrous and pregnancy [53]. Furthermore, transplant experiments indicated that the action of TGF-β was intrinsic to the epithelial cells and acted in an autocrine manner. For example, wild-type epithelium was unaffected and differentiated normally [53]. Taking full advantage of the mammary model, Kordon et al [53] used serial transplantation of WAP-TGF-β223-225 epithelium to show that the mammary stem cells were significantly compromised by the transgene expression. Because endogenous WAP protein or reporter expression is restricted to specific luminal epithelial cells and is regulated by the estrus cycle, those investigators postulated that WAP-TGF-β223-225 expression targeted the immediate daughters of mammary epithelial cells, causing inappropriate apoptosis and subsequent depletion of this compartment. The severity of the MMTV-TGF-β223-225 phenotype may also be influenced by the timing or cell specificity of expression relative to that of endogenous LTGF-β activation. Our preliminary data using antibodies that discriminate between active and latent TGF-β supports this concept in that TGF-β activation is suppressed by mid to late pregnancy. Thus, WAP-β223-225 would be driving inappropriate TGF-β activity at a point when it is normally inhibited.
The question of how mammary glands of TGF-β-null mice develop has not yet been addressed because the null mice die around the time of mammary gland development [54]. These mice have provided evidence of a previously unrecognized aspect of TGF-β biology, however. Letterio et al [55] demonstrated that homozygous TGF-β-null off-spring are protected from the gross inflammation that leads to their early death by maternal transfer of TGF-β via the placenta and milk. Another surprising aspect of the TGF-β1-null mouse is that the haploid genotype leads to greatly reduced TGF-β production, which is reflected by serum levels that are 11% of the levels in wild-type animals, indicating that TGF-β regulates its own production and/or stability. This chronic depletion of the latent complex pools presumably restricts TGF-β activity, and results in a subtly altered proliferative phenotype in liver and lung. A further consequence is increased tumor development, indicating that TGF-β is sufficiently depleted to impede its action as a tumor suppressor [12].
We recently evaluated the mammary gland phenotype of TGF-β-null heterozygote mutant mice (Barcellos-Hoff MH, et al, unpublished data). Mammary ductal outgrowth during puberty was accelerated, and mammary epithelial proliferation was increased in young TGF-β1 haploid genotype animals. This effect appeared to be stage-specific in that mammary gland whole mounts from adult nulliparous mice were grossly normal in morphology and functionally intact. Despite the apparently normal morphology, we found that the frequency of proliferating cells of the mammary epithelium is significantly elevated in mammary gland, as occurs in other tissues like liver from TGF-β-null mutant heterozygotes [12].
It is unknown at this time whether systemic, stromal, epithelial or all sources of TGF-β are critical in defining the rate of growth in mammary gland. Gorska et al [56] addressed this question by overexpressing a dominant-negative kinase-deficient TGF-β type II receptor driven by MMTV-long terminal repeat, resulting in primarily epithelial expression, or metallothionein-derived promoter, which is expressed in the stroma. The dominant-negative receptor impedes signaling by all TGF-β isoforms, but circumvents the issues raised by constitutively active TGF-β transgenes. Blunting the response to TGF-β in the mammary epitheliuem resulted in alveolar hyperplasia and premature functional differentiation. In contrast, inhibition of the stromal response to TGF-β by this means resulted in increased ductal branching [57]. Thus, both stromal and epithelial responses to TGF-β activity play a role in defining the maturation of mammary tissue.
The question of when and where LTGF-β is activated during normal development and homeostasis remains to be addressed, however. We recently developed dual immunofluorescence of LAP antibodies and activation-specific chNTGF-β antibodies [27] and have begun to study the pattern of localization during mouse mammary gland development and differentiation (Fig. 3). Although LAP immunoreactivity (green) is relatively uniform, a heterogeneous pattern of TGF-β staining (yellow-orange) is observed in the epithelium of adult mammary glands (Barcellos-Hoff MH et al, unpublished data). We have examined mammary glands from a variety of developmental states to determine the physiologic correlates of this pattern. We found that mammary epithelial TGF-β immunoreactivity was heterogeneous and most intense during periods of proliferation and morphogenesis, suggesting the presence of a distinct subpopulation. Further characterization of these cells and determination of their ultimate fate in terms of proliferation and differentiation may be informative regarding the cellular mechanisms by which pattern and function are established during mammary gland development.
Figure 3.
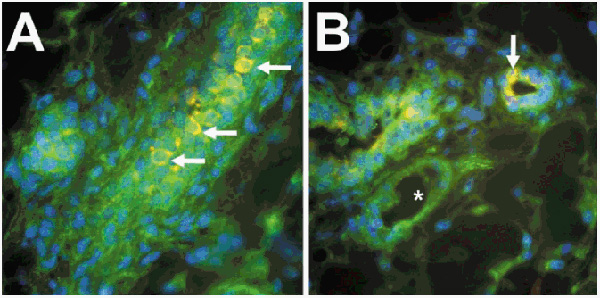
Dual immunolocalization of latency-associated peptide (LAP) and transforming growth factor (TGF)-β in murine mammary gland. Antibodies to LAP (green) to localize latent TGF (LTGF)-β, antigen-purified TGF-β antibodies that specifically detect active TGF-β (red), and DAPI stained nuclei (blue) were visualized using tricolor digital fluorescence microscopy. (A) A section tangential to a duct; (B) a transverse and cross-section of a duct of mammary gland from a nulliparous Balb/c mouse. Colocalization of LAP and TGF-β appears yellow in certain epithelial cells (arrows). Note that certain cells have prominent LAP staining, but are not immunoreactive for TGF-β and are adjacent to cells that exhibit staining of both LAP and TGF-β. Neither stromal, myoepithelial, or endothelial (B, asterisk) cells show prominent TGF-β immunoreactivity.
Conclusion
It is clear from a variety of studies in cell culture that TGF-β can have myriad effects, many of which complement each other, but others that appear paradoxic. In vivo studies using exposure to exogenous sources or transgenic expression of constitutively active protein have shown what TGF-β can do in the mammary gland. These have generally led to the conclusion that TGF-β has a prominent role in regulating pattern formation by the epithelium, perhaps via interactions with the stroma, and is involved in fate decisions by individual cells. In the great majority of animal and cell culture studies, however, activation of the ubiquitous latent TGF-β complex, which exercises fundamental control of TGF-β action, has been scarcely addressed, which is due in large part to the lack of appropriate tools and reagents. As evidenced by the WAP-TGF-β223-225 versus the MMTV-TGF-β223-225 transgenic phenotypes, spatial and temporal patterns of LTGF-β activation are key elements for to understanding the specific consequences of TGF-β activity during mammary gland development. Likewise, similar studies of the relative activity of the different TGF-β isoforms in the mammary gland may help shed light on the need for this apparent redundancy. Finally, studies that address mechanisms of activation may then contribute to strategies for manipulation of TGF-β in situ and lead to a better understanding of how its dysregulation contributes to carcinogenesis [58].
Acknowledgments
Acknowledgements
Work by the authors and cited in the present review was supported by the US Army Medical Research and Materiel Command under DAMD-17-96-1-6716, the California Breast Cancer Research Program and the Office of Health and Environmental Research, Health Effects Research Division, of the US Department of Energy Contract No DE-AC-03-76SF00098.
References
- Daniel CW, Robinson SD. Regulation of mammary growth and function by TGF-β. . Mol Reprod Dev. 1992;32:145–151. doi: 10.1002/mrd.1080320210. [DOI] [PubMed] [Google Scholar]
- Knabbe C, Lippman ME, Wakefield LM, et al. Evidence that transforming growth factor-β is a hormonally regulated negative growth factor in human breast cancer cells. Cell. 1987;48:417–428. doi: 10.1016/0092-8674(87)90193-0. [DOI] [PubMed] [Google Scholar]
- Wakefield l, Colletta AA, McCune BK, Sporn MB. Roles for transforming growth factors-β in the genesis, prevention and treatment of breast cancer. Genes, Oncogens, and Hormones: Advances in Cellular and Molecular Biology of Breast Cancer. Edited by Dickson RB, Lipman MF. Boston: Kluwer Academic Publishers, 1991. pp. 97–136.
- Silberstein GB, Daniel CW. Reversible inhibition of mammary gland growth by transforming growth factor-β. . Science. 1987;237:291–293. doi: 10.1126/science.3474783. [DOI] [PubMed] [Google Scholar]
- Daniel CW, Silberstein GB, Van Horn K, Strickland P, Robinson S. TGF-β1-induced inhibition of mouse mammary ductal growth: developmental specificity and characterization. . Dev Biol. 1989;135:20–30. doi: 10.1016/0012-1606(89)90154-1. [DOI] [PubMed] [Google Scholar]
- Daniel CW, Silberstein GB. Postnatal development of the rodent mammary gland. . The Mammary Gland: Development, Regulation and Function. Edited by Neville MC, Daniel CW. New York: Plenum Press Publishing Corp. 1987:3–36. [Google Scholar]
- Lawrence DA, Pircher R, Jullien P. Conversion of a high molecular weight latent beta-TGF from chicken embryo fibroblasts into a low molecular weight active beta-TGF under acidic conditions. . Biochem Biophys Res Commun. 1985;133:1026–1034. doi: 10.1016/0006-291x(85)91239-2. [DOI] [PubMed] [Google Scholar]
- Kim S-J, Park K, Koeller D, et al. Post-transcriptional regulation of the human transforming growth factor-β1 gene. . J Biol Chem. 1992;267:13702–13707. [PubMed] [Google Scholar]
- Lawrence DA. Identification and activation of latent transforming growth factor β. . Methods Enzymol. 1991;198:327–336. doi: 10.1016/0076-6879(91)98033-3. [DOI] [PubMed] [Google Scholar]
- Flaumenhaft R, Rifkin DB. The extracellular regulation of growth factor action. Mol Biol Cell. 1992;3:1057–1065. doi: 10.1091/mbc.3.10.1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierce DFJ, Johnson MD, Matsui Y, et al. Inhibition of mammary duct development but not alveolar outgrowth during pregnancy in transgenic mice expressing active TGF-β1. . Genes Dev. 1993;7:2308–2317. doi: 10.1101/gad.7.12a.2308. [DOI] [PubMed] [Google Scholar]
- Tang B, Bottinger EP, Jakowlew SB, et al. Transforming growth factor-beta1 is a new form of tumor suppressor with true haploid insufficiency. . Nature Med. 1998;4:802–807. doi: 10.1038/nm0798-802. [DOI] [PubMed] [Google Scholar]
- Gomm JJ, Smithe J, Ryall GK, et al. Localization of basic fibroblast growth factor and transforming growth factor β1 in the human mammary gland. . Cancer Res. 1991;51:4685–4692. [PubMed] [Google Scholar]
- Lu Y-J, Osin P, Lakhani SR, Palma SD, Gusterson BA, Shipley JM. Comparative genomic hybridization analysis of lobular carcinoma in situ and a typical lobular hyperplasia and potential roles for gains and losses of genetic material in breast neoplasia. . Cancer Res. 1998;58:4721–4727. [PubMed] [Google Scholar]
- Mizukami Y, Nonomura A, Yamada T, et al. Immunohistochemical demonstration of growth factors, TGF-alpha, TGF-beta, IGF-I and neu oncogene product in benign and malignant human breast tissues. . Anticancer Res. 1990;10:1115–1126. [PubMed] [Google Scholar]
- Stampfer MR, Yaswen P, Alhadeff M, Hosoda J. TGF-β induction of extracellular matrix associated proteins in normal and transformed human mammary epithelial cells in culture is independent of growth effects. . J Cell Physiol. 1993;155:210–221. doi: 10.1002/jcp.1041550127. [DOI] [PubMed] [Google Scholar]
- Baillie R, Coombes RC, Smith J. Multiple forms of TGF-beta 1 in breast tissues: a biologically active form of the small latent complex of TGF-beta 1. . Eur J Cancer. 1996;32A:1566–1573. doi: 10.1016/0959-8049(96)00115-3. [DOI] [PubMed] [Google Scholar]
- Dong-Le Bourhis X, Berthois Y, Millot G, et al. Effect of stromal and epithelial cells derived from normal and tumorous breast tissue on the proliferation of human breast cancer cell lines in co-culture. Int J Cancer. 1997;71:42–48. doi: 10.1002/(SICI)1097-0215(19970328)71:1<42::AID-IJC9>3.3.CO;2-X. [DOI] [PubMed] [Google Scholar]
- Sieuwerts AM, Klijn JG, Henzen-Logmand SC, et al. Urokinase-type-plasminogen-activator (uPA) production by human breast (myo) fibroblasts in vitro: influence of transforming growth factor-beta(1) [TGF beta(1)] compared with factor(s) released by human epithelial-carcinoma cells. . Int J Cancer. 1998;76:829–835. doi: 10.1002/(SICI)1097-0215(19980610)76:6<829::AID-IJC11>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- Chakravarthy D, Green AR, Green VL, Kerin MJ, Speirs V. Expression and secretion of TGF-beta isoforms and expression of TGF-beta-receptors I, II and III in normal and neoplastic human breast. . Int J Oncol. 1999;15:187–194. doi: 10.3892/ijo.15.1.187. [DOI] [PubMed] [Google Scholar]
- Robinson SD, Silberstein GB, Roberts AB, Flanders KC, Daniel CD. Regulated expression and growth inhibitory effects of transforming growth factor-β isoforms in mouse mamary gland development. . Development. 1991;113:867–878. doi: 10.1242/dev.113.3.867. [DOI] [PubMed] [Google Scholar]
- Silberstein GB, Flanders KC, Roberts AB, Daniel CW. Regulation of mammary morphogenesis: evidence for extracellular matrix-mediated inhibition of ductal budding by transforming growth factor-β1. . Dev Biol. 1992;152:354–362. doi: 10.1016/0012-1606(92)90142-4. [DOI] [PubMed] [Google Scholar]
- Barcellos-Hoff MH. Latency and activation in the regulation of TGF-β. . J Mam Gland Biol Neoplasia. 1996;3:353–363. [PubMed] [Google Scholar]
- Barcellos-Hoff MH, Derynck R, Tsang ML-S, Weatherbee JA. Transforming growth factor-β activation in irradiated murine mammary gland. . J Clin Invest. 1994;93:892–899. doi: 10.1172/JCI117045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barcellos-Hoff MH, Ehrhart EJ, Kalia M, et al. Immunohistochemical detection of active TGF-β in situ using engineered tissue. . Am J Pathol. 1995;147:1228–1237. [PMC free article] [PubMed] [Google Scholar]
- Barcellos-Hoff MH. Radiation-induced transforming growth factor β and subsequent extracellular matrix reorganization in murine mammary gland. . Cancer Res. 1993;53:3880–3886. [PubMed] [Google Scholar]
- Ehrhart EJ, Carroll A, Segarini P, Tsang ML-S, Barcellos-Hoff MH. Latent transforming growth factor-β activation in situ: quantitative and functional evidence following low dose irradiation. . FASEB J. 1997;11:991–1002. doi: 10.1096/fasebj.11.12.9337152. [DOI] [PubMed] [Google Scholar]
- Pertovaara L, Sistonen L, Bos TJ, et al. Enhanced jun gene expression is an early genomic response to transforming growth factor beta stimulation. . Mol Cell Biol. 1989;9:1255–1262. doi: 10.1128/mcb.9.3.1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahimi N, Tremblay E, McAdam L, Roberts A, Elliott B. Autocrine secretion of TGF-beta 1 and TGF-beta 2 by pre-adipocytes and adipocytes: a potent negative regulator of adipocyte differentiation and proliferation of mammary carcinoma cells. . In Vitro Cell Dev Biol Anim. 1998;34:412–420. doi: 10.1007/s11626-998-0023-z. [DOI] [PubMed] [Google Scholar]
- Ellis I, Banyard J, Schor SL. Differential response of fetal and adult fibroblasts to cytokines: cell migration and hyaluronan synthesis. . Development. 1997;124:1593–1600. doi: 10.1242/dev.124.8.1593. [DOI] [PubMed] [Google Scholar]
- McCaffrey TA, Falcone DJ. Evidence for an age-related dysfunction in the antiproliferative response to transforming growth factor-β in vascular smooth muscle cells. . Mol Biol Cell. 1993:315–322. doi: 10.1091/mbc.4.3.315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi K, Suzuki K, Ono T. Loss of growth inhibitory activity of TGF-β toward normal human mammary epithelial cells growth within collagen gel matrix. . Biochem Biophys Res Commun. 1990;173:1239–1247. doi: 10.1016/s0006-291x(05)80919-2. [DOI] [PubMed] [Google Scholar]
- Martikainen P, Kyprianou N, Isaacs JT. Effect of transforming growth factor-β1 on proliferation and death of rat prostatic cells. . Endocrinology. 1990;127:2963–2968. doi: 10.1210/endo-127-6-2963. [DOI] [PubMed] [Google Scholar]
- Rotello RJ, Lieberman RC, Purchio AF, Gerschenson LE. Coordinated regulation of apoptosis and cell proliferation by transforming growth factor β1 in cultured uterine epithelial cells. . Proc Natl Acad Sci USA. 1991;88:3412–3415. doi: 10.1073/pnas.88.8.3412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strange R, Li F, Saurer S, Burkhardt A, Friis RR. Apoptotic cell death and tissue remodeling during mouse mammary gland involution. . Development (Camb) 1992;115:49–58. doi: 10.1242/dev.115.1.49. [DOI] [PubMed] [Google Scholar]
- Ayoub IA, Yang TJ. The regulatory role of transforming growth factor-beta in activation of milk mononuclear cells. . Am J Reprod Immunol. 1997;38:121–128. doi: 10.1111/j.1600-0897.1997.tb00286.x. [DOI] [PubMed] [Google Scholar]
- Newton LK, Yung WKA, Pettigrew LC, Steck PA. Growth regulatory activities of endothelial extracellular matrix mediation by transforming growth factor-β. . Exp Cell Res. 1990;190:127–132. doi: 10.1016/0014-4827(90)90153-2. [DOI] [PubMed] [Google Scholar]
- Ferguson JE, Schor AM, Howell A, Feruson MWJ. Changes in the extracellular matrix of the normal human breast during the men-strual cycle. . Cell Tissue Res. 1992;268:167–177. doi: 10.1007/BF00338066. [DOI] [PubMed] [Google Scholar]
- Silberstein GB, Strickland P, Coleman S, Daniel CW. Epithelium-dependent extracellular matrix synthesis in transforming growth factor-β1-growth-inhibited mouse mammary gland. . J Cell Biol. 1990;110:2209–2219. doi: 10.1083/jcb.110.6.2209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Streuli CH, Schmidhauser C, Kobrin M, Bissell MJ, Derynck R. Extra-cellular matrix regulates expression of the TGF-β1 gene. . J Cell Biol. 1993;120:253–260. doi: 10.1083/jcb.120.1.253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie J, Haslam SZ. Extracellular matrix regulates ovarian hormone-dependent proliferation of mouse mammary epithelial cells. Endocrinology. 1997;138:2466–2473. doi: 10.1210/endo.138.6.5211. [DOI] [PubMed] [Google Scholar]
- Butta A, MacLennan K, Flanders KC, et al. Induction of transforming growth factor β1 in human breast cancer in vivo following tamoxifen treatment. . Cancer Res. 1992;52:4261–4264. [PubMed] [Google Scholar]
- Bruner KL, Rodgers WH, Gold LI, et al. Transforming growth factor β mediates the progesterone suppression of an epithelial metalloproteinase by adjacent stroma in the human endometrium. . Proc Natl Acad Sci USA. 1995;92:7362–7366. doi: 10.1073/pnas.92.16.7362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson SD, Roberts AB, Daniel CW. TGFβ suppresses casein synthesis in mouse mammary explants and may play a role in controlling milk levels. . J Cell Biol. 1993;120:245–251. doi: 10.1083/jcb.120.1.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergstraesser L, Sherer S, Panos R, Weitzman S. Stimulation and inhibition of human mammary epithelial cell duct morphogenesis in vitro. . Proc Assoc Am Phys. 1996;108:140–154. [PubMed] [Google Scholar]
- Soriano JV, Orci L, Montesano R. TGF-β1 induces morphogenesis of branching cords by cloned mammary epithelial cells at subpicomolar concentrations. . Biochem Biophys Res Commun. 1996;220:879–885. doi: 10.1006/bbrc.1996.0499. [DOI] [PubMed] [Google Scholar]
- Miettinen PJ, Ebner R, Lopez AR, Derynck R. TGF-β induced transdifferentiation of mammary epithelial cells to mesenchymal cells: involvement of type I receptors. . J Cell Biol. 1994;127:2021–2036. doi: 10.1083/jcb.127.6.2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desmouliere A, Geinoz A, Gabbiani F, Gabbiani G. Transforming growth factor-β1 induces α-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. . J Cell Biol. 1993;122:103–111. doi: 10.1083/jcb.122.1.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniel CW, Robinson S, Silberstein GB. The role of TGF-β in patterning and growth of the mammary ductal tree. . J Mam Gland Biol Neoplasia. 1996;1:331–341. doi: 10.1007/BF02017389. [DOI] [PubMed] [Google Scholar]
- Brunner AM, Marquardt H, Malacko AR, Lioubin MN, Purchio AF. Site-directed mutagenesis of cysteine residues in the pro region of the transforming growth factor β1 precursor. . J Biol Chem. 1989;264:13660–13664. [PubMed] [Google Scholar]
- Smith GH. TGF-β and functional differentiation. . J Mam Gland Biol Neoplasia. 1996;1:343–352. doi: 10.1007/BF02017390. [DOI] [PubMed] [Google Scholar]
- Jhappan C, Geiser AG, Kordon EC, et al. Targeting expression of a transforming growth factor β1 transgene to the pregnant mammary gland inhibits alveolar development and lactation. EMBO J. 1993;12:1835–1845. doi: 10.1002/j.1460-2075.1993.tb05832.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kordon EC, McKnight RA, Jhappan C, et al. Ectopic TGF beta 1 expression in the secretory mammary epithelium induces early senescence of the epithelial stem cell population. . Dev Biol. 1995;168:47–61. doi: 10.1006/dbio.1995.1060. [DOI] [PubMed] [Google Scholar]
- Kulkarni AB, Huh CG, Becker D, et al. Transforming growth factor beta 1 null mutation in mice causes excessive inflammatory response and early death. . Proc Natl Acad Sci USA. 1993;90:770–774. doi: 10.1073/pnas.90.2.770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letterio JJ, Geiser AG, Kulkarni AB, et al. Maternal rescue of transforming growth factor-β1 null mice. . Science. 1994;264:1936–1938. doi: 10.1126/science.8009224. [DOI] [PubMed] [Google Scholar]
- Gorska AE, Joseph H, Derynck R, Moses HL, Serra R. Dominant-negative interference of the transforming growth factor-beta type II receptor in mammary gland epithelium results in alveolar hyperplasia and differentiation in virgin mice. . Cell Growth Differ. 1998;9:229–238. [PubMed] [Google Scholar]
- Joseph H, Gorska AE, Sohn P, Moses HL, Serra R. Overexpression of a kinase-deficient transforming growth factor-β type II receptor in mouse mammary stroma results in increased epithelial branching. . Mol Biol Cell. 1999;10:1221–1234. doi: 10.1091/mbc.10.4.1221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiss M, Barcellos-Hoff MH. Transforming growth factor-β in breast cancer: a working hypothesis. . Br Cancer Res Treat. 1997;45:81–95. doi: 10.1023/a:1005865812918. [DOI] [PubMed] [Google Scholar]