Distinct Dynamin-dependent and -independent Mechanisms Target Structurally Homologous Dopamine Receptors to Different Endocytic Membranes (original) (raw)
Abstract
D1 and D2 dopamine receptors are structurally homologous G protein–coupled receptors that serve distinct physiological functions both in neurons and nonneural cell types. We have observed that these receptors are selectively endocytosed in HEK293 cells by distinct dynamin-dependent and -independent mechanisms. Although these endocytic mechanisms operate with similarly rapid kinetics, they differ in their regulation by agonist and deliver D1 and D2 receptors specifically to different primary endocytic vesicles. After this segregation into different endocytic membranes, both D1 and D2 receptors recycle to the plasma membrane. Similar results are observed in Neuro2A neuroblastoma cells coexpressing both receptors at high levels. These findings establish that “classical” dynamin-dependent and “alternative” dynamin-independent endocytic mechanisms differ in their physiological regulation, sort structurally homologous signaling receptors in the plasma membrane, and mediate distinct early endocytic pathways leading to recycling endosomes. Our results also refute the previous hypothesis that dynamin-independent endocytosis targets G protein–coupled receptors selectively to lysosomes, and they suggest a new role of endocytic sorting mechanisms in physically segregating structurally homologous signaling receptors at the cell surface.
Keywords: receptor, endocytosis, dopamine, cell membrane, cell physiology
Endocytosis of cell surface receptors has a well-established role in transporting ligands required for cellular nutrition and transcytosis (Trowbridge et al., 1993; Apodaca et al., 1994; Mostov and Cardone, 1995). Many receptors that mediate ligand-dependent signal transduction also undergo endocytosis. The endocytosis of signaling receptors serves a variety of important physiological functions in the rapid and long-term regulation of signal transduction (Lohse, 1993; Seaman et al., 1996; Lefkowitz et al., 1998), as well as in determining the physiological specificity of receptor-mediated signaling among distinct downstream effectors (Vieira et al., 1996; Daaka et al., 1998). Although these functions are clearly distinct from nutrient uptake, and different protein interactions can promote the association of signaling receptors with endocytic membranes, the endocytic pathways themselves that mediate the endocytic membrane trafficking of signaling receptors and nutritive receptors are remarkably similar (Bohm et al., 1996; Zhang et al., 1996).
Several receptor-tyrosine kinases and G protein–coupled receptors are endocytosed by a dynamin-dependent mechanism involving clathrin-coated pits, and these receptors follow an endocytic pathway similar to that mediating the constitutive endocytosis of LDL and transferrin receptors (Carpentier et al., 1982; Hopkins et al., 1985; von Zastrow and Kobilka, 1992; Goodman et al., 1996). However, dynamin-dependent endocytosis of signaling receptors tends to be tightly regulated by agonists and exhibits a high degree of biochemical specificity. For example, structurally homologous subtypes of G protein–coupled receptor differ significantly in their ability to enter clathrin-coated pits after activation, even when coexpressed together in the same cells at high levels. Moreover, the subtype specificity of G protein–coupled receptor endocytosis is a distinct functional property of individual receptors that does not correlate with the biochemical specificity of G protein activation (Nantel et al., 1993; von Zastrow et al., 1993; Mostafapour et al., 1996; Daunt et al., 1997).
In addition to agonist-induced endocytosis by clathrin-coated pits, there is increasing evidence that certain signaling receptors can be endocytosed by alternate, clathrin-independent endocytic mechanism(s). For example, G protein–coupled AT1A angiotensin receptors and M2 muscarinic receptors are endocytosed by a dynamin-independent mechanism (Zhang et al., 1996; Pals-Rylaarsdam et al., 1997). Furthermore, while G protein–coupled beta-2 adrenergic receptors are endocytosed by clathrin-coated pits in some cell types, previous studies suggest that these receptors may be endocytosed by a clathrin-independent mechanism in other cells (Raposo et al., 1989; Dupree et al., 1993). These observations raise fundamental questions about the role of distinct endocytic mechanisms in the cell biology of signaling receptors. In particular, it is not known whether structurally homologous receptors are endocytosed specifically by distinct mechanisms in the same cells. Furthermore, it is not known whether the endocytosis of signaling receptors by distinct mechanisms at the plasma membrane targets receptors to different endocytic pathways, or whether the endocytic vesicles formed by these endocytic mechanisms rapidly fuse together to deliver endocytosed receptors to the same early endosomes. The latter is true for certain nutrients endocytosed constitutively by clathrin-independent endocytosis, raising the question of what physiological role is served by endocytosis of receptors by distinct mechanisms (van Deurs et al., 1989; Hansen et al., 1993). In contrast, previous studies of G protein–coupled receptors suggest that distinct endocytic mechanisms may target receptors to different pathways directly from the plasma membrane (Valiquette et al., 1990; Hausdorff et al., 1991; Zhang et al., 1996), suggesting a primary role of these mechanisms in endocytic sorting. However, the role of distinct mechanisms in sorting signaling receptors is not clear because the beta-2 adrenergic receptor may be endocytosed by more than one mechanism in the same cell type (Valiquette et al., 1990; Hausdorff et al., 1991). Indeed, recent studies demonstrate that dynamin-dependent endocytosis can deliver the beta-2 adrenergic receptor to both recycling and degradative pathways, further raising the question of the physiological function served by distinct endocytic mechanisms (Gagnon et al., 1998).
We have examined the endocytic membrane trafficking of D1 and D2 dopamine receptors, two structurally homologous subtypes of G protein–coupled receptor that are closely related to adrenergic receptors. These receptors are of particular interest because they are prototypic postsynaptic and presynaptic receptors, respectively, in neurons and they also serve distinct physiological functions in nonneural cells (Jaber et al., 1996; Missale et al., 1998). Furthermore, previous studies suggest that differences in the membrane localization of these receptors may distinguish their functional regulation in heterologous cell types (Trogadis et al., 1995; Ng et al., 1997). Thus, we anticipated that examination of these receptors might provide fundamental new insight into the physiological diversity of membrane trafficking properties among individual, structurally homologous signaling receptors that serve different biological functions.
Indeed, our studies demonstrate that D1 and D2 dopamine receptors differ greatly in their endocytic membrane trafficking, both in neural and nonneural cell types. Specifically, we demonstrate that D1 and D2 receptors, even when overexpressed at high levels in transfected cells, are endocytosed specifically by distinct dynamin-dependent and -independent mechanisms. Furthermore, we demonstrate that these endocytic mechanisms differ in their physiological regulation and target structurally homologous receptors to different early endocytic pathways. Nevertheless, both D1 and D2 receptors are apparently able to recycle efficiently to the plasma membrane. These observations demonstrate that structurally homologous signaling receptors are efficiently sorted by distinct endocytic mechanisms, and they suggest that a primary function of these mechanisms is to physically segregate structurally homologous signaling receptors at or near the cell surface.
Materials and Methods
Construction of Epitope-tagged D1 and D2 Dopamine Receptors
Human D1 and D2 dopamine receptors (cDNAs provided by Dr. David Grandy, Vollum Institute, Portland, OR) were epitope-tagged in the amino-terminal extracellular domain with either a FLAG or HA epitope, as described previously for the epitope tagging of adrenergic and opiate receptors (Guan et al., 1992; Keith et al., 1996). The coding sequence of the human D1 receptor (Zhou et al., 1990) was digested with NcoI and SmaI and inserted into the NcoI and PvuII sites of a shuttle vector containing a sequence encoding the appropriate epitope tag following a favorable Kozak sequence and a cleavable signal sequence (Guan et al., 1992). This strategy substituted the first nine residues in the amino-terminal extracellular domain of the D1 receptor with the corresponding epitope tag. The coding sequence of the long isoform of the human D2 receptor (Grandy et al., 1989) was digested at the 5′-end using BamHI, blunt-ended with the Klenow fragment of DNA polymerase, and the 3′-end was then digested with XhoI. This fragment was inserted into the same shuttle vector as used above, resulting in the addition of the epitope tag sequence directly to the amino-terminal extracellular domain of the wild-type protein. Epitope-tagged receptor cDNAs were then subcloned into pcDNA3 (Invitrogen). All mutated sequences were confirmed by dideoxy sequencing of the completed constructs (Biomolecular Resource Center, UCSF).
Cell Culture and Transfection
Human embryonic kidney (HEK)1 293 fibroblastic cells and mouse Neuro2A neuroblastoma cells (American Type Culture Collection) were maintained in Dulbecco's minimal essential medium (DME H-21) supplemented with 10% fetal bovine calf serum and 100 U/ml pen/strep (UCSF Cell Culture Facility). Cell monolayers were transfected with receptor cDNAs cloned into pcDNA3 by calcium phosphate coprecipitation (Okayama and Berg, 1983). Clones of stably transfected cells were selected and cultured in 200 μg/ml G418 (geneticin; Life Technologies, Inc.), and screened for receptor expression using fluorescence microscopy and signaling assays (see below). To examine the effect of mutant dynamin on endocytosis, cDNA encoding HA-tagged wild-type or K44E dominant negative mutant dynamin I (provided by Dr. Richard Vallee, Worcester Foundation for Biomedical Research, Shrewsbury, MA) was cloned into pcDNA3 and expressed by transient transfection in cell clones expressing either FLAG-tagged D1 or D2 receptors. Experiments using transiently transfected cells were performed 48–72 h after transfection.
Adenylyl Cyclase Assay
Stably transfected HEK293 cells expressing epitope-tagged dopamine receptors were preincubated in multiwell plates with 1 mM isobutyl methylxanthine (Sigma Chemical Co.) for 30 min at 37°C in DME H-21. Cells expressing epitope-tagged D1 receptors were then incubated for an additional 30 min after the addition of various concentrations of dopamine (Research Biochemicals) freshly dissolved in water, or after the addition of an equal volume of water for control (unstimulated) incubations. Cells expressing epitope-tagged D2 receptors were preincubated in the same manner with isobutyl methylxanthine (1 mM), and then forskolin (10 μM) was added followed after 5 min by dopamine or an equal volume of water for an additional 30 min. After incubations carried out at 37°C, cell extracts were prepared and cAMP present in lysates was measured as described (Keith et al., 1996) using a radioimmunoassay kit (Diagnostic Products Corp.). Data are presented as mean dopamine-induced stimulation (D1 receptors) or inhibition (D2 receptors) of adenylyl cyclase activity ±SD, determined from six independent assays for each condition.
Confirmation of Functional Integrity of Epitope-tagged D1 and D2 Dopamine Receptors
The functional integrity of epitope-tagged D1 and D2 receptors was confirmed by ligand-dependent regulation of adenylyl cyclase (see below) in stably transfected HEK293 cells, a process which requires that expressed receptors are capable of binding agonists and coupling to the appropriate heterotrimeric G proteins. Receptor signaling was measured by assaying dopamine-dependent regulation of adenylyl cyclase activity in intact cells. Dopamine caused little change of adenylyl cyclase activity in untransfected HEK293 cells, indicating that these cells express a negligible background level of endogenous dopamine receptors. In cells transfected with epitope-tagged D1 receptors, dopamine caused a large stimulation of adenylyl cyclase activity (15 ± 1.7-fold stimulation for the FLAG-tagged D1 receptor and a 21 ± 2.0-fold stimulation for the HA-tagged receptor with 10 μM dopamine, n = 6), indicating that epitope-tagged D1 receptors mediate functional ligand-dependent signaling via Gs. Epitope-tagged D2 receptors mediated significant dopamine-dependent inhibition of adenylyl cyclase activity (5.0 ± 2.0-fold inhibition with 10 μM dopamine, n = 6), confirming that these receptors mediate functional ligand-dependent signaling via Gi.
Immunocytochemical Staining and Fluorescence Microscopy
Cells grown on glass coverslips were preincubated with M1 mAb (3 μg/ml; Kodak IBI), which specifically recognizes the FLAG epitope, or HA.11 mAb (3 μg/ml; Berkeley Antibody Co.), which specifically recognizes the HA epitope. After preincubation for 30 min to allow antibodies to bind to epitope-tagged receptors exposed at the plasma membrane, experiments were started by incubating cells with the indicated concentrations of dopamine (the physiological agonist of both D1 and D2 receptors), SCH23390 (a potent D1 receptor antagonist), or haloperidol (a potent D2 receptor antagonist) for the indicated times (all dopaminergic compounds were purchased from Research Biochemicals). Experiments were stopped by fixing cells for 15 min with 4% formaldehyde in PBS. Specimens were then permeabilized with 0.1% Triton X-100 (Sigma Chemical Co.), and antibody-labeled receptors were visualized by incubating permeabilized specimens in the presence of FITC-conjugated goat anti–mouse IgG or Cy3-conjugated donkey anti–mouse IgG (1:250 dilution of stock prepared according to manufacturer's recommendation; Jackson ImmunoResearch). For dual localization of mutant dynamin in cells expressing FLAG-tagged dopamine receptors, fixed and permeabilized specimens were incubated with mouse monoclonal anti-HA antibody (1:1,000 dilution; Berkeley Antibody Co.) and 5 μg/ml subtype-specific rabbit anti– mouse IgG2b antibody (Zymed), and washed. Then, receptors (labeled with M1 monoclonal and rabbit anti–mouse IgG2b) and dynamin (labeled with mouse anti-HA IgG1) were visualized specifically by incubating specimens with Texas red donkey anti–rabbit (1:250 dilution; Jackson ImmunoResearch) and anti–mouse IgG1 subtype-specific FITC (1:1,000 dilution; Boehringer Mannheim). Colocalization studies of FLAG-tagged D2 receptors with HA-tagged D1 receptors were performed in a similar manner. Epifluorescence microscopy was performed using a Nikon 60X NA1.4 objective, and confocal fluorescence microscopy was carried out using a BioRad MRC 1000 and a Zeiss 100X NA1.3 objective. Minimal bleedthrough in double-labeling experiments was verified by imaging single-labeled specimens under the same conditions.
Biochemical Analysis of Receptor Endocytosis and Degradation Using Cleavable and Noncleavable Biotin
Stably transfected HEK293 cells expressing either the FLAG-tagged D1 or D2 receptor were grown to confluence in 6-cm dishes, then chilled and washed three times with ice-cold PBS. For internalization assays, the cell surface was biotinylated with 30 mg/ml cleavable sulfo-NHS-_S_-_S_-biotin (Pierce Chemical Co.) for 30 min either at 4°C or room temperature, as specified in the figure legends. For degradation assays, the cell surface was biotinylated with 30 mg/ml noncleavable sulfo-NHS-biotin in PBS (Pierce Chemical Co.) for 30 min at room temperature. Unreacted biotin was quenched and removed by three successive washes of biotinylated cells in TBS supplemented with 10 mM glycine. Cells were then incubated for the indicated times in regular media (DME + 10% FBS) prewarmed to 37°C and supplemented with the indicated ligands. For internalization assays, the physiological agonist dopamine (10 μM) was used to activate both D1 and D2 receptors. Degradation assays, which required prolonged periods of incubation that caused significant oxidative degradation of dopamine, were conducted using the relatively stable D1-specific agonist 6-chloro-PB (SKF-81297) or the D2-specific agonist quinpirole at a final concentration of 10 μM. Assays were stopped by chilling cells immediately on ice and then washing three times with ice-cold PBS. For cleavage of surface-localized biotinylated receptors in the internalization assays, cells were then incubated at 4°C with two changes of glutathione strip buffer (50 mM glutathione, 75 mM NaCl, 75 mM NaOH, 10% FBS in H2O) for 20 min each (Cao et al., 1998). Excess glutathione was then quenched for 30 min at 4°C in iodoacetamide buffer (50 mM iodoacetamide, 1% BSA in PBS, pH 7.4). For both internalization and degradation assays, cells were then extracted with 0.5% (vol/vol) Triton X-100, 10 mM Tris-HCl, pH 7.5, 120 mM NaCl, 25 mM KCl, 1 μg/ml leupeptin, 1 μg/ml pepstatin, 2 μg/ml aprotinin, 2 μg/ml pefablock, and 1 mg/ml iodoacetamide. Extracts were then clarified by two rounds of centrifugation at 4°C (14,000 g) for 15 min, and receptors were purified from the extracts by immunoprecipitation.
D1 and D2 receptors were immunoprecipitated using 5 μg/ml anti-FLAG M2 mAb (Kodak) followed by 7 μg/ml rabbit anti–mouse IgG (Jackson ImmunoResearch) and 25 μl of protein A–Sepharose beads in PBS (Pharmacia Biotech). Immunoprecipitates were washed sequentially with HSB (0.1% SDS, 0.5% Triton X-100, 20 mM Tris-HCl, pH 7.5, 120 mM NaCl, 25 mM KCl), two washes with HSB layered with a 1 M sucrose cushion, one wash with 1 M NaCl in HSB, and one wash in low-salt wash buffer (0.5% Triton X-100, 10 mM Tris-HCl, pH 7.35, and 2 mM EDTA). Washed beads were extracted with SDS sample buffer and eluted proteins were resolved by SDS-PAGE under nonreducing conditions. Resolved proteins were transferred to nitrocellulose membranes (Micron Separations, Inc.) and blocked for 30 min in Blotto (5% dry milk, 0.1% Tween 20 in TBS). Biotinylated proteins were then complexed with horseradish peroxidase using the VectaStain ABC detection system (Vector Laboratories), then detected by enzyme-linked chemiluminescence (Amersham) and exposed to BioMax film (Eastman Kodak). Quantitation was performed by transmission densitometry of films exposed in the linear range, using a CCD video camera (Cohu Electronics) interfaced with a Macintosh computer running NIH Image software.
Results
D1 and D2 Dopamine Receptors Differ in Their Endocytic Membrane Trafficking in Neuroblastoma Cells
Receptors present in the plasma membrane of transfected Neuro2A neuroblastoma cells were epitope tagged on their extracellular domain, allowing these receptors to be labeled specifically in intact cells by adding mAb to the culture medium. The redistribution of antibody-labeled receptors was then visualized by fluorescence microscopy to examine specifically the endocytic trafficking of receptors from the plasma membrane. This approach has been used successfully to study the endocytosis and recycling of several G protein–coupled receptors without significantly perturbing their membrane trafficking properties (von Zastrow and Kobilka, 1992; von Zastrow et al., 1993; Keith et al., 1996; Daunt et al., 1997). Endocytosis of mAb was not observed in untransfected cells or in cells expressing receptors that do not possess the appropriate epitope tag (data not shown), indicating that endocytosis of antibody is specifically mediated by internalization from the plasma membrane.
D1 and D2 receptors were coexpressed in the same cells and selectively labeled at the cell surface by anti-HA (HA.11) and anti-FLAG (M1) mAbs, respectively. In cells incubated in the absence of dopamine, D1 receptors remained primarily in the plasma membrane (Fig. 1 A, arrowheads) and were observed only rarely in intracellular membranes. In contrast, D2 receptors endocytosed within minutes into numerous endocytic vesicles located throughout the cytoplasm (Fig. 1 B, arrows indicate examples of these structures). This difference in the membrane localization of D1 and D2 receptors is emphasized in the merged color image (Fig. 1 C). In the presence of the physiological agonist dopamine, both D1 and D2 receptors were observed in numerous endocytic vesicles (Fig. 1, D and E, respectively; Fig. 1 F shows the merged color image). Significantly, D1 and D2 receptors were differentially regulated in cells coexpressing both receptors, confirming that these observations reflect intrinsic differences in membrane trafficking properties of these closely homologous receptor proteins.
Figure 1.

Localization of D1 and D2 dopamine receptors in neuroblastoma cells. HA-tagged D1 receptors (arrowheads) and FLAG-tagged D2 receptors (arrows) present in the plasma membrane of transfected N2A neuroblastoma cells were specifically labeled at 37°C with HA.11 and M1 mAbs, respectively, as described in Materials and Methods. The localization of antibody-labeled receptors was then visualized by fluorescence microscopy after incubation of cells for 30 min in the absence (A–C) or presence (D–F) of 10 μM dopamine.
Subtype-specific Differences in Dopamine Receptor Trafficking Are Conserved in Nonneural Cell Types
To determine whether the differences in the endocytic membrane trafficking of D1 and D2 receptors are unique to neural cells, or if these differences in receptor internalization may be conserved in a wider variety of cell types, we examined the endocytic trafficking of D1 and D2 receptors in HEK293 fibroblast cells. These experiments were conducted in stably transfected cell clones expressing either D1 or D2 receptors tagged with the FLAG epitope, and cell clones were selected that expressed similar numbers of receptors in the plasma membrane (determined by fluorescence flow cytometry after labeling of surface receptors using FITC-conjugated antibody, data not shown).
As observed in neuroblastoma cells, FLAG-tagged D1 receptors labeled with mAb remained predominantly in the plasma membrane of cells incubated in the absence of agonist and did not internalize to a significant extent after 1 h (Fig. 2 A). Receptor activation caused rapid redistribution of antibody-labeled receptors from the plasma membrane to intracellular vesicles, which was readily detectable by fluorescence microscopy within several minutes after the addition of dopamine (Fig. 2 B). We confirmed this observation using a biochemical assay of receptor internalization based upon inaccessibility of surface-biotinylated receptors to membrane-impermeant reducing agent after endocytosis (Cao et al., 1998). D1 receptors biotinylated in the plasma membrane became inaccessible to extracellular glutathione within minutes after addition of dopamine, consistent with rapid net internalization of receptors. In contrast, in the absence of dopamine, very little internalized (glutathione-resistant) receptor was detected even after 60 min (Fig. 2 C). The D1 receptor antagonist SCH23390 (1 μM) blocked the ability of dopamine to induce the rapid internalization of D1 receptors, whereas SCH23390 had no effect by itself on the localization of receptors in the plasma membrane (Fig. 2 C, inset). Quantitation of these data by densitometric scanning confirmed the rapidity of D1 receptor internalization (t 1/2 <5 min) and revealed that, at steady state, dopamine induced ∼70% net internalization of the total surface pool of D1 receptors. In contrast, after incubation for 60 min in the absence of dopamine, <5% net internalization was observed (Fig. 2 D). These results suggest that endocytosis of D1 receptors in HEK293 cells, as in neuroblastoma cells, occurs rapidly and is mediated by an agonist-dependent mechanism.
Figure 2.

Localization and endocytosis of D1 receptors in HEK293 cells. The localization of FLAG-tagged D1 receptors expressed in stably transfected HEK293 cells was visualized by fluorescence microscopy in cells incubated for 30 min in the absence (A) or presence (B) of 10 μM dopamine. (C) Agonist- induced internalization of D1 receptors was determined biochemically using cell surface biotinylation assay, as described in Materials and Methods. The total amount of D1 receptor protein biotinylated at the cell surface is represented in lane 1 (Total). The efficiency of biotin cleavage from receptors present in the plasma membrane was indicated by the nearly quantitative removal of biotin observed upon incubating biotinylated cells on ice with glutathione, shown in lane 2 (Strip). Internalized receptors were detected by their resistance to cleavage by glutathione at various times after warming cells to 37°C in the absence or presence of 10 μM dopamine for 5 min (lanes 3 and 4), 30 min (lane 5 and 6), or 60 min (lanes 7 and 8). To test the role of antagonist on endocytosis, internalization of D1 receptors was compared in cells incubated for 30 min in the absence of ligand (inset, lane 1), in the presence of 1 μM SCH23390 (inset, lane 2), or in the presence of 1 μM SCH23390 combined with 10 μM dopamine (inset, lane 3). The antagonist SCH23390 by itself had no effect on receptor endocytosis, but it completely inhibited agonist-induced endocytosis caused by dopamine. (D) Receptor internalization was quantitated by densitometric scanning of streptavidin blot data determined in the absence (dashed line) or presence (solid line) of 10 μM dopamine. Each data point represents the mean fraction of endocytosed D1 receptors (expressed as a percentage relative to the total amount of biotinylated receptor present without cleavage, n = 3–4). Error bars represent standard error of the mean.
FLAG-tagged D2 receptors visualized by fluorescence microscopy exhibited pronounced internalization both in the absence (Fig. 3 A) and presence (Fig. 3 B) of dopamine, suggesting that subtype-specific differences in the regulation of dopamine receptor endocytosis are conserved in nonneural cells. These observations were confirmed using the surface biotinylation assay, in which a significant pool of internalized, glutathione-resistant D2 receptors was detected both in the absence and presence of dopamine (Fig. 3 C). Significant internalization of D2 receptors was also observed in the presence of high concentrations (1 μM) of the potent D2 antagonist haloperidol, further suggesting that D2 receptors undergo significant, constitutive (ligand-independent) endocytosis (Fig. 3 C, inset). Quantitation of these results revealed that D2 receptors internalized nearly as rapidly as D1 receptors (t 1/2 ≤5 min). However, in contrast to the exclusively agonist-induced internalization of D1 receptors, ∼25% of the surface population of D2 receptors was internalized at steady state in the absence of agonist, compared with <5% net internalization for the D1 receptor. In the presence of dopamine, net internalization of D2 receptors was stimulated two- to threefold (Fig. 3 D). These observations suggest that, in addition to the constitutive internalization of D2 receptors observed in the absence of ligand or presence of antagonist, D2 receptors also exhibited a significant agonist-induced component of rapid endocytosis. Importantly, these biochemical assays of receptor internalization were performed in the absence of bound antibodies, suggesting that neither the constitutive nor agonist-induced components of receptor endocytosis were induced by antibody binding.
Figure 3.

Localization and endocytosis of D2 receptors in HEK293 cells. The localization of FLAG-tagged D2 receptors expressed in stably transfected HEK293 cells was examined (as in Fig. 2) in cells incubated for 30 min in the absence (A) or presence (B) of 10 μM dopamine. (C) Constitutive and agonist- induced internalization of D2 receptors was assayed biochemically using cell surface biotinylation. The total amount of biotinylated surface D2 receptor and the efficiency of biotin cleavage are indicated in lanes 1 (Total) and 2 (Strip), respectively. Receptor internalization was determined in cells incubated at 37°C either in the absence or presence of 10 μM dopamine for 5 min (lanes 3 and 4), 15 min (lane 5 and 6), 30 min (lanes 7 and 8), or 60 min (lanes 9 and 10). To test the role of antagonist on endocytosis, internalization of D2 receptors was compared in cells incubated for 30 min in the absence of ligand (inset, lane 1), in the presence of 1 μM haloperidol (inset, lane 2), or in the presence of 1 μM haloperidol combined with 10 μM dopamine (inset, lane 3). The D2 antagonist haloperidol did not by itself induce endocytosis but was unable to inhibit endocytosis of receptors observed in the absence of agonist, confirming that this endocytosis occurs constitutively. (D) Quantitation of these results by densitometric scanning of biotinylation data obtained in the absence (dashed line) or presence (solid line) of 10 μM dopamine. Each data point represents the mean from multiple experiments (n = 3–4), and error bars represent the standard error of the mean.
Interestingly, in addition to gel bands corresponding to the molecular mass of the monomeric, complex-glycosylated D2 receptor (50–60 kD), we also observed specific bands of immunoreactive receptor protein resolving at higher apparent molecular mass. Two of these bands, resolving at ∼100 and 150 kD, are consistent with dimeric and trimeric forms of the receptor protein, which have been observed previously with several types of G protein– coupled receptor including the D2 receptor (Hebert et al., 1996; Ng et al., 1996; Cvejic and Devi, 1997). The relative amount of these bands varied from experiment to experiment, and these apparent oligomers were not unique to the D2 receptor (a faint band corresponding to dimeric D1 receptors is apparent in the gel shown in Fig. 2). However, the band electrophoresing at ∼70 kD was observed consistently, is unlikely to represent oligomeric receptor, and was not observed in similar analyses of the D1 receptor. These observations suggest the possibility of a specific difference in the posttranslational processing of the D2 receptor, which may be relevant to its mechanism of endocytosis (see below).
D1 and D2 Receptors Are Selectively Endocytosed by Distinct Dynamin-dependent and -independent Mechanisms
To investigate molecular mechanisms mediating the subtype-specific endocytosis of dopamine receptors, we examined receptor internalization in HEK293 cells expressing K44E mutant dynamin, which specifically inhibits clathrin-mediated endocytosis in a dominant negative manner (Herskovits et al., 1993; Damke et al., 1994). D1 receptors remained associated with the plasma membrane (Fig. 4, A and C, respectively) in cells expressing K44E mutant dynamin (Fig. 4, B and D), both in the absence and presence of dopamine (although dopamine-induced clustering of receptors at the cell surface was observed, see arrows). Confirming this hypothesis, K44E mutant dynamin strongly inhibited dopamine-induced internalization of D1 receptors measured biochemically using the biotinylation assay (Fig. 4, E and F). However, wild-type dynamin expressed at similar levels did not block endocytosis of D1 receptors (data not shown), indicating the specificity of this inhibition. Furthermore, many of the D1 receptor clusters observed in the plasma membrane of dopamine-treated cells colocalized with clathrin, and endocytosis of D1 receptors was strongly inhibited in cells exposed to mildly hypertonic media (0.4 M sucrose) which inhibits clathrin function in these cells (Heuser and Anderson, 1989; Keith et al., 1996). Taken together, these observations strongly suggest that D1 receptors are endocytosed exclusively by a dynamin-dependent mechanism involving clathrin-coated pits.
Figure 4.

Endocytosis of D1 receptors is dynamin dependent. Stably transfected HEK293 cells expressing FLAG-tagged D1 receptors were transiently transfected with an expression plasmid encoding HA-tagged wild-type or K44E mutant dynamin, as described in Materials and Methods, and receptor localization was visualized by fluorescence microscopy. In cells incubated in the absence of agonist, D1 receptors (A) were localized exclusively in the plasma membrane, whereas mutant dynamin was observed throughout the cytoplasm (B). In the presence of dopamine, D1 receptors failed to endocytose in cells expressing mutant dynamin (C and D, respectively) and instead clustered in small puncta that remained associated with the plasma membrane (C, arrows). Identical experiments performed using wild-type dynamin did not block internalization of D1 receptors (not shown), indicating that the inhibition caused by the K44E dominant negative mutant is biochemically specific. (E) The effect of mutant dynamin on D1 receptor endocytosis was analyzed biochemically using the surface biotinylation assay. Mutant dynamin blocked D1 receptor internalization observed in the presence of dopamine (compare lanes 4 and 6) and also blocked the small amount of residual endocytosis observed in the absence of agonist (compare lanes 3 and 5). (F) The effect of mutant dynamin on D1 receptor internalization was quantitated by densitometric scanning of streptavidin blots. This analysis indicated that K44E mutant dynamin reduced agonist-induced internalization of D1 receptors observed after 30 min in the presence of 10 μM dopamine by ∼85% (compare bars 2 and 4).
We obtained significantly different results when internalization of D2 receptors was examined in the same manner. Fluorescence microscopy demonstrated numerous endocytic vesicles containing antibody-tagged D2 receptors both in the absence and presence of dopamine (Fig. 5, A and C), even in cells expressing K44E mutant dynamin at high levels (Fig. 5, B and D). These experiments were performed in parallel with those shown in Fig. 4, using comparable levels of mutant dynamin expression and identical experimental conditions. Biochemical analysis of receptor internalization using the biotinylation assay further supported the observation that the D2 receptor endocytosed when coexpressed with mutant dynamin. Neither the constitutive nor dopamine-induced components of net internalization of D2 receptors were detectably inhibited by K44E mutant dynamin (Fig. 5, E and F). Interestingly, whereas agonist-induced internalization of D2 receptors was not significantly inhibited in cells overexpressing K44E mutant dynamin (Fig. 5 F, compare bars 2 and 4), constitutive internalization of D2 receptors measured in the same cells was actually enhanced by approximately twofold (Fig. 5 F, compare bars 1 and 3). This observation is reminiscent of previous studies demonstrating that blockade of dynamin-dependent endocytosis causes a compensatory upregulation of alternate endocytic mechanism(s) (Damke et al., 1995). However, as our results represent net internalization determined both by endocytosis and recycling of receptors (see below), we cannot rule out the possibility that this effect on net internalization of receptors may reflect a dynamin-dependent change in receptor recycling.
Figure 5.

Endocytosis of D2 receptors is dynamin independent. Stably transfected HEK293 cells expressing FLAG-tagged D2 receptors were transiently transfected with K44E mutant dynamin and analyzed by fluorescence microscopy after incubation for 30 min in the absence (A and B) or presence (C and D) of 10 μM dopamine. In contrast to the complete blockade of D1 receptor internalization caused by dominant negative mutant dynamin, endocytosis of D2 receptors, both in the absence and presence of agonist, was readily detected (A and C, arrows). Biochemical assay of D2 receptor internalization using the surface biotinylation assay confirmed that K44E mutant dynamin did not inhibit either the constitutive (compare lanes 3 and 5) or agonist-induced components (compare lanes 4 and 6) of D2 receptor endocytosis (E, quantitated in F).
Both D1 and D2 Receptors Recycle to the Plasma Membrane Rapidly After Endocytosis
Having observed endocytosis of D1 and D2 receptors by distinct mechanisms, we next questioned whether these mechanisms target receptors differentially to degradative or recycling pathways. A biochemical assay of surface-biotinylated receptors was used to examine receptor degradation. These experiments indicated that the rate of degradation of both D1 and D2 receptors (t 1/2 >7 h, Fig. 6, A and B) was significantly slower than the rate of receptor internalization (t 1/2 ≤5 min, Figs. 2 and 3), indicating that D1 and D2 receptors are targeted inefficiently to lysosomes and suggesting that both receptors recycle efficiently to the plasma membrane.
Figure 6.

Neither D1 nor D2 receptors undergo rapid degradation. The degradation rates of biotinylated D1 or D2 receptors (A and B, respectively) were determined in the absence of agonist (dashed line) or in the continuous presence (solid line) of the chemically stable D1 or D2 receptor agonists 6-chloro-PB (10 μM) or quinpirole (10 μM), respectively, as described in Materials and Methods. Each data point represents the mean fraction of biotinylated receptor remaining at various time points (expressed as a percentage of the total amount of biotinylated surface receptor determined at t = 0), determined from densitometric scanning of streptavidin blots from multiple experiments (n = 3). Error bars represent the standard error of the mean.
To detect receptor recycling, we assayed the reappearance of biotinylated receptors at the cell surface by recovery of their susceptibility to membrane-impermeant reducing agent. Internalized D1 receptors (Fig. 7 A, lane 1) rapidly recovered susceptibility to cleavage by extracellular glutathione during a subsequent incubation of cells in the absence of dopamine (Fig. 7 A, lanes 2–4). The same assay indicated that a significant proportion of internalized D2 receptors also returned to the plasma membrane under these conditions (Fig. 7 B). Quantitation of these results confirmed this conclusion and indicated that both D1 and D2 receptors recycle significantly more rapidly (t 1/2 ≤30 min) than they degrade (t 1/2 >7 h, Fig. 6). In addition, D1 and D2 receptors labeled with mAb were observed by fluorescence microscopy to redistribute from endocytic vesicles to the cell surface within 30 min (Fig. 7, D–G).
Figure 7.

Both D1 and D2 receptors recycle back to the plasma membrane after endocytosis. A biochemical assay using surface biotinylation was used to determine the recycling of endocytosed receptors (inaccessible to cleavage by glutathione) back to the plasma membrane (where they become susceptible to cleavage by glutathione). HEK293 cells expressing FLAG-tagged D1 receptors were stimulated for 30 min in the presence of 10 μM dopamine to drive endocytosis, chilled to 4°C to arrest membrane trafficking, and then reincubated at 37°C to allow recycling for 5 min (A, lane 2), 15 min (lane 3), or 30 min (lane 4). D1 receptor completely recycled within 15 min when compared with the total pool of endocytosed D1 receptor (compare lanes 1 and 3). The identical experiment was performed using HEK293 cells expressing FLAG-tagged D2 receptors at comparable levels (B). Total endocytosed D2 receptors observed after incubation of cells for 30 min in the presence of 10 μM dopamine were determined (lane 1) and compared with the amount of internalized D2 receptor remaining in identical cells that were washed extensively on ice to remove agonist after stimulating endocytosis, then reincubated at 37°C in the absence of agonist for either 15 min (lane 2) or 30 min (lane 3). This analysis indicated that a significant amount of D2 receptor recycles back to the plasma membrane within 30 min (compare lanes 1 and 3). These biochemical data were quantitated by densitometric scanning of streptavidin blots to compare the recycling of D1 and D2 receptors (C, solid line and dashed line, respectively). Each data point represents the mean fraction of receptors that became accessible to glutathione (expressed as a percentage of the total internalized pool determined at t = 0) from determinations performed in duplicate from samples prepared in two independent experiments. Error bars represent the standard error of the mean. (D–G) Recycling of endocytosed D1 and D2 receptors was visualized by fluorescence microscopy. FLAG-tagged D1 or D2 receptors expressed in HEK293 cells were labeled in the plasma membrane with M1 mAb, and receptor endocytosis was allowed to occur for 30 min in the presence of 10 μM dopamine. Cells were then chilled on ice to arrest membrane trafficking, washed extensively to remove residual dopamine, and antibodies attached to receptors remaining in the plasma membrane were eluted by washing intact cells in the presence of EDTA, thereby specifically labeling only D1 or D2 receptors located in endocytic vesicles (D and F, respectively). Cells were then incubated at 37°C for 30 min in the absence of agonist, and recycling of antibody-labeled D1 and D2 receptors back to the plasma membrane was visualized by fluorescence microscopy (E and G).
Interestingly, the apparent rate of D2 recycling measured using the biochemical assay was slower than that of the D1 receptor (Fig. 7 C). However, because of the continuous endocytosis of D2 (but not D1) receptors in the absence of agonist, these assays are expected to significantly underestimate the rate and extent of D2 receptor recycling. Indeed, using the steady-state assumption (fraction of internalized receptors = k endocytosis/k recycling; this neglects the much slower rate of D2 receptor degradation), the data in Fig. 3 B suggest that the D2 receptor recycles with similar rapid kinetics as that of the D1 receptor.
Endocytosis of D1 and D2 Receptors by Distinct Membrane Vesicles in Neuroblastoma Cells
We next examined whether D1 and D2 receptors are endocytosed by distinct membrane vesicles in neuroblastoma cells and, if so, whether this selectivity is maintained in cells overexpressing both receptors. To accomplish this, D1 and D2 receptors tagged with different epitopes were coexpressed in Neuro2A cells and preincubated in the presence of mAb and dopamine at 4°C. This allowed D1 and D2 receptors present in the plasma membrane to be labeled with antibodies and bind agonist under conditions that do not allow endocytosis of receptors. Then cells were rapidly warmed to 37°C in the continued presence of dopamine to allow endocytosis of both antibody-labeled receptors, and receptor localization was visualized in cells fixed after initiating the pulse of endocytosis. In cells warmed to 37°C for short time periods (e.g., 4 min, Fig. 8), endocytosis of antibody-labeled D1 and D2 receptors (Fig. 8, A and B, respectively) was readily detected in the same cells. Significantly, most endocytic vesicles observed at this time point were selectively enriched in D1 or D2 receptors, whereas colocalization of receptors in the same vesicles was observed only rarely at this time point [Fig. 8 C, note the differential membrane localization of D1 (green) and D2 (red) receptor immunoreactivity]. The distinct membrane localization of D1 and D2 receptors was particularly apparent by examination of regions close to the plasma membrane at higher magnification (Fig. 8 C, inset).
Figure 8.
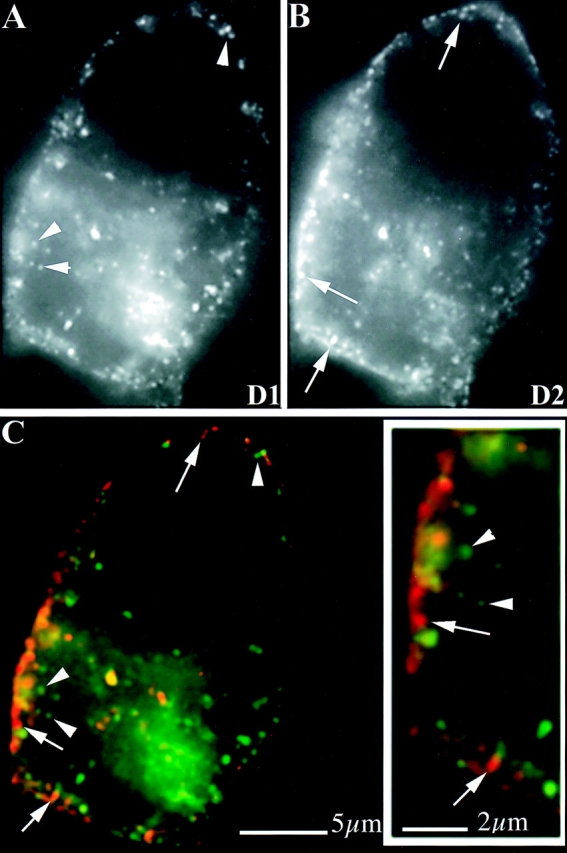
Selective endocytosis of D1 and D2 receptors by distinct membrane vesicles in neuroblastoma cells. Neuro2A neuroblastoma cells coexpressing the HA-tagged D1 and FLAG-tagged D2 receptors were surface labeled at 4°C with specific mAbs in the presence of 10 μM dopamine, as described in Materials and Methods. Cells were then warmed to 37°C for various time periods in the continued presence of dopamine, to allow endocytosis of both receptors to occur, then cells were fixed and processed for dual label fluorescence microscopy to visualize the localization of D1 and D2 receptors in the same cells. At short times after warming cells (e.g., 4 min), internalized D1 and D2 receptors were visualized predominantly in different endocytic vesicles, as apparent in the monochrome image pairs (A and B, respectively) and emphasized in the merged color image (C, D1 immunoreactivity is indicated in green and D2 in red). This differential localization of receptors was most pronounced upon examination of regions close to the plasma membrane at higher magnification (C, inset), where endocytic membranes differentially concentrated in D1 (arrowheads) or D2 (arrows) receptors were observed.
Discussion
In this study, we compared the endocytic trafficking of two structurally homologous subtypes of dopamine receptor that are of particular physiological interest. We observed that these receptors are selectively endocytosed in the same cells by distinct mechanisms and primary endocytic membranes. These observations constitute, to our knowledge, the first example of subtype-specific sorting of signaling receptors by distinct endocytic membranes when coexpressed in the same cells, and they provide a favorable system for examining the membrane trafficking of signaling receptors after endocytosis by distinct mechanisms.
In principle, molecular sorting operations that mediate the selective membrane trafficking of signaling receptors could occur at multiple locations within the cell. In the case of constitutively endocytosed receptors that transport nutrient ligands (e.g., LDL and mannose-6-phosphate receptors), individual receptors are endocytosed by similar clathrin-coated pits but differ subsequently in their sorting between different membrane trafficking pathways after delivery to the same early endosomes (Trowbridge et al., 1993). Thus, studies of nutrient receptors have clearly established that a great deal of molecular sorting occurs in early endocytic vesicles after the initial endocytosis of receptors (Kornfeld and Mellman, 1989). Conversely, endocytosis mediated by a clathrin-independent mechanism can deliver nutrient ligands and toxins to the same early endosomes as endocytosis mediated by clathrin-coated pits, suggesting that distinct endocytic mechanisms may play a redundant function in delivering cargo to the same early endosomes (Hansen et al., 1993). The present observations indicate that structurally homologous G protein– coupled signaling receptors are efficiently sorted in the plasma membrane by distinct endocytic mechanisms, even when overexpressed in heterologous cell types. Such efficient sorting at the cell surface may be of particular importance for controlling the activation-dependent membrane trafficking of signaling receptors, including dopamine receptors, which can be inactivated by protein phosphorylation significantly more rapidly than they are endocytosed (Tiberi et al., 1996). Thus, the molecular machinery required to appropriately target receptors in response to agonist stimulation may be able to recognize activated receptors only in the plasma membrane and not after endocytosis.
Dynamin-dependent endocytosis of certain G protein– coupled receptors is mediated by phosphorylation-dependent association of activated receptors with beta-arrestin, which can then serve as an adapter protein for recruiting receptors into clathrin-coated pits (Goodman et al., 1996; Goodman et al., 1998). This mechanism may apply to the agonist-induced endocytosis of the D1 receptor, as ligand-dependent phosphorylation of this receptor has been demonstrated previously (Ng et al., 1994a; Tiberi et al., 1996). The mechanism of dynamin-independent endocytosis of D2 receptors remains to be elucidated. Sequence examination indicates that, despite extensive conservation in transmembrane and extracellular domains, the cytoplasmic domains of the D1 and D2 receptors are highly divergent. Furthermore, our biochemical studies suggest that D2 receptors may differ in their posttranslational modification from D1 receptors (Fig. 3). In particular, the ∼70-kD form of the receptor protein is not consistent with oligomeric receptor complexes, and does not correspond to phosphorylated receptor forms identified in previous studies (Ng et al., 1994b). The ∼70-kD form of D2 receptor can also be observed using metabolic labeling of cells with [35S]methionine and is not reduced to monomeric size by digestion with PNGase F, suggesting that this difference in apparent molecular mass is not caused by N-linked glycosylation (Vickery, R.G., and M. von Zastrow, unpublished observations). Thus, other posttranslational modifications may distinguish the D2 receptor and play a role in the alternate mechanism of D2 receptor endocytosis. The possible role of ubiquitination is particularly interesting in view of the well established role of this posttranslational modification in promoting dynamin-independent endocytosis of the G protein–coupled receptor Ste2p in Saccharomyces cerevisiae (Hicke and Riezman, 1996; Hicke et al., 1998). However, this process is ligand induced and can be facilitated by clathrin (Tan et al., 1993). Therefore, it will be interesting to determine to what extent the alternate mechanism of D2 receptor endocytosis observed in mammalian cells may be similar to endocytosis of mating factor receptors in yeast. Interestingly, the D2 receptor contains 20 lysine residues located in various cytoplasmic domains, most of which are not conserved in the D1 receptor. However, we have not yet determined whether any of these residues is ubiquitinated in mammalian cells, or indeed whether ubiquitination plays any role in the membrane trafficking of D2 receptors. Studies in progress, which are attempting to define specific cytoplasmic residues that distinguish the endocytic properties of D1 and D2 receptors, may provide further insight into these questions.
We initially expected that the selective endocytosis of D1 and D2 receptors by distinct mechanisms might target individual receptors directly from the cell surface to different recycling and degradative pathways, as predicted from the results of previous studies (Valiquette et al., 1990; Hausdorff et al., 1991; Zhang et al., 1996). Our biochemical studies suggest that this is not the case because both D1 and D2 receptors, although endocytosed by distinct mechanisms, recycle to the plasma membrane with similarly high efficiency and are degraded relatively inefficiently. However, we cannot state at the present time the extent to which these early endocytic pathways remain separated or converge during the process of D1 and D2 receptor recycling.
What physiological role might this efficient sorting of signaling receptors in the plasma membrane serve? In the case of neurons, where D1 and D2 receptors mediate distinct postsynaptic and presynaptic functions, it is tempting to speculate that the selective endocytosis of D2 receptors by a distinct, dynamin-independent mechanism may play an important role in presynaptic physiology. Postsynaptic D1 receptors are located in the somatodendritic plasma membrane (Hersch et al., 1995; Yung et al., 1995; Hersch et al., 1997), as are constitutively endocytosed nutrient receptors (Cameron et al., 1991; Jareb and Banker, 1998). Presynaptic D2 receptors, in contrast, are localized in the same presynaptic membrane specializations at which synaptic vesicle release and recycling occur. Despite this close proximity, the extremely high concentration of dopamine present in synaptic vesicles makes it physiologically imperative to keep D2 autoreceptors physically separated from endocytosed synaptic vesicle membranes. Indeed, immunohistochemical studies suggest that synaptic vesicle membranes are not enriched in presynaptic D2 receptors, even under conditions of strong synaptic activation that greatly increase the rate of synaptic vesicle membrane recycling (Sesack et al., 1994). Thus, the selective endocytosis of D2 receptors by a distinct mechanism could efficiently separate presynaptic autoreceptors from synaptic vesicle membrane components immediately upon endocytosis, thereby circumventing the need to sort autoreceptors from a common endocytic intermediate (Cameron et al., 1991; Cameron et al., 1993). Although further studies will be necessary to test this hypothesis, it is interesting to note that D1 and D2 receptors are differentially distributed among distinct endocytic vesicles at the cell periphery and in extended cell processes of neuroblastoma cells at the earliest stages of endocytosis.
These studies indicate that D1 and D2 receptors are also efficiently sorted when overexpressed in nonneural cells, suggesting that the endocytosis of structurally homologous signaling receptors by distinct membranes is highly conserved and may mediate other important physiological function(s). Recent studies of both G protein–coupled receptors and receptor tyrosine kinases suggest that dynamin-dependent endocytosis delivers signaling receptors to endocytic membranes required for mitogenic signal transduction via a subset of downstream effectors (Vieira et al., 1996; Daaka et al., 1998). Thus, it is possible that the efficient segregation of D1 and D2 dopamine receptors among distinct endocytic membranes may play an important role in determining the fidelity of subtype-specific receptor signaling. Indeed, whereas D2 receptors can be demonstrated to couple promiscuously to multiple downstream effectors when studied in vitro, receptor-mediated signal transduction observed in vivo is remarkably specific (Missale et al., 1998). Therefore, our results suggest a new role of distinct endocytic mechanisms in physically segregating structurally homologous signaling receptors in the plasma membrane and shortly after endocytosis, where functional interaction between receptors and specific effector molecules is required for physiologically appropriate signal transduction.
In summary, these results establish a new level of molecular selectivity in the endocytic membrane trafficking of signaling receptors. This selectivity is highly conserved in diverse cell types and segregates structurally homologous subtypes of G protein–coupled signaling receptor at the plasma membrane and at a very early stage in the endocytic pathway. So far, studies of the endocytosis of signaling receptors have focused primarily on dynamin-dependent mechanisms, even though there is increasing evidence that distinct, dynamin-independent mechanism(s) mediate the endocytosis of certain signaling receptors. These results provide the first evidence that dynamin-dependent and -independent endocytic mechanisms play a role in distinguishing the properties of structurally homologous receptors coexpressed in the same cells, suggesting a new role of distinct endocytic membrane trafficking mechanisms in mediating the physiological specificity of cellular signal transduction.
Acknowledgments
We are grateful to Steven Murray, Uwe Klein, Tracy Cao, and Heather Deacon for insightful suggestions during the course of these studies and for critical comments on the manuscript. We thank David Grandy for providing cDNA encoding human D1 and D2 receptors, David Sibley for providing cDNA encoding the rat D1 receptor used in preliminary studies, Richard Vallee for cDNAs encoding mutant dynamin, Uwe Klein for preparing biotinylated-M1 mAb used in preliminary studies, Peter Chu for performing adenylyl cyclase assays and preparing FITC-conjugated M1 antibody, and Alexander Hillel for assistance with cell culture.
Abbreviation used in this paper
HEK
human embryonic kidney
Footnotes
These studies were funded by a grant from the National Alliance for Research on Schizophrenia and Depression (to R.G. Vickery), a postdoctoral training fellowship from the National Institutes of Health (to R.G. Vickery), and a research grant from the Tourette Syndrome Association (to M. von Zastrow). R.G. Vickery is a Staglin Fellow in Molecular Neurobiology.
References
- Apodaca G, Katz LA, Mostov KE. Receptor-mediated transcytosis of IgA in MDCK cells is via apical recycling endosomes. J Cell Biol. 1994;125:67–86. doi: 10.1083/jcb.125.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bohm SK, Khitin LM, Grady EF, Aponte G, Payan DG, Bunnett NW. Mechanisms of desensitization and resensitization of proteinase-activated receptor-2. J Biol Chem. 1996;271:22003–22016. doi: 10.1074/jbc.271.36.22003. [DOI] [PubMed] [Google Scholar]
- Cameron P, Mundigl O, De Camilli P. Traffic of synaptic vesicle proteins in polarized and nonpolarized cells. J Cell Sci Suppl. 1993;17:93–100. doi: 10.1242/jcs.1993.supplement_17.14. [DOI] [PubMed] [Google Scholar]
- Cameron PL, Südhof TC, Jahn R, De Camilli P. Colocalization of synaptophysin with transferrin receptors: implications for synaptic vesicle biogenesis. J Cell Biol. 1991;115:151–164. doi: 10.1083/jcb.115.1.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao TC, Mays RW, von Zastrow M. Regulated endocytosis of G protein–coupled receptors by a biochemically and functionally distinct subpopulation of clathrin-coated pits. J Biol Chem. 1998;273:24592–24602. doi: 10.1074/jbc.273.38.24592. [DOI] [PubMed] [Google Scholar]
- Carpentier JL, Gorden P, Anderson RG, Goldstein JL, Brown MS, Cohen S, Orci L. Co-localization of 125I-epidermal growth factor and ferritin low-density lipoprotein in coated pits: a quantitative electron microscopic study in normal and mutant human fibroblasts. J Cell Biol. 1982;95:73–77. doi: 10.1083/jcb.95.1.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cvejic S, Devi LA. Dimerization of the delta opioid receptor: implication for a role in receptor internalization. J Biol Chem. 1997;272:26959–26964. doi: 10.1074/jbc.272.43.26959. [DOI] [PubMed] [Google Scholar]
- Daaka Y, Luttrell LM, Ahn S, Della GJ, Rocca, Ferguson SS, Caron MG, Lefkowitz RJ. Essential role for G protein–coupled receptor endocytosis in the activation of mitogen-activated protein kinase. J Biol Chem. 1998;273:685–688. doi: 10.1074/jbc.273.2.685. [DOI] [PubMed] [Google Scholar]
- Damke H, Baba T, Warnock DE, Schmid SL. Induction of mutant dynamin specifically blocks endocytic coated vesicle formation. J Cell Biol. 1994;127:915–934. doi: 10.1083/jcb.127.4.915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damke H, Baba T, van der Bliek AM, Schmid SL. Clathrin-independent pinocytosis is induced in cells overexpressing a temperature-sensitive mutant of dynamin. J Cell Biol. 1995;131:69–80. doi: 10.1083/jcb.131.1.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daunt DA, Hurt C, Hein L, Kallio J, Feng F, Kobilka BK. Subtype-specific intracellular trafficking of alpha2-adrenergic receptors. Mol Pharmacol. 1997;51:711–720. doi: 10.1124/mol.51.5.711. [DOI] [PubMed] [Google Scholar]
- Dupree P, Parton RG, Raposo G, Kurzchalia TV, Simons K. Caveolae and sorting in the trans-Golgi network of epithelial cells. EMBO (Eur Mol Biol Organ) J. 1993;12:1597–1605. doi: 10.1002/j.1460-2075.1993.tb05804.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gagnon AW, Kallal L, Benovic JL. Role of clathrin-mediated endocytosis in agonist-induced down-regulation of the beta2-adrenergic receptor. J Biol Chem. 1998;273:6976–6981. doi: 10.1074/jbc.273.12.6976. [DOI] [PubMed] [Google Scholar]
- Goodman OJ, Krupnick JG, Santini F, Gurevich VV, Penn RB, Gagnon AW, Keen JH, Benovic JL. Beta-arrestin acts as a clathrin adaptor in endocytosis of the beta2-adrenergic receptor. Nature. 1996;383:447–450. doi: 10.1038/383447a0. [DOI] [PubMed] [Google Scholar]
- Goodman OJ, Krupnick JG, Santini F, Gurevich VV, Penn RB, Gagnon AW, Keen JH, Benovic JL. Role of arrestins in G-protein-coupled receptor endocytosis. Adv Pharmacol. 1998;42:429–433. doi: 10.1016/s1054-3589(08)60780-2. [DOI] [PubMed] [Google Scholar]
- Grandy DK, Marchionni MA, Makam H, Stofko RE, Alfano M, Frothingham L, Fischer JB, Burke-Howie KJ, Bunzow JR, Server AC, et al. Cloning of the cDNA and gene for a human D2 dopamine receptor. Proc Natl Acad Sci USA. 1989;86:9762–9766. doi: 10.1073/pnas.86.24.9762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guan XM, Kobilka TS, Kobilka BK. Enhancement of membrane insertion and function in a type IIIb membrane protein following introduction of a cleavable signal peptide. J Biol Chem. 1992;267:21995–21998. [PubMed] [Google Scholar]
- Hansen SH, Sandvig K, van Deurs B. Molecules internalized by clathrin-independent endocytosis are delivered to endosomes containing transferrin receptors. J Cell Biol. 1993;123:89–97. doi: 10.1083/jcb.123.1.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hausdorff WP, Campbell PT, Ostrowski J, Yu SS, Caron MG, Lefkowitz RJ. A small region of the beta-adrenergic receptor is selectively involved in its rapid regulation. Proc Natl Acad Sci USA. 1991;88:2979–2983. doi: 10.1073/pnas.88.8.2979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hebert TE, Moffett S, Morello JP, Loisel TP, Bichet DG, Barret C, Bouvier M. A peptide derived from a beta2-adrenergic receptor transmembrane domain inhibits both receptor dimerization and activation. J Biol Chem. 1996;271:16384–16392. doi: 10.1074/jbc.271.27.16384. [DOI] [PubMed] [Google Scholar]
- Hersch SM, Ciliax BJ, Gutekunst CA, Rees HD, Heilman CJ, Yung KK, Bolam JP, Ince E, Yi H, Levey AI. Electron microscopic analysis of D1 and D2 dopamine receptor proteins in the dorsal striatum and their synaptic relationships with motor corticostriatal afferents. J Neurosci. 1995;15:5222–5237. doi: 10.1523/JNEUROSCI.15-07-05222.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hersch SM, Yi H, Heilman CJ, Edwards RH, Levey AI. Subcellular localization and molecular topology of the dopamine transporter in the striatum and substantia nigra. J Comp Neurol. 1997;388:211–227. [PubMed] [Google Scholar]
- Herskovits JS, Burgess CC, Obar RA, Vallee RB. Effects of mutant rat dynamin on endocytosis. J Cell Biol. 1993;122:565–578. doi: 10.1083/jcb.122.3.565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heuser JE, Anderson RG. Hypertonic media inhibit receptor-mediated endocytosis by blocking clathrin-coated pit formation. J Cell Biol. 1989;108:389–400. doi: 10.1083/jcb.108.2.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hicke L, Riezman H. Ubiquitination of a yeast plasma membrane receptor signals its ligand-stimulated endocytosis. Cell. 1996;84:277–287. doi: 10.1016/s0092-8674(00)80982-4. [DOI] [PubMed] [Google Scholar]
- Hicke L, Zanolari B, Riezman H. Cytoplasmic tail phosphorylation of the alpha-factor receptor is required for its ubiquitination and internalization. J Cell Biol. 1998;141:349–358. doi: 10.1083/jcb.141.2.349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopkins CR, Miller K, Beardmore JM. Receptor-mediated endocytosis of transferrin and epidermal growth factor receptors: a comparison of constitutive and ligand-induced uptake. J Cell Sci Suppl. 1985;3:173–186. doi: 10.1242/jcs.1985.supplement_3.17. [DOI] [PubMed] [Google Scholar]
- Jaber M, Robinson SW, Missale C, Caron MG. Dopamine receptors and brain function. Neuropharmacology. 1996;35:1503–1519. doi: 10.1016/s0028-3908(96)00100-1. [DOI] [PubMed] [Google Scholar]
- Jareb M, Banker G. The polarized sorting of membrane proteins expressed in cultured hippocampal neurons using viral vectors. Neuron. 1998;20:855–867. doi: 10.1016/s0896-6273(00)80468-7. [DOI] [PubMed] [Google Scholar]
- Keith DE, Murray SR, Zaki PA, Chu PC, Lissin DV, Kang L, Evans CJ, von Zastrow M. Morphine activates opioid receptors without causing their rapid internalization. J Biol Chem. 1996;271:19021–19024. doi: 10.1074/jbc.271.32.19021. [DOI] [PubMed] [Google Scholar]
- Kornfeld S, Mellman I. The biogenesis of lysosomes. Annu Rev Cell Biol. 1989;5:483–525. doi: 10.1146/annurev.cb.05.110189.002411. [DOI] [PubMed] [Google Scholar]
- Lefkowitz RJ, Pitcher J, Krueger K, Daaka Y. Mechanisms of beta-adrenergic receptor desensitization and resensitization. Adv Pharmacol. 1998;42:416–420. doi: 10.1016/s1054-3589(08)60777-2. [DOI] [PubMed] [Google Scholar]
- Lohse MJ. Molecular mechanisms of membrane receptor desensitization. Biochim Biophys Acta. 1993;1179:171–188. doi: 10.1016/0167-4889(93)90139-g. [DOI] [PubMed] [Google Scholar]
- Missale C, Nash SR, Robinson SW, Jaber M, Caron MG. Dopamine receptors: from structure to function. Physiol Rev. 1998;78:189–225. doi: 10.1152/physrev.1998.78.1.189. [DOI] [PubMed] [Google Scholar]
- Mostafapour S, Kobilka BK, von Zastrow M. Pharmacological sequestration of a chimeric beta 3/beta 2 adrenergic receptor occurs without a corresponding amount of receptor internalization. Recept Signal Transduct. 1996;6:151–163. [PubMed] [Google Scholar]
- Mostov KE, Cardone MH. Regulation of protein traffic in polarized epithelial cells. Bioessays. 1995;17:129–138. doi: 10.1002/bies.950170208. [DOI] [PubMed] [Google Scholar]
- Nantel F, Bonin H, Emorine LJ, Zilberfarb V, Strosberg AD, Bouvier M, Marullo S. The human beta 3-adrenergic receptor is resistant to short term agonist-promoted desensitization. Mol Pharmacol. 1993;43:548–555. [PubMed] [Google Scholar]
- Ng GY, Mouillac B, George SR, Caron M, Dennis M, Bouvier M, O'Dowd BF. Desensitization, phosphorylation and palmitoylation of the human dopamine D1 receptor. Eur J Pharmacol. 1994a;267:7–19. doi: 10.1016/0922-4106(94)90219-4. [DOI] [PubMed] [Google Scholar]
- Ng GY, O'Dowd BF, Caron M, Dennis M, Brann MR, George SR. Phosphorylation and palmitoylation of the human D2L dopamine receptor in Sf9 cells. J Neurochem. 1994b;63:1589–1595. doi: 10.1046/j.1471-4159.1994.63051589.x. [DOI] [PubMed] [Google Scholar]
- Ng GY, O'Dowd BF, Lee SP, Chung HT, Brann MR, Seeman P, George SR. Dopamine D2 receptor dimers and receptor-blocking peptides. Biochem Biophys Res Commun. 1996;227:200–204. doi: 10.1006/bbrc.1996.1489. [DOI] [PubMed] [Google Scholar]
- Ng GY, Varghese G, Chung HT, Trogadis J, Seeman P, O'Dowd BF, George SR. Resistance of the dopamine D2L receptor to desensitization accompanies the up-regulation of receptors on to the surface of Sf9 cells. Endocrinology. 1997;138:4199–4206. doi: 10.1210/endo.138.10.5433. [DOI] [PubMed] [Google Scholar]
- Okayama H, Berg P. A cDNA cloning vector that permits expression of cDNA inserts in mammalian cells. Mol Cell Biol. 1983;3:280–289. doi: 10.1128/mcb.3.2.280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pals-Rylaarsdam R, Gurevich VV, Lee KB, Ptasienski JA, Benovic JL, Hosey MM. Internalization of the m2 muscarinic acetylcholine receptor. Arrestin-independent and -dependent pathways. J Biol Chem. 1997;272:23682–23689. doi: 10.1074/jbc.272.38.23682. [DOI] [PubMed] [Google Scholar]
- Raposo G, Dunia I, Delavier KC, Kaveri S, Strosberg AD, Benedetti EL. Internalization of beta-adrenergic receptor in A431 cells involves non-coated vesicles. Eur J Cell Biol. 1989;50:340–352. [PubMed] [Google Scholar]
- Seaman MN, Burd CG, Emr SD. Receptor signalling and the regulation of endocytic membrane transport. Curr Opin Cell Biol. 1996;8:549–556. doi: 10.1016/s0955-0674(96)80034-2. [DOI] [PubMed] [Google Scholar]
- Sesack SR, Aoki C, Pickel VM. Ultrastructural localization of D2 receptor-like immunoreactivity in midbrain dopamine neurons and their striatal targets. J Neurosci. 1994;14:88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan PK, Davis NG, Sprague GF, Payne GS. Clathrin facilitates the internalization of seven transmembrane segment receptors for mating pheromones in yeast. J Cell Biol. 1993;123:1707–1716. doi: 10.1083/jcb.123.6.1707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiberi M, Nash SR, Bertrand L, Lefkowitz RJ, Caron MG. Differential regulation of dopamine D1A receptor responsiveness by various G protein–coupled receptor kinases. J Biol Chem. 1996;271:3771–3778. doi: 10.1074/jbc.271.7.3771. [DOI] [PubMed] [Google Scholar]
- Trogadis JE, Ng GY, O'Dowd BF, George SR, Stevens JK. Dopamine D1 receptor distribution in Sf9 cells imaged by confocal microscopy: a quantitative evaluation. J Histochem Cytochem. 1995;43:497–506. doi: 10.1177/43.5.7730588. [DOI] [PubMed] [Google Scholar]
- Trowbridge IS, Collawn JF, Hopkins CR. Signal-dependent membrane protein trafficking in the endocytic pathway. Annu Rev Cell Biol. 1993;9:129–161. doi: 10.1146/annurev.cb.09.110193.001021. [DOI] [PubMed] [Google Scholar]
- Valiquette M, Bonin H, Hnatowich M, Caron MG, Lefkowitz RJ, Bouvier M. Involvement of tyrosine residues located in the carboxyl tail of the human beta 2-adrenergic receptor in agonist-induced down-regulation of the receptor. Proc Natl Acad Sci USA. 1990;87:5089–5093. doi: 10.1073/pnas.87.13.5089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Deurs B, Petersen OW, Olsnes S, Sandvig K. The ways of endocytosis. Int Rev Cytol. 1989;117:131–177. doi: 10.1016/s0074-7696(08)61336-4. [DOI] [PubMed] [Google Scholar]
- Vieira AV, Lamaze C, Schmid SL. Control of EGF receptor signaling by clathrin-mediated endocytosis. Science. 1996;274:2086–2089. doi: 10.1126/science.274.5295.2086. [DOI] [PubMed] [Google Scholar]
- von Zastrow M, Kobilka BK. Ligand-regulated internalization and recycling of human beta 2-adrenergic receptors between the plasma membrane and endosomes containing transferrin receptors. J Biol Chem. 1992;267:3530–3538. [PubMed] [Google Scholar]
- von Zastrow M, Link R, Daunt D, Barsh G, Kobilka B. Subtype-specific differences in the intracellular sorting of G protein–coupled receptors. J Biol Chem. 1993;268:763–766. [PubMed] [Google Scholar]
- Yung KK, Bolam JP, Smith AD, Hersch SM, Ciliax BJ, Levey AI. Immunocytochemical localization of D1 and D2 dopamine receptors in the basal ganglia of the rat: light and electron microscopy. Neuroscience. 1995;65:709–730. doi: 10.1016/0306-4522(94)00536-e. [DOI] [PubMed] [Google Scholar]
- Zhang J, Ferguson S, Barak LS, Menard L, Caron MG. Dynamin and beta-arrestin reveal distinct mechanisms for G protein–coupled receptor internalization. J Biol Chem. 1996;271:18302–18305. doi: 10.1074/jbc.271.31.18302. [DOI] [PubMed] [Google Scholar]
- Zhou QY, Grandy DK, Thambi L, Kushner JA, Van Tol HH, Cone R, Pribnow D, Salon J, Bunzow JR, Civelli O. Cloning and expression of human and rat D1 dopamine receptors. Nature. 1990;347:76–80. doi: 10.1038/347076a0. [DOI] [PubMed] [Google Scholar]