Regulation of body temperature by the nervous system (original) (raw)
. Author manuscript; available in PMC: 2019 Apr 4.
Summary
The regulation of body temperature is one of the most critical functions of the nervous system. Here we review our current understanding of thermoregulation in mammals. We outline the molecules and cells that measure body temperature in the periphery, the neural pathways that communicate this information to the brain, and the central circuits that coordinate the homeostatic response. We also discuss some of the key unresolved questions in this field, including: the role of temperature sensing in the brain; the molecular identity of the warm-sensor; the central representation of the labelled line for cold; and the neural substrates of thermoregulatory behavior. We suggest that approaches for molecularly-defined circuit analysis will provide new insight into these questions in the near future.
Introduction
Birds and mammals have the remarkable ability to regulate their internal temperature within a narrow range that is higher than the surroundings. The reason for this is unknown. One hypothesis is that elevated body temperature evolved as a secondary consequence of the higher metabolic rates needed for sustained activity (e.g. flight) or occupation of new ecological niches (e.g. nocturnal foraging and cold climates) (Bennett and Ruben, 1979; Crompton et al., 1978; Heinrich, 1977). Over time, this elevated body temperature may have become defended as a means to enable the optimization of cellular processes for a specific temperature range (Heinrich, 1977). Whatever the reason, the emergence of elevated but stable body temperature was a key event that accompanied the proliferation of birds and mammals across the globe, and an understanding of the thermoregulatory system is central to understanding our own physiology.
In this review, we describe the neural mechanisms that regulate body temperature in mammals. First, we outline some of the basic principles of the thermoregulatory system as a whole. Next, we summarize what is known about the molecules, cells, and tissues that measure temperature at different sites in the body, and the pathways by which they communicate this information to the brain. We then describe our current understanding of the circuits in the brain that integrate temperature information and coordinate the behavioral and autonomic response. Finally, we highlight some of the key questions that remain to be answered.
1. The organization of the thermoregulatory system
Feed-forward and feedback regulation of body temperature
Body temperature is not a single value but varies depending on where it is measured. In studies of thermoregulation, it is common to divide the body into two compartments: (1) the external shell, which includes the skin and largely fluctuates in temperature along with the environment, and (2) the internal core, which includes the central nervous system and viscera and has relatively stable temperature (Jessen, 1985; Romanovsky et al., 2009).
The core temperature is the regulated variable in the thermoregulatory system (Hensel, 1973) and is maintained by a combination of feedback and feed-forward mechanisms (Kanosue et al., 2010). Feedback responses are those that are triggered when core temperature deviates from the defended range: for example, exercise generates heat that can increase internal temperature by several degrees Celsius (Fuller et al., 1998; Walters et al., 2000) (Figure 1). Such changes in internal temperature are detected by specialized thermoreceptors located throughout the body core, including the viscera, brain, and spinal cord (Jessen, 1985). Localized heating or cooling of any of these internal structures induces global feedback responses that oppose the applied temperature change.
Figure 1. Core temperature during challenges to thermal homeostasis.
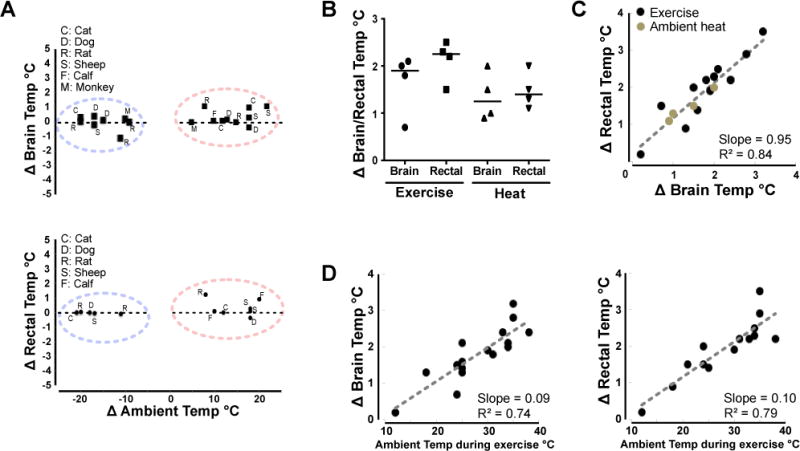
A. Changes in brain or rectal temperatures are typically small during acute external temperature challenges (30-60 minutes) in a range of mammals.
B. Changes in brain and rectal temperature in the rat after 30 minutes of exercise (treadmill, average speed 20 m/min, average ambient temp 27°C) or 30 minutes of heat exposure (average 45°C). Solid lines – median
C. Rat brain and rectal temperatures are tightly correlated during either exercise or external heating.
D. Exercise-induced warming in the rat brain and core are sensitive to prevailing ambient temperatures
Feed-forward mechanisms are triggered in the absence of any change in core temperature and instead enable preemptive responses to anticipated thermal challenges. The most common example of feed-forward control is the detection of a change in air temperature by thermoreceptors in the skin, which triggers thermoregulatory responses that precede and prevent any change in core temperature (Nakamura and Morrison, 2008, 2010; Romanovsky, 2014). Although feed-forward and feedback signals convey different kinds of information about body temperature, they are thought to converge on a common set of neural substrates in the preoptic area (POA) of the hypothalamus.
Physiologic versus behavioral thermoregulation
Body temperature is regulated by two types of mechanisms, physiologic and behavioral (Figure 2). Physiologic effectors are involuntary, mostly autonomic responses that generate or dissipate heat. The primary physiologic responses to cold exposure are brown adipose tissue (BAT) thermogenesis and skeletal muscle shivering, which generate heat, and the constriction of blood vessels (vasoconstriction), which prevents heat loss. Exposure to warmth triggers a complementary set of autonomic responses, including suppression of thermogenesis and facilitation of heat loss through water evaporation (e.g. sweating) and dilation of blood vessels (vasodilation).
Figure 2. Types of thermoregulatory effectors.

Examples of physiological and behavioral strategies for controlling body temperature.
Different species sometimes use different strategies to achieve the same physiologic effect. For example, humans achieve evaporative heat loss primarily by sweating, whereas dogs rely on panting and rodents spread saliva on their fur (Jessen, 1985). Likewise the effects of vasodilation are enhanced in species that have specialized thermoregulatory organs, such as the rat tail or rabbit ears, that can rapidly dissipate heat due to their large surface area. Despite these superficial differences, the major classes of physiologic responses are thought to be governed by a common set of neural substrates that are conserved across mammals.
Behavior is also an important mechanism for body temperature control. Whereas physiologic responses are involuntary, thermoregulatory behaviors are motivated, meaning that they are flexible, goal-oriented actions that are learned by reinforcement and driven by the expectation of reward (Carlton and Marks, 1958; Epstein and Milestone, 1968; Weiss and Laties, 1961). The most basic thermoregulatory behaviors are cold and warm-seeking, in which animals move between microenvironments in their habitat in order to alter the rate of heat loss or absorption. More complex thermoregulatory behaviors include nest or burrow making, in which animals create their own thermal microenvironment (Terrien et al., 2011); social behaviors such as huddling between conspecifics (Batchelder et al., 1983); and human behaviors such as wearing clothing or using air-conditioning.
The engagement of specific thermoregulatory mechanisms is hierarchical, meaning that different effectors become activated at different temperature thresholds. In general, behavioral responses are utilized in preference to autonomic effectors, and autonomic effectors are activated in a stereotyped sequence. This sequence is thought to reflect the “cost” of activating different responses, either in terms of their energy use or the trade-offs they require with competing physiologic systems. For example, heat challenge triggers vasodilation at lower temperatures than sweating, possibly because sweating results in water loss that upsets fluid balance (Costill and Fink, 1974). Similarly cold challenge activates vasoconstriction before shivering or BAT thermogenesis, in accordance with the relative energy cost of these different mechanisms. The existence of these distinct temperature thresholds has been interpreted as evidence that the thermoregulatory circuit contains multiple effector loops, each of which operates to some extent independently (McAllen et al., 2010; Satinoff, 1978).
Interactions between thermoregulation and other physiologic systems
The core temperature defended by the thermoregulatory system (the “balance point” or “set point”) is not a fixed value but fluctuates in response to internal and external factors. Many of these factors are unrelated to temperature per se and instead reflect interactions with other physiologic systems. One example is fever, which is the controlled increase of body temperature that occurs most commonly in response to an infection (Figure 3). Fever is triggered by bacterial lipids and other molecules (“pyrogens”) that directly or indirectly induce the production of prostaglandin E2 (PGE2) by endothelial cells lining the POA (Evans et al., 2015). PGE2 is thought to inhibit the activity of POA neurons that function to reduce body temperature, thereby producing a regulated hyperthermia that increases the likelihood of surviving an infection.
Figure 3. The generation of fever.
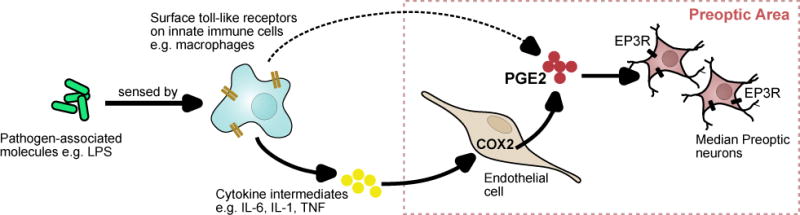
The presence of molecules associated with pathogens like bacteria and viruses is sensed by innate immune cells in the blood and lead to the production of pyrogenic intermediates like cytokines and prostaglandins that act on the preoptic area. In the preoptic area, COX2 expression in endothelial cells result in local PGE2 production, which is the dominant source of fever-inducing PGE2.
PGE2 acts through EP3 receptors expressed in the median preoptic (MnPO) to effect changes in body temperature. LPS – lipopolysaccharide; COX2 – cyclooxygenase 2.
Sleep is a second example of a physiologic process that modulates, and is modulated by, the thermoregulatory system (Krueger and Takahashi, 1997). The onset of sleep tracks closely the rate of decline in body temperature, and, during sleep, entry into epochs of rapid eye-movement (REM) is accompanied by near complete inhibition of thermoregulatory responses in many species (Krueger and Takahashi, 1997). Overlaid on these effects of sleep are slower timescale, diurnal fluctuations in body temperature that arise from circadian rhythms (Heller et al., 2010). Sleep, circadian rhythms, and body temperature are all controlled by dedicated neural circuits in the anterior hypothalamus, but the interconnections between these circuits have not been defined.
Thermoregulation is also tightly interconnected with the energy and fluid homeostasis systems, due to the substantial demands that thermoregulatory effectors place on bodily resources. For example, cold-induced thermogenesis consumes approximately 60% of total energy expenditure when mice are maintained at an ambient temperature of 4°C (Abreu-Vieira et al., 2015). To satisfy this energy need, mice exposed to cold will double their daily food intake (Bauwens et al., 2011) and, if supplied with adequate food, can live and proliferate at cold temperatures indefinitely. However when food is scarce, mice sacrifice the defense of body temperature and go into torpor, a state of regulated hypothermia and inactivity in which core body temperature can fall below 31°C (Webb et al., 1982). Similarly, in dehydrated animals, evaporative heat loss is attenuated in favor of thermoregulatory effectors that do not require water (Baker and Doris, 1982; Fortney et al., 1984; Morimoto, 1990). How these trade-offs between competing physiologic needs are resolved in the brain is an important open question (Nakamura et al., 2017).
2. Sources of input into the thermoregulatory system
The primary input into thermoregulatory system comes from sensory neurons that measure the temperature of the body. Most of these sensory neurons have cell bodies located in peripheral ganglia and axons that extend out to measure the temperature of key thermoregulatory tissues (e.g. the skin, spinal cord, and abdominal viscera; discussed below). A separate set of sensory neurons are located within the brain itself and measure the temperature of the hypothalamus.
Peripheral temperature sensing
Peripheral temperature sensing is mediated primarily by two classes of sensory neurons that are activated by innocuous warmth (~34-42°C) or col d (~14-30°C). These neurons have cell bodies located in trigeminal ganglion (for innervation of the head and face) and dorsal root ganglia (DRG; for innervation of the rest of the body). They are pseudounipolar, meaning that their axons split into two branches, one of which innervates the skin or viscera and the other projects to the dorsal horn of the spinal cord or to the spinal trigeminal nucleus in the brainstem (Figure 4).
Figure 4. Ascending neural pathways that transmit warm and cool signals from the periphery.
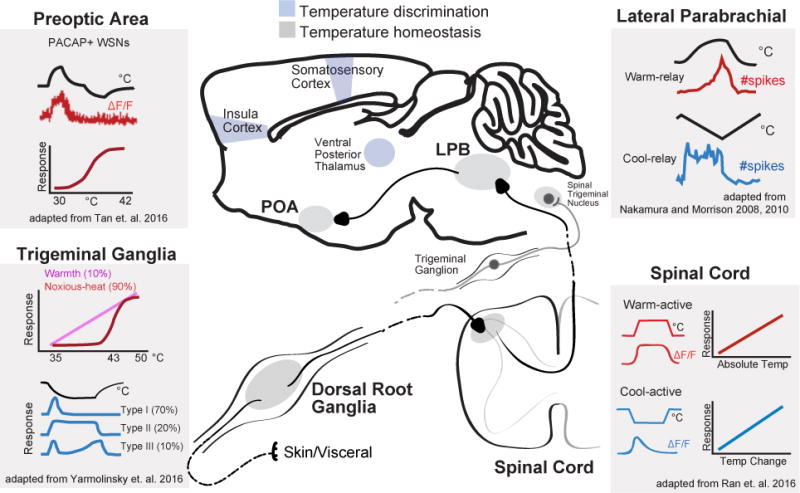
Structures involved in transmission of thermosensory input from the viscera and skin. Temperature information is sensed by neurons with cell bodies in primary sensory ganglia (or trigeminal ganglia), and then transmitted to the dorsal horn of the spinal cord (or chief sensory nucleus of V), the lateral parabrachial nuclei and finally the preoptic area. Brain regions involved in homeostatic control are shown in gray and those involved in temperature discrimination are shown in blue. Simplified schematics show the responses of neurons in this pathway to external heating and cooling. Sensory ganglia: adapted from (Yarmolinsky et al., 2016). Imaging of neural activity in the trigeminal ganglion shows that over 90% of thermal responsive cells responded to either heating or cooling with 2-5% of cells showing bimodal responses. Upper Line - typical normalized response over 35-50°C tem perature range for the 2 classes of heat sensitive neurons: warmth-sensing neurons with graded responses and broad dynamic range, and noxious heat-sensing neurons with high threshold and narrow dynamic range. Lower plot shows typical responses to cooling to 10°C for the 3 classes of cold-sensing neurons: Type 1 with tonic response to mild-cooling and rapid inactivation by noxious cold; Type 2 with sustained response to noxious cold; and Type 3 with a hybrid response. Spinal cord: adapted from Ran 2016. Imaging of neural activity in dorsal horn showed that cool-active neurons were rapidly adapting and responses scaled with the magnitude of temperature change. Warm-active neurons were non-adapting and responses reflect absolute target temperature. Broadly tuned neurons (not-shown) that responded to both cooling and heating were also present. Lateral parabrachial nucleus: Top – adapted from (Nakamura and Morrison, 2010). Single unit extracellular recording from warm-responsive neurons in the dorsal LPB that project to the preoptic area revealed that activity is increased by skin warming (14 out of 17 cells). Bottom – adapted from (Nakamura and Morrison, 2008). Single unit extracellular recording from cooling-responsive neurons in the external lateral LPB that project to the preoptic area showed that activity is induced in response to skin cooling (11 out of 14). Preoptic area: Adapted from (Tan et al., 2016). Population activity responses of warm-activated PACAP+ neurons in response to external temperature measured by fiber photometry. Preoptic PACAP neurons are progressive activated by increasing temperature from 30 to 42°C. They show no further activity increase in response to noxious heat (>42°C) or activity decrease in response to cold (<30°C).
Peripheral thermosensation has been comprehensively reviewed elsewhere (Ma, 2010; Vriens et al., 2014). Here we briefly outline the key facts relevant to thermoregulation.
Cold-sensing
TRPM8 is the primary peripheral cold sensor in the thermoregulatory system. This channel is activated in vitro by mild cooling (<26-28° C) and its expression is required for cold perception (Bautista et al., 2007; Dhaka et al., 2007; McKemy et al., 2002; Peier et al., 2002). TRPM8 is expressed in essentially all cold-sensitive neurons, and ablation of these TRPM8+ cells abolishes the behavioral and neural responses to cooling (Knowlton et al., 2013; Pogorzala et al., 2013; Yarmolinsky et al., 2016). As would be expected for thermosensor with a role in thermoregulation, treatment with TRPM8 agonists causes hyperthermia, whereas TRPM8 antagonists cause hypothermia (Almeida et al., 2012; Gavva et al., 2012). In addition, TRPM8 antagonists block the ability of environmental cold to induce Fos in brain regions that mediate thermoregulation (Almeida et al., 2012). Thus TRPM8 and the neurons that express it delineate a labelled line that communicates cold information from the periphery into the CNS.
Warm-sensing
The molecular identity of the peripheral warm sensor is controversial. Several TRP channels have been proposed to play this role, including TRPV1, TRPV3, TRPV4, and TRPM2, but there is conflicting evidence for and against all of these candidates (summarized in Table 1). At the level of the sensory neurons, the cells that mediate warm-sensing are a subset of TRPV1+ primary afferents. Treatment of these cells with a TRPV1 antagonist blocks their activation in vivo by innocuous warmth (Yarmolinsky et al., 2016), which is counterintuitive given that TRPV1 is activated in vitro only at higher temperatures (>42° C). However this may reflect the presence of co-agonists or post-translational modifications that can lower the TRPV1 temperature threshold in vivo (Tominaga et al., 1998; Vellani et al., 2001). Peripheral TRPV1 antagonists induce hyperthermia, whereas TRPV1 agonists induce hypothermia, consistent with a role in thermoregulation ((Gavva, 2008; Gavva et al., 2007; Hori, 1984; Steiner et al., 2007) but see counterarguments in (Romanovsky et al., 2009) and Table 1). Nevertheless the fact that TRPV1 knockout mice have normal body temperature indicates that, at a minimum, other channels can compensate for its thermoregulatory function (Caterina et al., 2000; Iida et al., 2005; Szelenyi et al., 2004). Knockout of other TRP channels, alone or in combination, has yielded inconsistent effects in some cases and in others the thermoregulatory phenotype has not been fully characterized (Huang et al., 2011; Song et al., 2016; Tan and McNaughton, 2016; Vriens et al., 2014).
Table 1.
| Gene | Location | Evidence For | References | Evidence Against | References |
|---|---|---|---|---|---|
| TRPV1 | Peripheral | Peripheral TRPV1 agonists induce hypothermia and activate preoptic warm-sensitive neurons | (Hori, 1984; Nakayama et al., 1978; Tan et al., 2016) | Some effects of peripheral TRPV1 agonists may be centrally mediated | (Hori, 1984; Romanovsky et al., 2009) |
| Peripheral TRPV1 antagonists induce hyperthermia | (Gavva, 2008; Gavva et al., 2007) | TRPV1 antagonist induced hyperthermia is independent of temperature | (Romanovsky et al., 2009; Steiner et al., 2007) | ||
| TRPV1 can be activated by warm temperatures following sensitization by endogeneous co-agonists | (Cao et al., 2013; Tominaga et al., 1998; Vellani et al., 2001) | Temperature threshold of purified TRPV1 (~42 C) is above the range of innocuous warmth | (Cao et al., 2013; Caterina et al., 1997) | ||
| TRPV1 antagonists block the activation of sensory neurons by innocuous warmth in vivo | (Yarmolinsky et al., 2016) | TRPV1 knockout or ablation of TRPV1 + neurons has little or no effect on body temperature or thermoregulation | (Caterina et al., 2000; Garami et al., 2011; Iida et al., 2005; Pogorzala et al., 2013) | ||
| TRPV1 | Central | Central capsaicin can induce hypothermia | (Hori, 1984) | TRPV1 expression is extremely sparse in the brain and absent from preoptic area | (Cavanaugh et al., 2011) |
| The requirement for TRPV1 in the response to brain warming or central capsaicin has not been reported | |||||
| TRPM2 | Central or peripheral | TRPM2 KO diminishes the activation of neurons by warming in hypothalamic slices or DRG cultures | (Song et al., 2016; Tan and McNaughton, 2016) | TRPM2 is broadly expressed in the brain and periphery and does not specifically label thermoregulatory cells | (Song et al., 2016; Tan and McNaughton, 2016) |
| TRPM2 KO mice have a mildly elevated fever in response to PGE2 and show reduced preference for warm temperatures | (Song et al., 2016; Tan and McNaughton, 2016) | TRPM2 KO mice have normal core body temperature, and the response to brain warming and other thermoregulatory challenges has not been reported | |||
| TRPV3 | Peripheral | Heterologous TRPV3 is activated by innocuous warmth (threshold ~32 C), | (Peier et al., 2002; Smith et al., 2002; Xu et al., 2002) | The thermosensory deficits in TRPV3 KO mice are strain and sex-dependent | (Huang et al., 2011; Miyamoto et al., 2011) |
| TRPV3 KO mice have impaired thermosensation | (Moqrich et al., 2005) | TRPV3/TRPV4 double KO mice have normal thermosensation | (Huang et al., 2011) | ||
| TRPV4 | Peripheral | Heterologous TRPV4 is activated by innocuous warmth (threshold ~34 C) | (Guler et al., 2002) | TRPV4 KO mice and TRPV3/TRPV4 double KO mice have normal thermoregulation | (Huang et al., 2011; Liedtke and Friedman, 2003) |
| TRPV4 KO mice have impaired thermosensation and a peripheral TRPV4 antagonist increases body temperature | (Lee et al., 2005; Vizin et al., 2015) |
Tissues that provide thermoregulatory input
The relative contribution of different tissues to the overall body temperature signal has been investigated by manipulating their temperature and then measuring the thermoregulatory response (Figure 5). This has identified four tissues that provide particularly important input: the skin, spinal cord, abdominal viscera, and brain (Cabanac, 1975; Jessen, 1985). In general, the POA is the most thermosensitive site (i.e. largest effector response per degree of warming or cooling), whereas the skin undergoes the largest temperature fluctuations. These inputs from different tissues are summed to determine the magnitude of the thermoregulatory response; this summation can be simply additive or more complex, depending on the context (Figure 5).
Figure 5. Interaction between core (brain or spinal cord) and ambient (skin) temperature in the control of thermoregulatory effectors.
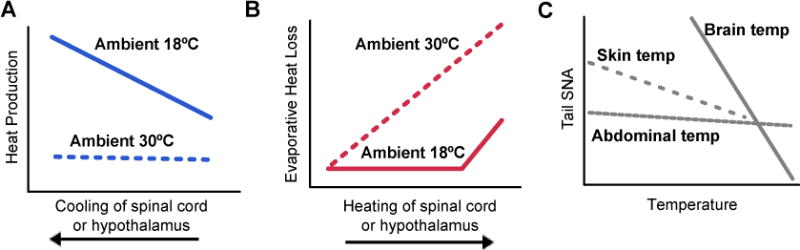
A and B. Adapted from (Jessen and Ludwig, 1971). Spinal cord (SC) and hypothalamic (Hypo) temperatures in the dog were independently manipulated at varying ambient temperatures and resultant effects of heat production and evaporative heat loss are shown.
C. Adapted from (Shafton et al., 2014). Changes in rat tail sympathetic nerve activity (SNA), which is a measure of vasoconstriction (low SNA – vasodilation) as abdominal, skin or brain temperature is altered.
Skin temperature functions both as an input that activates thermoregulatory effectors (e.g. shivering when the air is cold) and as a discriminative signal that guides behavior (e.g. this object is warm). For this reason different parts of the skin contribute to thermoregulation in different ways. Non-hairy (glabrous) skin in mammals is restricted to a few sites, such as parts of the hands, feet, and face, that are more important for discriminating the temperature of external objects. Hairy skin covers the majority of the body and, due to this larger surface area, contributes relatively more of the input signal that drives thermoregulatory effectors (Romanovsky, 2014). However there are exceptions to this rule. For example, heating of the face (Nadel et al., 1973) or the scrotum (Waites, 1962) drives panting and sweating to a greater extent than would be predicted based on their surface area, whereas heating of the extremities (e.g. arms and legs) has proportionally less effect.
Outside of the brain, the spinal cord is the most well characterized contributor to the core body temperature signal, and numerous studies have shown that selectively heating or cooling the spinal cord can trigger appropriate thermoregulatory responses (Figure 5) (Cabanac, 1975; Jessen, 1985). The thermosensitivity of the spinal cord is thought to be mediated by the same sensory neurons that measure the temperature of the skin and viscera (Brock and McAllen, 2016). This is possible because the axons of these primary sensory afferents terminate in the dorsal horn of the spinal cord, such that heating or cooling of the spinal cord potentiates neurotransmitter release from their thermosensitive terminals.
Temperature sensing in the brain
In addition to peripheral tissues, the temperature of the brain itself is an input into the thermoregulatory system (Figure 5). The most sensitive site in the brain is a hotspot in the midline POA, located between the anterior commissure and optic chiasm, that when heated elicits dramatic and coordinated heat defensive responses such as panting, sweating, vasodilation, and cold-seeking behavior (Andersson et al., 1956; Carlisle, 1966; Carlisle and Laudenslager, 1979; Hemingway et al., 1954; Magoun et al., 1938). Cooling of this structure has the opposite effect, promoting vasoconstriction, BAT thermogenesis, shivering, and operant responses for heat (Hammel et al., 1960). These observations suggest that the midline POA contains intrinsically thermosensitive neurons that are important for body temperature control.
Brain temperature can increase by 2-3°C in response to exercise or fever, which provides a context in which POA warm-sensing may be important (Figure 1) (Fuller et al., 1998; Walters et al., 2000). On the other hand, acute exposure to environmental heat or cold does not affect brain temperature in most animals (although there are exceptions, Figure 1) (Bratincsak and Palkovits, 2005; Hammel, 1968; Hammel et al., 1963; Hellstrom and Hammel, 1967; Nakamura and Morrison, 2008, 2010). In addition to sensing local brain temperature, POA neurons also receive information about peripheral temperature via an ascending neural pathway (Figure 4), and 25-50% of the POA neurons that are activated by local brain warming are also activated by warming of the skin or spinal cord (Boulant and Hardy, 1974; Wit and Wang, 1968). Thus many POA cells integrate central and peripheral thermal information.
While it is likely that intrinsically thermosensitive POA neurons play a role in thermoregulation, it is important to emphasize that thermosensitive neurons are found in many brain regions, and most of these neurons presumably have no role in body temperature regulation (Barker and Carpenter, 1970; Eisenman et al., 1971; Hellon, 1986). For example, cooling of the HVC, a premotor nucleus in the zebra finch, can selectively slow the bird’s song speed, even though songbird singing is not naturally controlled by changes in brain temperature (Long and Fee, 2008). To definitively establish the physiologic relevance of intrinsic POA thermosensing, it will likely be necessary to identify and disrupt the brain temperature sensor.
Molecules that sense brain temperature
If warm-sensitivity is defined as an increase in spontaneous firing rate of >0.8 action potentials per second per degree Celsius, then approximately 20% of POA neurons are warm-sensitive in vitro (Boulant, 2006; Nakayama et al., 1978) or in vivo (Hellon, 1970; Knox et al., 1973; Nakayama et al., 1963). This warm-sensitivity is an intrinsic property of POA neurons (Kelso and Boulant, 1982) and has been proposed to be mediated by either a heat-activated non-selective cation current (Kobayashi et al., 2006) or a heat-inactivated potassium current (Boulant, 2006). In neither case has the identity of the relevant ion channel been established.
Two TRP channels, TRPV1 and TRPM2, have been proposed to function as warm-sensors in the brain (Table 1). The case for TRPV1 is based on the fact that central injection of the TRPV1 agonist capsaicin can induce hypothermia (Hori, 1984; Romanovsky et al., 2009). However the site of action of capsaicin in these experiments is unclear, since TRPV1 is expressed at extremely low levels in the brain and is absent from the POA (Cavanaugh et al., 2011). The evidence supporting a role for TRPM2 includes the fact that TRPM2 knockout mice have an exacerbated fever response, and the fact that neurons from these knockout mice have attenuated thermosensitivity in vitro (Song et al., 2016). However TRPM2 KO mice have normal core body temperature, and TRPM2 is broadly expressed in the brain and periphery (Song et al., 2016; Tan and McNaughton, 2016), suggesting that TRPM2 is unlikely to be the molecule that confers warm-sensitivity on a specific subset of neurons. For both of these candidates, a critical test will be to measure whether deletion of the channel in the brain can abrogate the thermoregulatory response to POA warming.
3. Afferent pathways from the periphery to the POA
The POA receives ascending signals from thermoreceptors in the skin, viscera, and spinal cord, which are then presumably integrated with information about the temperature of the brain in order to enact thermoregulatory responses (Jessen, 1985; Vriens et al., 2014). Temperature information from sensory afferents that innervate the skin and viscera is transmitted to the POA by a neural pathway with relays in spinal cord and parabrachial nucleus (Figure 4). A separate pathway involving vagal afferents may also contribute, but this is less well characterized.
Coding of temperature in the spinal cord
Warm and cold-sensitive sensory neurons innervate superficial laminae of the dorsal horn, where they synapse on spinal cord projection neurons (Figure 4). Electrophysiological recordings of spinal neurons have demonstrated the existence of distinct neuronal populations that respond to warmth and cold, as well as polymodal cells involved that respond to temperatures in the noxious range (Ma, 2010). Recently, this work has been extended to include population-level responses by in vivo calcium imaging in spinal cord (Ran et al., 2016). This has revealed that cold responsive dorsal horn neurons encode primarily temperature change and are rapidly adapting, whereas heat responsive spinal neurons encode absolute temperature and are non-adapting (Ran et al., 2016). Selective ablation of TRPV1 or TRPM8 expressing sensory afferents confirmed that the spinal response to mild cooling was mediated by TRPM8+ cells, whereas TRPV1+ cells drove spinal responses to noxious heat and a combination of TRPV1+ and TRPM8+ inputs were involved in the representation of innocuous warmth (Ran et al., 2016). Of note, combined ablation of all TRPV1+ and TRPM8+ neurons also abolishes the behavioral responses to temperature between 10 and 50°C, indicating that these two subsets together define the necessary set of thermoreceptors (Pogorzala et al., 2013).
Segregated warm and cold relays in the lateral parabrachial nucleus
Dorsal horn neurons send glutamatergic projections to the brain that collateralize to the thalamus and lateral parabrachial nucleus (LPB; Figure 4) (Hylden et al., 1989). Thermal information received in thalamus is relayed to somatosensory cortex, where it mediates the perception and discrimination of temperature (Craig et al., 1994). However thalamic lesions do not block behavioral or autonomic thermoregulatory responses (Nakamura and Morrison, 2008; Yahiro et al., 2017), suggesting that the spinothalamocortical pathway is dispensable for body temperature regulation in some contexts. In contrast, lesioning or silencing of the LPB abolishes the autonomic responses to skin warming and cooling as well as temperature preference in a behavioral assay (Kobayashi and Osaka, 2003; Nakamura and Morrison, 2008, 2010; Yahiro et al., 2017). Thus ascending input to the LPB, which in turn is relayed to the POA, is critical for the activation of thermoregulatory responses to environmental temperature.
The LPB has several subdivisions, each of which contains a mixture of cell types implicated in different aspects of homeostasis including feeding, salt appetite, thirst, and cardiovascular function, in addition to thermoregulation (Davern, 2014). Ascending cold and warm signals terminate in two anatomically distinct subdivisions of the LPB: the external lateral LPB (LPBel, cold) and the dorsal LPB (LPBd, warm). Consistent with this anatomic segregation, cold and warm challenge induce Fos expression primarily in the LPBel and LPBd, respectively (Bratincsak and Palkovits, 2004; Geerling et al., 2016; Nakamura and Morrison, 2008, 2010). Warmth-activated LPBd neurons express the neuropeptide dynorphin, whereas cold-activated LPBel neurons are a subset of a larger Foxp2+ population that does not express dynorphin (Geerling et al., 2016). Single-unit recordings from antidromically identified LPB → POA neurons have demonstrated that LPBd neurons are activated by skin warming from 34 to 38°C, whereas LPBel neurons are activated by skin cooling across the same range (Figure 4) (Nakamura and Morrison, 2008, 2010).
Warm and cold-activated LPB neurons send dense glutamatergic projections to the midline POA and particularly the MnPO (Geerling et al., 2016; Nakamura and Morrison, 2008, 2010). This direct projection is likely to be an important pathway by which thermal information received in the LPB is transmitted to the POA, but the connectivity between specific LPB and POA cell types has not been established.
4. Thermoregulatory neurons in the preoptic hypothalamus
The POA is thought to be the key integratory site for thermoregulation in the brain. This is supported by many lines of evidence, including: (1) POA lesioning or pharmacologic silencing results in animals that cannot defend their core body temperature in either a hot or cold environment (Ishiwata et al., 2005; Lipton, 1968; Osaka, 2004; Satinoff et al., 1976; Van Zoeren and Stricker, 1976), (2) local warming of the POA causes hypothermia and heat-defensive responses that mirror the response to environmental heat (Andersson et al., 1956; Carlisle, 1966; Carlisle and Laudenslager, 1979; Hemingway et al., 1954; Magoun et al., 1938), (3) injection of the pyrogen PGE2 into the POA causes fever (Elmquist et al., 1996; Scammell et al., 1996), (4) the POA contains neurons that are selectively activated in vivo by warming of the skin, spinal cord, or brain (Hellon, 1970; Knox et al., 1973; Nakayama et al., 1961; Nakayama et al., 1963; Tan et al., 2016), and (5) the POA is densely connected to brain regions that receive thermal information from the periphery as well as structures that control thermoregulatory effectors.
Although the POA is critical for thermoregulation, it is also associated with many other functions, including the regulation of fluid balance, sleep, mating, and parental behaviors. These functions are likely mediated by distinct cell types, which raises the question of which cell types in the POA are specifically involved in regulation of body temperature. Recent work has begun to investigate this question by using genetic approaches for neural manipulation and recording.
Genetic identification of thermoregulatory neurons in the POA
Exposure to a warm environment induces Fos expression in a medial region of the POA that includes the VMPO and MnPO (Bachtell et al., 2003; Bratincsak and Palkovits, 2004; Harikai et al., 2003; Scammell et al., 1993; Yoshida et al., 2005). This activated region overlaps with the area where local warming is most effective at inducing thermoregulatory responses (Magoun et al., 1938), and where the pyrogen PGE2 acts to induce fever (Elmquist et al., 1996; Scammell et al., 1996). Thus these Fos+ cells are likely to be important for thermoregulation.
To identify these cells, an unbiased approach for molecular profiling of activated neurons (Knight et al., 2012) was used to capture and sequence mRNA from POA neurons activated by warmth (Tan et al., 2016). This revealed that exposure of mice to ambient warmth (37°C) activates a specific subpopulation of POA neurons identified by co-expression of the neuropeptides PACAP and BDNF (Tan et al., 2016). Optical recordings of the activity of these POAPACAP/BDNF neurons in awake, behaving mice demonstrated that they are rapidly (seconds) and progressively activated when mice are exposed to ambient warmth (~30-42°C; Figure 4) or challenged with peripheral injection of capsaicin, a TRPV1 agonist. In contrast, these neurons are unresponsive to cold temperatures, the TRPM8 agonist icilin, or several non-thermal stimuli. Consistent with this selective regulation by warmth, optical stimulation of these neurons induced hypothermia mediated by combination of autonomic and behavioral mechanisms (Table 2) (Tan et al., 2016). Thus POAPACAP/BDNF neurons are activated by environmental warmth, and their activity is sufficient to drive the coordinated homeostatic response to heat.
Table 2.
Optogenetic and Chemogenetic Manipulations
| POA Region | Cell Type Marker | Experimental Manipulation | Neural Activity | Core Temperature | Tail Vasodilation | BAT Temperature | Behavior Change | Reference |
|---|---|---|---|---|---|---|---|---|
| POA | TRPM2 | hM3Dq | ↑ | ↓ | ↑ | ↓ | ND | Song et al., 2016 |
| POA | TRPM2 | hM4Di | ↓ | ↑ | NS | NS | ND | Song et al., 2016 |
| POA | Vglut2 | hM3Dq | ↑ | ↓ | ND | ND | ND | Song et al., 2016 |
| POA | Vgat | hM3Dq | ↑ | NS | ND | ND | ND | Song et al., 2016 |
| vLPO | Vgat | ChR | ↑ | ↓ | ND | ND | ↓ activity | Zhao et al., 2017 |
| vLPO | Vgat | hGtACR1 | ↓ | ↑ | ND | ND | ↑ activity | Zhao et al., 2017 |
| vLPO | Vglut2 | ChR | ↑ | ↓ | ND | ND | ↓ activity | Zhao et al., 2017 |
| vLPO | Vglut2 | hM3Dq | ↑ | ↓ | ND | ND | ND | Zhao et al., 2017 |
| MPO | Vgat | ChR | ↑ | NS | ND | ND | no change in activity | Zhao et al., 2017 |
| MnPO,VMPO | LepRb | hM3Dq | ↑ | ↓ | ND | ND | ↓ activity,↑ postural extension | Yu et al., 2016 |
| MnPO,VMPO | Vglut2 | hM3Dq | ↑ | ↓ | ND | ND | NS activity | Yu et al., 2016 |
| MnPO,VMPO | Vgat | hM3Dq | ↑ | NS | ND | ND | NS activity | Yu et al., 2016 |
| MnPO,VMPO | PACAP | SSFO | ↑ | ↓ | ↑ | ↓ | ↑cold seeking,↓ nest building | Tan et al., 2016 |
| MnPO,VMPO | BDNF | SSFO | ↑ | ↓ | ↑ | ↓ | ↑cold seeking,↓ nest building | Tan et al., 2016 |
| MnPO | Vglut2 | ChR | ↑ | ↓ | ↑ | ND | ↑ drinking in subset | Abbott and Saper, 2017 |
| DMH | ChAT | ChR | ↑ | ↓ | ND | ↓ | ND | Jeong et al., 2015 |
| DMH | ChAT | ArCH | ↓ | ↑ | ND | ↑ | ND | Jeong et al., 2015 |
| DMH | LepRb | hM3Dq | ↑ | ↑ | ND | ↑ | ↑ activity | Rezai-Zadeh et al., 2014 |
| DMH | Vglut2 | ChR | ↑ | ↑ | ND | ↑ | ND | Tan et al., 2016 |
| DMH | Vglut2 | hM3Dq | ↑ | ↑ | ND | ↑ | ND | Tan et al., 2016 |
| DMH | Vglut2 | hM3Dq | ↑ | ↑ | ND | ND | ↑ activity | Zhao et al., 2017 |
| DMH | Vglut2 | hGtACR1 | ↓ | ↓ | ND | ND | ↓ activity | Zhao et al., 2017 |
| DMH | Vgat | ChR | ↑ | ↑ | ND | ND | ↑ activity | Zhao et al., 2017 |
| DMH | Vgat | hM3Dq | ↑ | ↑ | ND | ND | ↑ activity | Zhao et al., 2017 |
| DMH | Vgat | hGtACR1 | ↓ | ↓ | ND | ND | ↓ activity | Zhao et al., 2017 |
| POA→DMH | POAPACAP | ChR | ↑ | ↓ | NS | ↓ | no change in cold seeking | Tan et al., 2016 |
| POA→DMH | VLPOVGAT | ChR | ↑ | ↓ | ND | ND | ↓ activity | Zhao et al., 2017 |
| DMH→RMR | non-specific | ChR | ↑ | NS | ND | ↑ | ND | Kataoka et al., 2014 |
Warmth-activated neurons: Glutamatergic or GABAergic?
Approximately two-thirds of POAPACAP/BDNF neurons express GAD2, suggesting that these cells are mostly GABAergic (Tan et al., 2016). This finding is consistent with traditional models for the thermoregulatory circuit, which posit that warmth-activated POA neurons are GABAergic and tonically inhibit downstream structures that drive thermogenesis (Morrison and Nakamura, 2011).
However, in apparent contradiction to this model, two recent studies reported that optogenetic or chemogenetic stimulation of GABAergic (Vgat+) cells in the medial POA had no effect on body temperature (Song et al., 2016; Yu et al., 2016). Instead, stimulation of glutamatergic (Vglut2+) POA cells induced hypothermia, via a combination of increased tail vasodilation and reduced thermogenesis and locomotor activity (Table 2) (Abbott and Saper, 2017; Song et al., 2016; Yu et al., 2016). These glutamatergic cells are partially overlapping with neurons that express the leptin receptor (LepR), and stimulation of POALepR cells was also sufficient to induce hypothermia (Yu et al., 2016). Given that the hormone leptin is important for the regulation of energy expenditure, these POALepR cells may play a role in linking body temperature to changes in nutritional state (Yu et al., 2016; Zhang et al., 2011).
The lack of theremogulatory response to stimulation of GABAergic POA cells is puzzling. One possibility is that there are subsets of GABAergic POA neurons that have different roles in thermoregulation, and the observed phenotype depends on precisely which combinations of neurons are stimulated. Consistent with this, optogenetic activation of a subset of GABAergic cells localized to the ventrolateral preoptic area (vlPOA) has been shown to induce hypothermia through a specific projection to the DMH, as predicted by earlier models (Table 2) (Zhao et al., 2017).
Alternatively, it is possible that warmth-activated POAPACAP/BDNF neurons do not regulate body temperature through release of GABA. In this regard, although most POAPACAP/BDNF neurons express GAD2, many also express Vglut2 (Tan et al., 2016). In most brain regions these markers are mutually exclusive, but recent reports have described hypothalamic neurons that co-express GAD2 with Vglut2, rather than Vgat (Leib et al., 2017; Romanov et al., 2017). It will be important to clarify this issue by directly recording the post-synaptic currents induced by terminal stimulation of POA warmth-activated neurons innervating different targets.
Efferent pathways from the POA to thermoregulatory effectors
Thermal information received in the POA is communicated to downstream structures that control physiologic and behavioral effectors (Figure 6). Here we briefly outline what is known about the neural mechanisms and pathways that control each of these responses.
Figure 6. Descending circuits controlling thermoregulatory effectors.
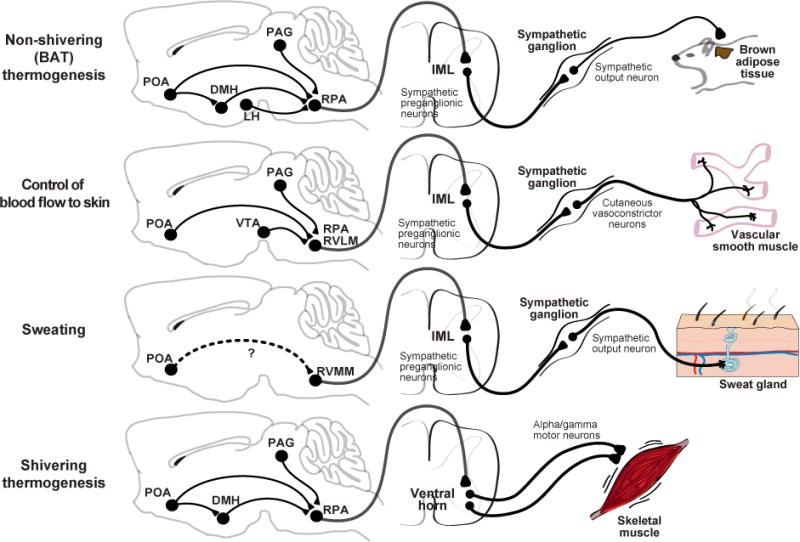
The CNS/PNS regions involved invarious thermoregulatory effector responses and the proposed descending pathway from the POA to motor output. Note that many of the connections in the brain that are drawn are postulated based on indirect evidence. Dashed arrows indicate that a functional connection exists, but that the anatomic pathway is unknown and may involve multiple synapses and additional brain regions. POA – preoptic area, DMH – dorsomedial hypothalamus, LH – lateral hypothalamus, PAG – periaqueductal gray, VTA – ventral tegmental area, RMR – raphe medullary region, RPA – raphe pallidus, RVLM – rostral ventrolateral medulla, RVMM – rostral ventromedial medulla, IML – interomediolateral column, SSN – superior salivary nucleus.
A. Control of physiologic responses
Physiologic effectors are involuntary responses that generate or dissipate heat. Four physiologic effectors are particularly important for thermoregulation in mammals: BAT thermogenesis, control of skin blood flow, shivering, and evaporative cooling. While the central circuits that control each of these responses is distinct, they are thought to share a common organization in which thermal information is received and integrated in the POA and then transmitted to effectors through a descending pathway that exits the brain via the rostral medulla (Figure 6). These medullary output neurons then activate peripheral sympathetic or parasympathetic circuits, or in the case of shivering, somatic motor neurons, that induce the physiologic response.
BAT Thermogenesis
BAT is a specialized organ for the rapid production of heat. In mice, BAT is found most prominently in the interscapular region, where it is highly innervated by sympathetic nerves. Release of norepinephrine from this sympathetic innervation induces mitochondrial leak in BAT that produces heat (known as non-shivering or BAT thermogenesis). The mechanisms that control BAT thermogenesis have been comprehensively reviewed elsewhere (Morrison et al., 2012; Morrison and Nakamura, 2011). Here we briefly summarize key features of the efferent circuit.
The rostral raphe pallidus (rRPA) is the primary site where descending signals driving BAT thermogenesis exit the brain (Figure 6). Exposure to cold or pyrogens activates premotor neurons in the rRPA that project to the spinal cord, many of which express either the glutamate transporter (Vglut3) or serotonin (Morrison et al., 1999; Nakamura et al., 2004; Nakamura et al., 2002). These rRPA projections activate preganglionic neurons in the intermediolateral (IML) nucleus of the spinal cord that in turn regulate sympathetic outflow and thereby BAT activity (Figure 6). Chemical stimulation of the rRPA is sufficient to induce BAT thermogenesis (Morrison et al., 1999), whereas chemical inhibition of the rRPA blocks BAT thermogenesis induced by skin cooling, central PGE2, or stress (Kataoka et al., 2014; Madden and Morrison, 2003; Morrison, 2003; Nakamura et al., 2002; Nakamura and Morrison, 2007). Thus the rRPA is a common final output of multiple thermogenic pathways.
The rRPA receives extensive innervation from the DMH (Hermann et al., 1997; Hosoya et al., 1989). Chemical stimulation of the DMH increases BAT thermogenesis (Zaretskaia et al., 2002), which can be blocked by inhibition of rRPA (Cao et al., 2004; Cao and Morrison, 2006), whereas silencing of the DMH attenuates BAT activity (Madden and Morrison, 2004). Skin cooling, PGE2, and stress activate DMH neurons that project to the rRPA (Kataoka et al., 2014; Yoshida et al., 2009). Optogenetic stimulation of the direct DMH → rRPA projection elicits BAT thermogenesis (Kataoka et al., 2014) but the precise identity and connectivity of these neurons remains unclear. Cold activates both GABAergic and glutamatergic DMH neurons in vivo (Zhao et al., 2017), including a subpopulation that co-expresses the leptin receptor (LepR) and prolactin-releasing peptide (PrRP) (Dodd et al., 2014; Zhang et al., 2011). BAT temperature is increased by stimulation of glutamatergic or LepR neurons in the DMH (Table 2) (Rezai-Zadeh et al., 2014; Tan et al., 2016). Conversely, cholinergic neurons in DMH are activated by warmth, project to rRPA, and inhibit BAT thermogenesis in the mouse (Jeong et al., 2015), although this was not observed in the rat (Conceicao et al., 2017). Given that “glutamatergic” and “GABAergic” DMH neurons are heterogeneous populations that contain many different cell types, it will be necessary to identify better molecular markers in order to dissect the efferent circuitry.
The POA provides a major thermoregulatory input to the DMH (Figure 6). The traditional model has been that a GABAergic POA→DMH projection supplies tonic input that inhibits the DMH and suppresses thermogenesis (Morrison and Nakamura, 2011). This is supported by the observation that knife cuts that separate the POA from the DMH increase BAT thermogenesis (Chen et al., 1998). The thermoregulatory effect of this loss of POA→DMH input can be replicated by pharmacologic activation of the DMH (Cao et al., 2004; Morrison et al., 1999; Zaretskaia et al., 2002) and blocked by pharmacologic inhibition of the DMH (Nakamura et al., 2005). GABAergic neurons in the mPOA that express the EP3 receptor (which binds to PGE2 to stimulate fever) have been reported to densely innervate the DMH (Nakamura et al., 2005; Nakamura et al., 2009).
Consistent with this model, warmth-activated POAPACAP/BDNF neurons that innervate the DMH express GAD2, and optogenetic stimulation of this POA→DMH projection suppresses BAT thermogenesis (Tan et al., 2016); however, as described above, whether GABA is released by these cells has not been measured directly. Body temperature is also reduced by stimulation of glutamatergic POA neurons (either POAVGLUT2 or POALepR neurons, which are 60% glutamatergic) (Yu et al., 2016) and POALepR neurons are known to project to the DMH. However it is unknown whether their effects on body temperature are due to direct modulation of BAT thermogenesis via neurons in the DMH or via other mechanisms.
In addition to this canonical POA → DMH → rRPA pathway, there are several other routes by which thermoregulatory signals may be communicated to rRPA. POA neurons expressing EP3R or LepR send direct projections to the rRPA that may contribute to the control of BAT thermogenesis (Nakamura et al., 2009; Yoshida et al., 2009; Zhang et al., 2011). There is likewise a pathway between the DMH, caudal periaqueductal gray (cPAG), and rRPA that has been proposed to regulate BAT thermogenesis (Chen et al., 2002; Yoshida et al., 2005). Finally, orexin neurons in the lateral hypothalamus innervate the rRPA, and this pathway is important for BAT thermogenesis induced by cold, PGE2, and stress (Takahashi et al., 2013; Tupone et al., 2011; Zhang et al., 2010). How orexin neurons connect to upstream elements of the thermoregulatory circuit (e.g. the POA) is not well defined.
Skin blood flow
The rate of heat exchange between the skin and environment depends on blood flow to the skin. Decreasing skin blood flow by cutaneous vasoconstriction is a thermoregulatory mechanism for preventing heat loss, whereas increasing blood flow (cutaneous vasodilation) has the opposite effect. In rodents, these vasomotor responses are controlled primarily by the release of norepinephrine from sympathetic fibers innervating vascular smooth muscle in the skin, which promotes vasoconstriction (Ootsuka and Tanaka, 2015). The effects of vasomotion are particularly prominent in the rodent tail, which can undergo large temperature fluctuations in order to increase or decrease heat loss.
The rRPA and adjacent rostral ventrolateral medulla (RVLM) contain the sympathetic premotor neurons that are critical for the regulation of cutaneous vasomotor responses (Figure 6). Excitation of the rRPA increases vasoconstriction and decreases tail skin temperature (Blessing and Nalivaiko, 2001; Rathner and McAllen, 1999; Tanaka et al., 2002), whereas inhibition of the rRPA blocks the vasoconstriction caused by cold (Blessing and Nalivaiko, 2001; Ootsuka et al., 2004; Ootsuka and McAllen, 2005). Similar but smaller responses are observed following bidirectional manipulation of the RVLM (Key and Wigfield, 1994; Ootsuka and McAllen, 2005; Rathner et al., 2008; Tanaka et al., 2002).
The POA is a major regulator of cutaneous vasomotor responses. Inhibition or cooling of the POA induces vasoconstriction (Osborne and Kurosawa, 1994), whereas excitation or warming induces vasodilation (Carlisle and Laudenslager, 1979; Tanaka et al., 2002). These effects are mediated in part by POAPACAP/BDNF neurons, which drive vasodilation when optically stimulated (Tan et al., 2016). Drug microinjection studies suggest there is a second population located in the caudolateral POA also regulates vasomotor responses. (Tanaka et al., 2009, 2011).
The vasomotor responses to POA manipulations require the rRPA (Tanaka et al., 2013; Tanaka et al., 2002), and are likely mediated by both direct POA → rRPA projections (Nakamura et al., 2009; Tanaka et al., 2011; Yoshida et al., 2009) as well as indirect pathways. Activation of the DMH is sufficient to induce vasoconstriction but, unlike BAT thermogenesis, the DMH is not required for the vasomotor response to POA cooling (Rathner et al., 2008) and stimulation of POAPACAP/BDNF neuron terminals in the DMH fails to induce tail vasodilation (Tan et al., 2016). Instead, two other brain regions, the ventral tegmental area (VTA) and rostroventrolateral PAG, have been proposed to be part of the downstream circuit (Ootsuka and Tanaka, 2015). How these two brain regions are functionally connected to the POA and rRPa is unknown (Figure 6).
Shivering
Shivering is the fast, repetitive contraction of skeletal muscle to generate heat that is triggered by cold exposure or fever (“chills”). The regulation of shivering involves a similar set of structures to those that regulate other physiologic responses, including the LPB, POA, DMH, and rRPA (Figure 6). Shivering induced by skin cooling is blocked by inhibition of the ascending cutaneous pathway in the LPB (Nakamura and Morrison, 2008) and its target in the MnPO (Nakamura and Morrison, 2011). Direct cooling of the POA facilitates shivering (Andersen et al., 1962; Hammel et al., 1960), whereas POA warming or stimulation blocks shivering induced by environmental cold (Hemingway et al., 1954; Kanosue et al., 1991; Zhang et al., 1995). Consistent with a model in which tonic, descending inhibition from warmth-activated neurons in the POA suppresses shivering, shivering is increased by stimulation of the DMH, rRPA, and adjacent structures (Nakamura and Morrison, 2011; Nason and Mason, 2004; Stuart et al., 1961) whereas inhibition or lesioning of those sites blocks shivering induced by cold and PGE2 (Brown et al., 2008; Nakamura and Morrison, 2011; Stuart et al., 1962; Tanaka et al., 2006; Tanaka et al., 2001). The circuitry that connects these and other structures to control shivering is unknown, as is the precise pathway that leads to motor neuron activation. However retrograde tracing has shown that the rRPA and adjacent structures provide polysynaptic input into skeletal muscle, suggesting that they may serve as a common output (Kerman et al., 2003).
Evaporative heat loss
Water evaporation is a thermoregulatory strategy for dissipating heat. Evaporative cooling in humans is achieved primarily by sweating, whereas most non-primates rely on panting but may sweat in restricted locations, such as the footpad of the cat (Jessen, 1985). In cats and rodents, sweating is controlled by the release of acetylcholine from sympathetic innervation of peripheral sweat glands (Figure 6). These sympathetic ganglia, in turn, are controlled by innervation from preganglionic neurons located in the intermediolateral (IML) cell column of the spinal cord. Within the brain, the premotor neurons for sweating that project to the IML appear to be located in the rostral ventromedial medulla (RVMM): stimulation of the RVMM induces sweating in the cat (Davison and Koss, 1975; Shafton and McAllen, 2013) and activation of the RVMM correlates with sweating in humans (Farrell et al., 2013). As with other heat defensive responses, sweating can also be elicited by heating of the POA (Magoun et al., 1938) and sweating correlates with POA activation measured by fMRI in humans (Farrell et al., 2014). The specific circuitry that connects the POA and RVMM is unknown.
Thermoregulatory Behaviors
Animals engage in voluntary behaviors that alter their local thermal environment. These include warm and cold seeking, nesting and burrowing, huddling, basking, postural extension, and saliva spreading, as well as more complex strategies used by humans (Terrien et al., 2011). Thermoregulatory behavior is ancient and widespread: it occurs not only in endotherms (birds and mammals), but also in reptiles, fish, and many invertebrates that rely almost exclusively on behavior to respond to changes in external temperature. Thermoregulatory behavior is also motivated, at least in mammals, which means that temperature can serve as a reward that trains animals perform new tasks. For example, rats exposed to cold will learn to lever press to turn on a heat lamp (Carlton and Marks, 1958; Weiss and Laties, 1961), whereas rats exposed to heat will lever press to turn on a cold shower (Epstein and Milestone, 1968) or a cooling fan (Lipton, 1968). This suggests that thermoregulatory behaviors are driven by the same motivational systems that subserve other behaviors, such as eating and drinking, that arise from homeostatic needs.
The neural circuitry underlying these behavioral responses is poorly understood (Almeida et al., 2015). The POA is sufficient but not necessary for activation of most thermoregulatory behaviors (Table 3). The evidence for sufficiency includes the fact that cooling of the POA stimulates operant responses for heat (Gale et al., 1970; Laudenslager, 1976; Satinoff, 1964), whereas warming of the POA inhibits those responses (Carlisle, 1966; Carlisle and Laudenslager, 1979; Laudenslager, 1976). Optogenetic stimulation of warmth-activated POAPACAP/BDNF neurons promotes cold-seeking behavior and inhibits nest building in the cold (Tan et al., 2016), whereas chemogenetic stimulation of POALepR neurons promotes postural extension, a behavioral strategy for heat dissipation (Yu et al., 2016). It is important to note that, in all of these cases, the behavioral and autonomic responses to POA manipulation function in the same direction, indicating that they are part of a coordinated, homeostatic response.
Table 3.
| Behavior | POA stimulation (opto/chemogenetic) | POA warming | POA cooling | POA lesion |
|---|---|---|---|---|
| Temperature preference | Lower (cold-seeking) (Tan et al., 2016) | Lower (cold-seeking) (Adair, 1977) | Higher (warm-seeking) (Adair, 1977) | No effect on LPS-induced cold-seeking (Almeida et al., 2006) |
| Operant responses for heat reward | ND | Decrease in cold (Carlisle, 1966; Laudenslager, 1976) | Increase at baseline (Satinoff, 1964) and in cold (Laudenslager, 1976) | Increase in cold (Carlisle, 1969; Schulze et al., 1981) |
| Operant responses for cool reward | ND | Increase at baseline (Cabanac and Dib, 1983) | No effect at baseline (Cabanac and Dib, 1983) | Increase in heat (Lipton, 1968) |
| Postural extension | Increased at baseline (Yu et al., 2016) | Increased at baseline (Roberts and Mooney, 1974) | ND | Reduced in heat (Roberts and Martin, 1977; Whyte et al., 2006) |
Nevertheless, lesions that ablate the POA leave most thermoregulatory behaviors intact (Table 3) (Almeida et al., 2006; Carlisle, 1969; Lipton, 1968; Roberts and Martin, 1977; Satinoff and Rutstein, 1970). In fact POA lesions often enhance operant responding for thermal rewards, likely in order to compensate for the loss of autonomic thermoregulation (Carlisle, 1969). In general, lesioning experiments have failed to identify any forebrain brain region that is necessary for thermoregulatory behaviors in the way that the POA is necessary for autonomic responses, although a few special cases have been identified. These include a requirement for the POA in the postural extension induced by warmth (Roberts and Martin, 1977); for the DMH in cold-seeking induced by systemic inflammation, but not other stimuli (Almeida et al., 2006; Wanner et al., 2017), and for the MnPO in cold-seeking in response to salt challenge (Konishi et al., 2007).
The failure of POA lesions to block thermoregulatory behaviors has generally been interpreted to imply that the POA is “not involved” in these responses (Almeida et al., 2015). However this seems unlikely given the broad sufficiency of POA stimulation for orchestrating a variety of thermoregulatory behaviors. An alternative explanation is that the POA circuitry is complex, containing many intermingled cell types, and for this reason it is difficult to interpret the results of lesioning experiments that lack cell-type-specificity. There is ample precedent for this. For example, non-specific lesions of the hypothalamic arcuate nucleus (ARC) are famous for causing hyperphagia and obesity, suggesting the ARC functions as a “satiety center” (Choi and Dallman, 1999). However specific ablation of one ARC cell type (AgRP neurons) causes starvation (Luquet et al., 2005). In the future, it will be important to use approaches for cell-type-specific manipulation to re-investigate the role of the POA and downstream structures in the control of thermoregulatory behaviors.
6. Open questions
Many fundamental questions about the thermoregulatory system remain unanswered. Below we discuss four.
What is the mechanism and functional significance of hypothalamic temperature sensing?
Our current understanding of how the brain regulates body temperature has been strongly influenced by the seminal discovery that POA warming induces hypothermia (Magoun et al., 1938). Yet 80 years later there is still no agreement about the physiologic significance of this observation or its underlying molecular mechanism. Recent work has identified specific POA neurons that are selectively activated by ambient warmth (Tan et al., 2016), but whether these cells also sense brain temperature is unclear. Conversely the candidate warm-sensor TRPM2 has been identified in the hypothalamus (Song et al., 2016), but its broad expression raises the question of how it could function as a specific warm-sensor and, furthermore, in which neural cell types it acts. Addressing these questions will require experiments that combine exogenous control of brain temperature with cell-type-specific neural recording and manipulation.
What are the cell types that orchestrate the homeostatic response to cold?
The POA is required for thermoregulatory responses to cold, and cold exposure activates neurons in the POA (Bachtell et al., 2003; Bratincsak and Palkovits, 2004; Yoshida et al., 2005). However the specific cell types that mediate the thermoregulatory response to cold have not been identified. Optical recordings of warmth-activated POAPACAP/BDNF neurons revealed that these cells are selectively tuned to innocuous warmth, showing no response to peripheral cooling below 30°C, at least at the level that coul d be detected by fiber photometry (Tan et al., 2016). This suggests that the POA may contain a distinct population of cold-responsive cells that are the targets of the ascending cold channel. Molecular identification of these cells and elucidation of their interactions with POA warmth-activated neurons will be an important area for investigation.
What are the neural substrates of thermoregulatory behavior?
Thermoregulatory behavior remains the most enigmatic of the classic motivated behaviors that include eating and drinking. No forebrain region or cell type has yet been shown to be required for these responses. While the dogma has been that the POA is not involved, the fact that stimulation of specific POA cell types can drive robust heat-defensive behaviors reveals that these cells can serve as a genetic entry point into the underlying circuit (Tan et al., 2016; Yu et al., 2016). Moreover, the recent finding that lesions of the LPB, but not thalamus, block temperature selection behavior suggests that downstream targets of the LPB, such as the POA, are involved (Yahiro et al., 2017). It will be illuminating to identify these circuits and understand how they connect to the broader motivational system that drives other homeostatic behaviors.
To what extent does thermoregulation in rodents accurately model human physiology?
We have emphasized in this review the power of mouse genetics to probe the neural circuitry that controls body temperature, but it is important to acknowledge that there are differences in thermoregulation between rodents and humans. For example, mice subjected to food deprivation enter into torpor, a state of prolonged, regulated hypothermia, whereas rats and humans do not. Likewise large animals such as humans have much greater thermal inertia than rodents and consequently are less affected by transient changes in environmental temperature (Romanovsky, 2014). At the level of neural circuits, it remains an open question to what extent the specific cell types and interconnections that control body temperature in mice will be conserved in other species, although it is clear that many of the same brain regions are involved. Addressing these questions will require a comparative approach that investigates thermoregulation across species, possibly enabled by new technologies for gene-editing.
Acknowledgments
Z.A.K. is a New York Stem Cell Foundation-Robertson Investigator and acknowledges support from the New York Stem Cell Foundation, the American Diabetes Association Pathway Program, the Rita Allen Foundation, the Program for Breakthrough Biological Research, and the UCSF DERC (P30-DK06372) and NORC (P30-DK098722). This work was supported by DP2-DK109533, R01-NS094781, and R01-DK106399 (Z.A.K.).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Abbott SBG, Saper CB. Median preoptic glutamatergic neurons promote thermoregulatory heat loss and water consumption in mice. J Physiol. 2017;595:6569–6583. doi: 10.1113/JP274667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abreu-Vieira G, Xiao C, Gavrilova O, Reitman ML. Integration of body temperature into the analysis of energy expenditure in the mouse. Mol Metab. 2015;4:461–470. doi: 10.1016/j.molmet.2015.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida MC, Hew-Butler T, Soriano RN, Rao S, Wang W, Wang J, Tamayo N, Oliveira DL, Nucci TB, Aryal P, et al. Pharmacological blockade of the cold receptor TRPM8 attenuates autonomic and behavioral cold defenses and decreases deep body temperature. J Neurosci. 2012;32:2086–2099. doi: 10.1523/JNEUROSCI.5606-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida MC, Steiner AA, Branco LG, Romanovsky AA. Neural substrate of cold-seeking behavior in endotoxin shock. PLoS One. 2006;1:e1. doi: 10.1371/journal.pone.0000001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida MC, Vizin RC, Carrettiero DC. Current understanding on the neurophysiology of behavioral thermoregulation. Temperature (Austin) 2015;2:483–490. doi: 10.1080/23328940.2015.1095270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersen HT, Anderson B, Gale C. Central control of cold defence mechanisms and the release of ‘endopyrogen’ in the goat. Acta Physiologica Scandinavica. 1962;54:159–174. doi: 10.1111/j.1748-1716.1962.tb02341.x. [DOI] [PubMed] [Google Scholar]
- Andersson B, Grant R, Larsson S. Central control of heat loss mechanisms in the goat. Acta Physiol Scand. 1956;37:261–280. doi: 10.1111/j.1748-1716.1956.tb01362.x. [DOI] [PubMed] [Google Scholar]
- Bachtell RK, Tsivkovskaia NO, Ryabinin AE. Identification of temperature-sensitive neural circuits in mice using c-Fos expression mapping. Brain Res. 2003;960:157–164. doi: 10.1016/s0006-8993(02)03807-6. [DOI] [PubMed] [Google Scholar]
- Baker MA, Doris PA. Effect of dehydration on hypothalamic control of evaporation in the cat. J Physiol. 1982;322:457–468. doi: 10.1113/jphysiol.1982.sp014049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barker JL, Carpenter DO. Thermosensitivity of neurons in the sensorimotor cortex of the cat. Science. 1970;169:597–598. doi: 10.1126/science.169.3945.597. [DOI] [PubMed] [Google Scholar]
- Batchelder P, Kinney RO, Demlow L, Lynch CB. Effects of temperature and social interactions on huddling behavior in Mus musculus. Physiol Behav. 1983;31:97–102. doi: 10.1016/0031-9384(83)90102-6. [DOI] [PubMed] [Google Scholar]
- Bautista DM, Siemens J, Glazer JM, Tsuruda PR, Basbaum AI, Stucky CL, Jordt SE, Julius D. The menthol receptor TRPM8 is the principal detector of environmental cold. Nature. 2007;448:204–208. doi: 10.1038/nature05910. [DOI] [PubMed] [Google Scholar]
- Bauwens JD, Schmuck EG, Lindholm CR, Ertel RL, Mulligan JD, Hovis I, Viollet B, Saupe KW. Cold tolerance, cold-induced hyperphagia, and nonshivering thermogenesis are normal in alpha(1)-AMPK−/− mice. Am J Physiol Regul Integr Comp Physiol. 2011;301:R473–483. doi: 10.1152/ajpregu.00444.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett AF, Ruben JA. Endothermy and activity in vertebrates. Science. 1979;206:649–654. doi: 10.1126/science.493968. [DOI] [PubMed] [Google Scholar]
- Blessing WW, Nalivaiko E. Raphe magnus/pallidus neurons regulate tail but not mesenteric arterial blood flow in rats. Neuroscience. 2001;105:923–929. doi: 10.1016/s0306-4522(01)00251-2. [DOI] [PubMed] [Google Scholar]
- Boulant JA. Counterpoint: Heat-induced membrane depolarization of hypothalamic neurons: an unlikely mechanism of central thermosensitivity. Am J Physiol Regul Integr Comp Physiol. 2006;290:R1481–1484. doi: 10.1152/ajpregu.00655.2005. discussion R1484. [DOI] [PubMed] [Google Scholar]
- Boulant JA, Hardy JD. The effect of spinal and skin temperatures on the firing rate and thermosensitivity of preoptic neurones. J Physiol. 1974;240:639–660. doi: 10.1113/jphysiol.1974.sp010627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bratincsak A, Palkovits M. Activation of brain areas in rat following warm and cold ambient exposure. Neuroscience. 2004;127:385–397. doi: 10.1016/j.neuroscience.2004.05.016. [DOI] [PubMed] [Google Scholar]
- Bratincsak A, Palkovits M. Evidence that peripheral rather than intracranial thermal signals induce thermoregulation. Neuroscience. 2005;135:525–532. doi: 10.1016/j.neuroscience.2005.06.028. [DOI] [PubMed] [Google Scholar]
- Brock JA, McAllen RM. Spinal cord thermosensitivity: An afferent phenomenon? Temperature (Austin) 2016;3:232–239. doi: 10.1080/23328940.2016.1157665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown JW, Sirlin EA, Benoit AM, Hoffman JM, Darnall RA. Activation of 5-HT1A receptors in medullary raphe disrupts sleep and decreases shivering during cooling in the conscious piglet. Am J Physiol Regul Integr Comp Physiol. 2008;294:R884–894. doi: 10.1152/ajpregu.00655.2007. [DOI] [PubMed] [Google Scholar]
- Cabanac M. Temperature regulation. Annu Rev Physiol. 1975;37:415–439. doi: 10.1146/annurev.ph.37.030175.002215. [DOI] [PubMed] [Google Scholar]
- Cao WH, Fan W, Morrison SF. Medullary pathways mediating specific sympathetic responses to activation of dorsomedial hypothalamus. Neuroscience. 2004;126:229–240. doi: 10.1016/j.neuroscience.2004.03.013. [DOI] [PubMed] [Google Scholar]
- Cao WH, Morrison SF. Glutamate receptors in the raphe pallidus mediate brown adipose tissue thermogenesis evoked by activation of dorsomedial hypothalamic neurons. Neuropharmacology. 2006;51:426–437. doi: 10.1016/j.neuropharm.2006.03.031. [DOI] [PubMed] [Google Scholar]
- Carlisle HJ. Behavioural significance of hypothalamic temperature-sensitive cells. Nature. 1966;209:1324–1325. doi: 10.1038/2091324a0. [DOI] [PubMed] [Google Scholar]
- Carlisle HJ. Effect of preoptic and anterior hypothalamic lesions on behavioral thermoregulation in the cold. J Comp Physiol Psychol. 1969;69:391–402. doi: 10.1037/h0028170. [DOI] [PubMed] [Google Scholar]
- Carlisle HJ, Laudenslager ML. Observations on the thermoregulatory effects of preoptic warming in rats. Physiology & behavior. 1979;23:723–732. doi: 10.1016/0031-9384(79)90166-5. [DOI] [PubMed] [Google Scholar]
- Carlton PL, Marks RA. Cold exposure and heat reinforced operant behavior. Science. 1958;128:1344. doi: 10.1126/science.128.3335.1344. [DOI] [PubMed] [Google Scholar]
- Caterina MJ, Leffler A, Malmberg AB, Martin WJ, Trafton J, Petersen-Zeitz KR, Koltzenburg M, Basbaum AI, Julius D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science. 2000;288:306–313. doi: 10.1126/science.288.5464.306. [DOI] [PubMed] [Google Scholar]
- Cavanaugh DJ, Chesler AT, Jackson AC, Sigal YM, Yamanaka H, Grant R, O’Donnell D, Nicoll RA, Shah NM, Julius D, et al. Trpv1 reporter mice reveal highly restricted brain distribution and functional expression in arteriolar smooth muscle cells. J Neurosci. 2011;31:5067–5077. doi: 10.1523/JNEUROSCI.6451-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen XM, Hosono T, Yoda T, Fukuda Y, Kanosue K. Efferent projection from the preoptic area for the control of non-shivering thermogenesis in rats. J Physiol. 1998;512(Pt 3):883–892. doi: 10.1111/j.1469-7793.1998.883bd.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen XM, Nishi M, Taniguchi A, Nagashima K, Shibata M, Kanosue K. The caudal periaqueductal gray participates in the activation of brown adipose tissue in rats. Neurosci Lett. 2002;331:17–20. doi: 10.1016/s0304-3940(02)00757-7. [DOI] [PubMed] [Google Scholar]
- Choi S, Dallman MF. Hypothalamic obesity: multiple routes mediated by loss of function in medial cell groups. Endocrinology. 1999;140:4081–4088. doi: 10.1210/endo.140.9.6964. [DOI] [PubMed] [Google Scholar]
- Conceicao EPS, Madden CJ, Morrison SF. Tonic inhibition of brown adipose tissue sympathetic nerve activity via muscarinic acetylcholine receptors in the rostral raphe pallidus. J Physiol. 2017;595:7495–7508. doi: 10.1113/JP275299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costill DL, Fink WJ. Plasma volume changes following exercise and thermal dehydration. J Appl Physiol. 1974;37:521–525. doi: 10.1152/jappl.1974.37.4.521. [DOI] [PubMed] [Google Scholar]
- Craig AD, Bushnell MC, Zhang ET, Blomqvist A. A thalamic nucleus specific for pain and temperature sensation. Nature. 1994;372:770–773. doi: 10.1038/372770a0. [DOI] [PubMed] [Google Scholar]
- Crompton AW, Taylor CR, Jagger JA. Evolution of homeothermy in mammals. Nature. 1978;272:333–336. doi: 10.1038/272333a0. [DOI] [PubMed] [Google Scholar]
- Davern PJ. A role for the lateral parabrachial nucleus in cardiovascular function and fluid homeostasis. Front Physiol. 2014;5:436. doi: 10.3389/fphys.2014.00436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davison MA, Koss MC. Brainstem loci for activation of electrodermal response in the cat. Am J Physiol. 1975;229:930–934. doi: 10.1152/ajplegacy.1975.229.4.930. [DOI] [PubMed] [Google Scholar]
- Dhaka A, Murray AN, Mathur J, Earley TJ, Petrus MJ, Patapoutian A. TRPM8 is required for cold sensation in mice. Neuron. 2007;54:371–378. doi: 10.1016/j.neuron.2007.02.024. [DOI] [PubMed] [Google Scholar]
- Dodd GT, Worth AA, Nunn N, Korpal AK, Bechtold DA, Allison MB, Myers MG, Jr, Statnick MA, Luckman SM. The thermogenic effect of leptin is dependent on a distinct population of prolactin-releasing peptide neurons in the dorsomedial hypothalamus. Cell Metab. 2014;20:639–649. doi: 10.1016/j.cmet.2014.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenman JS, Edinger HM, Barker JL, Carpenter DO. Neuronal thermosensitivity. Science. 1971;172:1360–1362. doi: 10.1126/science.172.3990.1360. [DOI] [PubMed] [Google Scholar]
- Elmquist JK, Scammell TE, Jacobson CD, Saper CB. Distribution of Fos-like immunoreactivity in the rat brain following intravenous lipopolysaccharide administration. J Comp Neurol. 1996;371:85–103. doi: 10.1002/(SICI)1096-9861(19960715)371:1<85::AID-CNE5>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- Epstein AN, Milestone R. Showering as a coolant for rats exposed to heat. Science. 1968;160:895–896. doi: 10.1126/science.160.3830.895. [DOI] [PubMed] [Google Scholar]
- Evans SS, Repasky EA, Fisher DT. Fever and the thermal regulation of immunity: the immune system feels the heat. Nat Rev Immunol. 2015;15:335–349. doi: 10.1038/nri3843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrell MJ, Trevaks D, McAllen RM. Preoptic activation and connectivity during thermal sweating in humans. Temperature (Austin) 2014;1:135–141. doi: 10.4161/temp.29667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrell MJ, Trevaks D, Taylor NA, McAllen RM. Brain stem representation of thermal and psychogenic sweating in humans. Am J Physiol Regul Integr Comp Physiol. 2013;304:R810–817. doi: 10.1152/ajpregu.00041.2013. [DOI] [PubMed] [Google Scholar]
- Fortney SM, Wenger CB, Bove JR, Nadel ER. Effect of hyperosmolality on control of blood flow and sweating. J Appl Physiol Respir Environ Exerc Physiol. 1984;57:1688–1695. doi: 10.1152/jappl.1984.57.6.1688. [DOI] [PubMed] [Google Scholar]
- Fuller A, Carter RN, Mitchell D. Brain and abdominal temperatures at fatigue in rats exercising in the heat. J Appl Physiol (1985) 1998;84:877–883. doi: 10.1152/jappl.1998.84.3.877. [DOI] [PubMed] [Google Scholar]
- Gale CC, Mathews M, Young J. Behavioral thermoregulatory responses to hypothalamic cooling and warming in baboons. Physiol Behav. 1970;5:1–6. doi: 10.1016/0031-9384(70)90003-x. [DOI] [PubMed] [Google Scholar]
- Gavva NR. Body-temperature maintenance as the predominant function of the vanilloid receptor TRPV1. Trends Pharmacol Sci. 2008;29:550–557. doi: 10.1016/j.tips.2008.08.003. [DOI] [PubMed] [Google Scholar]
- Gavva NR, Bannon AW, Surapaneni S, Hovland DN, Jr, Lehto SG, Gore A, Juan T, Deng H, Han B, Klionsky L, et al. The vanilloid receptor TRPV1 is tonically activated in vivo and involved in body temperature regulation. J Neurosci. 2007;27:3366–3374. doi: 10.1523/JNEUROSCI.4833-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gavva NR, Davis C, Lehto SG, Rao S, Wang W, Zhu DX. Transient receptor potential melastatin 8 (TRPM8) channels are involved in body temperature regulation. Mol Pain. 2012;8:36. doi: 10.1186/1744-8069-8-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geerling JC, Kim M, Mahoney CE, Abbott SB, Agostinelli LJ, Garfield AS, Krashes MJ, Lowell BB, Scammell TE. Genetic identity of thermosensory relay neurons in the lateral parabrachial nucleus. Am J Physiol Regul Integr Comp Physiol. 2016;310:R41–54. doi: 10.1152/ajpregu.00094.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammel HT. Regulation of internal body temperature. Annual review of physiology. 1968;30:641–710. doi: 10.1146/annurev.ph.30.030168.003233. [DOI] [PubMed] [Google Scholar]
- Hammel HT, Hardy JD, Fusco MM. Thermoregulatory responses to hypothalamic cooling in unanesthetized dogs. The American journal of physiology. 1960;198:481–486. doi: 10.1152/ajplegacy.1960.198.3.481. [DOI] [PubMed] [Google Scholar]
- Hammel HT, Jackson DC, Stolwijk JA, Hardy JD, Stromme SB. Temperature Regulation by Hypothalamic Proportional Control with an Adjustable Set Point. J Appl Physiol. 1963;18:1146–1154. doi: 10.1152/jappl.1963.18.6.1146. [DOI] [PubMed] [Google Scholar]
- Harikai N, Tomogane K, Sugawara T, Tashiro S. Differences in hypothalamic Fos expressions between two heat stress conditions in conscious mice. Brain Res Bull. 2003;61:617–626. doi: 10.1016/j.brainresbull.2003.07.001. [DOI] [PubMed] [Google Scholar]
- Heinrich B. Why have some animals evolved to regulate a high body temperature? The American Naturalist. 1977;111:623–640. [Google Scholar]
- Heller HC, Edgar DM, Grahn DA, Glotzbach SF. Comprehensive Physiology. John Wiley & Sons, Inc.; 2010. Sleep, Thermoregulation, and Circadian Rhythms. [Google Scholar]
- Hellon RF. The stimulation of hypothalamic neurones by changes in ambient temperature. Pflugers Arch. 1970;321:56–66. doi: 10.1007/BF00594122. [DOI] [PubMed] [Google Scholar]
- Hellon RF. Are single-unit recordings useful in understanding thermoregulation? Yale J Biol Med. 1986;59:197–203. [PMC free article] [PubMed] [Google Scholar]
- Hellstrom B, Hammel HT. Some characteristics of temperature regulation in the unanesthetized dog. The American journal of physiology. 1967;213:547–556. doi: 10.1152/ajplegacy.1967.213.2.547. [DOI] [PubMed] [Google Scholar]
- Hemingway A, Forgrave P, Birzis L. Shivering suppression by hypothalamic stimulation. J Neurophysiol. 1954;17:375–386. doi: 10.1152/jn.1954.17.4.375. [DOI] [PubMed] [Google Scholar]
- Hensel H. Neural processes in thermoregulation. Physiol Rev. 1973;53:948–1017. doi: 10.1152/physrev.1973.53.4.948. [DOI] [PubMed] [Google Scholar]
- Hermann DM, Luppi PH, Peyron C, Hinckel P, Jouvet M. Afferent projections to the rat nuclei raphe magnus, raphe pallidus and reticularis gigantocellularis pars alpha demonstrated by iontophoretic application of choleratoxin (subunit b) J Chem Neuroanat. 1997;13:1–21. doi: 10.1016/s0891-0618(97)00019-7. [DOI] [PubMed] [Google Scholar]
- Hori T. Capsaicin and central control of thermoregulation. Pharmacol Ther. 1984;26:389–416. doi: 10.1016/0163-7258(84)90041-x. [DOI] [PubMed] [Google Scholar]
- Hosoya Y, Sugiura Y, Zhang FZ, Ito R, Kohno K. Direct projection from the dorsal hypothalamic area to the nucleus raphe pallidus: a study using anterograde transport with Phaseolus vulgaris leucoagglutinin in the rat. Exp Brain Res. 1989;75:40–46. doi: 10.1007/BF00248528. [DOI] [PubMed] [Google Scholar]
- Huang SM, Li X, Yu Y, Wang J, Caterina MJ. TRPV3 and TRPV4 ion channels are not major contributors to mouse heat sensation. Mol Pain. 2011;7:37. doi: 10.1186/1744-8069-7-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hylden JL, Anton F, Nahin RL. Spinal lamina I projection neurons in the rat: collateral innervation of parabrachial area and thalamus. Neuroscience. 1989;28:27–37. doi: 10.1016/0306-4522(89)90229-7. [DOI] [PubMed] [Google Scholar]
- Iida T, Shimizu I, Nealen ML, Campbell A, Caterina M. Attenuated fever response in mice lacking TRPV1. Neurosci Lett. 2005;378:28–33. doi: 10.1016/j.neulet.2004.12.007. [DOI] [PubMed] [Google Scholar]
- Ishiwata T, Saito T, Hasegawa H, Yazawa T, Kotani Y, Otokawa M, Aihara Y. Changes of body temperature and thermoregulatory responses of freely moving rats during GABAergic pharmacological stimulation to the preoptic area and anterior hypothalamus in several ambient temperatures. Brain Res. 2005;1048:32–40. doi: 10.1016/j.brainres.2005.04.027. [DOI] [PubMed] [Google Scholar]
- Jeong JH, Lee DK, Blouet C, Ruiz HH, Buettner C, Chua S, Jr, Schwartz GJ, Jo YH. Cholinergic neurons in the dorsomedial hypothalamus regulate mouse brown adipose tissue metabolism. Mol Metab. 2015;4:483–492. doi: 10.1016/j.molmet.2015.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jessen C. Thermal afferents in the control of body temperature. Pharmacol Ther. 1985;28:107–134. doi: 10.1016/0163-7258(85)90085-3. [DOI] [PubMed] [Google Scholar]
- Jessen C, Ludwig O. Spinal cord and hypothalamus as core sensors of temperature in the conscious dog. II. Addition of signals. Pflugers Arch. 1971;324:205–216. doi: 10.1007/BF00586419. [DOI] [PubMed] [Google Scholar]
- Kanosue K, Crawshaw LI, Nagashima K, Yoda T. Concepts to utilize in describing thermoregulation and neurophysiological evidence for how the system works. Eur J Appl Physiol. 2010;109:5–11. doi: 10.1007/s00421-009-1256-6. [DOI] [PubMed] [Google Scholar]
- Kanosue K, Niwa K, Andrew PD, Yasuda H, Yanase M, Tanaka H, Matsumura K. Lateral distribution of hypothalamic signals controlling thermoregulatory vasomotor activity and shivering in rats. Am J Physiol. 1991;260:R486–493. doi: 10.1152/ajpregu.1991.260.3.R486. [DOI] [PubMed] [Google Scholar]
- Kataoka N, Hioki H, Kaneko T, Nakamura K. Psychological stress activates a dorsomedial hypothalamus-medullary raphe circuit driving brown adipose tissue thermogenesis and hyperthermia. Cell Metab. 2014;20:346–358. doi: 10.1016/j.cmet.2014.05.018. [DOI] [PubMed] [Google Scholar]
- Kelso SR, Boulant JA. Effect of synaptic blockade on thermosensitive neurons in hypothalamic tissue slices. Am J Physiol. 1982;243:R480–490. doi: 10.1152/ajpregu.1982.243.5.R480. [DOI] [PubMed] [Google Scholar]
- Kerman IA, Enquist LW, Watson SJ, Yates BJ. Brainstem substrates of sympatho-motor circuitry identified using trans-synaptic tracing with pseudorabies virus recombinants. J Neurosci. 2003;23:4657–4666. doi: 10.1523/JNEUROSCI.23-11-04657.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Key BJ, Wigfield CC. The influence of the ventrolateral medulla on thermoregulatory circulations in the rat. J Auton Nerv Syst. 1994;48:79–89. doi: 10.1016/0165-1838(94)90162-7. [DOI] [PubMed] [Google Scholar]
- Knight ZA, Tan K, Birsoy K, Schmidt S, Garrison JL, Wysocki RW, Emiliano A, Ekstrand MI, Friedman JM. Molecular profiling of activated neurons by phosphorylated ribosome capture. Cell. 2012;151:1126–1137. doi: 10.1016/j.cell.2012.10.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knowlton WM, Palkar R, Lippoldt EK, McCoy DD, Baluch F, Chen J, McKemy DD. A sensory-labeled line for cold: TRPM8-expressing sensory neurons define the cellular basis for cold, cold pain, and cooling-mediated analgesia. J Neurosci. 2013;33:2837–2848. doi: 10.1523/JNEUROSCI.1943-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knox GV, Campbell C, Lomax P. Cutaneous temperature and unit activity in the hypothalamic thermoregulatory centers. Exp Neurol. 1973;40:717–730. doi: 10.1016/0014-4886(73)90106-4. [DOI] [PubMed] [Google Scholar]
- Kobayashi A, Osaka T. Involvement of the parabrachial nucleus in thermogenesis induced by environmental cooling in the rat. Pflugers Arch. 2003;446:760–765. doi: 10.1007/s00424-003-1119-7. [DOI] [PubMed] [Google Scholar]
- Kobayashi S, Hori A, Matsumura K, Hosokawa H. Point: Heat-induced membrane depolarization of hypothalamic neurons: a putative mechanism of central thermosensitivity. Am J Physiol Regul Integr Comp Physiol. 2006;290:R1479–1480. doi: 10.1152/ajpregu.00655.2005. discussion R1484. [DOI] [PubMed] [Google Scholar]
- Konishi M, Kanosue K, Kano M, Kobayashi A, Nagashima K. The median preoptic nucleus is involved in the facilitation of heat-escape/cold-seeking behavior during systemic salt loading in rats. Am J Physiol Regul Integr Comp Physiol. 2007;292:R150–159. doi: 10.1152/ajpregu.00769.2005. [DOI] [PubMed] [Google Scholar]
- Krueger JM, Takahashi S. Thermoregulation and sleep. Closely linked but separable. Ann N Y Acad Sci. 1997;813:281–286. doi: 10.1111/j.1749-6632.1997.tb51706.x. [DOI] [PubMed] [Google Scholar]
- Laudenslager ML. Proportional hypothalamic control of behavioral thermoregulation in the squirrel monkey. Physiol Behav. 1976;17:383–390. doi: 10.1016/0031-9384(76)90095-0. [DOI] [PubMed] [Google Scholar]
- Leib DE, Zimmerman CA, Poormoghaddam A, Huey EL, Ahn JS, Lin YC, Tan CL, Chen Y, Knight ZA. The Forebrain Thirst Circuit Drives Drinking through Negative Reinforcement. Neuron. 2017;96:1272–1281 e1274. doi: 10.1016/j.neuron.2017.11.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipton JM. Effects of preoptic lesions on heat-escape responding and colonic temperature in the rat. Physiol Behav. 1968;3:165–169. [Google Scholar]
- Long MA, Fee MS. Using temperature to analyse temporal dynamics in the songbird motor pathway. Nature. 2008;456:189–194. doi: 10.1038/nature07448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luquet S, Perez FA, Hnasko TS, Palmiter RD. NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science. 2005;310:683–685. doi: 10.1126/science.1115524. [DOI] [PubMed] [Google Scholar]
- Ma Q. Labeled lines meet and talk: population coding of somatic sensations. J Clin Invest. 2010;120:3773–3778. doi: 10.1172/JCI43426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madden CJ, Morrison SF. Excitatory amino acid receptor activation in the raphe pallidus area mediates prostaglandin-evoked thermogenesis. Neuroscience. 2003;122:5–15. doi: 10.1016/s0306-4522(03)00527-x. [DOI] [PubMed] [Google Scholar]
- Madden CJ, Morrison SF. Excitatory amino acid receptors in the dorsomedial hypothalamus mediate prostaglandin-evoked thermogenesis in brown adipose tissue. Am J Physiol Regul Integr Comp Physiol. 2004;286:R320–325. doi: 10.1152/ajpregu.00515.2003. [DOI] [PubMed] [Google Scholar]
- Magoun HW, Harrison F, Brobeck JR, Ranson SW. Activation of Heat Loss Mechanisms by Local Heating of the Brain. Journal of Neurophysiology. 1938;1:101–114. [Google Scholar]
- McAllen RM, Tanaka M, Ootsuka Y, McKinley MJ. Multiple thermoregulatory effectors with independent central controls. Eur J Appl Physiol. 2010;109:27–33. doi: 10.1007/s00421-009-1295-z. [DOI] [PubMed] [Google Scholar]
- McKemy DD, Neuhausser WM, Julius D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature. 2002;416:52–58. doi: 10.1038/nature719. [DOI] [PubMed] [Google Scholar]
- Morimoto T. Thermoregulation and body fluids: role of blood volume and central venous pressure. Jpn J Physiol. 1990;40:165–179. doi: 10.2170/jjphysiol.40.165. [DOI] [PubMed] [Google Scholar]
- Morrison SF. Raphe pallidus neurons mediate prostaglandin E2-evoked increases in brown adipose tissue thermogenesis. Neuroscience. 2003;121:17–24. doi: 10.1016/s0306-4522(03)00363-4. [DOI] [PubMed] [Google Scholar]
- Morrison SF, Madden CJ, Tupone D. Central control of brown adipose tissue thermogenesis. Front Endocrinol (Lausanne) 2012;3 doi: 10.3389/fendo.2012.00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison SF, Nakamura K. Central neural pathways for thermoregulation. Front Biosci (Landmark Ed) 2011;16:74–104. doi: 10.2741/3677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison SF, Sved AF, Passerin AM. GABA-mediated inhibition of raphe pallidus neurons regulates sympathetic outflow to brown adipose tissue. Am J Physiol. 1999;276:R290–297. doi: 10.1152/ajpregu.1999.276.2.R290. [DOI] [PubMed] [Google Scholar]
- Nadel ER, Mitchell JW, Stolwijk JA. Differential thermal sensitivity in the human skin. Pflugers Arch. 1973;340:71–76. doi: 10.1007/BF00592198. [DOI] [PubMed] [Google Scholar]
- Nakamura K, Matsumura K, Hubschle T, Nakamura Y, Hioki H, Fujiyama F, Boldogkoi Z, Konig M, Thiel HJ, Gerstberger R, et al. Identification of sympathetic premotor neurons in medullary raphe regions mediating fever and other thermoregulatory functions. J Neurosci. 2004;24:5370–5380. doi: 10.1523/JNEUROSCI.1219-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura K, Matsumura K, Kaneko T, Kobayashi S, Katoh H, Negishi M. The rostral raphe pallidus nucleus mediates pyrogenic transmission from the preoptic area. J Neurosci. 2002;22:4600–4610. doi: 10.1523/JNEUROSCI.22-11-04600.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura K, Morrison SF. Central efferent pathways mediating skin cooling-evoked sympathetic thermogenesis in brown adipose tissue. Am J Physiol Regul Integr Comp Physiol. 2007;292:R127–136. doi: 10.1152/ajpregu.00427.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura K, Morrison SF. A thermosensory pathway that controls body temperature. Nat Neurosci. 2008;11:62–71. doi: 10.1038/nn2027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura K, Morrison SF. A thermosensory pathway mediating heat-defense responses. Proc Natl Acad Sci U S A. 2010;107:8848–8853. doi: 10.1073/pnas.0913358107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura K, Morrison SF. Central efferent pathways for cold-defensive and febrile shivering. J Physiol. 2011;589:3641–3658. doi: 10.1113/jphysiol.2011.210047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura Y, Nakamura K, Matsumura K, Kobayashi S, Kaneko T, Morrison SF. Direct pyrogenic input from prostaglandin EP3 receptor-expressing preoptic neurons to the dorsomedial hypothalamus. Eur J Neurosci. 2005;22:3137–3146. doi: 10.1111/j.1460-9568.2005.04515.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura Y, Nakamura K, Morrison SF. Different populations of prostaglandin EP3 receptor-expressing preoptic neurons project to two fever-mediating sympathoexcitatory brain regions. Neuroscience. 2009;161:614–620. doi: 10.1016/j.neuroscience.2009.03.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakayama T, Eisenman JS, Hardy JD. Single unit activity of anterior hypothalamus during local heating. Science. 1961;134:560–561. doi: 10.1126/science.134.3478.560. [DOI] [PubMed] [Google Scholar]
- Nakayama T, Hammel HT, Hardy JD, Eisenman JS. Thermal stimulation of electrical activity of single units of the preoptic region. American Journal of Physiology. 1963;204:1122–1126. [Google Scholar]
- Nakayama T, Hori Y, Suzuki M, Yonezawa T, Yamamoto K. Thermo-sensitive neurons in preoptic and anterior hypothalamic tissue cultures in vitro. Neurosci Lett. 1978;9:23–26. doi: 10.1016/0304-3940(78)90042-3. [DOI] [PubMed] [Google Scholar]
- Nason MW, Jr, Mason P. Modulation of sympathetic and somatomotor function by the ventromedial medulla. J Neurophysiol. 2004;92:510–522. doi: 10.1152/jn.00089.2004. [DOI] [PubMed] [Google Scholar]
- Ootsuka Y, Blessing WW, McAllen RM. Inhibition of rostral medullary raphe neurons prevents cold-induced activity in sympathetic nerves to rat tail and rabbit ear arteries. Neurosci Lett. 2004;357:58–62. doi: 10.1016/j.neulet.2003.11.067. [DOI] [PubMed] [Google Scholar]
- Ootsuka Y, McAllen RM. Interactive drives from two brain stem premotor nuclei are essential to support rat tail sympathetic activity. Am J Physiol Regul Integr Comp Physiol. 2005;289:R1107–1115. doi: 10.1152/ajpregu.00005.2005. [DOI] [PubMed] [Google Scholar]
- Ootsuka Y, Tanaka M. Control of cutaneous blood flow by central nervous system. Temperature (Austin) 2015;2:392–405. doi: 10.1080/23328940.2015.1069437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osaka T. Cold-induced thermogenesis mediated by GABA in the preoptic area of anesthetized rats. Am J Physiol Regul Integr Comp Physiol. 2004;287:R306–313. doi: 10.1152/ajpregu.00003.2004. [DOI] [PubMed] [Google Scholar]
- Osborne PG, Kurosawa M. Perfusion of the preoptic area with muscimol or prostaglandin E2 stimulates cardiovascular function in anesthetized rats. J Auton Nerv Syst. 1994;46:199–205. doi: 10.1016/0165-1838(94)90037-x. [DOI] [PubMed] [Google Scholar]
- Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, Story GM, Earley TJ, Dragoni I, McIntyre P, Bevan S, et al. A TRP channel that senses cold stimuli and menthol. Cell. 2002;108:705–715. doi: 10.1016/s0092-8674(02)00652-9. [DOI] [PubMed] [Google Scholar]
- Pogorzala LA, Mishra SK, Hoon MA. The cellular code for mammalian thermosensation. The Journal of Neuroscience: the official journal of the Society for Neuroscience. 2013;33:5533–5541. doi: 10.1523/JNEUROSCI.5788-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ran C, Hoon MA, Chen X. The coding of cutaneous temperature in the spinal cord. Nat Neurosci. 2016;19:1201–1209. doi: 10.1038/nn.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rathner JA, Madden CJ, Morrison SF. Central pathway for spontaneous and prostaglandin E2-evoked cutaneous vasoconstriction. Am J Physiol Regul Integr Comp Physiol. 2008;295:R343–354. doi: 10.1152/ajpregu.00115.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rathner JA, McAllen RM. Differential control of sympathetic drive to the rat tail artery and kidney by medullary premotor cell groups. Brain Res. 1999;834:196–199. doi: 10.1016/s0006-8993(99)01568-1. [DOI] [PubMed] [Google Scholar]
- Rezai-Zadeh K, Yu S, Jiang Y, Laque A, Schwartzenburg C, Morrison CD, Derbenev AV, Zsombok A, Munzberg H. Leptin receptor neurons in the dorsomedial hypothalamus are key regulators of energy expenditure and body weight, but not food intake. Mol Metab. 2014;3:681–693. doi: 10.1016/j.molmet.2014.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts WW, Martin JR. Effects of lesions in central thermosensitive areas on thermoregulatory responses in rat. Physiol Behav. 1977;19:503–511. doi: 10.1016/0031-9384(77)90226-8. [DOI] [PubMed] [Google Scholar]
- Romanov RA, Zeisel A, Bakker J, Girach F, Hellysaz A, Tomer R, Alpar A, Mulder J, Clotman F, Keimpema E, et al. Molecular interrogation of hypothalamic organization reveals distinct dopamine neuronal subtypes. Nat Neurosci. 2017;20:176–188. doi: 10.1038/nn.4462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romanovsky AA. Skin temperature: its role in thermoregulation. Acta Physiol (Oxf) 2014;210:498–507. doi: 10.1111/apha.12231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romanovsky AA, Almeida MC, Garami A, Steiner AA, Norman MH, Morrison SF, Nakamura K, Burmeister JJ, Nucci TB. The transient receptor potential vanilloid-1 channel in thermoregulation: a thermosensor it is not. Pharmacol Rev. 2009;61:228–261. doi: 10.1124/pr.109.001263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satinoff E. Behavioral Thermoregulation in Response to Local Cooling of the Rat Brain. Am J Physiol. 1964;206:1389–1394. doi: 10.1152/ajplegacy.1964.206.6.1389. [DOI] [PubMed] [Google Scholar]
- Satinoff E. Neural organization and evolution of thermal regulation in mammals. Science. 1978;201:16–22. doi: 10.1126/science.351802. [DOI] [PubMed] [Google Scholar]
- Satinoff E, Rutstein J. Behavioral thermoregulation in rats with anterior hypothalamic lesions. Journal of comparative and physiological psychology. 1970;71:77–82. doi: 10.1037/h0028959. [DOI] [PubMed] [Google Scholar]
- Satinoff E, Valentino D, Teitelbaum P. Thermoregulatory cold-defense deficits in rats with preoptic/anterior hypothalamic lesions. Brain Res Bull. 1976;1:553–565. doi: 10.1016/0361-9230(76)90082-4. [DOI] [PubMed] [Google Scholar]
- Scammell TE, Elmquist JK, Griffin JD, Saper CB. Ventromedial preoptic prostaglandin E2 activates fever-producing autonomic pathways. J Neurosci. 1996;16:6246–6254. doi: 10.1523/JNEUROSCI.16-19-06246.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scammell TE, Price KJ, Sagar SM. Hyperthermia induces c-fos expression in the preoptic area. Brain Res. 1993;618:303–307. doi: 10.1016/0006-8993(93)91280-6. [DOI] [PubMed] [Google Scholar]
- Shafton AD, Kitchener P, McKinley MJ, McAllen RM. Reflex control of rat tail sympathetic nerve activity by abdominal temperature. Temperature (Austin) 2014;1:37–41. doi: 10.4161/temp.29597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shafton AD, McAllen RM. Location of cat brain stem neurons that drive sweating. Am J Physiol Regul Integr Comp Physiol. 2013;304:R804–809. doi: 10.1152/ajpregu.00040.2013. [DOI] [PubMed] [Google Scholar]
- Song K, Wang H, Kamm GB, Pohle J, Reis FC, Heppenstall P, Wende H, Siemens J. The TRPM2 channel is a hypothalamic heat sensor that limits fever and can drive hypothermia. Science. 2016;353:1393–1398. doi: 10.1126/science.aaf7537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steiner AA, Turek VF, Almeida MC, Burmeister JJ, Oliveira DL, Roberts JL, Bannon AW, Norman MH, Louis JC, Treanor JJ, et al. Nonthermal activation of transient receptor potential vanilloid-1 channels in abdominal viscera tonically inhibits autonomic cold-defense effectors. The Journal of Neuroscience: the official journal of the Society for Neuroscience. 2007;27:7459–7468. doi: 10.1523/JNEUROSCI.1483-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuart DG, Kawamura Y, Hemingway A. Activation and suppression of shivering during septal and hypothalamic stimulation. Exp Neurol. 1961;4:485–506. doi: 10.1016/0014-4886(61)90048-6. [DOI] [PubMed] [Google Scholar]
- Stuart DG, Kawamura Y, Hemingway A, Price WM. Effects of septal and hypothalamic lesions on shivering. Exp Neurol. 1962;5:335–347. doi: 10.1016/0014-4886(62)90042-0. [DOI] [PubMed] [Google Scholar]
- Szelenyi Z, Hummel Z, Szolcsanyi J, Davis JB. Daily body temperature rhythm and heat tolerance in TRPV1 knockout and capsaicin pretreated mice. Eur J Neurosci. 2004;19:1421–1424. doi: 10.1111/j.1460-9568.2004.03221.x. [DOI] [PubMed] [Google Scholar]
- Takahashi Y, Zhang W, Sameshima K, Kuroki C, Matsumoto A, Sunanaga J, Kono Y, Sakurai T, Kanmura Y, Kuwaki T. Orexin neurons are indispensable for prostaglandin E2-induced fever and defence against environmental cooling in mice. J Physiol. 2013;591:5623–5643. doi: 10.1113/jphysiol.2013.261271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan CH, McNaughton PA. The TRPM2 ion channel is required for sensitivity to warmth. Nature. 2016;536:460–463. doi: 10.1038/nature19074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan CL, Cooke EK, Leib DE, Lin YC, Daly GE, Zimmerman CA, Knight ZA. Warm-Sensitive Neurons that Control Body Temperature. Cell. 2016;167:47–59 e15. doi: 10.1016/j.cell.2016.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka M, McKinley MJ, McAllen RM. Roles of two preoptic cell groups in tonic and febrile control of rat tail sympathetic fibers. Am J Physiol Regul Integr Comp Physiol. 2009;296:R1248–1257. doi: 10.1152/ajpregu.91010.2008. [DOI] [PubMed] [Google Scholar]
- Tanaka M, McKinley MJ, McAllen RM. Preoptic-raphe connections for thermoregulatory vasomotor control. J Neurosci. 2011;31:5078–5088. doi: 10.1523/JNEUROSCI.6433-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka M, McKinley MJ, McAllen RM. Role of an excitatory preoptic-raphe pathway in febrile vasoconstriction of the rat’s tail. Am J Physiol Regul Integr Comp Physiol. 2013;305:R1479–1489. doi: 10.1152/ajpregu.00401.2013. [DOI] [PubMed] [Google Scholar]
- Tanaka M, Nagashima K, McAllen RM, Kanosue K. Role of the medullary raphe in thermoregulatory vasomotor control in rats. J Physiol. 2002;540:657–664. doi: 10.1113/jphysiol.2001.012989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka M, Owens NC, Nagashima K, Kanosue K, McAllen RM. Reflex activation of rat fusimotor neurons by body surface cooling, and its dependence on the medullary raphe. J Physiol. 2006;572:569–583. doi: 10.1113/jphysiol.2005.102400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka M, Tonouchi M, Hosono T, Nagashima K, Yanase-Fujiwara M, Kanosue K. Hypothalamic region facilitating shivering in rats. Jpn J Physiol. 2001;51:625–629. doi: 10.2170/jjphysiol.51.625. [DOI] [PubMed] [Google Scholar]
- Terrien J, Perret M, Aujard F. Behavioral thermoregulation in mammals: a review. Front Biosci (Landmark Ed) 2011;16:1428–1444. doi: 10.2741/3797. [DOI] [PubMed] [Google Scholar]
- Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, Skinner K, Raumann BE, Basbaum AI, Julius D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron. 1998;21:531–543. doi: 10.1016/s0896-6273(00)80564-4. [DOI] [PubMed] [Google Scholar]
- Tupone D, Madden CJ, Cano G, Morrison SF. An orexinergic projection from perifornical hypothalamus to raphe pallidus increases rat brown adipose tissue thermogenesis. J Neurosci. 2011;31:15944–15955. doi: 10.1523/JNEUROSCI.3909-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Zoeren JG, Stricker EM. Thermal homeostasis in rats after intrahypothalamic injections of 6-hyroxydopamine. Am J Physiol. 1976;230:932–939. doi: 10.1152/ajplegacy.1976.230.4.932. [DOI] [PubMed] [Google Scholar]
- Vellani V, Mapplebeck S, Moriondo A, Davis JB, McNaughton PA. Protein kinase C activation potentiates gating of the vanilloid receptor VR1 by capsaicin, protons, heat and anandamide. J Physiol. 2001;534:813–825. doi: 10.1111/j.1469-7793.2001.00813.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vriens J, Nilius B, Voets T. Peripheral thermosensation in mammals. Nat Rev Neurosci. 2014;15:573–589. doi: 10.1038/nrn3784. [DOI] [PubMed] [Google Scholar]
- Waites GM. The effect of heating the scrotum of the ram on respiration and body temperature. Q J Exp Physiol Cogn Med Sci. 1962;47:314–323. doi: 10.1113/expphysiol.1962.sp001615. [DOI] [PubMed] [Google Scholar]
- Walters TJ, Ryan KL, Tate LM, Mason PA. Exercise in the heat is limited by a critical internal temperature. J Appl Physiol (1985) 2000;89:799–806. doi: 10.1152/jappl.2000.89.2.799. [DOI] [PubMed] [Google Scholar]
- Wanner SP, Almeida MC, Shimansky YP, Oliveira DL, Eales JR, Coimbra CC, Romanovsky AA. Cold-Induced Thermogenesis and Inflammation-Associated Cold-Seeking Behavior Are Represented by Different Dorsomedial Hypothalamic Sites: A Three-Dimensional Functional Topography Study in Conscious Rats. J Neurosci. 2017;37:6956–6971. doi: 10.1523/JNEUROSCI.0100-17.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webb GP, Jagot SA, Jakobson ME. Fasting-induced torpor in Mus musculus and its implications in the use of murine models for human obesity studies. Comp Biochem Physiol A Comp Physiol. 1982;72:211–219. doi: 10.1016/0300-9629(82)90035-4. [DOI] [PubMed] [Google Scholar]
- Weiss B, Laties VG. Behavioral thermoregulation. Science. 1961;133:1338–1344. doi: 10.1126/science.133.3461.1338. [DOI] [PubMed] [Google Scholar]
- Wit A, Wang SC. Temperature-sensitive neurons in preoptic-anterior hypothalamic region: effects of increasing ambient temperature. Am J Physiol. 1968;215:1151–1159. doi: 10.1152/ajplegacy.1968.215.5.1151. [DOI] [PubMed] [Google Scholar]
- Yahiro T, Kataoka N, Nakamura Y, Nakamura K. The lateral parabrachial nucleus, but not the thalamus, mediates thermosensory pathways for behavioural thermoregulation. Sci Rep. 2017;7:5031. doi: 10.1038/s41598-017-05327-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yarmolinsky DA, Peng Y, Pogorzala LA, Rutlin M, Hoon MA, Zuker CS. Coding and Plasticity in the Mammalian Thermosensory System. Neuron. 2016;92:1079–1092. doi: 10.1016/j.neuron.2016.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida K, Konishi M, Nagashima K, Saper CB, Kanosue K. Fos activation in hypothalamic neurons during cold or warm exposure: projections to periaqueductal gray matter. Neuroscience. 2005;133:1039–1046. doi: 10.1016/j.neuroscience.2005.03.044. [DOI] [PubMed] [Google Scholar]
- Yoshida K, Li X, Cano G, Lazarus M, Saper CB. Parallel preoptic pathways for thermoregulation. J Neurosci. 2009;29:11954–11964. doi: 10.1523/JNEUROSCI.2643-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu S, Qualls-Creekmore E, Rezai-Zadeh K, Jiang Y, Berthoud HR, Morrison CD, Derbenev AV, Zsombok A, Munzberg H. Glutamatergic Preoptic Area Neurons That Express Leptin Receptors Drive Temperature-Dependent Body Weight Homeostasis. J Neurosci. 2016;36:5034–5046. doi: 10.1523/JNEUROSCI.0213-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaretskaia MV, Zaretsky DV, Shekhar A, DiMicco JA. Chemical stimulation of the dorsomedial hypothalamus evokes non-shivering thermogenesis in anesthetized rats. Brain Res. 2002;928:113–125. doi: 10.1016/s0006-8993(01)03369-8. [DOI] [PubMed] [Google Scholar]
- Zhang W, Sunanaga J, Takahashi Y, Mori T, Sakurai T, Kanmura Y, Kuwaki T. Orexin neurons are indispensable for stress-induced thermogenesis in mice. J Physiol. 2010;588:4117–4129. doi: 10.1113/jphysiol.2010.195099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Kerman IA, Laque A, Nguyen P, Faouzi M, Louis GW, Jones JC, Rhodes C, Munzberg H. Leptin-receptor-expressing neurons in the dorsomedial hypothalamus and median preoptic area regulate sympathetic brown adipose tissue circuits. J Neurosci. 2011;31:1873–1884. doi: 10.1523/JNEUROSCI.3223-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang YH, Yanase-Fujiwara M, Hosono T, Kanosue K. Warm and cold signals from the preoptic area: which contribute more to the control of shivering in rats? J Physiol. 1995;485(Pt 1):195–202. doi: 10.1113/jphysiol.1995.sp020723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao ZD, Yang WZ, Gao C, Fu X, Zhang W, Zhou Q, Chen W, Ni X, Lin JK, Yang J, et al. A hypothalamic circuit that controls body temperature. Proc Natl Acad Sci U S A. 2017;114:2042–2047. doi: 10.1073/pnas.1616255114. [DOI] [PMC free article] [PubMed] [Google Scholar]