Targeted transcriptional repression using a chimeric TALE-SRDX repressor protein (original) (raw)
Abstract
Transcriptional activator-like effectors (TALEs) are proteins secreted by Xanthomonas bacteria when they infect plants. TALEs contain a modular DNA binding domain that can be easily engineered to bind any sequence of interest, and have been used to provide user-selected DNA-binding modules to generate chimeric nucleases and transcriptional activators in mammalian cells and plants. Here we report the use of TALEs to generate chimeric sequence-specific transcriptional repressors. The dHax3 TALE was used as a scaffold to provide a DNA-binding module fused to the EAR-repression domain (SRDX) to generate a chimeric repressor that targets the RD29A promoter. The dHax3.SRDX protein efficiently repressed the transcription of the RD29A::LUC transgene and endogenous RD29A gene in Arabidopsis. Genome wide expression profiling showed that the chimeric repressor also inhibited the expression of several other genes that contain the designer TALE-target sequence in their promoters. Our data suggest that TALEs can be used to generate chimeric repressors to specifically repress the transcription of genes of interest in plants. This sequence-specific transcriptional repression by direct on promoter effector technology is a powerful tool for functional genomics studies and biotechnological applications.
Similar content being viewed by others
Introduction
Plants are sessile organisms and rely on a wide array of molecular mechanisms to control and adjust their adaptive responses to developmental and environmental cues. Understanding these responses requires that we understand the functioning of plant genes. Although such understanding has become increasingly possible with rapid advances in DNA sequencing technologies, the challenge is to turn information about gene sequence into knowledge about gene function. The analysis of the completed plant genome sequences revealed that transcriptional regulation plays a very pronounced role in plants (Riechmann et al. 2000). The Arabidopsis genome is predicted to contain 25,498 genes, and more than 2000 of these are transcription factors. Consequently, the percentage of transcription factor genes relative to the whole gene content is higher in Arabidopsis (6–10%) than in organisms with genomes of similar size including Drosophila melanogaster (4.7%) and Caenorhabditis elegans (3.6%) (Riechmann et al. 2000). Plants use many different transcription factors to modulate gene expression and achieve the correct spaciotemporal control of gene activation and repression. Researchers use factors that activate or repress gene expression to understand gene function and phenotypic effects (Krogan and Long 2009). Transcriptional repression effects occur at several levels and involve active and passive repressors. Active repressors exhibit an intrinsic repression activity because they have defined repression domains that can interfere with the formation of the transcription pre-initiation complex and basal transcription (Krogan and Long 2009; Mahfouz 2010). Passive repressors, in contrast, interfere with transcriptional activators and prevent their binding to the DNA to cause transcription repression (Kazan 2006; Krogan and Long 2009).
Chimeric repressors, which have been used to study gene function in animal and plant systems, have a DNA-binding domain or a transcription factor fused to a variety of repression domains (Beerli et al. 2000; de Haan et al. 2000; Ohta et al. 2001). Transcription repressors that contain the EAR-repression domain (ERF-associated amphiphilic repression) were the first to be identified in plants (Kagale and Rozwadowski 2011; Ohta et al. 2001). The exact mechanism of function of the transcriptional repressors is not known but may involve interaction with co-repressors (Szemenyei et al. 2008). Researchers have fused the EAR motif to a number of transcription factors to generate dominant chimeric repressors (Hiratsu et al. 2003; Ikeda and Ohme-Takagi 2009; Matsui et al. 2008; Mito et al. 2010; Tsutsui et al. 2009). The application of this technology, however, is limited to the pre-existing transcription factors. In the chimeric repressor silencing technology or “CRES-T” system, the transcription factors were used as a DNA-binding module to repress the transcription factor’s target genes but because many of the transcription factor targets are not known, it is difficult or impossible to achieve repression to specific genes. This repression technology would be much more powerful if it could be applied to a user-selected and defined single locus or multiple loci.
A chimeric transcription repressor or activator, in principle, is composed of a DNA-binding domain, an activation or repression domain, and a nuclear-localization signal (Guan et al. 2002; Yaghmai and Cutting 2002). Exploiting this fundamental principle to generate chimeric repressors or activators with genome-wide applicability requires a DNA-binding domain that can be modified and adapted to selectively and specifically bind to any DNA sequence in the genome. This kind of DNA-binding domain exists in phytopathogenic bacteria in the genus Xanthomonas. Phytopathogenic Xanthomonas spp. use the type III secretion system to inject transcription activator-like effectors (TALEs) into plant cells (Boch and Bonas 2010). These TALEs, which act as virulence factors, translocate to the nucleus and function as transcription factors by binding to their DNA targets in the promoter regions and reprograming the gene expression of the host (Kay et al. 2007; Romer et al. 2007). The repeats of the DNA-binding domains (DBD) dictate the specificity of TALEs to their DNA target (Boch et al. 2009; Moscou and Bogdanove 2009). The repeats are nearly identical, and each repeat is composed of 34 or 35 amino acids and has a repeat variable diresidue (RVD) at positions 12 and 13. There is a one-to-one correspondence between each RVD and a single nucleotide in the DNA target, and hence the identity and the sequence of the RVDs determine the specificity of each TALE to its target (Boch et al. 2009; Moscou and Bogdanove 2009).
The Brassicaceae pathogen X. campestris pv. armoraciae strain 5 produces Hax3, which is a member of the AvrBs3 family of TALEs (Kay et al. 2005). Hax3 TALE protein has all the structural features of TALEs including a DNA-binding domain composed of 11.5 repeat units, a nuclear-localization signal, and an acidic activation domain. The chief advantage of TALEs for the design of transcriptional repressors or activators is their versatility; they could be used, in principle, to target any functional domain to any gene locus by fusing the functional domain to an appropriately modified TALE DBD (Christian et al. 2010; Li et al. 2011a; Mahfouz et al. 2011; Morbitzer et al. 2010; Weber et al. 2011). Thus, TALEs could be used to take advantage of the entire repertoire of the transcription-regulatory domains to achieve the desired level and type of regulation for any gene of interest. Hax3 recognizes a 12-bp DNA-binding element and has been used to generate a de novo engineered hybrid nuclease capable of generating double-strand breaks in vitro and in vivo in plants (Mahfouz et al. 2011).
Here, we report the generation of a chimeric transcriptional repressor that targets an element in the RD29A promoter. Our data demonstrate that the dHax3.SRDX chimeric repressor selectively suppresses the expression of the RD29A::LUC transgene and the RD29A endogenous gene as well as several other genes with the target element in their promoters in Arabidopsis. This gene repression technology will be useful for functional genomics studies in crops and should have significant biotechnological applications.
Results
Design and generation of the dHax3.SRDX TALE-based transcriptional repressor to target an RD29A promoter element
To generate a chimeric TALE-based repressor, we selected the Hax3 TALE as a scaffold and the SRDX repression domain; the Hax3 TALE provided a precise and selective DNA-binding module while the SRDX repression domain (Heyl et al. 2008) provided transcriptional repression. The natural Hax3 TALE binds to a 12-bp DNA box that contains the TACACCCAAACAT nucleotide sequence. The 11.5 repeats of dHax3 were modified to bind a 12-bp sequence (TCCCTTTATCTCT) in the RD29A promoter; this sequence overlaps by two nucleotides with the abscisic acid-responsive element (ABRE) (Fig. 1a) (Mundy et al. 1990). The dHax3 recognition box is preceded by the T nucleotide, 13-bp binding box, which was shown to be essential for TALEs binding to their targets (Boch et al. 2009). To generate the hybrid dHax3.SRDX repressor, we fused in frame the nucleotide sequence corresponding to 12 aa of the SRDX dominant repressor (LDLDLELRLGFA) to the full-length dHax3 sequence (Fig. 1b) (Hiratsu et al. 2003; Matsui and Ohme-Takagi 2010). As we previously reported, the dHax3 fragments were assembled and fused to generate the full-length dHax3(Mahfouz et al. 2011), which was then fused at its C-terminus to a fragment containing the SRDX domain. The dHax3.SRDX sequence was cloned in the pENTR/D gateway vector, and the construct was confirmed by sequencing. The in silico-designed chimeric repressor, herein referred to as dHax3.SRDX, has 972 aa residues and an estimated molecular mass of 102.77 kDa. The dHax3.SRDX was subsequently sub-cloned in the pET32a gateway compatible protein expression vector by LR reaction to generate thioredoxin.6His fusion protein (Trx.His.dHax3.SRDX). The bacterially expressed Trx.6His.dHax3.SRDX was shown to migrate in SDS polyacrylamide gel electrophoresis according to its expected size (Fig. 2a).
Fig. 1
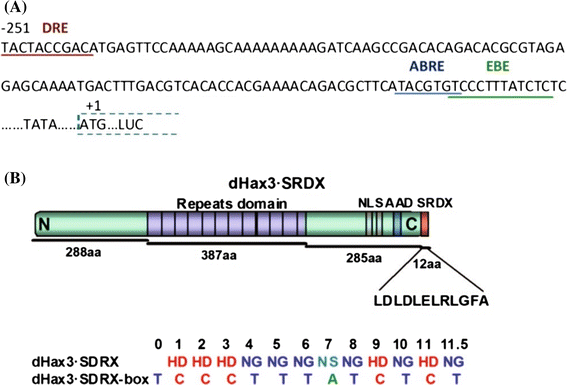
Sequence of RD29A promoter showing effector-binding elements and schematic diagram of dHax3.SRDX. a The sequence of the RD29A promoter showing the dHax3.SRDX-binding element (EBE), the dehydration-responsive element (DRE), and the ABA responsive element (ABRE). b Structural representation of dHax3.SRDX showing the N-terminal domain, repeat domain, a linker domain, a nuclear-localization signal, acidic activation domain, and the SRDX domain with its amino acid sequence shown below. Also shown below are the repeat variable diresidues (RVDs) of dHax3.SRDX with the sequence of its DNA-binding element (EBE)
Fig. 2
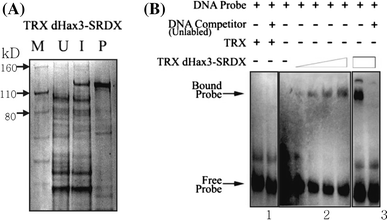
Purification of dHax3.SRDX and EMSA. a Bacterial expression and purification of dHax3.SRDX protein. Lane 1 shows the protein marker, lane 2 shows the un-induced cell extract, lane 3 shows the IPTG-induced cell extract, and lane 4 shows the purified dHax3.SRDX fraction. b EMSA of dHax3.SRDX binding to its target sequence. Panel 1: the thioredoxin tag does not bind to the biotin-labeled probe. Panel 2: the dHax3.SRDX protein binds to the biotin-labeled probe sequence in a concentrationdependent manner. Panel 3: the unlabeled probe competitively reduces the binding of the biotin-labeled probe to the dHax3.SRDX protein
Hax3.SRDX chimeric repressor protein binds to its target sequence in vitro
To examine the ability of the bacterially expressed dHax3.SRDX chimeric repressor to bind to its DNA target, we performed electrophoretic mobility shift assays (EMSA) using the bacterially expressed dHax3.SRDX and a biotin-labeled double-stranded oligo-nucleotide containing the 12-bp DNA target sequence preceded by the T nucleotide. The assays showed that the dHax3.SRDX was capable of binding to its target DNA sequence and that the binding was competitively reduced by the addition of the same but unlabeled double-stranded oligo-nucleotides. The results suggest that the fusion of the SRDX domain did not affect the ability of the dHax3.SRDX chimeric repressor protein to bind to its target site (Fig. 2b).
Expression of dHax3.SRDX repressor results in repression of the RD29A promoter in vivo
To test whether the dHax3.SRDX causes transcriptional repression of the luciferase transgene driven by the RD29A promoter and the endogenous RD29A gene in their chromosomal contexts, we generated stable transgenic lines of Arabidopsis ectopically expressing Hax3.SRDX under the cauliflower mosaic virus 35S promoter in a background containing the RD29A::LUC transgene (ecotype C24, referred to as the WT). Several transgenic lines over-expressing 35S::dHAX3.SRDX were selected on MS nutrient medium supplemented with hygromycin at 30 mg/L. Homozygous T3 plants expressing 35S::dHAX3.SRDX were used to analyze the transcriptional repression of the RD29A promoter in both the luciferase transgene and the endogenous RD29A gene.
RD29A is one of the most well studied promoters (Yamaguchi-Shinozaki and Shinozaki 1994). This promoter contains the ABRE and dehydration responsive element (DRE) (Narusaka et al. 2003). The promoter can be activated by osmotic and cold stresses as well as by ABA(Ishitani et al. 1997). Because the DRE element can be activated by osmotic and cold stresses but not by ABA, this promoter has been used to analyze ABA-dependent and ABA-independent signaling (Ishitani et al. 1997; Xiong et al. 1999a). The DNA-binding box of the dHax3.SRDX hybrid repressor overlaps with 2 bp of the ABRE in the RD29A promoter and no overlap with the DRE box (Fig. 1a). We tested the transcriptional repression of the RD29A promoter by single or combined treatments of Arabidopsis plants expressing dHax3.SRDX with ABA and cold. Six lines of Arabidopsis over-expressing dHax3.SRDX showed significant repression of the endogenous RD29A gene and the luciferase transgene (Fig. 3a, b). The treatment of WT with 100 μm ABA resulted in the activation of the RD29A promoter and strong luminescence as shown in Fig. 3a. In contrast to the WT, ectopic expression of the dHax3.SRDX led to the suppression of the RD29A promoter, resulting in minimal or no luminescence. Our data show that the dHax3.SRDX strongly represses the RD29A promoter.
Fig. 3
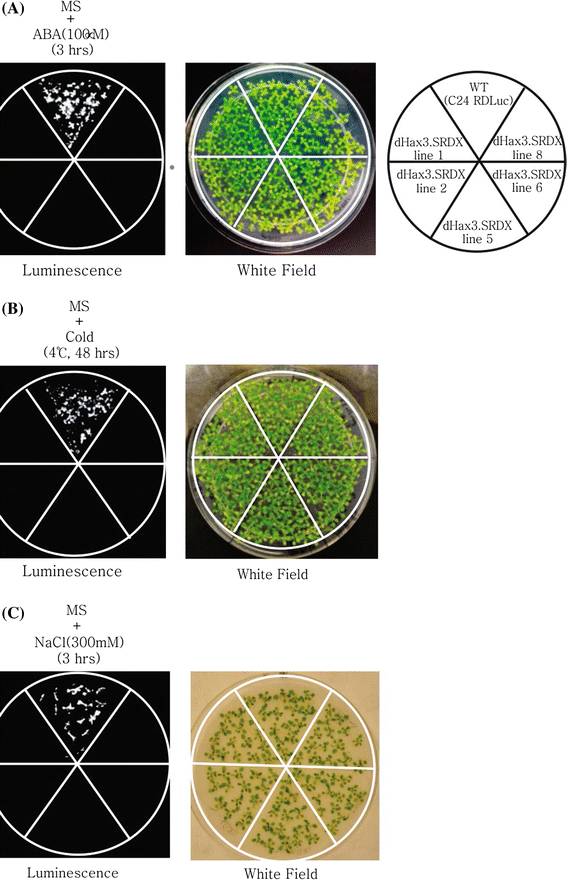
Transcriptional repression of RD29A::LUC in dHax3.SRDX transgenic plants. a WT (C24 RD29A::LUC) and dHax3.SRDX expressing lines 1, 2, 5, and 6 grown on MS agar plates for 1 week were treated with 100 μm ABA for 3 h. Luminescence images were taken after the ABA treatment (left). The right panel shows the label details of dHax3.SRDX expressing lines. b Two-week old plants from the same lines as (a) were treated with cold (4°C) for 48 h. Luminescence images were taken after the cold treatment (left)
Because the RD29A promoter is also induced by salt, we tested the ability of the dHax3.SRDX chimeric repressor to repress the RD29A promoter after salt treatment (300 mM NaCl, 3 h). Our data show that the dHax3.SRDX chimeric repressor can significantly repress the RD29A promoter after salt treatment (Fig. 3c).
It was previously reported that low temperature and ABA treatments have additive effects on the transcriptional induction of the RD29A::LUC transgene and the RD29A endogene (Xiong et al. 1999b). Because cold and ABA activate the RD29A promoter via two different signaling pathways, we examined the ability of the dHax3.SRDX chimeric repressor to simultaneously repress gene transcription activated by both signaling pathways. We performed a combined treatment of cold (48 h at 4C°) and ABA (100 μm for 3 h) on the same dHax3.SRDX lines. Our data indicate that the designer chimeric repressor is capable of repressing the LUC expression in response to two independent signaling pathways (Fig. 4a).
Fig. 4

The ability of dHax3.SRDX to repress RD29A::LUC after a combined treatment with ABA and cold and the transcriptional repression is due to the presence of the SRDX domain. a One-week-old plants grown on MS agar plates were treated with cold (4°C, 48 h) followed by ABA (100 μm, 3 h). Luminescence images were taken after the combined treatments. All dHax3.SRDX expressing lines showed a significant repression of RD29A::LUC after the combined treatments. WT (control) luminescence indicates the transcriptional activities of the RD29A::LUC promoter. b uidA repression is mediated by the SRDX repression domain. In planta functional analysis of dHax3 activator and dHax3.SRDX chimeric repressor. The constructs uidA driven by Bs3 WT or Bs3_∆_dHax3 promoters were co-delivered via Agrobacterium tumefaciens into N. benthamiana leaves and the GUS assays were performd 48 hpi
Since the repression might be mediated merely by the binding of the dHax3 effector, to its promoter, and not by SRDX repression domain of dHax3.SRDX repressor, we tested this possibility by functional analysis in N. benthamiana transient assays. We inserted the dHax3.SRDX effector-binding element in the Bs3 minimal promoter to generate ∆d_Hax3Bs3::uidA_ and co-delivered this construct or its WT version (Bs3::uidA) with 35S::dHax3 or 35S::dHax3.SRDX, respectively (Morbitzer et al. [2011](/article/10.1007/s11103-011-9866-x#ref-CR33 "Morbitzer R, Elsaesser J, Hausner J, Lahaye T (2011) Assembly of custom TALE-type DNA binding domains by modular cloning. Nucleic Acids Res. doi: 10.1093/nar/gkr151
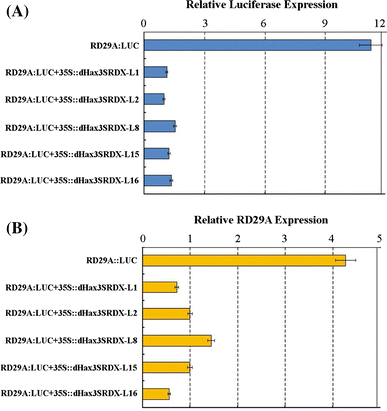
")). Our data clearly indicate that the dHax3 activates the ∆d_Hax3Bs3::uidA_ expression (Fig. [4](/article/10.1007/s11103-011-9866-x#Fig4)b, panel 2). dHax3.SRDX, however, strongly repressed the expression of the ∆d_Hax3Bs3::uidA_ (Fig. [4](/article/10.1007/s11103-011-9866-x#Fig4)b, panel 7).To confirm the luminescence data at the molecular level, we analyzed the abundance of the luciferase and RD29A transcripts using real-time RT-PCR. The endogenous RD29A gene and RD29A::LUC transgene were induced and highly expressed in the WT control plants treated with 100 μm ABA for 3 h (Fig. 5a, b). The dHax3.SRDX chimeric repressor lines, however, showed substantially lower levels of the RD29A and LUC transcripts, indicating that their expression was repressed. These data are consistent with the bioluminescence data and indicate that the dHax3.SRDX protein functions as a strong chimeric repressor in planta.
Fig. 5
Q-PCR analysis of RNA extracted from ABA-treated C24 RD29A:: LUC WT control and dHax3.SRDX expressing lines. Experiments were normalized relative to actin2, and two or more biological replicates were tested with similar results
Genome-wide effect of the dHax3.SRDX chimeric repressor on gene expression
Several reports have shown that TALEs can be used as DNA-binding modules with high specificity and precision (Li et al. 2011a; Morbitzer et al. 2010; Zhang et al. 2011). TALEs with a higher number of repeats and longer DNA targets might be more specific than those with shorter repeats and DNA targets (Boch et al. 2009). To test the specificity of the dHax3.SRDX chimeric repressor, we used the Patmatch algorithm (http://is.gd/g31n9) to scan the Arabidopsis genome for sequences that contain matching boxes specifically in the promoter regions. We identified in the −500 bp upstream sequence only one perfect match that corresponds to the RD29A promoter (AT5G52310). The code of the RVDs DNA-binding specificities, however, is somewhat degenerate. For example, the NS RVD could bind to A, C, G, or T nucleotide, with higher affinity to A and reduced affinity to C, G, and T nucleotides (Boch et al. 2009; Miller et al. 2011). We scanned the Arabidopsis genome for potential dHax3.SRDX binding to DNA boxes that include TCCCTTTATCTCT, TCCCTTTCTCTCT, TCCCTTTGTCTCT, or TCCCTTTTTCTCT in the −500 bp upstream promoter region. We found one perfect match, in the RD29A gene promoter that includes the TCCCTTTATCTCT box, and 20 other possible binding hits on either strand, including C, G, or T nucleotide corresponding to the 7th repeat in the tandem repeat region of the DNA-binding module. Moreover, we identified 70 additional possible targets when we extended the search to −3,000 bp upstream promoter regions and 82 binding hits in the genes including introns and UTR regions.
To examine the selectivity and specificity of the dHax3.SRDX repressor, we studied the expression patterns of these genes in the dHax3.SRDX transgenic lines. We performed a microarray experiment to investigate the transcript levels of these potential targets in the WT control and the dHax3.SRDX lines. We found that the gene hits in the −500 bp upstream promoter regions were under-expressed by at least twofold across all dHax3.SRDX lines. The repressed genes that contain the binding box (ATX1, AT2G07280, AT5G08390, AAP6, AT2G41230, ATTAP1, AT5G45580, LTI78) are shown in blue boxes and their relative distance from the transcriptional start is shown at left and the nucleotides that bind to the NS 7th repeat are shown at the right (Fig. 6). The distance from the transcriptional start seems to be important since in most of the repressed genes the target sequence lies within the −500 bp of the promoter region. Interestingly, in most of the lines we found >20-fold repression of two gene hits (AT2G41230 and AT2G07280), which have the target sequence within −3,000 bp upstream promoter regions. The strong repression of these two genes might indicate the importance of the chromatin environment, the distance between the binding sequence and transcriptional start or the presence of multiple potential binding sequences that may lead to the recruitment of more repressor proteins or tighter binding. We also noticed that one predicted target (AAP6) was over-expressed by twofold (Fig. 6). The over-expression of this gene hit might be an indirect consequence of the repression of one or more of the repressed targets. Importantly, we did not find transcriptional modulation in genes that do not have the target sequence in their promoters. Our data demonstrate that the dHax3.SRDX chimeric repressor inhibits the expression of genes with the designed target sequences in their promoters, and suggest that the off-target effects can be predicted and avoided in the design of such chimeric repressors.
Fig. 6
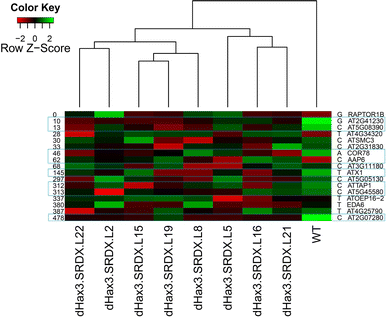
Microarray expression analysis of dHax3.SRDX over-expression lines showing the repression of target genes containing the TCCCTTT[A/C/G/T]TCTCT DNA-binding box within the promoter region. Samples (columns) were clustered using Euclidean distance and complete linkage. Genes (rows) with motif variations were sorted in ascending order according to the distance to the 5′UTR start. Genes in the blue frames (discussed in text) compared to the WT control are showing transcriptional repression across all tested dHax3.SRDX lines
Discussion
In the present study, we demonstrate that the TALE-based dHax3.SRDX chimeric protein designed to target the RD29A promoter can repress the RD29A::LUC transgene and RD29A endogenous gene in a chromosomal context and thus can be used as an effective artificial repressor. The repeats of the dHax3 DNA-binding domain were modified so that it can bind to a DNA box in the RD29A promoter. The dHax3 was fused in frame to the SRDX domain to generate the dHax3.SRDX. Our data suggest that the fusion of the SRDX domain does not affect the conformation of the dHax3 since the fusion protein was capable of binding to its target sequence in vitro (Fig. 2b). dH_ax3.SRDX_, driven by the CaMV 35S promoter, was ectopically expressed in Arabidopsis containing the RD29A::LUC transgene. The RD29A promoter can be activated by a variety of environmental stresses including salt, drought, cold, and also the phytohormone ABA (Chinnusamy et al. 2002; Hua et al. 2006; Ishitani et al. 1997; Kasuga et al. 2004). To test the ability of the dHax3.SRDX to repress its target genes, we used luciferase imaging to analyze LUC expression in several Arabidopsis lines expressing dHax3.SRDX. Arabidopsis dHax3.SRDX lines grown on MS media and treated with 100 μm ABA for 3 h showed a significant suppression of the luciferase signal relative to the wild type control, indicating the repression of the RD29A::LUC transgene by the dHax3.SRDX chimeric repressor protein (Fig. 3a). The dHax3.SRDX also suppressed the expression of RD29A::LUC transgene in response to cold or salt treatments (Fig. 3b, c). Moreover, our data demonstrate the ability of the dHax3.SRDX to suppress the expression of the RD29A::LUC transgene after a combined ABA and cold treatment, indicating its ability to simultaneously inhibit the activities of multiple transcriptional activators, which integrate signals from different signaling pathways (Fig. 4).
Our data indicate that the dHax3.SRDX chimeric repressor protein suppressed the transcription of a RD29A::LUC transgene and RD29A endogenous gene in a chromatin context. These data demonstrate the versatility of the repression mechanism and the ability of the TALE-based repressors to suppress the expression of more than one gene simultaneously. It should be noted that the transient and stable expression of TALE protein alone did not suppress the expression of RD29A promoter indicating that the repression is due to the presence of the SRDX repression domain (Fig. 4b) (Mahfouz et al. 2011). Moreover, the use of the strong 35S promoter might not be necessary to obtain gene repression effects. A lower ratio of repressor to activator was able to cause efficient suppression of the reporter gene in Arabidopsis transient assays (Fujimoto et al. 2000). Our data demonstrate a significant reduction in the expression of the endogenous RD29A gene, although the repression of RD29A was generally less than that of the RD29A::LUC transgene as evidenced by real-time RT-PCR (Fig. 5a, b). The ability of the TALE repressor to bind to its target might be affected by the chromatin status or the distance of the TALE binding motif from the transcriptional start site may influence its repression activity.
Because the dHax3.SRDX chimeric repressor contains the NS-type RVD at the 7th position, we tested the ability of the repressor to repress genes that contain binding boxes where the 7th nucleotide in the DNA target is A, C, G, or T. We identified 21 potential hits in the −500 bp upstream promoter region and 70 more gene hits in the −3,000 bp upstream region. It should be mentioned that these are not off-targets since they have the TALE-SRDX DNA binding box according to the TALE DNA binding code. Off-targets are defined as the genes whose level of expression is modulated due to the expression of TALE-SRDX and may contain a non-perfect match of the DNA binding box. Our microarray data indicate that the expression of 5 gene hits in the −500 bp regions is repressed in all tested lines. The repression levels vary among the targets perhaps because of the chromosomal position effect or the distance from the transcription start site. Moreover, two gene hits (AT2G41230 and AT2G07280) containing the dHax3.SRDX DNA-binding box on the opposite strand in −3,000 bp upstream region, show significantly higher levels of repression. These two genes contain multiple repeats with similar sequence to the DNA-binding box TCCCTTT[ACGT]TCTCT within −5 kb upstream of the transcription start site. In fact, there are eight repeats of 50-bp motifs with a DNA sequence similar to that of the DNA binding box (TCCCTTGTAAATCATTCT). The high level of repression of these two genes could be due to their local chromatin environments, the presence of the DNA-binding box on the opposite strand, the distance of the DNA-binding box relative to the transcriptional start site, or the presence of multiple copies of a motif similar to the DNA-binding box. The presence of multiple sequences similar to the DNA-binding box might help recruit a large number of the chimeric repressor to result in strong transcriptional repression. It appears that the transcriptional repression occurred regardless of the dHax3.SRDX binding orientation because the presence of the EBE on the forward or reverse strand relative to the gene resulted in repression activities. This is consistent with proposed models of SRDX-mediated repression where SRDX might function by disrupting the formation of the transcription complex or recruiting repressors and co-repressors. These data indicate that the chromatic status may affect the binding of the dHax3.SRDX or the function of the SRDX domain. Moreover, many details are unknown concerning the nature of TALE binding, including the chromatin environments and optimal distance from the transcription start site. These details are of paramount importance for the future design of effective TALE-based activators or repressors.
Because de novo TALEs (dTALEs) with higher repeat units might be expected to possess higher target specificity (Morbitzer et al. [2011](/article/10.1007/s11103-011-9866-x#ref-CR33 "Morbitzer R, Elsaesser J, Hausner J, Lahaye T (2011) Assembly of custom TALE-type DNA binding domains by modular cloning. Nucleic Acids Res. doi: 10.1093/nar/gkr151
")), dHax3.SRDX chimeric repressors should be designed with at least 18 repeat units if a single gene is to be targeted. However, designing dTALEs with variable repeat lengths and RVDs with degenerate nucleotide binding would be useful for analyzing the nature and details of the binding and activity of dTALE activators and repressors. Several parameters for TALE target designs were suggested based on the study of positional biases, neighbor effects, and the binding of RVDs to nucleotides in the overall target (Cermak et al. [2011](/article/10.1007/s11103-011-9866-x#ref-CR5 "Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, Baller JA, Somia NV, Bogdanove AJ, Voytas DF (2011) Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res 39:e82. doi:
10.1093/nar/gkr218
")). According to these suggestions, there is strong bias against T at position 1, A at position 2, and G at position 3 and the next to the last position, and a moderate bias for T at the last position (Cermak et al. [2011](/article/10.1007/s11103-011-9866-x#ref-CR5 "Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, Baller JA, Somia NV, Bogdanove AJ, Voytas DF (2011) Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res 39:e82. doi:
10.1093/nar/gkr218
")). These and other suggestions may be useful for designing effective TALE DNA-binding modules. Moreover, several TALEs repeats assembly protocols were recently reported, and these should facilitate the efficient design and generation of TALEs for user-selected targets (Cermak et al. [2011](/article/10.1007/s11103-011-9866-x#ref-CR5 "Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, Baller JA, Somia NV, Bogdanove AJ, Voytas DF (2011) Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res 39:e82. doi:
10.1093/nar/gkr218
"); Li et al. [2011b](/article/10.1007/s11103-011-9866-x#ref-CR23 "Li T, Huang S, Zhao X, Wright DA, Carpenter S, Spalding MH, Weeks DP, Yang B (2011b). Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes. Nucleic Acids Res 39:6315–6325"); Morbitzer et al. [2011](/article/10.1007/s11103-011-9866-x#ref-CR33 "Morbitzer R, Elsaesser J, Hausner J, Lahaye T (2011) Assembly of custom TALE-type DNA binding domains by modular cloning. Nucleic Acids Res. doi:
10.1093/nar/gkr151
"); Zhang et al. [2011](/article/10.1007/s11103-011-9866-x#ref-CR49 "Zhang F, Cong L, Lodato S, Kosuri S, Church GM, Arlotta P (2011) Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription. Nat Biotechnol 29:149–153")).Our present study suggests that a single Hax3.SRDX protein can be designed to silence a single gene or a group of genes sharing DNA-binding elements in the promoter regions, making it possible to functionally characterize single genes, multiple genes or gene families. It remains to be determined whether the TALE-based repression technology could be a useful alternative to RNA interference (RNAi), a widely used technology that suffers from a variety of disadvantages and pitfalls. The demonstration of the effectiveness of the TALE-based repression technology to different promoters, gene families and chromosomal loci compared to RNAi is required to draw a conclusion. The primary disadvantage of RNAi is the off-target effects, i.e., sequence complementarities between an siRNA and the mRNA involving fewer than 10 nucleotides can lead to the reduction of expression (Birmingham et al. 2006). dTALE repressors, in contrast, can be designed to bind specifically to one genomic target. RNAi-based silencing exhibits variable effects depending on the target gene, the targeted region of the transcript, and even between different lines carrying the same RNAi construct (Wang et al. 2005). Moreover, RNAi phenotypes can be lost over several generations. Because RNAi suffers from problems related to efficacy, specificity and stability, silencing by TALE chimeric repressors might represent an attractive alternative with enormous potential for basic and applied research. For example, silencing by TALE chimeric repressors could be used for the genetic manipulation of commercially important crop plants (Mahfouz and Li 2011). This gene repression technology may also be applied to crop plants to identify genes responsible for traits of interest including biotic and abiotic stress resistance, nitrogen use efficiency, high yield, and control of fertility.
Materials and methods
dHax3.SRDX design and vector construction
pET32a.dHax3.SRDX was generated by performing LR recombination reactions between the dHax3.SRDX entry clone and the gateway-compatible pET32a expression vector according to manufacturer’s protocol (Invitrogen). The expression clone was transformed into Escherichia coli BL21, and the protein expression was induced at 25°C for 5 h with 1 mM isopropyl β-d-1thiogalactopyranoside (IPTG). The TRX.dHax3.SRDX protein was purified using Qiagen Ni–NTA agarose resin according to the manufacturer’s instructions. LR reactions were performed between the pENTR/dHax3.SRDX entry clone and the pMDC32 gateway compatible binary vector to generate the final construct for in planta analysis. The pMDC32/dHax3.SRDX clone was confirmed by sequencing and transformed into Agrobacterium tumefaciens GV3101 and then to Arabidopsis plants by the floral dipping method (Zhang et al. 2006).
Electrophoretic mobility shift assay (EMSA)
EMSA was performed using the Lightshift Chemiluminescent kit (Pierce) and following the manufacturer’s instructions. For dHax3.SRDX DNA-binding studies, the TRX.dHax3.SRDX protein was purified as described above, and the protein concentration was measured with a Bradford protein assay kit (Bio-Rad). Complementary pairs of 5′ biotin-labeled or non-labeled oligonucleotides were annealed and used as probes for the binding studies. EMSA binding-reaction buffer contained 12 mM Tris–HCL, 60 mM KCl, 1 mM DTT, 2.5% glycerol, 5 mM MgCl2, 50 ng/μL poly(dI.dC), 0.05% Nonidet P-40, 0.2 mM EDTA, 35 fmol biotin-labeled DNA, 10 pmol unlabeled DNA, and 30–480 fmol TRX.His fusion proteins. The EMSA binding reactions were kept on ice for 10 min before the biotin-labeled probe was added. Gel electrophoresis was performed using 8% Tris/Borate/EDTA (TBE) native ready-made gels from Invitrogen. Blotting was performed on a positively charged nylon membrane, and the membrane was cross-linked using CL-1000 UV cross-linker for 30 s.
Agrobacterium and Arabidopsis transformation
Arabidopsis thaliana (ecotype C24) expressing the chimeric RD29A::LUC reporter gene (referred to as the wild type or WT in this study) was used for the in planta analysis of the dHax3.SRDX repressor. pMDC32.dHax3.SRDX was transformed into Agrobacterium strain GV3101 by the freeze-and-thaw method. A transformed single colony of Agrobacterium GV3101 was used to transform Arabidopsis C24 RD29A::LUC plants as described previously (Zhang et al. 2006). Transformants were selected on MS agar plates supplemented with 30 mg/L of hygromycin B. T3 plants were used for the transcriptional repression analysis.
Assay of RD29A::LUC expression
WT (C24 RD29A::LUC) and dHax3.SRDX over-expression lines were grown on MS agar media for 1 week. Seedlings were sprayed with 100 μm ABA for 3 h or kept at 4°C for 24 h (cold treatment) or incubated on filter papers soaked with 300 mM NaCl (salt treatment) (Chinnusamy et al. 2002). Plates with treated plants were sprayed with Luciferin at room temperature and kept in the dark for 5 min before they were placed in the dark chamber of the CCD camera (PIXIS 2048). Image quantification was carried out using the Winview software supplied by the manufacturer.
RNA isolation and quantitative real-time PCR
The expression levels for the LUC transgene and RD29A endogenous gene were analyzed in the treated and untreated WT and dHax3.SRDX lines. Total RNA was purified using the RNeasy kit (Qiagen SA). For the cDNA synthesis, 1 μg of total RNA was used for the reverse transcription using reverse trancriptase (Life Technologies, Applied Bioscience).
All cDNAs were measured by real-time PCR. Quantitative PCR was performed in an ABI prism 7,900 using a SYBR Green. The β-actin gene served as the internal control for normalization of data. All experiments are done in at least duplicate.
Microarray analysis
The RNeasy plant mini kit (Qiagen) was used to extract total RNAs from 2-week-old WT and dHax3.SRDX over-expression lines that were grown on MS agar plates and treated with 100 μM ABA. The total RNA samples were pretreated with RNase-free DNase I and cleaned using the Plant Total RNA isolation kit (Qiagen). Probe preparation, hybridization to the Arabidopsis ATH1 Gene expression arrays (Agilent Technologies), and subsequent processing steps were performed according to the manufacturer’s procedures. Raw signals were extracted from the scanned images; the background was subtracted and the data were normalized using the Feature Extraction Software (10.7.1.1; Agilent technologies). These signals were then log 2 transformed and subjected to percentile shift-based normalization and median-based baseline transformation using Genespring GX (Agilent Technologies). WT (C24 RD29A::LUC) plant values were used as the baseline to calculate the intensity ratio/fold changes of the dHax3.SRDX plant lines. False discovery rates (FDRs) were calculated by significance analysis of microarrays algorithm, and genes with fold change of at least 2 and a FDR-corrected P value lower than 0.05 were identified.
References
- Beerli RR, Dreier B, Barbas CF 3rd (2000) Positive and negative regulation of endogenous genes by designed transcription factors. Proc Natl Acad Sci USA 97:1495–1500
Article PubMed CAS Google Scholar - Birmingham A, Anderson EM, Reynolds A, Ilsley-Tyree D, Leake D, Fedorov Y, Baskerville S, Maksimova E, Robinson K, Karpilow J et al (2006) 3’ UTR seed matches, but not overall identity, are associated with RNAi off-targets. Nat Methods 3:199–204
Article PubMed CAS Google Scholar - Boch J, Bonas U (2010) Xanthomonas AvrBs3 family-type III effectors: discovery and function. Annu Rev Phytopathol 48:419–436
Article PubMed CAS Google Scholar - Boch J, Scholze H, Schornack S, Landgraf A, Hahn S, Kay S, Lahaye T, Nickstadt A, Bonas U (2009) Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326:1509–1512
Article PubMed CAS Google Scholar - Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, Baller JA, Somia NV, Bogdanove AJ, Voytas DF (2011) Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res 39:e82. doi: 10.1093/nar/gkr218
- Chinnusamy V, Stevenson B, Lee BH, Zhu JK (2002) Screening for gene regulation mutants by bioluminescence imaging. Sci STKE 36:pl10
Google Scholar - Christian M, Cermak T, Doyle EL, Schmidt C, Zhang F, Hummel A, Bogdanove AJ, Voytas DF (2010) Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 186:757–761
Article PubMed CAS Google Scholar - de Haan G, Chusacultanachai S, Mao C, Katzenellenbogen BS, Shapiro DJ (2000) Estrogen receptor-KRAB chimeras are potent ligand-dependent repressors of estrogen-regulated gene expression. J Biol Chem 275:13493–13501
Article PubMed Google Scholar - Fujimoto SY, Ohta M, Usui A, Shinshi H, Ohme-Takagi M (2000) Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 12:393–404
Article PubMed CAS Google Scholar - Guan X, Stege J, Kim M, Dahmani Z, Fan N, Heifetz P, Barbas CF 3rd, Briggs SP (2002) Heritable endogenous gene regulation in plants with designed polydactyl zinc finger transcription factors. Proc Natl Acad Sci USA 99:13296–13301
Article PubMed CAS Google Scholar - Heyl A, Ramireddy E, Brenner WG, Riefler M, Allemeersch J, Schmulling T (2008) The transcriptional repressor ARR1-SRDX suppresses pleiotropic cytokinin activities in Arabidopsis. Plant Physiol 147:1380–1395
Article PubMed CAS Google Scholar - Hiratsu K, Matsui K, Koyama T, Ohme-Takagi M (2003) Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J 34:733–739
Article PubMed CAS Google Scholar - Hua ZM, Yang X, Fromm ME (2006) Activation of the NaCl- and drought-induced RD29A and RD29B promoters by constitutively active Arabidopsis MAPKK or MAPK proteins. Plant Cell Environ 29:1761–1770
Article PubMed CAS Google Scholar - Ikeda M, Ohme-Takagi M (2009) A novel group of transcriptional repressors in Arabidopsis. Plant Cell Physiol 50:970–975
Article PubMed CAS Google Scholar - Ishitani M, Xiong L, Stevenson B, Zhu JK (1997) Genetic analysis of osmotic and cold stress signal transduction in Arabidopsis: interactions and convergence of abscisic acid-dependent and abscisic acid-independent pathways. Plant Cell 9:1935–1949
Article PubMed CAS Google Scholar - Kagale S, Rozwadowski K (2011) EAR motif-mediated transcriptional repression in plants: an underlying mechanism for epigenetic regulation of gene expression. Epigenetics 6(2):141–146
Google Scholar - Kasuga M, Miura S, Shinozaki K, Yamaguchi-Shinozaki K (2004) A combination of the Arabidopsis DREB1A gene and stress-inducible rd29A promoter improved drought- and low-temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol 45:346–350
Article PubMed CAS Google Scholar - Kay S, Boch J, Bonas U (2005) Characterization of AvrBs3-like effectors from a Brassicaceae pathogen reveals virulence and avirulence activities and a protein with a novel repeat architecture. Mol Plant Microbe Interact 18:838–848
Article PubMed CAS Google Scholar - Kay S, Hahn S, Marois E, Hause G, Bonas U (2007) A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science 318:648–651
Article PubMed CAS Google Scholar - Kazan K (2006) Negative regulation of defence and stress genes by EAR-motif-containing repressors. Trends Plant Sci 11:109–112
Article PubMed CAS Google Scholar - Krogan NT, Long JA (2009) Why so repressed? Turning off transcription during plant growth and development. Curr Opin Plant Biol 12:628–636
Article PubMed CAS Google Scholar - Li T, Huang S, Jiang WZ, Wright D, Spalding MH, Weeks DP, Yang B (2011a) TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res 39:359–372
Article PubMed Google Scholar - Li T, Huang S, Zhao X, Wright DA, Carpenter S, Spalding MH, Weeks DP, Yang B (2011b). Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes. Nucleic Acids Res 39:6315–6325
Google Scholar - Mahfouz MM (2010) RNA-directed DNA methylation: mechanisms and functions. Plant Signal Behav 5:806–816
Article PubMed CAS Google Scholar - Mahfouz MM, Li L (2011) TALE nucleases and next generation GM crops. GM Crop 2:99–103
Article Google Scholar - Mahfouz MM, Li L, Shamimuzzaman M, Wibowo A, Fang X, Zhu JK (2011) De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks. Proc Natl Acad Sci USA 108:2623–2628
Article PubMed CAS Google Scholar - Matsui K, Ohme-Takagi M (2010) Detection of protein–protein interactions in plants using the transrepressive activity of the EAR motif repression domain. Plant J 61:570–578
Article PubMed CAS Google Scholar - Matsui K, Umemura Y, Ohme-Takagi M (2008) AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J 55:954–967
Article PubMed CAS Google Scholar - Miller JC, Tan S, Qiao G, Barlow KA, Wang J, Xia DF, Meng X, Paschon DE, Leung E, Hinkley SJ et al (2011) A TALE nuclease architecture for efficient genome editing. Nat Biotechnol 29:143–148
Article PubMed CAS Google Scholar - Mito T, Seki M, Shinozaki K, Ohme-Takagi M, Matsui K (2010) Generation of chimeric repressors that confer salt tolerance in Arabidopsis and rice. Plant Biotechnol J 9:736–746
Google Scholar - Morbitzer R, Romer P, Boch J, Lahaye T (2010) Regulation of selected genome loci using de novo-engineered transcription activator-like effector (TALE)-type transcription factors. Proc Natl Acad Sci USA 107:21617–21622
Google Scholar - Morbitzer R, Elsaesser J, Hausner J, Lahaye T (2011) Assembly of custom TALE-type DNA binding domains by modular cloning. Nucleic Acids Res. doi: 10.1093/nar/gkr151
- Moscou MJ, Bogdanove AJ (2009) A simple cipher governs DNA recognition by TAL effectors. Science 326:1501
Article PubMed CAS Google Scholar - Mundy J, Yamaguchi-Shinozaki K, Chua NH (1990) Nuclear proteins bind conserved elements in the abscisic acid-responsive promoter of a rice rab gene. Proc Natl Acad Sci USA 87:1406–1410
Article PubMed CAS Google Scholar - Narusaka Y, Nakashima K, Shinwari ZK, Sakuma Y, Furihata T, Abe H, Narusaka M, Shinozaki K, Yamaguchi-Shinozaki K (2003) Interaction between two cis-acting elements, ABRE and DRE, in ABA-dependent expression of Arabidopsis rd29A gene in response to dehydration and high-salinity stresses. Plant J 34:137–148
Article PubMed CAS Google Scholar - Ohta M, Matsui K, Hiratsu K, Shinshi H, Ohme-Takagi M (2001) Repression domains of class II ERF transcriptional repressors share an essential motif for active repression. Plant Cell 13:1959–1968
Article PubMed CAS Google Scholar - Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, Adam L, Pineda O, Ratcliffe OJ, Samaha RR et al (2000) Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science 290:2105–2110
Article PubMed CAS Google Scholar - Romer P, Hahn S, Jordan T, Strauss T, Bonas U, Lahaye T (2007) Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science 318:645–648
Article PubMed Google Scholar - Szemenyei H, Hannon M, Long JA (2008) TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 319:1384–1386
Article PubMed CAS Google Scholar - Tsutsui T, Kato W, Asada Y, Sako K, Sato T, Sonoda Y, Kidokoro S, Yamaguchi-Shinozaki K, Tamaoki M, Arakawa K et al (2009) DEAR1, a transcriptional repressor of DREB protein that mediates plant defense and freezing stress responses in Arabidopsis. J Plant Res 122:633–643
Article PubMed CAS Google Scholar - Wang T, Iyer LM, Pancholy R, Shi X, Hall TC (2005) Assessment of penetrance and expressivity of RNAi-mediated silencing of the Arabidopsis phytoene desaturase gene. New Phytol 167:751–760
Article PubMed CAS Google Scholar - Weber E, Gruetzner R, Werner S, Engler C, Marillonnet S (2011) Assembly of Designer TAL Effectors by Golden Gate Cloning. PLoS One 6:e19722
Article PubMed CAS Google Scholar - Xiong L, Ishitani M, Lee H, Zhu JK (1999a) HOS5-a negative regulator of osmotic stress-induced gene expression in Arabidopsis thaliana. Plant J 19:569–578
Article PubMed CAS Google Scholar - Xiong L, Ishitani M, Zhu JK (1999b) Interaction of osmotic stress, temperature, and abscisic acid in the regulation of gene expression in Arabidopsis. Plant Physiol 119:205–212
Article PubMed CAS Google Scholar - Yaghmai R, Cutting GR (2002) Optimized regulation of gene expression using artificial transcription factors. Mol Ther 5:685–694
Article PubMed CAS Google Scholar - Yamaguchi-Shinozaki K, Shinozaki K (1994) A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 6:251–264
Article PubMed CAS Google Scholar - Zhang X, Henriques R, Lin SS, Niu QW, Chua NH (2006) Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat Protoc 1:641–646
Article PubMed CAS Google Scholar - Zhang F, Cong L, Lodato S, Kosuri S, Church GM, Arlotta P (2011) Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription. Nat Biotechnol 29:149–153
Article PubMed Google Scholar
Acknowledgments
We thank Thomas Lahaye for providing the Bs3::uidA binary construct. We also thank Jose M. Pardo and Ray Bressan for helpful discussions and members of the genome-engineering group at KAUST for their feedback and technical assistance.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
- Center for Plant Stress Genomics and Technology, King Abdullah University of Science and Technology, Thuwal, 23955-6900, Kingdom of Saudi Arabia
Magdy M. Mahfouz, Lixin Li, Marek Piatek & Xiaoyun Fang - BioScience Core Laboratory, King Abdullah University of Science and Technology, Thuwal, 23955-6900, Kingdom of Saudi Arabia
Hicham Mansour & Dhinoth K. Bangarusamy - Department of Horticulture and Landscape Architecture, Purdue University, West Lafayette, IN, 47907, USA
Jian-Kang Zhu
Authors
- Magdy M. Mahfouz
You can also search for this author inPubMed Google Scholar - Lixin Li
You can also search for this author inPubMed Google Scholar - Marek Piatek
You can also search for this author inPubMed Google Scholar - Xiaoyun Fang
You can also search for this author inPubMed Google Scholar - Hicham Mansour
You can also search for this author inPubMed Google Scholar - Dhinoth K. Bangarusamy
You can also search for this author inPubMed Google Scholar - Jian-Kang Zhu
You can also search for this author inPubMed Google Scholar
Corresponding authors
Correspondence toMagdy M. Mahfouz or Jian-Kang Zhu.
Electronic supplementary material
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Mahfouz, M.M., Li, L., Piatek, M. et al. Targeted transcriptional repression using a chimeric TALE-SRDX repressor protein.Plant Mol Biol 78, 311–321 (2012). https://doi.org/10.1007/s11103-011-9866-x
- Received: 27 October 2011
- Accepted: 27 November 2011
- Published: 14 December 2011
- Issue Date: February 2012
- DOI: https://doi.org/10.1007/s11103-011-9866-x