Differential Effects of σ1 Receptor Blockade on Self-Administration and Conditioned Reinstatement Motivated by Cocaine vs Natural Reward (original) (raw)
INTRODUCTION
The sigma1 (_σ_1) receptor is a 223-amino-acid protein localized in several organs, such as liver, heart, testis, gastrointestinal tract, and brain (Maurice et al, 2002). Immunohistochemical studies confirmed earlier in vitro binding assays indicating that _σ_1 receptors are present in numerous brain structures, with particularly high concentrations in specific areas within the limbic system as well as brainstem motor structures (for review, see Maurice et al, 2002). The highest levels of _σ_1 receptor immunostaining have been observed in the granular layer of the olfactory bulb, hypothalamic nuclei, hippocampus, and pyramidal layers of the hippocampus, caudate putamen, septum, nucleus accumbens, and amygdala (Alonso et al, 2000; but see also Maurice et al, 2002). At the subcellular level, the _σ_1 receptor is present in neurons, on the endoplasmic reticulum as well as mitochondrial, nuclear, and plasma membranes (Maurice et al, 2002; Su and Hayashi, 2003). Physiologically, _σ_1 receptor activation leads to cellular restructuring by translocating cholesterol and cytoskeletal proteins from the endoplasmic reticulum to the plasma membrane and nucleus, and regulates synaptic transmission by modulating intracellular Ca2+ mobilization (Hayashi and Su, 2001, 2003a, 2003b; Monnet et al, 2003). The endogenous ligand of _σ_1 receptors has not been fully identified yet. However, several lines of evidence suggest that neuroactive steroids such as dehydroepiandrosterone (DHEA) and progesterone act as potent endogenous _σ_1 receptor modulators (for review, see Monnet and Maurice, 2006). Behaviorally, _σ_1 receptors have been implicated in cognitive function, anxiety, depression, and regulation of stress responses (eg Maurice et al, 2001; Urani et al, 2001). Additionally, _σ_1 receptors have been found to modulate several neurobehavioral effects of cocaine, including the drug's subjective (Katz et al, 2003), psychomotor stimulant (Menkel et al, 1991; Ujike et al, 1996), rewarding (Romieu et al, 2000), and toxic (Matsumoto et al, 2001) actions. Of interest with regard to a _σ_1 receptor role in cocaine addiction is that pharmacological blockade of this receptor attenuates expression of cocaine-conditioned place preference (CPP) (Romieu et al, 2000, 2002), suggesting that _σ_1 receptors participate in mediating the conditioned incentive effects of cocaine-related environmental stimuli. It is well established that such stimuli can evoke craving or lead to relapse in abstinent individuals (eg O'Brien et al, 1998) and, in animals, consistently elicit reinstatement of extinguished cocaine-seeking (See, 2002; Shaham et al, 2003; Weiss, 2005).
To extend our understanding of the role of _σ_1 receptors in addiction-relevant conditioned effects of cocaine, the effects of a potent selective _σ_1 receptor antagonist, BD1047 (Matsumoto et al, 1995; McCracken et al, 1999b), were examined on reinstatement of cocaine-seeking induced by drug-related contextual stimuli. To establish whether BD1047 preferentially modifies drug-directed behavior or exerts general suppressant effects on motivated behavior, BD1047 effects were tested also on responding induced by stimuli conditioned to a potent conventional reinforcer, sweetened condensed milk (SCM). Because _σ_1 receptors have been implicated not only in the expression but also acquisition (Romieu et al, 2000, 2002) of cocaine CPP (ie a process linked to the acute reinforcing actions of cocaine), tests of BD1047 effects on self-administration of cocaine and SCM were conducted as well.
MATERIALS AND METHODS
Subjects
Eighty-seven male Wistar rats (Charles River, Wilmington, MA; 200–250 g upon arrival) were housed 2–3/cage in a temperature- and humidity-controlled vivarium on a reverse 12:12 h light/dark cycle with ad libitum access to food and water. All procedures were conducted in strict adherence to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee of The Scripps Research Institute.
Drugs
Cocaine hydrochloride (National Institute on Drug Abuse, Bethesda, MD) was dissolved in sterile physiological saline. Cocaine or saline vehicle was intravenously (i.v.) infused at a volume of 0.1 ml over 4 s. _N_-[2-(3,4-dichlorophenyl)ethyl]-_N_-methyl-2-(dimethylamino)ethylamine (BD1047), obtained from Dr WD Bowen (Brown University, Providence, RI), was dissolved in distilled water and intraperioneally (i.p.) administered in a volume of 1 ml/kg.
Effects of BD1047 on Conditioned Reinstatement: Procedures
Behavioral training and testing were conducted according to previously described procedures (Baptista et al, 2004; Caine et al, 1993; Weiss et al, 2000). Briefly, operant responding maintained by food or SCM was initially established using a continuous reinforcement schedule in daily 60- and 40-min sessions, respectively. Rats designated for cocaine self-administration were surgically prepared with jugular catheters (Caine et al, 1993; Weiss et al, 2000) and given 7 days of recovery before commencing self-administration training. Self-administration of cocaine (0.25 mg/0.1 ml; i.v.) or SCM (2 : 1 v/v in distilled water; 0.1 ml delivered into a 0.2 ml receptacle) began on a fixed ratio 1 (FR1) schedule of reinforcement in daily 120-min cocaine or 40-min SCM sessions, 5 days/week. Responses at the right, active lever were reinforced and followed by a 20-s time-out (TO) period signaled by illumination of a cue light above the lever. During this time, the lever remained inactive. Responses at the left, inactive lever had no scheduled consequences.
Following 2 weeks of training, responses at the active lever were differentially reinforced in the presence of distinct discriminative stimuli (SD). A constant 70 dB white noise (S+) signaled availability of the reinforcer whereas illumination of a 2.8 W house light (S−), located at the top of the chamber's front panel, signaled the absence of the reinforcer (ie, saline solution instead of cocaine or no consequence instead of SCM; see Baptista et al, 2004). Three daily sessions (lasting 1 h for the cocaine group, and restricted to 20 min for the SCM group to avoid satiety effects by excessive ingestion of SCM), separated by 30-min intervals, were conducted, with two ‘reward’ sessions and one ‘non-reward’ session sequenced in random order. Sessions were initiated by presentation of the respective SD and extension of the levers. The SD remained present until termination of the session by retraction of the levers. After eight training days (ie, a total of 16 ‘reward’ and 8 ‘non-reward’ sessions), both the cocaine and SCM groups were placed on extinction conditions in daily 1-h sessions during which the reinforcers and SD were withheld until a criterion of ⩽4 responses/session for 3 consecutive days was reached. One day after each animal reached the extinction criterion, reinstatement tests began. These 1-h tests were conducted under extinction conditions, but with reintroduction of the SD as during the conditioning phase. Rats were tested first in the presence of the S− to verify the behavioral selectivity of the SD. Two days later, tests of BD1047 (0, 1, 3, 10, 20, or 30 mg/kg, i.p.) on S+-induced reinstatement began. BD1047 was administered 10 min before the onset of sessions. Each animal was tested with only one dose of BD1047 according to a between-subjects design.
Effect of BD1047 on Self-Administration: Procedures
After acquisition of cocaine- or SCM-reinforced responding on an FR1 schedule as above, but without being subjected to conditioning procedures, rats received further daily 120-min access to cocaine on an FR 5 schedule or 30-min access to SCM on an FR1 schedule until stable intake developed (±10%). Effects of BD1047 (0, 1, 3, 10, 20, and 30 mg/kg, i.p.) on cocaine or SCM self-administration were then determined. Each rat was tested once with each BD1047 dose on separate days according to a Latin square design (_n_=6 animals/group). Each drug test was preceded by five baseline cocaine or SCM self-administration sessions.
Statistics
Differences in responding at the active lever between the respective reward and non-reward conditions during the training phase were analyzed by paired _t_-tests. Differences in the number of responses during the extinction and reinstatement phases, including the effects of BD1047 on reinstatement responses, were analyzed separately for the cocaine and SCM groups by one-way ANOVA. Cumulative responses were analyzed by mixed-factorial ANOVA, followed by simple effects analysis. Effects of BD1047 on responses reinforced directly by cocaine or SCM were analyzed separately by one-way within-subjects ANOVA. Significant omnibus tests were followed by Fisher's PLSD post hoc tests.
RESULTS
Effect of BD1047 on Conditioned Reinstatement
Cocaine
All rats (_n_=33) acquired cocaine-reinforced responding, maintained stable cocaine self-administration during the conditioning phase, and ceased responding during non-reward (saline) sessions (Figure 1a, left panel). Following initiation of the extinction contingency, rats required, on average, 9.6±1.3 (mean±SEM) sessions to reach the criterion. During subsequent reinstatement tests, the cocaine S+ (in BD1047 vehicle-treated rats) but not the saline-associated S− elicited strong recovery of responding (p<0.001, Fisher PLSD tests after ANOVA: F2,8=31.2; p<0.001; Figure 1a). BD1047 dose-dependently reduced the response reinstatement induced by the S+ (Figure 1a, right panel) with significant effects at the 20 (p<0.05) and 30 mg/kg (p<0.01) doses (Fisher PLSD tests after ANOVA: F5,27=3.2; p<0.05). Further examination of the BD1047 effects revealed that this agent modified the cumulative response profile associated with S+-induced reinstatement (Figure 1a, inset) as reflected by a main effect for BD1047 doses (F5,27=4.2; p<0.01) and a dose × time (10 min intervals) interaction (F25,135=1.9; p<0.01). Moreover, this analysis indicated that only the 20 mg/kg and 30 mg/kg BD1047 doses decreased responding at all time points compared to vehicle-treated rats (simple effects, p<0.05; Figure 1a, inset). Inactive lever responses remained low (⩽4 responses) throughout the experiment and unaltered by BD1047.
Figure 1
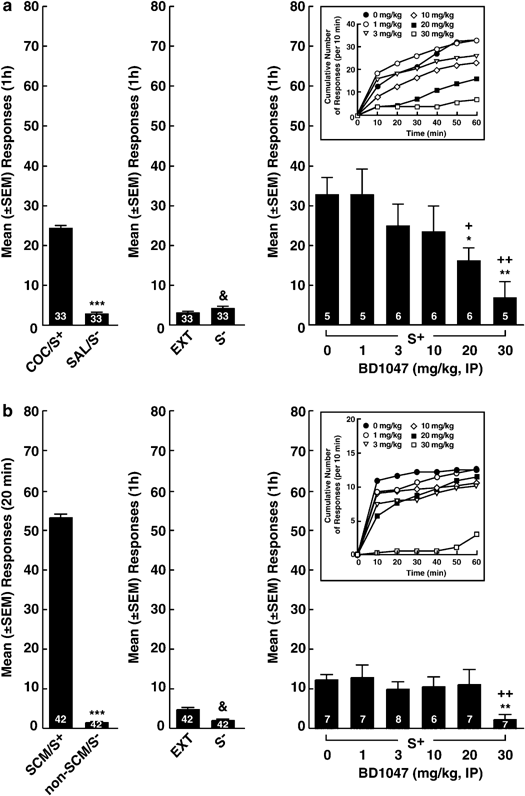
Effects of BD1047 on reinstatement induced by discriminative stimuli associated with (a) cocaine and (b) SCM. (a) Left panels: active lever responses during conditioning sessions in the presence of stimuli paired with cocaine (upper panel: COC/S+) or SCM (lower panel: SCM/S+) vs non-availability of these reinforcers (upper panel: SAL/S−; lower panel: non-SCM/S−). Center panels: Extinction (EXT) responses at criterion and responses during an initial reinstatement test in the presence of the stimulus paired with reward non-availability (S−). Right panels: reinstatement responses in the presence of the stimuli previously associated with reward (cocaine: upper panel; SCM: lower panel) availability (S+) in vehicle-treated rats (0), and modification of conditioned reinstatement across doses of BD1047. Insets: cumulative number of responses throughout the 60-min reinstatement periods (error bars omitted for clarity). Upper panels: ***paired _t_-test _t_32=14.3; p<0.001 vs COC/S+; &p<0.001 vs S+; *p<0.05 and **p<0.01 vs vehicle; +p<0.05 and ++p<0.01 vs BD1047 1 mg/kg. Lower panels: ***paired _t_-test _t_42=91.1; p<0.001 vs SCM/S+; &p<0.001 vs S+; **p<0.01 vs vehicle; ++p<0.01 vs BD1047 1 mg/kg. Numbers inside bars represent sample sizes.
Sweetened condensed milk
As in the case of cocaine, all rats (_n_=42) acquired robust SCM-reinforced responding, maintained stable responding during the conditioning phase, and ceased responding during non-reward sessions (Figure 1b, left panel). Rats required on average 10.3±0.7 (mean±SEM) sessions to reach the extinction criterion. Presentation of the SCM S+ (in vehicle-injected rats), but not the S−, produced reliable reinstatement (p<0.001, Fisher PLSD tests after ANOVA: F2,12=53.3; p<0.001; Figure 1b, right panel). Only the highest dose of BD1047 attenuated the S+-induced response reinstatement (p<0.01, Fisher PLSD tests after ANOVA: F5,36=2.6; p<0.05; Figure 1b). Examination of BD1047's effects on cumulative responses confirmed that the _σ_1 antagonist modified the shape of the cumulative response profile (main effect of BD1047 dose: F5,36=3.5; p<0.05), but only at the highest dose (Figure 1b, inset). The failure to obtain a significant dose × time interaction reflects the suppressant effects on reinstatement of only a single (ie the 30 mg/kg) dose that persisted across the entire session (Figure 1b, inset). Responses at the inactive lever remained low (4 responses) throughout training and testing and were not modified by BD1047.
Effect of BD1047 on Cocaine and SCM-Reinforced Behavior
All rats acquired stable cocaine (_n_=6) or SCM self-administration (_n_=6) after 14 days of training. BD1047 did not alter either cocaine (F5,25=1.9; _p_>0.05; Figure 2a) or SCM-reinforced responding (F5,25=1.7; _p_>0.05; Figure 2b). Responses at the inactive lever remained low (⩽2 responses) throughout testing, and were not modified by BD1047.
Figure 2
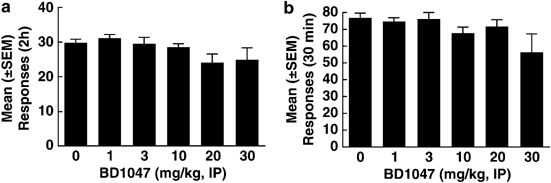
Effects of BD1047 on self-administration of cocaine or SCM. Cocaine (a) and SCM- (b) reinforced responses after vehicle vs BD1047 administration.
DISCUSSION
BD1047 dose-dependently reduced reinstatement induced by a cocaine-related contextual stimulus, whereas responding elicited by a stimulus conditioned to a palatable conventional reinforcer, SCM, was attenuated at the highest BD1047 dose only. The _σ_1 antagonist did not modify self-administration of cocaine or SCM. This observation suggests that pharmacological blockade of _σ_1 receptors does not modify the acute reinforcing effects of cocaine or palatable natural reward, but selectively attenuates the incentive-motivational effects of reward-paired contextual stimuli with a preferential action on stimuli conditioned to cocaine vs natural reward.
The preferential interference by BD1047 with cocaine S+-induced reinstatement resembles earlier observations where a Group II metabotropic glutamate receptor agonist (LY379268) selectively attenuated reinstatement induced by stimuli conditioned to cocaine vs conventional reinforcers including food and SCM (Baptista et al, 2004). This effect of BD1047 is unlikely to be the result of differences in response-reinstating efficacy between the cocaine and SCM S+ per se. While SCM conditioning sessions were restricted to 20 min to avoid satiety and, thus, shorter than cocaine conditioning sessions, the number of responses per session was considerably higher in the SCM than in the cocaine group such that these animals experienced a greater number of reinforcer presentations in the presence of the SCM S+ compared to the cocaine S+ condition. Nonetheless, as in previous reports where stimuli conditioned to conventional reinforcers were less effective in eliciting reward-seeking than cocaine-associated stimuli (Baptista et al, 2004; Grimm et al, 2002), the SCM S+ produced weaker reinstatement than the cocaine S+. Moreover, the SCM S+ induced and maintained responding only during the first 10–20 min of the reinstatement test, whereas the cocaine S+ produced strong and sustained responding throughout the 60 min session.
BD1047 produced dose-dependent effects on reinstatement only in the case of the cocaine S+. However, at the highest dose, reinstatement elicited by the SCM S+ was significantly suppressed as well. Further scrutiny of the time course of responding during the reinstatement tests (Figure 1a and b, insets) shows that the highest doses of BD1047 induced a delayed onset of responding. Specifically, the effects of the 20 mg/kg dose appeared to dissipate 30 min into the test (Figure 1a), whereas the effects of the 30 mg/kg dose did not show signs of dissipation until 50–60 min after the beginning of the test (see insets of Figure 1a and b). The pharmacokinetic profile of BD1047 has not yet been precisely described. However, these observations in conjunction with earlier data (Matsumoto et al, 1995; Romieu et al, 2004; Urani et al, 2001) strongly suggest that the behavioral effects of BD1047 last for at least 30 min following administration and that BD1047's duration of action may vary as a function of dose. The reason for the failure of BD1047 to alter SCM S+-induced behavior at low doses, paired with the full suppression of this behavior only at the highest dose is presently unclear. This effect cannot be attributed to motor impairment or sedation, because BD1047 did not interfere, even at the highest dose, with the high rate of responding in rats self-administering SCM (Figure 2b), confirming previous reports that _σ_1 receptor antagonists are devoid of general suppressant effects on behavior (McCracken et al, 1999a; Romieu et al, 2006). In particular, while BD1047 reduces the acute locomotor stimulant effects of cocaine as well as cocaine-induced convulsions and lethality, the _σ_1 receptor antagonist does not interfere with spontaneous locomotor activity (McCracken et al, 1999a).
As BD1047 reliably attenuated conditioned cocaine-seeking, one might expect that this drug would exert at least similar, if not greater, effects on behavior that is less robust (ie conditioned SCM-seeking). One interpretation of this finding is that _σ_1 receptors participate in mediating behavior motivated by drug-related but not non-drug-related stimuli and therefore, possibly, point toward the existence of separate neural substrates for drug-related vs conventional learning. It is also possible that _σ_1 receptors participate in regulating conditioning processes independent of the nature of the unconditioned stimulus (ie cocaine vs SCM), but that the effects of non-drug cues are more resistant to disruption than the effects of drug cues. A more likely interpretation for the differential effects of BD1047 on cocaine S+ vs SCM S+-induced reinstatement is related to upregulation of _σ_1 receptor function after cocaine treatment. Acute cocaine administration has been shown to upregulate _σ_1 receptor gene expression and protein levels in whole brain, striatum, and cortex (Liu et al, 2005). Moreover, 4 days of intermittent cocaine exposure during place conditioning increased in vivo binding levels of the _σ_1 receptor agonist [3H](+)-SKF-10,047 in the olfactory bulb, hippocampus, hypothalamus, cortex, and striatum—a change that was sustained during extinction of CPP—indicative of persistent functional _σ_1 receptor upregulation (Romieu et al, 2004). It is possible then that over the course of cocaine self-administration and cue conditioning, followed by a prolonged extinction period, sustained upregulation of _σ_1 receptor function may have occurred, an effect that would not be expected to develop in the cocaine-naïve SCM group. Consequently, increases in _σ_1 receptor expression or affinity may have amplified the effects of BD1047 in the cocaine-exposed but not drug-naïve SCM group, providing a possible explanation for the differential efficacy of BD1047 in reversing behavior induced by the cocaine vs SCM S+. Several endogenous systems have been shown to interact with the _σ_1 receptor, including peptides of the neuropeptide Y and calcitonin gene-related peptide families and neuroactive steroids that have attracted much interest recently as putative endogenous ligands for the _σ_1 receptor (for reviews, see Maurice, 2004; Monnet and Maurice, 2006). Specifically, it has been shown that pregnenolone and DHEA act as _σ_1 receptor agonists, progesterone being the most potent endogenous _σ_1 receptor antagonist known to date (Maurice, 2004; Maurice et al, 2002). These neuroactive steroids have been shown to modulate cocaine-induced locomotor stimulation or CPP through a direct interaction with _σ_1 receptors (Romieu et al, 2003), and in vivo assay also showed that circulating levels of cortisol and DHEA sulfate were increased following days of abstinence in cocaine addicts (Buydens-Branchey and Branchey, 2004; Buydens-Branchey et al, 2002). Finally, studies in rats have confirmed that DHEA attenuated cocaine self-administration and cocaine-seeking, an effect possibly resulting from an interaction with the _σ_1 receptor (Doron et al, 2006; Maayan et al, 2006). On the basis of these findings, it would seem justified to speculate that cocaine during self-administration training and conditioning produced persistent neuroendocrine perturbations, which could in turn have resulted in upregulation of _σ_1 receptors increasing susceptibility to relapse (ie, conditioned reinstatement) in the cocaine group.
BD1047 did not modify the primary reinforcing effects of cocaine and SCM at a dose range that progressively reduced the conditioned effects of the cocaine S+. Sigma1 receptors are highly expressed in the hippocampus (Alonso et al, 2000), a brain site with an established role in the occasion-setting action of contextual stimuli (eg Holland and Bouton, 1999). Indeed, transient inactivation of the dorsal hippocampus was shown to specifically block contextual reinstatement of cocaine-seeking, but not reinstatement induced by cocaine or discrete cocaine-paired cues (Fuchs et al, 2005). Therefore, one may speculate that, in the present study, the _σ_1 receptor antagonist interfered with the processing of reward-related contextual information at the hippocampal level, reducing its motivating impact, an effect that would not interfere with the direct reinforcing actions of drug or natural reward. It is important to note, however, that BD1047 has been reported to also block cocaine priming-induced reactivation of cocaine CPP (Romieu et al, 2004)—an effect thought to be mediated by the nucleus accumbens shell and the ventral tegmental area (see Kalivas and McFarland, 2003 for review)—suggesting that _σ_1 receptors in the nucleus accumbens and the ventral tegmental area may play a role in reinstatement induced by cocaine priming manipulations (Alonso et al, 2000; Maurice et al, 2002; McFarland et al, 2003). However, a more comprehensive understanding of the pharmacological profile of BD1047 relevant for reinstatement and relapse, including its effects on priming-induced reinstatement as well as the assessment of specific links between effects of _σ_1 manipulations in anatomically distinct brain regions on cocaine-seeking associated with distinct risk factors (ie contextual vs discrete cocaine cues vs cocaine priming vs stress exposure) will remain for further research.
The lack of BD1047 effect on cocaine self-administration observed here is in apparent contradiction to earlier findings showing that BD1047 blocks the acquisition of cocaine CPP, suggesting that the drug can antagonize the acute reinforcing actions of cocaine (Romieu et al, 2000, 2002). Several explanations can be offered for this discrepancy. First, acquisition of cocaine CPP was obtained with involuntary cocaine administration rather than self-administration. It is well established that the behavioral and neurochemical effects of voluntary vs involuntary cocaine administration differ (eg Hemby et al, 1997; Jacobs et al, 2003; Wise, 2000). Moreover, the duration and amount of cocaine exposure in the self-administering rats of the present study was considerably greater compared to that in the CPP studies. Therefore, both the reinforcing quality and relevant neuroadaptive changes are likely to differ in rats subjected to involuntary vs self-administration of cocaine. Consistent with this hypothesis is recent evidence on differential regulation of the _σ_1 receptor as a function of the mode of administration of methamphetamine (active vs passive). Rats self-administering methamphetamine showed increased expression of _σ_1 receptor mRNA in the hippocampus and decreased expression in the frontal cortex compared to yoked-methamphetamine and yoked-saline controls (Stefanski et al, 2004). Second, rats in the present study self-administered cocaine for 2 h after receiving BD1047. Therefore, the effects of BD1047 may have been surmountable by continued response-contingent administration of cocaine in the present case, but not in the CPP studies in which rats received a single i.p. cocaine dose before conditioning sessions. A possible alternative explanation for these discrepant findings may be that self-administration and CPP procedures also differ with respect to the significance of contextual learning in the acquisition of cocaine-reinforced behavior. The contextual associative learning component relevant for CPP through which environmental stimuli in the drug-paired environment eventually establish secondary reinforcing properties is less critical in the establishment of drug-reinforced behavior in self-administration studies. In fact, self-administration depends on intact operant learning abilities of the organism (ie animals learn the association between a response and an outcome rather than between a previously neutral set of environmental stimuli and drug-induced internal state). According to this possibility, BD1047 may be effective in disrupting the acquisition of associations between previously neutral cues now paired with cocaine, but does not disrupt the maintenance of response–outcome behavior. A comparative study testing the effects of chronic BD1047 administration during the induction of CPP vs acquisition of cocaine self-administration will be needed to pursue this hypothesis.
What may constrain interpretation of the findings in terms of a specific role of _σ_1 receptors in conditioned cocaine reinstatement is that only a single _σ_1 antagonist was tested. However, few _σ_1-selective ligands are presently available, and BD1047, which has nanomolar affinity for the _σ_1 receptor, is the most selective and potent _σ_1 receptor antagonist described to date ([Daniels et al, 2006](/articles/1301323#ref-CR6 "Daniels A, Ayala E, Chen W, Coop A, Matsumoto RR (2006). N-[2-(m-methoxyphenyl)ethyl]-N-ethyl-2-(1-pyrrolidinyl)ethylamine (UMB 116) is a novel antagonist for cocaine-induced effects. Eur J Pharmacol, print copy in press (originally published online April 5, 2006, at www.sciencedirect.com
)."); [Matsumoto et al, 1995](/articles/1301323#ref-CR20 "Matsumoto RR, Bowen WD, Tom MA, Vo VN, Truong DD, De Costa BR (1995). Characterization of two novel sigma receptor ligands: antidystonic effects in rats suggest sigma receptor antagonism. Eur J Pharmacol 280: 301–310."); [McCracken et al, 1999a](/articles/1301323#ref-CR25 "McCracken KA, Bowen WD, de Costa BR, Matsumoto RR (1999a). Two novel sigma receptor ligands, BD1047 and LR172, attenuate cocaine- induced toxicity and locomotor activity. Eur J Pharmacol 370: 225–232."), [1999b](/articles/1301323#ref-CR26 "McCracken KA, Bowen WD, Matsumoto RR (1999b). Novel sigma receptor ligands attenuate the locomotor stimulatory effects of cocaine. Eur J Pharmacol 365: 35–38.")). Moreover, although BD1047 also has moderate affinity for _σ_2 receptors and _β_\-adrenergic receptors, it shows substantial selectivity for _σ_1 receptors (≈50 times over _σ_2 receptors, and ≈150 times over _β_\-adrenergic receptors), providing confidence that, at the dose range used, the behavioral effects obtained here are attributable to an action at _σ_1 receptors ([Matsumoto et al, 1995](/articles/1301323#ref-CR20 "Matsumoto RR, Bowen WD, Tom MA, Vo VN, Truong DD, De Costa BR (1995). Characterization of two novel sigma receptor ligands: antidystonic effects in rats suggest sigma receptor antagonism. Eur J Pharmacol 280: 301–310.")). In addition, evidence exists that reduction of _σ_1 receptor expression following antisense treatment prevents the acquisition of cocaine CPP, a finding that directly implicates _σ_1 receptors in contextual cocaine conditioning ([Romieu et al, 2000](/articles/1301323#ref-CR34 "Romieu P, Martin-Fardon R, Maurice T (2000). Involvement of the sigma1 receptor in the cocaine-induced conditioned place preference. Neuroreport 11: 2885–2888.")). Moreover, BD1047 produced effects on CPP highly similar to those of _σ_1 antisense treatment ([Romieu et al, 2000](/articles/1301323#ref-CR34 "Romieu P, Martin-Fardon R, Maurice T (2000). Involvement of the sigma1 receptor in the cocaine-induced conditioned place preference. Neuroreport 11: 2885–2888.")). Together, these observations provide strong support for the interpretation that the present findings resulted from an action of BD1047 at _σ_1 receptors.In summary, the selective _σ_1 receptor antagonist, BD1047, attenuated conditioned reinstatement by reward-paired contextual stimuli with a preferential action on stimuli conditioned to cocaine vs natural reward. BD1047 had little effect on consummatory behavior maintained by drug or natural reward. The presumably selective _σ_1 receptor antagonist action of BD1047, therefore, appears to attenuate the motivating effects of cocaine cues without interfering with normal motivational function. Thus, the results identify BD1047 as a promising agent for further scrutiny with regard to therapeutic potential and, by inference, that _σ_1 receptors may represent a promising novel target for the prevention of cocaine craving and relapse.
References
- Alonso G, Phan V, Guillemain I, Saunier M, Legrand A, Anoal M et al (2000). Immunocytochemical localization of the sigma(1) receptor in the adult rat central nervous system. Neuroscience 97: 155–170.
CAS Google Scholar - Baptista MA, Martin-Fardon R, Weiss F (2004). Preferential effects of the metabotropic glutamate 2/3 receptor agonist LY379268 on conditioned reinstatement versus primary reinforcement: comparison between cocaine and a potent conventional reinforcer. J Neurosci 24: 4723–4727.
Article CAS Google Scholar - Buydens-Branchey L, Branchey M (2004). Cocaine addicts with conduct disorder are typified by decreased cortisol responsivity and high plasma levels of DHEA-S. Neuropsychobiology 50: 161–166.
Article CAS Google Scholar - Buydens-Branchey L, Branchey M, Hudson J, Dorota Majewska M (2002). Perturbations of plasma cortisol and DHEA-S following discontinuation of cocaine use in cocaine addicts. Psychoneuroendocrinology 27: 83–97.
Article CAS Google Scholar - Caine SB, Lintz R, Koob GF (1993). Intravenous drug self-administration techniques in animals. In: Sahgal A (ed). Behavioral Neuroscience: A Practical Approach, Vol. 2. University Press: Oxford. pp 117–143.
Google Scholar - Daniels A, Ayala E, Chen W, Coop A, Matsumoto RR (2006). _N_-[2-(_m_-methoxyphenyl)ethyl]-_N_-ethyl-2-(1-pyrrolidinyl)ethylamine (UMB 116) is a novel antagonist for cocaine-induced effects. Eur J Pharmacol, print copy in press (originally published online April 5, 2006, at www.sciencedirect.com ).
- Doron R, Fridman L, Gispan-Herman I, Maayan R, Weizman A, Yadid G (2006). DHEA, a neurosteroid, decreases cocaine self-administration and reinstatement of cocaine-seeking behavior in rats. Neuropsychopharmacology 31: 2231–2236.
Article CAS Google Scholar - Fuchs RA, Evans KA, Ledford CC, Parker MP, Case JM, Mehta RH et al (2005). The role of the dorsomedial prefrontal cortex, basolateral amygdala, and dorsal hippocampus in contextual reinstatement of cocaine seeking in rats. Neuropsychopharmacology 30: 296–309.
Article CAS Google Scholar - Grimm JW, Shaham Y, Hope BT (2002). Effect of cocaine and sucrose withdrawal period on extinction behavior, cue-induced reinstatement, and protein levels of the dopamine transporter and tyrosine hydroxylase in limbic and cortical areas in rats. Behav Pharmacol 13: 379–388.
Article CAS Google Scholar - Hayashi T, Su TP (2001). Regulating ankyrin dynamics: roles of sigma-1 receptors. Proc Natl Acad Sci USA 98: 491–496.
Article CAS Google Scholar - Hayashi T, Su TP (2003a). Intracellular dynamics of sigma-1 receptors (sigma(1) binding sites) in NG108-15 cells. J Pharmacol Exp Ther 306: 726–733.
Article CAS Google Scholar - Hayashi T, Su TP (2003b). Sigma-1 receptors (sigma(1) binding sites) form raft-like microdomains and target lipid droplets on the endoplasmic reticulum: roles in endoplasmic reticulum lipid compartmentalization and export. J Pharmacol Exp Ther 306: 718–725.
Article CAS Google Scholar - Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997). Differences in extracellular dopamine concentrations in the nucleus accumbens during response-dependent and response-independent cocaine administration in the rat. Psychopharmacology 133: 7–16.
Article CAS Google Scholar - Holland PC, Bouton ME (1999). Hippocampus and context in classical conditioning. Curr Opin Neurobiol 9: 195–202.
Article CAS Google Scholar - Jacobs EH, Smit AB, de Vries TJ, Schoffelmeer AN (2003). Neuroadaptive effects of active versus passive drug administration in addiction research. Trends Pharmacol Sci 24: 566–573.
Article CAS Google Scholar - Kalivas PW, McFarland K (2003). Brain circuitry and the reinstatement of cocaine-seeking behavior. Psychopharmacology (Berl) 168: 44–56.
Article CAS Google Scholar - Katz JL, Libby TA, Kopajtic T, Husbands SM, Newman AH (2003). Behavioral effects of rimcazole analogues alone and in combination with cocaine. Eur J Pharmacol 468: 109–119.
Article CAS Google Scholar - Liu Y, Chen GD, Lerner MR, Brackett DJ, Matsumoto RR (2005). Cocaine up-regulates Fra-2 and sigma-1 receptor gene and protein expression in brain regions involved in addiction and reward. J Pharmacol Exp Ther 314: 770–779.
Article CAS Google Scholar - Maayan R, Lotan S, Doron R, Shabat-Simon M, Gispan-Herman I, Weizman A et al (2006). Dehydroepiandrosterone (DHEA) attenuates cocaine-seeking behavior in the self-administration model in rats. Eur Neuropsychopharmacol 16: 329–339.
Article CAS Google Scholar - Matsumoto RR, Bowen WD, Tom MA, Vo VN, Truong DD, De Costa BR (1995). Characterization of two novel sigma receptor ligands: antidystonic effects in rats suggest sigma receptor antagonism. Eur J Pharmacol 280: 301–310.
Article CAS Google Scholar - Matsumoto RR, McCracken KA, Pouw B, Miller J, Bowen WD, Williams W et al (2001). N-alkyl substituted analogs of the sigma receptor ligand BD1008 and traditional sigma receptor ligands affect cocaine-induced convulsions and lethality in mice. Eur J Pharmacol 411: 261–273.
Article CAS Google Scholar - Maurice T (2004). Neurosteroids and sigma1 receptors, biochemical and behavioral relevance. Pharmacopsychiatry 37 (Suppl 3): S171–S182.
Article CAS Google Scholar - Maurice T, Martin-Fardon R, Romieu P, Matsumoto RR (2002). Sigma(1) (sigma(1)) receptor antagonists represent a new strategy against cocaine addiction and toxicity. Neurosci Biobehav Rev 26: 499–527.
Article CAS Google Scholar - Maurice T, Phan VL, Privat A (2001). The anti-amnesic effects of sigma1 (sigma1) receptor agonists confirmed by in vivo antisense strategy in the mouse. Brain Res 898: 113–121.
Article CAS Google Scholar - McCracken KA, Bowen WD, de Costa BR, Matsumoto RR (1999a). Two novel sigma receptor ligands, BD1047 and LR172, attenuate cocaine- induced toxicity and locomotor activity. Eur J Pharmacol 370: 225–232.
Article CAS Google Scholar - McCracken KA, Bowen WD, Matsumoto RR (1999b). Novel sigma receptor ligands attenuate the locomotor stimulatory effects of cocaine. Eur J Pharmacol 365: 35–38.
Article CAS Google Scholar - McFarland K, Lapish CC, Kalivas PW (2003). Prefrontal glutamate release into the core of the nucleus accumbens mediates cocaine-induced reinstatement of drug-seeking behavior. J Neurosci 23: 3531–3537.
Article CAS Google Scholar - Menkel M, Terry P, Pontecorvo M, Katz JL, Witkin JM (1991). Selective sigma ligands block stimulant effects of cocaine. Eur J Pharmacol 201: 251–252.
Article CAS Google Scholar - Monnet FP, Maurice T (2006). The σ1 protein as a target for the non-genomic effects of neuro(active)steroids: molecular, physiological, and behavioral aspects. J Pharmacol Sci 100: 93–118.
Article CAS Google Scholar - Monnet FP, Morin-Surun MP, Leger J, Combettes L (2003). Protein kinase C-dependent potentiation of intracellular calcium influx by sigma1 receptor agonists in rat hippocampal neurons. J Pharmacol Exp Ther 307: 705–712.
Article CAS Google Scholar - O'Brien CP, Childress AR, Ehrman R, Robbins SJ (1998). Conditioning factors in drug abuse: can they explain compulsion? J Psychopharmacol 12: 15–22.
Article CAS Google Scholar - Romieu P, Lucas M, Maurice T (2006). Sigma(1) receptor ligands and related neuroactive steroids interfere with the cocaine-induced state of memory. Neuropsychopharmacology 31: 1431–1443.
Article CAS Google Scholar - Romieu P, Martin-Fardon R, Bowen WD, Maurice T (2003). Sigma 1 receptor-related neuroactive steroids modulate cocaine-induced reward. J Neurosci 23: 3572–3576.
Article CAS Google Scholar - Romieu P, Martin-Fardon R, Maurice T (2000). Involvement of the sigma1 receptor in the cocaine-induced conditioned place preference. Neuroreport 11: 2885–2888.
Article CAS Google Scholar - Romieu P, Meunier J, Garcia D, Zozime N, Martin-Fardon R, Bowen WD et al (2004). The sigma1 (sigma1) receptor activation is a key step for the reactivation of cocaine conditioned place preference by drug priming. Psychopharmacology (Berl) 175: 154–162.
Article CAS Google Scholar - Romieu P, Phan VL, Martin-Fardon R, Maurice T (2002). Involvement of the sigma(1) receptor in cocaine-induced conditioned place preference: possible dependence on dopamine uptake blockade. Neuropsychopharmacology 26: 444–455.
Article CAS Google Scholar - See RE (2002). Neural substrates of conditioned-cued relapse to drug-seeking behavior. Pharmacol Biochem Behav 71: 517–529.
Article CAS Google Scholar - Shaham Y, Shalev U, Lu L, De Wit H, Stewart J (2003). The reinstatement model of drug relapse: history, methodology and major findings. Psychopharmacology (Berl) 168: 3–20.
Article CAS Google Scholar - Stefanski R, Justinova Z, Hayashi T, Takebayashi M, Goldberg SR, Su TP (2004). Sigma1 receptor upregulation after chronic methamphetamine self-administration in rats: a study with yoked controls. Psychopharmacology (Berl) 175: 68–75.
Article CAS Google Scholar - Su TP, Hayashi T (2003). Understanding the molecular mechanism of sigma-1 receptors: towards a hypothesis that sigma-1 receptors are intracellular amplifiers for signal transduction. Curr Med Chem 10: 2073–2080.
Article CAS Google Scholar - Ujike H, Kuroda S, Otsuki S (1996). sigma Receptor antagonists block the development of sensitization to cocaine. Eur J Pharmacol 296: 123–128.
Article CAS Google Scholar - Urani A, Roman FJ, Phan VL, Su TP, Maurice T (2001). The antidepressant-like effect induced by sigma(1)-receptor agonists and neuroactive steroids in mice submitted to the forced swimming test. J Pharmacol Exp Ther 298: 1269–1279.
CAS Google Scholar - Weiss F (2005). Neurobiology of craving, conditioned reward and relapse. Curr Opin Pharmacol 5: 9–19.
Article CAS Google Scholar - Weiss F, Maldonado-Vlaar CS, Parsons LH, Kerr TM, Smith DL, Ben-Shahar O (2000). Control of cocaine-seeking behavior by drug-associated stimuli in rats: effects on recovery of extinguished operant responding and extracellular dopamine levels in amygdala and nucleus accumbens. Proc Natl Acad Sci USA 97: 4321–4326.
Article CAS Google Scholar - Wise RA (2000). Addiction becomes a brain disease. Neuron 26: 27–33.
Article CAS Google Scholar
Acknowledgements
This is publication number 17316-NP from The Scripps Research Institute. This research was supported by NIH/NIDA Grants DA07348 and DA08467 (FW). We thank ND Stuempfig for technical assistance, M Arends for help with manuscript preparation, and Dr P Romieu (INSERM U. 575, Strasbourg, France) for constructive suggestions.
Author information
Authors and Affiliations
- Molecular and Integrative Neurosciences Department, The Scripps Research Institute, La Jolla, CA, USA
Rémi Martin-Fardon, Harinder Aujla & Friedbert Weiss - INSERM U. 710, EPHE, University of Montpellier II, Montpellier, France
Tangui Maurice - Department of Molecular Pharmacology, Physiology and Biotechnology, Brown University, Providence, RI, USA
Wayne D Bowen
Authors
- Rémi Martin-Fardon
You can also search for this author inPubMed Google Scholar - Tangui Maurice
You can also search for this author inPubMed Google Scholar - Harinder Aujla
You can also search for this author inPubMed Google Scholar - Wayne D Bowen
You can also search for this author inPubMed Google Scholar - Friedbert Weiss
You can also search for this author inPubMed Google Scholar
Corresponding author
Correspondence toRémi Martin-Fardon.
Rights and permissions
About this article
Cite this article
Martin-Fardon, R., Maurice, T., Aujla, H. et al. Differential Effects of _σ_1 Receptor Blockade on Self-Administration and Conditioned Reinstatement Motivated by Cocaine vs Natural Reward.Neuropsychopharmacol 32, 1967–1973 (2007). https://doi.org/10.1038/sj.npp.1301323
- Received: 01 August 2006
- Revised: 03 November 2006
- Accepted: 29 November 2006
- Published: 31 January 2007
- Issue Date: September 2007
- DOI: https://doi.org/10.1038/sj.npp.1301323