Complete genome sequence of Dehalogenimonas lykanthroporepellens type strain (BL-DC-9T) and comparison to “Dehalococcoides” strains (original) (raw)
- Open access
- Published: 25 May 2012
- Jean F. Challacombe1,
- Susana F. Delano1,
- Lance D. Green1,
- Hajnalka Daligault1,
- David Bruce1,
- Chris Detter1,
- Roxanne Tapia1,
- Shunsheng Han1,
- Lynne Goodwin1,
- James Han2,
- Tanja Woyke2,
- Sam Pitluck2,
- Len Pennacchio2,
- Matt Nolan2,
- Miriam Land3,
- Yun-Juan Chang3,
- Nikos C. Kyrpides2,
- Galina Ovchinnikova2,
- Loren Hauser3,
- Alla Lapidus2,4,
- Jun Yan5,
- Kimberly S. Bowman5,
- Milton S. da Costa6,
- Fred A. Rainey7 &
- …
- William M. Moe5
Standards in Genomic Sciences volume 6, pages 251–264 (2012)Cite this article
- 1305 Accesses
- 49 Citations
- 3 Altmetric
- Metrics details
Abstract
Dehalogenimonas lykanthroporepellens is the type species of the genus Dehalogenimonas, which belongs to a deeply branching lineage within the phylum Chloroflexi. This strictly anaerobic, mesophilic, non spore-forming, Gram-negative staining bacterium was first isolated from chlorinated solvent contaminated groundwater at a Superfund site located near Baton Rouge, Louisiana, USA. D. lykanthroporepellens was of interest for genome sequencing for two reasons: (a) an unusual ability to couple growth with reductive dechlorination of environmentally important polychlorinated aliphatic alkanes and (b) a phylogenetic position that is distant from previously sequenced bacteria. The 1,686,510 bp circular chromosome of strain BL-DC-9T contains 1,720 predicted protein coding genes, 47 tRNA genes, a single large subunit rRNA (23S-5S) locus, and a single, orphan, small subunit rRNA (16S) locus.
Introduction
Strain BL-DC-9T (=JCM 15061, =ATCC BAA-1523) is the type strain of the species Dehalogenimonas lykanthroporepellens, which is the type species of the genus Dehalogenimonas [[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed http://dx.doi.org/10.1099/ijs.0.011502-0
")\]. At the time of publication, _D. lykanthroporepellens_ is the only validly named species in this genus. The type strain was isolated from moderately acidic groundwater (pH=5.1) collected at a waste recovery well at the Petro-Processors of Louisiana, Inc. Superfund Site, near Baton Rouge, Louisiana (USA), in an area contaminated by high concentrations of several chlorinated alkanes and alkenes \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
")–[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. The strain is able to reductively dehalogenate a variety of environmentally important polychlorinated alkanes but not monochlorinated alkanes, chlorinated alkenes, or chlorinated benzenes \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
"),[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. Quantitative real-time PCR experiments indicate that bacteria closely related or identical to _D. lykanthroporepellens_ are present throughout the contaminated site from which strain BL-DC-9T was first isolated \[[4](/articles/10.4056/sigs.2806097#ref-CR4 "Yan J, Rash BA, Rainey FA, Moe WM. Detection and quantification of Dehalogenimonas and “Dehalococcoides” populations via PCR-based protocols targeting 16S rRNA genes. Appl Environ Microbiol 2009; 75:7560–7564. PubMed
http://dx.doi.org/10.1128/AEM.01938-09
")\]. In this report, we present a summary classification and a set of features for _D. lykanthroporepellens_ BL-DC-9T together with the description of the complete genomic sequencing and annotation.Classification and features
At present, D. lykanthroporepellens strain BL-DC-9T is phylogenetically isolated within the domain Bacteria, with no other species assigned to the genus Dehalogenimonas. On the basis of 16S rRNA gene sequences, strain BL-DC-9T clusters within the phylum Chloroflexi. Based on 16S rRNA gene sequences, the closest related type strains are Caldilinea tarbellica D1-25-10-4T [[5](/articles/10.4056/sigs.2806097#ref-CR5 "Grégoire P, Bohli M, Cayol JL, Joseph M, Guasco S, Dubourg K, Cambar J, Michotey V, Bonin P, Fardeau ML, Ollivier B. Caldilinea tarbellica sp. nov., a filamentous, thermophilic, anaerobic bacterium isolated from a deep hot aquifer in the Aquitaine Basin. Int J Syst Evol Microbiol 2011; 61:1436–1441. PubMed http://dx.doi.org/10.1099/ijs.0.025676-0
")\] and _Caldilinea aerophila_ STL-6-O1T \[[6](/articles/10.4056/sigs.2806097#ref-CR6 "Sekiguchi Y, Yamada T, Hanada S, Ohashi A, Harada H, Kamagata Y. Anaerolinea thermophila gen. nov., sp. nov. and Caldilinea aerophila gen. nov., sp. nov., novel filamentous thermophiles that represent a previously uncultured lineage of the domain Bacteria at the subphylum level. Int J Syst Evol Microbiol 2003; 53:1843–1851. PubMed
http://dx.doi.org/10.1099/ijs.0.02699-0
")\], with sequence identities of 81.7% and 81.5%, respectively \[[7](/articles/10.4056/sigs.2806097#ref-CR7 "Chun J, Lee JH, Jung Y, Kim M, Kim S, Kim BK, Lim YW. EzTaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int J Syst Evol Microbiol 2007; 57:2259–2261. PubMed
http://dx.doi.org/10.1099/ijs.0.64915-0
")\]. Aside from the closely related strain BL-DC-8 that was isolated from the same groundwater source as strain BL-DC-9T, the closest previously cultured phylogenetic relatives of strain BL-DC-9T are “_Dehalococcoides_” strains \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
"),[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. Although some variable regions of the 16S rRNA genes of _D. lykanthroporepellens_ and “_Dehalococcoides_” strains are highly homologous \[[4](/articles/10.4056/sigs.2806097#ref-CR4 "Yan J, Rash BA, Rainey FA, Moe WM. Detection and quantification of Dehalogenimonas and “Dehalococcoides” populations via PCR-based protocols targeting 16S rRNA genes. Appl Environ Microbiol 2009; 75:7560–7564. PubMed
http://dx.doi.org/10.1128/AEM.01938-09
")\], the overall identity of these genes is ∼90%, indicating a distant relationship \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
")\].Figure 1 shows the phylogenetic neighborhood of D. lykanthroporepellens strain BL-DC-9T in a 16S rRNA gene based phylogenetic dendrogram. The sequence of the lone 16S rRNA gene copy in the genome differs from the previously published 16S rRNA gene sequence (EU679419) by a single nucleotide position.
Figure 1.

Phylogenetic tree showing the position of D. lykanthroporepellens BL-DC-9T relative to other strains within the phylum Chloroflexi. The tree was inferred from 1,214 aligned nucleotide positions of the 16S rRNA gene sequence using the Neighbor-Joining method within the MEGA v4.0.2 package [[8](/articles/10.4056/sigs.2806097#ref-CR8 "Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol 2007; 24:1596–1599. PubMed http://dx.doi.org/10.1093/molbev/msm092
")\]. _Truepera radiovictrix_ RQ-24T is shown as an outgroup and was used to root the tree. Scale bar represents 2 substitutions per 100 nucleotide positions. Numbers at branching points denote support values from 1,000 bootstrap replicates if larger than 70%. Lineages with genome sequencing projects registered in GOLD \[[9](/articles/10.4056/sigs.2806097#ref-CR9 "Liolios K, Chen IM, Mavromatis K, Tavernarakis N, Hugenholtz P, Markowitz VM, Kyrpides NC. The Genomes On Line Database (GOLD) in 2009: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res 2010; 38:D346–D354. PubMed
http://dx.doi.org/10.1093/nar/gkp848
")\] are shown in blue, published genomes in bold: “_Dehalococcoides_” GT (CP001924), “_Dehalococcoides_” BAV1 (CP000688), “_Dehalococcoides_” CBDB1 (AJ965256), “_Dehalococcoides ethenogenes_” 195 (CP000027), “_Dehalococcoides_” VS (CP001827), _Dehalogenimonas lykanthroporepellens_ BL-DC-9T (CP002084), _Caldilinea aerophila_ STL-6-O1T (AP012337), _Anaerolinea thermophila_ UNI-1T (AP012029), “_Thermobaculum terrenum_” YNP1 (CP001825), _Sphaerobacter thermophilus_ DSM 20745T (CP001823), _Thermomicrobium roseum_ DSM 5159T (CP001275), _Herpetosiphon aurantiacus_ DSM 785T (CP000875), _Roseiflexus castenholzii_ DSM 13941T (CP000804), _Chloroflexus aurantiacus_ J-10-flT (CP000909), and _Chloroflexus aggregans_ MD-66T (CP001337).The cells of D. lykanthroporepellens stain Gram-negative, and are non-spore forming, irregular cocci with a diameter of 0.3–0.6 µm (Figure 2). Strains of D. lykanthroporepellens were isolated in liquid medium using a dilution-to-extinction approach. Growth was not observed on agar plates or on medium solidified with gellan gum, even after long term (2 months) incubation [[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. The temperature range for growth of strain BL-DC-9T is between 20°C and 37°C with an optimum between 28°C and 34°C \[[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. The pH range for growth is 6.0 to 8.0 with an optimum of 7.0 to 7.5 \[[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. The organism grows in the presence of 2% (w/v) NaCl and is resistant to ampicillin and vancomycin at concentrations of 1.0 and 0.1 g/l, respectively \[[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\].Figure 2.

Scanning electron micrograph of cells of D. lykanthroporepellens strain BL-DC-9T
D. lykanthroporepellens strain BL-DC-9T is a strictly anaerobic chemotroph, coupling utilization of H2 as an electron donor and several environmentally important polychlorinated aliphatic alkanes as electron acceptors for growth (Table 1). The chlorinated compounds known to be reductively dehalogenated include 1,2,3-trichloropropane, 1,2-dichloropropane, 1,1,2,2-tetrachloroethane, 1,1,2-trichloroethane, and 1,2-dichloroethane [[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. In all of the reductive dechlorination reactions characterized to date, strain BL-DC-9T appears to exclusively utilize vicinally halogenated alkanes as electron acceptors _via_ dihaloelimination reactions (i.e., simultaneous removal of two chlorine atoms from adjacent carbon atoms with concomitant formation of a carbon-carbon double bond) \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
"),[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. Strain BL-DC-9T does not utilize 1-chlorobenzene, 1-chloropropane, 2-chloropropane, 1,2-dichlorobenzene, _cis_\-1,2-dichloroethene, _trans_\-1,2-dichloroethene, tetrachloroethene, or vinyl chloride as electron acceptors for growth \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
"),[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\]. Growth is not supported by acetate, butyrate, citrate, ethanol, fructose, fumarate, glucose, lactate, lactose, malate, methanol, methyl ethyl ketone, propionate, pyruvate, succinate, or yeast extract in the absence of H2 \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
"),[3](/articles/10.4056/sigs.2806097#ref-CR3 "Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed
http://dx.doi.org/10.1111/j.14622920.2008.01804.x
")\].Table 1. Classification and general features of D. lykanthroporepellens strain BL-DC-9T according to the MIGS recommendations [[10](/articles/10.4056/sigs.2806097#ref-CR10 "Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, Tatusova T, Tompson N, Allen MJ, Angiuoli SV, et al. The minimum information about a genome sequence” (MIGS) specification. Nat Biotechnol 2008; 26:541–547. PubMed http://dx.doi.org/10.1038/nbt1360
")]
Chemotaxonomy
The major cellular fatty acids of strain BL-DC-9T, as identified and quantified with the Sherlock MIS v. 6.0 system (Microbial Identification, Inc.) using the Aerobe (TSBA) and MOORE libraries, are C18:1 ω 9c, C16:1 ω 9c, C16:0, and C14:0 [[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed http://dx.doi.org/10.1099/ijs.0.011502-0
")\]. The same fatty acids were also present in the closely related strain BL-DC-8 \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
")\]. Cellular fatty acids present in lower proportions include C18:0, C12:0, and the summed features listed in the MIDI Sherlock system as summed feature 5 (C18:2 _ω_ 6,9c and/or anteiso-C18:0) and summed feature 3 (one or more of C16:1 _ω_ 7 _c_ , C16:1 _ω_ 6 _c_ , iso-C15:03OH) \[[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed
http://dx.doi.org/10.1099/ijs.0.011502-0
")\].Genome sequencing and annotation
Genome project history
D. lykanthroporepellens strain BL-DC-9T was selected for sequencing on the basis of its phylogenetic position and the importance of reductive dechlorination in the field of environmental microbiology and bioremediation. A detailed understanding of the metabolic capabilities of chloroalkane-dehalogenating bacteria such as D. lykanthroporepellens has the potential to impact decision-making regarding site clean-up at thousands of DOE and non-DOE sites across the USA and around the world. D. lykanthroporepellens strain BL-DC-9T genome project is deposited in the Genomes OnLine Database [[9](/articles/10.4056/sigs.2806097#ref-CR9 "Liolios K, Chen IM, Mavromatis K, Tavernarakis N, Hugenholtz P, Markowitz VM, Kyrpides NC. The Genomes On Line Database (GOLD) in 2009: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res 2010; 38:D346–D354. PubMed http://dx.doi.org/10.1093/nar/gkp848
")\] and the complete genome sequence is available from GenBank. Sequencing, finishing, and annotation were performed by the DOE Joint Genome Institute (JGI). A summary of the project information is shown in Table [2](/articles/10.4056/sigs.2806097#Tab2).Table 2. Genome sequencing project information
Growth conditions and DNA isolation
D. lykanthroporepellens strain BL-DC-9T (=JCM 15061, =ATCC BAA-1523) was cultured in liquid anaerobic basal medium supplemented with 1,1,2-trichloroethane as described previously [[1](/articles/10.4056/sigs.2806097#ref-CR1 "Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed http://dx.doi.org/10.1099/ijs.0.011502-0
")\]. Cells were harvested from 2.0 L of culture medium by centrifugation (10,000×g, 10 min, 4°C). Total DNA was extracted from the resulting cell pellet using a combination of lysozyme/SDS/proteinase K treatment, followed by purification using hexadecyltrimethyl ammonium bromide (CTAB) in conjunction with phenol-chloroform-isoamyl alcohol purification, and ethanol precipitation \[[15](/articles/10.4056/sigs.2806097#ref-CR15 "IMG web site.
http://img.jgi.doe.gov/cgi-bin/w/main.cgi?section=TaxonDetail&page=cogs&cat=cat&taxon oid=648028022
")\].Genome sequencing and assembly
The genome of D. lykanthroporepellens BL-DC-9T was sequenced at the JGI using a combination of Illumina [[16](/articles/10.4056/sigs.2806097#ref-CR16 "Bennett S. Solexa Ltd. Pharmacogenomics 2004; 5:433–438. PubMed http://dx.doi.org/10.1517/14622416.5.4.433
")\] and 454 technologies \[[17](/articles/10.4056/sigs.2806097#ref-CR17 "Margulies M, Egholm M, Altman WE, Attiya S, Bader JS, Bemben LA, Berka J, Braverman MS, Chen YJ, Chen Z. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005; 437:376–380. PubMed")\]. An Illumina GAii shotgun library with reads of 152.4 Mb, a 454 Titanium draft library with average read length of 356.7±167 bases, and a paired end 454 library with average insert size of 19,767±4,941 kb were generated for this genome. All general aspects of library construction and sequencing performed at the JGI can be found at the JGI website \[[18](/articles/10.4056/sigs.2806097#ref-CR18 "DOE Joint Genome Institute.
http://www.jgi.doe.gov
")\]. Illumina sequencing data was assembled with VELVET \[[19](/articles/10.4056/sigs.2806097#ref-CR19 "Zerbino DR, Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res 2008; 18:821–829. PubMed
http://dx.doi.org/10.1101/gr.074492.107
")\], and the consensus sequences were shredded into 1.5 kb overlapped fake reads and assembled together with the 454 data. Draft assemblies were based on 103.1 Mb 454 standard draft data and all of the 454 paired end data (38,136 reads that were both mapped, non-redundant). Newbler parameters are -consed -a 50 -l 350 -g -m -ml 20.The initial Newbler assembly contained 64 contigs in 1 scaffold. The initial 454 assembly was converted into a phrap [[20](/articles/10.4056/sigs.2806097#ref-CR20 "The Phred/Phrap/Consed software package. http://www.phrap.com
")\] assembly by making fake reads from the consensus, and collecting the read pairs in the 454 paired end library. The Phred/Phrap/Consed software package \[[18](/articles/10.4056/sigs.2806097#ref-CR18 "DOE Joint Genome Institute.
http://www.jgi.doe.gov
")\] was used for sequence assembly and quality assessment \[[21](/articles/10.4056/sigs.2806097#ref-CR21 "Ewing B, Hillier L, Wendl MC, Green P. Base-calling of automated sequencer traces using phred. I. Accuracy assessment. Genome Res 1998; 8:175–185. PubMed")–[23](/articles/10.4056/sigs.2806097#ref-CR23 "Gordon D, Abajian C, Green P. Consed: a graphical tool for sequence finishing. Genome Res 1998; 8:195–202. PubMed")\] during the finishing process. After the shotgun stage, reads were assembled with parallel phrap (High Performance Software, LLC). Possible mis-assemblies were corrected with gapResolution \[Cliff Han, unpublished\], Dupfinisher \[[24](/articles/10.4056/sigs.2806097#ref-CR24 "Han CS, Chain P. Finishing repeat regions automatically with Dupfinisher. in Proceedings of the 2006 international conference on bioinformatics & computational biology. Arabnia HR, Valafar H (eds), CSREA Press. June 26–29, 2006: 141–146.")\], or sequencing cloned bridging PCR fragments with subcloning (Epicentre Biotechnologies, Madison, WI). Gaps between contigs were closed by editing in Consed, by PCR and by Bubble PCR primer walks. A total of 296 additional reactions were necessary to close gaps and to raise the quality of the finished sequence.The error rate of the completed genome sequence is less than 1 in 100 kb. Together, the combination of the Illumina and 454 sequencing platforms provided 170 × coverage of the genome. The final assembly contained 36,976 reads.
Genome annotation
Genes were identified using Prodigal [[25](/articles/10.4056/sigs.2806097#ref-CR25 "Hyatt D, Chen GL, Locascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 2010; 11:119. PubMed http://dx.doi.org/10.1186/1471-2105-11-119
")\] as part of the Oak Ridge National Laboratory genome annotation pipeline, followed by a round of manual curation using the JGI GenePRIMP pipeline \[[26](/articles/10.4056/sigs.2806097#ref-CR26 "Pati A, Ivanova N, Mikhailova N, Ovchinikova G, Hooper SD, Lykidis A, Kyrpides NC. GenePRIMP: A Gene Prediction Improvement Pipeline for microbial genomes. Nat Methods 2010; 7:455–457. PubMed
http://dx.doi.org/10.1038/nmeth.1457
")\]. The predicted CDSs were translated and used to search the National Center for Biotechnology Information (NCBI) nonredundant database, UniProt, TIGRFam, Pfam, PRIAM, KEGG, COG, and InterPro databases. These data sources were combined to assert a product description for each predicted protein. Non-coding genes and miscellaneous features were predicted using tRNAscan-SE \[[27](/articles/10.4056/sigs.2806097#ref-CR27 "Lowe TM, Eddy SR. tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res 1997; 25:955–964. PubMed")\], RNAMMer \[[28](/articles/10.4056/sigs.2806097#ref-CR28 "Lagesen K, Hallin PF, Rødland E, Stærfeldt HH, Rognes T, Ussery DW. RNammer: consistent annotation of rRNA genes in genomic sequences. Nucleic Acids Res 2007; 35:3100–3108. PubMed
http://dx.doi.org/10.1093/nar/gkm160
")\], Rfam \[[29](/articles/10.4056/sigs.2806097#ref-CR29 "Griffiths-Jones S, Bateman A, Marshall M, Khanna A, Eddy SR. Rfam: an RNA family database. Nucleic Acids Res 2003; 31:439–441. PubMed
http://dx.doi.org/10.1093/nar/gkg006
")\], TMHMM \[[30](/articles/10.4056/sigs.2806097#ref-CR30 "Krogh A, Larsson B, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J Mol Biol 2001; 305:567–580. PubMed
http://dx.doi.org/10.1006/jmbi.2000.4315
")\], and signal \[[31](/articles/10.4056/sigs.2806097#ref-CR31 "Bendtsen JD, Nielsen H, von Heijne G, Brunak S. Improved prediction of signal peptides: SignalP 3.0. J Mol Biol 2004; 340:783–795. PubMed
http://dx.doi.org/10.1016/j.jmb.2004.05.028
")\]. Additional gene prediction analysis and manual functional annotation were performed within the Integrated Microbial Genomes - Expert Review (IMG-ER) platform \[[32](/articles/10.4056/sigs.2806097#ref-CR32 "Markowitz VM, Mavromatis K, Ivanova NN, Chen IM, Chu K, Kyrpides NC. IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics 2009; 25:2271–2278. PubMed
http://dx.doi.org/10.1093/bioinformatics/btp393
")\].Genome properties
The genome of D. lykanthroporepellens strain BL-DC-9T comprises a single circular chromosome of 1,686,510 bp with 50.04% G+C content (Table 3 and Figure 3). Of the 1,771 genes predicted, 1,720 were protein-coding genes and 51 were RNAs; 61 pseudogenes were also identified. The majority of the protein-coding genes (68.8%) were assigned a putative function and those remaining were annotated as hypothetical proteins. The distribution of the predicted protein coding genes into COG functional categories is presented in Table 4.
Figure 3.
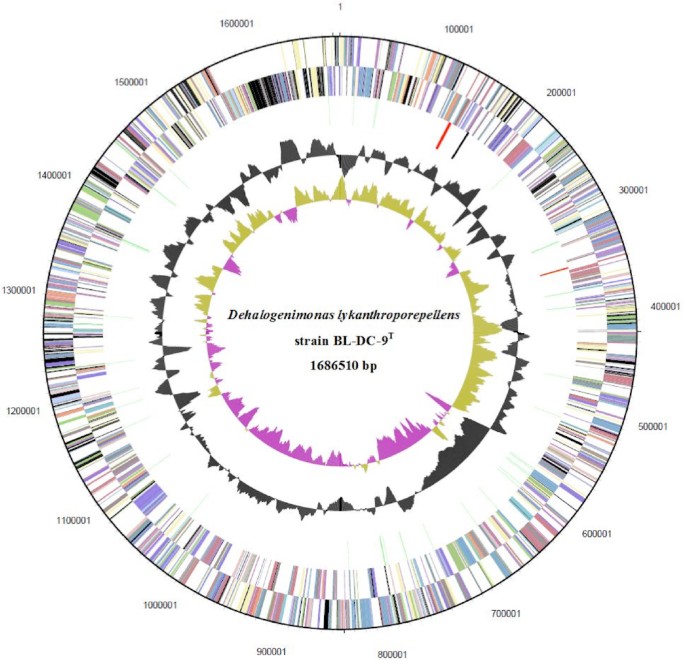
Graphical circular map of the chromosome of D. lykanthroporepellens strain BL-DC-9T. From outside to the center: Genes on the forward strand (color by COG categories), Genes on the reverse strand (color by COG categories), RNA genes (tRNAs green, rRNAs red, other RNAs black), GC content (peaks outside the circle indicate above average and peaks inside the circle indicate below average GC content), GC skew [calculated as (G − C) / (G + C); purple indicates values less than 1 and olive indicates values greater than 1]. Numbers outside the map denote nucleotide positions within the chromosome.
Table 3. Genome Statistics
Table 4. Number of genes associated with the general COG functional categories*
Insights from the genome sequence
Analysis of the complete genome sequence of strain BL-DC-9T and its comparison with the genomes of “_Dehalococcoides_” strains sequenced previously provide several insights into the evolution and adaptation of the organism to its niche. Transposon-mediated horizontal gene transfer appears to have played a major role in creating the genomic diversity and metabolic versatility in strain BL-DC-9T.
Horizontal gene transfer
Strain BL-DC-9T contains a prophage region (1,604,159 to 1,672,879 bp, ∼60% GC), which accounts for ∼4% of the chromosome. Of the 76 ORFs identified within this region, 45 have been annotated as encoding hypothetical proteins and a vast majority of these have no homologs in the public databases. In addition to the prophage region, manual curation indicated that ∼4.3% of the genome of strain BL-DC-9T (∼73,000 bp) is comprised of insertion sequence (IS) elements encoding 74 full-length or truncated transposases. These IS elements are scattered throughout the chromosome and their GC content varies from 47% to 57%. The IS elements of strain BL-DC-9T belong to the families IS256 (29 of 74), IS3/IS911 (14 of 74), IS3/IS600 (10 of 74), IS4/IS5 (7 of 74), IS4/IS5/ISMca7 (5 of 74), IS1182 (4 of 74), IS116/IS110/IS902 (2 of 74), IS204/IS1001/ISL3 (2 of 74), and IS6/ISCpe7 (1 of 74).
tRNAs and Selenocysteine utilization
The chromosome of strain BL-DC-9T contains 47 tRNA genes, including those for all 20 standard amino acids as well as the unusual amino acid selenocysteine. Proteins containing selenocysteine are found in all three domains of life and many organisms contain genes encoding the complex molecular machinery required for the incorporation of this modified amino acid during the translation process [33,[34](/articles/10.4056/sigs.2806097#ref-CR34 "Hendrickson TL. Easing selenocysteine into proteins. Nat Struct Mol Biol 2007; 14:100–101. PubMed http://dx.doi.org/10.1038/nsmb02 07-100
")\]. Strain BL-DC-9T contains an operon (_selCDAB_) putatively involved in selenocysteine biosynthesis. _selC_ encodes a selenocysteine-inserting tRNA (tRNAsec), which contains the complementary UCA anticodon for the internal UGA stop codon (Dehly\_R0051). A gene that is not part of this operon encodes a seryl-tRNA synthetase (Dehly\_0621), which catalyzes the aminoacylation of tRNAsec with serine. _selD_ encodes a selenophosphate synthetase (Dehly\_1500), an enzyme that produces monoselenophosphate using selenide and ATP as substrates. _selA_ encodes a selenocysteine synthase (Dehly\_1501), which utilizes monoselenophosphate as the selenium donor during the conversion of serine-acylated tRNAsec into selenocysteine-tRNAsec. _selB_ encodes a GTP-dependent selenocysteine-specific elongation factor (Dehly\_1502), which forms a quaternary complex with selenocysteine-tRNAsec and the selenocysteine inserting sequence (SECIS), which is a hairpin loop found immediately downstream of the UGA codon in the selenoprotein-encoding mRNA molecule \[[35](/articles/10.4056/sigs.2806097#ref-CR35 "Donovan J, Copeland PR. Evolutionary history of selenocysteine incorporation from the perspective of SECIS binding proteins. BMC Evol Biol 2009; 9:229. PubMed
http://dx.doi.org/10.1186/1471-2148-9-229
")\]. This complex ensures reading through the UGA codon and incorporation of selenocysteine, instead of termination of translation \[[36](/articles/10.4056/sigs.2806097#ref-CR36 "Zavacki AM, Mansell JB, Chung M, Klimovitsky B, Harney JW, Berry MJ. Coupled tRNA(Sec)-dependent assembly of the selenocysteine decoding apparatus. Mol Cell 2003; 11:773–781. PubMed
http://dx.doi.org/10.1016/S1097-2765(03)00064-9
")\]. Consistent with the presence of the genes encoding the synthesis and incorporation of selenocysteine, strain BL-DC-9T also contains a gene encoding a selenocysteine-containing formate dehydrogenase (Dehly\_0033). This gene has an internal in-frame UGA stop codon (574 bp from the AUG start codon), which is followed by a 48 bp putative SECIS element.Strain BL-DC-9T contains a putative IS256 element immediately downstream of the selCDAB operon (Dehly_1503, transposase). Previous phylogenetic analysis has provided evidence for horizontal transfer of these traits [[37](/articles/10.4056/sigs.2806097#ref-CR37 "Romero H, Zhang Y, Gladyshev VN, Salinas G. Evolution of selenium utilization traits. Genome Biol 2005; 6:R66. PubMed http://dx.doi.org/10.1186/gb-2005-6-8-r66
")\]. The presence of an IS element adjacent to the _sel_ genes in strain BL-DC-9T suggests horizontal transfer and explains the absence of this locus in “_Dehalococcoides_” strains sequenced previously \[[38](/articles/10.4056/sigs.2806097#ref-CR38 "Kube M, Beck A, Zinder SH, Kuhl H, Reinhardt R, Adrian L. Genome sequence of the chlorinated compound-respiring bacterium Dehalococcoides species strain CBDB1. Nat Biotechnol 2005; 23:1269–1273. PubMed
http://dx.doi.org/10.1038/nbt1131
")–[40](/articles/10.4056/sigs.2806097#ref-CR40 "McMurdie PJ, Behrens SF, Muller JA, Göke J, Ritalahti KM, Wagner R, Goltsman E, Lapidus A, Holmes S, Löffler FE, Spormann AM. Localized plasticity in the streamlined genomes of vinyl chloride respiring Dehalococcoides. PLoS Genet 2009; 5:e1000714. PubMed
http://dx.doi.org/10.1371/journal.pgen.1000714
")\].Comparative genomics
The chromosome of strain BL-DC-9T is 216,790 bp larger than that of “_Dehalococcoides ethenogenes_” strain 195, which has the largest genome among “_Dehalococcoides_” strains sequenced to date. The difference in the size of the chromosomes of strain BL-DC-9T and “_Dehalococcoides_” strains is partly due to the presence of multiple IS elements and IS element-associated genes in the former. These putative horizontally transferred genes appear to have played a major role in creating genomic diversity and phenotypic variability of strain BL-DC-9T vis-à-vis “_Dehalococcoides_” strains.
Although the chromosomes of strain BL-DC-9T and “_Dehalococcoides_” strains contain similar number of rRNA and tRNA encoding genes, they differ in their GC content, gene density, and percentage of sequence that encodes proteins. The lack of synteny between the chromosomes as well as the observed differences in the general features of the genomes of strain BL-DC-9T and “_Dehalococcoides_” strains insinuate their highly divergent evolutionary paths. This divergent evolutionary past is further supported by a phylogenetic tree constructed based on 432 core orthologous protein encoding genes shared between D. lykanthroporepellens BL-DC-9T and “_Dehalococcoides_” (meta)genomes [[41](/articles/10.4056/sigs.2806097#ref-CR41 "McMurdie PJ, Hug LA, Edwards EA, Holmes S, Spormann AM. Site-specific mobilization of vinyl chloride respiration islands by a mechanism common in Dehalococcoides. BMC Genomics 2011; 12:287. PubMed http://dx.doi.org/10.1186/14712164-12-287
")\]. In contrast, the chromosomes of four “_Dehalococcoides_” strains obtained from diverse geographic regions share a conserved core (1,029 orthologous groups of protein encoding genes conserved across all four genomes, with genes generally sharing the same order, orientation and synteny) that is interrupted by two high plasticity regions, indicating comparatively recent divergence from a common ancestor \[[40](/articles/10.4056/sigs.2806097#ref-CR40 "McMurdie PJ, Behrens SF, Muller JA, Göke J, Ritalahti KM, Wagner R, Goltsman E, Lapidus A, Holmes S, Löffler FE, Spormann AM. Localized plasticity in the streamlined genomes of vinyl chloride respiring Dehalococcoides. PLoS Genet 2009; 5:e1000714. PubMed
http://dx.doi.org/10.1371/journal.pgen.1000714
")\].BLAST comparisons of protein sets of strain BL-DC-9T and “_Dehalococcoides ethenogenes_” strain 195 revealed that the two strains contain ∼950 protein coding genes in common (bidirectional best hits, 20–90% identity at the predicted protein level). Pairwise blast comparisons indicated that strain BL-DC-9T contains ∼700 protein-coding genes with no homologs in strain 195. The latter contained ∼600 protein-coding genes with no homologs in BL-DC-9T. Genome-specific genes identified in strains BL-DC-9T and 195 encoded transposases, DNA endonucleases/methylases, heterodisulfide reductases, acetyltransferases, kinases, phosphatases, and dehalogenases. Some of these strain-specific genes were found within prophage-like regions or were associated with IS elements.
Biosynthesis and transport of compatible solutes
A number of microorganisms accumulate low molecular weight organic compounds alternately referred to as osmolytes, osmoprotectants, or “compatible solutes” to convey the notion that the compounds help the microorganisms survive osmotic stress but do not interfere with metabolism [[42](/articles/10.4056/sigs.2806097#ref-CR42 "Santos H, da Costa MS. Compatible solutes in organisms that live in hot saline environments. Environ Microbiol 2002; 4:501–509. PubMed http://dx.doi.org/10.1046/j.14622920.2002.00335.x
")\]. Ectoine, glycine-betaine, and proline are compatible solutes of many mesophilic bacteria capable of survival at high salt concentrations \[[42](/articles/10.4056/sigs.2806097#ref-CR42 "Santos H, da Costa MS. Compatible solutes in organisms that live in hot saline environments. Environ Microbiol 2002; 4:501–509. PubMed
http://dx.doi.org/10.1046/j.14622920.2002.00335.x
")\]. Many thermophilic organisms accumulate compatible solutes, such as mannosylglycerate and di-myo-inositol phosphate, which generally do not occur in mesophilic organisms \[[43](/articles/10.4056/sigs.2806097#ref-CR43 "Empadinhas N, Albuquerque L, Costa J, Zinder SH, Santos MA, Santos H, da Costa MS. A gene from the mesophilic bacterium Dehalococcoides ethenogenes encodes a novel mannosylglycerate synthase. J Bacteriol 2004; 186:4075–4084. PubMed
http://dx.doi.org/10.1128/JB.186.13.4075-4084.2004
")\].Strain BL-DC-9T contains an operon (ectABC) encoding putative homologs of the enzymes involved in ectoine biosynthesis and regulation (Dehly_1306, Dehly_1307, Dehly_1308). The closest homologs of strain BL-DC-9T_ectABC_ are found in Halomonas elongata, Wolinella succinogenes, and Desulfococcus oleovorans (48-75% identity at the predicted protein level). At least two putative transport systems for the compatible solutes proline/glycine-betaine have been identified in strain BL-DC-9T (proVWX and opuABCD). proV, proW, and proX encode an ATPase subunit (Dehly_0378), a permease protein (Dehly_0377), and a periplasmic subunit (Dehly_0376), respectively. opuA, opuB, opuC, and opuD encode a periplasmic substrate-binding protein (Dehly_0909), a permease protein (Dehly_0908), an ATPase subunit (Dehly_0907), and a permease protein (Dehly_0906), respectively. Although the permeases encoded by opuB, opuD, and proW as well as the ATPase subunits encoded by opuC and proV appear to be related to each other (34-40% identity at the predicted protein level), the periplasmic proteins encoded by opuA and proX are unrelated. The closest homologs of proVWX are found in Trichodesmium erythraeum, Marinomonas sp. MED121, and Fulvimarina pelagi (50% identity at the predicted protein level), whereas those of opuABCD are found in Pseudovibrio sp. JE062, Chromohalobacter salexigens DSM 3043, and Denitrovibrio acetiphilus DSM 12809 (44-60% identity at the predicted protein level). Strain BL-DC-9T also contains genes involved in the biosynthesis of proline (Dehly_0299, Dehly_0308). “_Dehalococcoides_” strains lack homologs of ectABC, proVWX, and opuABCD, but contain homologs of Dehly_0299 and Dehly_0308 (57 and 68% protein identity, respectively).
Homologs of a gene encoding a bifunctional mannosylglycerate synthase (mgsD) are found in “_Dehalococcoides_” strains (e.g., DET1363), an unusual occurrence for mesophilic bacteria [[43](/articles/10.4056/sigs.2806097#ref-CR43 "Empadinhas N, Albuquerque L, Costa J, Zinder SH, Santos MA, Santos H, da Costa MS. A gene from the mesophilic bacterium Dehalococcoides ethenogenes encodes a novel mannosylglycerate synthase. J Bacteriol 2004; 186:4075–4084. PubMed http://dx.doi.org/10.1128/JB.186.13.4075-4084.2004
")\]. Although the synthesis and accumulation of mannosylglycerate could not be proven to occur in “_D. ethenogenes_” because of insufficient biomass, the role of the bifunctional _mgsD_ was confirmed by cloning and expression in _Saccharomyces cerevisiae_ \[[43](/articles/10.4056/sigs.2806097#ref-CR43 "Empadinhas N, Albuquerque L, Costa J, Zinder SH, Santos MA, Santos H, da Costa MS. A gene from the mesophilic bacterium Dehalococcoides ethenogenes encodes a novel mannosylglycerate synthase. J Bacteriol 2004; 186:4075–4084. PubMed
http://dx.doi.org/10.1128/JB.186.13.4075-4084.2004
")\]. Comparative analysis revealed that BL-DC-9T contains a homologous gene (Dehly\_0877, 54% protein identity). This expands the range of species containing genes putatively involved in the biosynthesis of compatible solutes and may offer _D. lykanthroporepellens_ a stress response mechanism that allows growth under conditions of changing osmolarity.Reductive dehalogenases
Genes encoding the enzymes that are involved in catalyzing the reductive dehalogenation of chlorinated solvents are organized in rdhAB operons encoding two components: a 50–65 kDa protein (RdhA) that functions as a reductive dehalogenase and a ∼10 kDa hydrophobic protein with transmembrane helices (RdhB) that is thought to anchor the RdhA to the cytoplasmic membrane [[44](/articles/10.4056/sigs.2806097#ref-CR44 "Adrian L, Rahnenfuhrer J, Gobom J, Holscher T. Identification of a chlorobenzene reductive dehalogenase in Dehalococcoides sp. strain CBDB1. Appl Environ Microbiol 2007; 73:7717–7724. PubMed http://dx.doi.org/10.1128/AEM.01649-07
")–[53](/articles/10.4056/sigs.2806097#ref-CR53 "Thibodeau J, Gauthier A, Duguay M, Villemur R, Lepine F, Juteau P, Beaudet R. Purification, cloning, and sequencing of a 3,5-dichlorophenol reductive dehalogenase from Desulfitobacterium frappieri PCP-1. Appl Environ Microbiol 2004; 70:4532–4537. PubMed
http://dx.doi.org/10.1128/AEM.70.8.4532-4537.2004
")\]. Comparative genomic analyses revealed that strain BL-DC-9T contains several loci related to _rdhA_ and/or _rdhB_ genes scattered throughout the chromosome. The multiple _rdhA_ and _rdhB_ ORFs of strain BL-DC-9T have 24–74% and 24–65% identities at the predicted protein level, respectively. The closest homologs of _rdhA_ ORFs are found among “_Dehalococcoides_” strains (31-78% identity at the predicted protein level). A twin-arginine motif, with the predicted amino acid sequence SRRXFMK followed by a stretch of hydrophobic amino acids, was identified in the N-terminus of a large majority (19 of 25) of predicted RdhA sequences. Consistent with the presence of the twin-arginine sequence in the N-terminus of most of its RdhA sequences, strain BL-DC-9T contains an operon encoding proteins that constitute a putative twin-arginine translocation (TAT) system (Dehly\_1346-1349). This specialized system is involved in the secretion of folded proteins across the bacterial inner membrane into the periplasmic space \[[54](/articles/10.4056/sigs.2806097#ref-CR54 "Stanley NR, Palmer T, Berks BC. The twin arginine consensus motif of Tat signal peptides is involved in Sec-independent protein targeting in Escherichia coli. J Biol Chem 2000; 275:11591–11596. PubMed
http://dx.doi.org/10.1074/jbc.275.16.11591
"),[55](/articles/10.4056/sigs.2806097#ref-CR55 "Sargent F, Berks BC, Palmer T. Pathfinders and trailblazers: a prokaryotic targeting system for transport of folded proteins. FEMS Microbiol Lett 2006; 254:198–207. PubMed
http://dx.doi.org/10.1111/j.1574-6968.2005.00049.x
")\]. “_Dehalococcoides_” strains also contain an operon encoding an analogous TAT system that is partially related to the TAT system of strain BL-DC-9T (38 and 64% protein identity).Two conserved motifs, each containing four cysteine residues, a feature associated with binding iron-sulfur clusters [56], were identified near the C-terminus of 22 of the predicted RdhA sequences of strain BL-DC-9T. The first of these motifs had a consistent number of amino acids between the cysteine residues (CX2CX2CX3C). In the second motif, there were variable numbers of intervening amino acids after the first and second cysteine residues (CX2-21CX2-7CX3C). If a “full-length” rdhA is predicted to encode a protein containing a twin-arginine sequence in the N-terminus, two iron-sulfur cluster binding motifs in the C-terminus, and an intervening sequence of ∼450-500 aa, then strain BL-DC-9T contains 17 such genes. Two rdhA genes (Dehyl_0069 and 1582) appear to be truncated and are predicted to encode proteins lacking the twin-arginine sequences in their N-termini. Five rdhA genes appear to be substantially truncated and are predicted to encode proteins consisting of only the N-terminus (Dehly_0479 and 1520) or the C-terminus (Dehly_0075, 1523, and 1534).
Within strain BL-DC-9T, only six of the rdhA ORFs have a cognate rdhB, and an additional rdhB gene (Dehly_1504) appears to be an orphan with no cognate rdhA ORF nearby. The predicted RdhB sequences of strain BL-DC-9T contain two or three transmembrane helices. Similar features have been observed among the predicted RdhB sequences of “_Dehalococcoides_” strains [[38](/articles/10.4056/sigs.2806097#ref-CR38 "Kube M, Beck A, Zinder SH, Kuhl H, Reinhardt R, Adrian L. Genome sequence of the chlorinated compound-respiring bacterium Dehalococcoides species strain CBDB1. Nat Biotechnol 2005; 23:1269–1273. PubMed http://dx.doi.org/10.1038/nbt1131
")–[40](/articles/10.4056/sigs.2806097#ref-CR40 "McMurdie PJ, Behrens SF, Muller JA, Göke J, Ritalahti KM, Wagner R, Goltsman E, Lapidus A, Holmes S, Löffler FE, Spormann AM. Localized plasticity in the streamlined genomes of vinyl chloride respiring Dehalococcoides. PLoS Genet 2009; 5:e1000714. PubMed
http://dx.doi.org/10.1371/journal.pgen.1000714
")\]. Furthermore, the predicted RdhB sequences of strain BL-DC-9T and the “_Dehalococcoides_” strains share 40–72% identity. In at least seven loci (Dehly\_0069, 0075, 0479, 1504, 1530, 1534, and 1541), it appears that transposon insertion has truncated one or both of the _rdh_ genes. Interestingly, genes involved in the regulation of _rdhAB_ operons (_e.g._, MarR-type or two-component transcriptional regulators) were present only in seven loci (Dehly\_0121, 0274, 0479, 1148, 1355, 1530, and 1582). In addition to the genes encoding reductive dehalogenases, the strain BL-DC-9T genome contains two genes encoding putative haloacid dehalogenases (Dehly\_0588, Dehly\_1126) that have homologs among the “_Dehalococcoides_” strains (40-44% identity at the predicted protein level).The presence of IS elements adjacent to some rdhA/rdhB loci in strain BL-DC-9T indicates their acquisition from an unknown host. Previous studies of “_Dehalococcoides_” strains have also suggested horizontal transfer of reductive dehalogenase genes [[41](/articles/10.4056/sigs.2806097#ref-CR41 "McMurdie PJ, Hug LA, Edwards EA, Holmes S, Spormann AM. Site-specific mobilization of vinyl chloride respiration islands by a mechanism common in Dehalococcoides. BMC Genomics 2011; 12:287. PubMed http://dx.doi.org/10.1186/14712164-12-287
"),[57](/articles/10.4056/sigs.2806097#ref-CR57 "Krajmalnik-Brown R, Sung Y, Ritalahti KM, Saunders FM, Löffler FE. Environmental distribution of the trichloroethene reductive dehalogenase gene (tceA) suggests lateral gene transfer among Dehalococcoides. FEMS Microbiol Ecol 2007; 59:206–214. PubMed
http://dx.doi.org/10.1111/j.1574-6941.2006.00243.x
")\]. It remains to be determined if strain BL-DC-9T_rdhA_ genes lacking an _rdhB_ ORF downstream encode functional reductive dehalogenases and whether/how they are membrane-bound. It is possible that an incognate or a non-contiguous _rdhB_ (_e.g._, the orphan Dehly\_1504) could complement one or more of strain BL-DC-9T_rdhA_ genes lacking an _rdhB_ ORF downstream. Alternatively, some of these genes may encode reductive dehalogenases that function by an unknown mechanism. An enzyme involved in the reductive dehalogenation of tetrachloroethene by _Dehalospirillum multivorans_ was found in the cytoplasmic fraction \[[58](/articles/10.4056/sigs.2806097#ref-CR58 "Miller E, Wohlfarth G, Diekert G. Studies on tetrachloroethene respiration in Dehalospirillum multivorans. Arch Microbiol 1996; 166:379–387. PubMed
http://dx.doi.org/10.1007/BF01682983
")\], suggesting that some reductive dehalogenases are either loosely membrane-bound or soluble entities. The same may be the case for the majority of reductive dehalogenases of strain BL-DC-9T. Regardless, the repertoire of _rdhA_/_rdhB_ loci identified by complete genome sequencing sets the stage for future efforts to elucidate the mechanism of reductive dehalogenation by strain BL-DC-9T and other _Dehalogenimonas_ strains.References
- Moe WM, Yan J, Nobre MF, da Costa MS, Rainey FA. Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductive dehalogenating bacterium isolated from chlorinated solvent contaminated groundwater. Int J Syst Evol Microbiol 2009; 59:2692–2697. PubMed http://dx.doi.org/10.1099/ijs.0.011502-0
Article CAS PubMed Google Scholar - Bowman KS, Moe WM, Rash BA, Bae HS, Rainey FA. Bacterial diversity of an acidic Louisiana groundwater contaminated by DNAPL containing chloroethanes and other solvents. FEMS Microbiol Ecol 2006; 58:120–133. PubMed http://dx.doi.org/10.1111/j.15746941.2006.00146.x
Article CAS PubMed Google Scholar - Yan J, Rash BA, Rainey FA, Moe WM. Isolation of novel bacteria within the Chloroflexi capable of reductive dechlorination of 1,2,3-trichloropropane. Environ Microbiol 2009; 11:833–843. PubMed http://dx.doi.org/10.1111/j.14622920.2008.01804.x
Article CAS PubMed Google Scholar - Yan J, Rash BA, Rainey FA, Moe WM. Detection and quantification of Dehalogenimonas and “_Dehalococcoides_” populations via PCR-based protocols targeting 16S rRNA genes. Appl Environ Microbiol 2009; 75:7560–7564. PubMed http://dx.doi.org/10.1128/AEM.01938-09
Article PubMed Central CAS PubMed Google Scholar - Grégoire P, Bohli M, Cayol JL, Joseph M, Guasco S, Dubourg K, Cambar J, Michotey V, Bonin P, Fardeau ML, Ollivier B. Caldilinea tarbellica sp. nov., a filamentous, thermophilic, anaerobic bacterium isolated from a deep hot aquifer in the Aquitaine Basin. Int J Syst Evol Microbiol 2011; 61:1436–1441. PubMed http://dx.doi.org/10.1099/ijs.0.025676-0
Article PubMed Google Scholar - Sekiguchi Y, Yamada T, Hanada S, Ohashi A, Harada H, Kamagata Y. Anaerolinea thermophila gen. nov., sp. nov. and Caldilinea aerophila gen. nov., sp. nov., novel filamentous thermophiles that represent a previously uncultured lineage of the domain Bacteria at the subphylum level. Int J Syst Evol Microbiol 2003; 53:1843–1851. PubMed http://dx.doi.org/10.1099/ijs.0.02699-0
Article CAS PubMed Google Scholar - Chun J, Lee JH, Jung Y, Kim M, Kim S, Kim BK, Lim YW. EzTaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int J Syst Evol Microbiol 2007; 57:2259–2261. PubMed http://dx.doi.org/10.1099/ijs.0.64915-0
Article CAS PubMed Google Scholar - Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol 2007; 24:1596–1599. PubMed http://dx.doi.org/10.1093/molbev/msm092
Article CAS PubMed Google Scholar - Liolios K, Chen IM, Mavromatis K, Tavernarakis N, Hugenholtz P, Markowitz VM, Kyrpides NC. The Genomes On Line Database (GOLD) in 2009: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res 2010; 38:D346–D354. PubMed http://dx.doi.org/10.1093/nar/gkp848
Article PubMed Central CAS PubMed Google Scholar - Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, Tatusova T, Tompson N, Allen MJ, Angiuoli SV, et al. The minimum information about a genome sequence” (MIGS) specification. Nat Biotechnol 2008; 26:541–547. PubMed http://dx.doi.org/10.1038/nbt1360
Article PubMed Central CAS PubMed Google Scholar - Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc Natl Acad Sci USA 1990; 87:4576–4579. PubMed http://dx.doi.org/10.1073/pnas.87.12.4576
Article PubMed Central CAS PubMed Google Scholar - Garrity GM, Holt JG. Phylum BVI. Chloroflexi phy. nov. In: Garrity GM, Boone DR, Castenholz RW (eds), Bergey’s Manual of Systematic Bacteriology, Second Edition, Volume 1, Springer, New York, 2001, p. 427–446.
Chapter Google Scholar - Hugenholtz P, Stackebrandt E. Reclassification of Sphaerobacter thermophilus from the subclass Sphaerobacteridae in the phylum Actinobacteria to the class Thermomicrobia (emended description) in the phylum Chloroflexi (emended description). Int J Syst Evol Microbiol 2004; 54:2049–2051. PubMed http://dx.doi.org/10.1099/ijs.0.03028-0
Article PubMed Google Scholar - Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 2000; 25:25–29. PubMed http://dx.doi.org/10.1038/75556
Article PubMed Central CAS PubMed Google Scholar - IMG web site. http://img.jgi.doe.gov/cgi-bin/w/main.cgi?section=TaxonDetail&page=cogs&cat=cat&taxon oid=648028022
- Bennett S. Solexa Ltd. Pharmacogenomics 2004; 5:433–438. PubMed http://dx.doi.org/10.1517/14622416.5.4.433
Article PubMed Google Scholar - Margulies M, Egholm M, Altman WE, Attiya S, Bader JS, Bemben LA, Berka J, Braverman MS, Chen YJ, Chen Z. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005; 437:376–380. PubMed
PubMed Central CAS PubMed Google Scholar - DOE Joint Genome Institute. http://www.jgi.doe.gov
- Zerbino DR, Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res 2008; 18:821–829. PubMed http://dx.doi.org/10.1101/gr.074492.107
Article PubMed Central CAS PubMed Google Scholar - The Phred/Phrap/Consed software package. http://www.phrap.com
- Ewing B, Hillier L, Wendl MC, Green P. Base-calling of automated sequencer traces using phred. I. Accuracy assessment. Genome Res 1998; 8:175–185. PubMed
Article CAS PubMed Google Scholar - Ewing B, Green P. Base-calling of automated sequencer traces using phred. II. Error probabilities. Genome Res 1998; 8:186–194. PubMed
Article CAS PubMed Google Scholar - Gordon D, Abajian C, Green P. Consed: a graphical tool for sequence finishing. Genome Res 1998; 8:195–202. PubMed
Article CAS PubMed Google Scholar - Han CS, Chain P. Finishing repeat regions automatically with Dupfinisher. in Proceedings of the 2006 international conference on bioinformatics & computational biology. Arabnia HR, Valafar H (eds), CSREA Press. June 26–29, 2006: 141–146.
- Hyatt D, Chen GL, Locascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 2010; 11:119. PubMed http://dx.doi.org/10.1186/1471-2105-11-119
Article PubMed Central PubMed Google Scholar - Pati A, Ivanova N, Mikhailova N, Ovchinikova G, Hooper SD, Lykidis A, Kyrpides NC. GenePRIMP: A Gene Prediction Improvement Pipeline for microbial genomes. Nat Methods 2010; 7:455–457. PubMed http://dx.doi.org/10.1038/nmeth.1457
Article CAS PubMed Google Scholar - Lowe TM, Eddy SR. tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res 1997; 25:955–964. PubMed
Article PubMed Central CAS PubMed Google Scholar - Lagesen K, Hallin PF, Rødland E, Stærfeldt HH, Rognes T, Ussery DW. RNammer: consistent annotation of rRNA genes in genomic sequences. Nucleic Acids Res 2007; 35:3100–3108. PubMed http://dx.doi.org/10.1093/nar/gkm160
Article PubMed Central CAS PubMed Google Scholar - Griffiths-Jones S, Bateman A, Marshall M, Khanna A, Eddy SR. Rfam: an RNA family database. Nucleic Acids Res 2003; 31:439–441. PubMed http://dx.doi.org/10.1093/nar/gkg006
Article PubMed Central CAS PubMed Google Scholar - Krogh A, Larsson B, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J Mol Biol 2001; 305:567–580. PubMed http://dx.doi.org/10.1006/jmbi.2000.4315
Article CAS PubMed Google Scholar - Bendtsen JD, Nielsen H, von Heijne G, Brunak S. Improved prediction of signal peptides: SignalP 3.0. J Mol Biol 2004; 340:783–795. PubMed http://dx.doi.org/10.1016/j.jmb.2004.05.028
Article PubMed Google Scholar - Markowitz VM, Mavromatis K, Ivanova NN, Chen IM, Chu K, Kyrpides NC. IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics 2009; 25:2271–2278. PubMed http://dx.doi.org/10.1093/bioinformatics/btp393
Article CAS PubMed Google Scholar - Böck A. Selenium proteins containing selenocysteine. In: Crabtree RH (ed.), Encyclopedia of inorganic chemistry. 2006. John Wiley & Sons, Ltd, New York.
Google Scholar - Hendrickson TL. Easing selenocysteine into proteins. Nat Struct Mol Biol 2007; 14:100–101. PubMed http://dx.doi.org/10.1038/nsmb02 07-100
Article CAS PubMed Google Scholar - Donovan J, Copeland PR. Evolutionary history of selenocysteine incorporation from the perspective of SECIS binding proteins. BMC Evol Biol 2009; 9:229. PubMed http://dx.doi.org/10.1186/1471-2148-9-229
Article PubMed Central PubMed Google Scholar - Zavacki AM, Mansell JB, Chung M, Klimovitsky B, Harney JW, Berry MJ. Coupled tRNA(Sec)-dependent assembly of the selenocysteine decoding apparatus. Mol Cell 2003; 11:773–781. PubMed http://dx.doi.org/10.1016/S1097-2765(03)00064-9
Article CAS PubMed Google Scholar - Romero H, Zhang Y, Gladyshev VN, Salinas G. Evolution of selenium utilization traits. Genome Biol 2005; 6:R66. PubMed http://dx.doi.org/10.1186/gb-2005-6-8-r66
Article PubMed Central PubMed Google Scholar - Kube M, Beck A, Zinder SH, Kuhl H, Reinhardt R, Adrian L. Genome sequence of the chlorinated compound-respiring bacterium Dehalococcoides species strain CBDB1. Nat Biotechnol 2005; 23:1269–1273. PubMed http://dx.doi.org/10.1038/nbt1131
Article CAS PubMed Google Scholar - Seshadri R, Adrian L, Fouts DE, Eisen JA, Phillippy AM, Methe BA, Ward NL, Nelson WC, Deboy RT, Khouri HM, et al. Genome sequence of the PCE-dechlorinating bacterium Dehalococcoides ethenogenes. Science 2005; 307:105–108. PubMed http://dx.doi.org/10.1126/science.1102226
Article CAS PubMed Google Scholar - McMurdie PJ, Behrens SF, Muller JA, Göke J, Ritalahti KM, Wagner R, Goltsman E, Lapidus A, Holmes S, Löffler FE, Spormann AM. Localized plasticity in the streamlined genomes of vinyl chloride respiring Dehalococcoides. PLoS Genet 2009; 5:e1000714. PubMed http://dx.doi.org/10.1371/journal.pgen.1000714
Article PubMed Central PubMed Google Scholar - McMurdie PJ, Hug LA, Edwards EA, Holmes S, Spormann AM. Site-specific mobilization of vinyl chloride respiration islands by a mechanism common in Dehalococcoides. BMC Genomics 2011; 12:287. PubMed http://dx.doi.org/10.1186/14712164-12-287
Article PubMed Central CAS PubMed Google Scholar - Santos H, da Costa MS. Compatible solutes in organisms that live in hot saline environments. Environ Microbiol 2002; 4:501–509. PubMed http://dx.doi.org/10.1046/j.14622920.2002.00335.x
Article CAS PubMed Google Scholar - Empadinhas N, Albuquerque L, Costa J, Zinder SH, Santos MA, Santos H, da Costa MS. A gene from the mesophilic bacterium Dehalococcoides ethenogenes encodes a novel mannosylglycerate synthase. J Bacteriol 2004; 186:4075–4084. PubMed http://dx.doi.org/10.1128/JB.186.13.4075-4084.2004
Article PubMed Central CAS PubMed Google Scholar - Adrian L, Rahnenfuhrer J, Gobom J, Holscher T. Identification of a chlorobenzene reductive dehalogenase in Dehalococcoides sp. strain CBDB1. Appl Environ Microbiol 2007; 73:7717–7724. PubMed http://dx.doi.org/10.1128/AEM.01649-07
Article PubMed Central CAS PubMed Google Scholar - Fung JM, Morris RM, Adrian L, Zinder SH. Expression of reductive dehalogenase genes in Dehalococcoides ethenogenes strain 195 growing on tetrachloroethene, trichloroethene, or 2,3-dichlorophenol. Appl Environ Microbiol 2007; 73:4439–4445. PubMed http://dx.doi.org/10.1128/AEM.00215-07
Article PubMed Central CAS PubMed Google Scholar - Hölscher T, Gorisch H, Adrian L. Reductive dehalogenation of chlorobenzene congeners in cell extracts of Dehalococcoides sp. strain CBDB1. Appl Environ Microbiol 2003; 69:2999–3001. PubMed http://dx.doi.org/10.1128/AEM.69.5.2999-3001.2003
Article PubMed Central PubMed Google Scholar - Magnuson JK, Romine MF, Burris DR, Kingsley MT. Trichloroethene reductive dehalogenase from Dehalococcoides ethenogenes: sequence of tceA and substrate range characterization. Appl Environ Microbiol 2000; 66:5141–5147. PubMed http://dx.doi.org/10.1128/AEM.66.12.5141-5147.2000
Article PubMed Central CAS PubMed Google Scholar - Müller JA, Rosner BM, von Abendroth G, Meshulam-Simon G, McCarty PL, Spormann AM. Molecular identification of the catabolic vinyl chloride reductase from Dehalococcoides sp. strain VS and its environmental distribution. Appl Environ Microbiol 2004; 70:4880–4888. PubMed http://dx.doi.org/10.1128/AEM.70.8.4880-4888.2004
Article PubMed Central PubMed Google Scholar - Maillard J, Schumacher W, Vazquez F, Regeard C, Hagen WR, Holliger C. Characterization of the corrinoid iron-sulfur protein tetrachloroethene reductive dehalogenase of Dehalobacter restrictus. Appl Environ Microbiol 2003; 69:4628–4638. PubMed http://dx.doi.org/10.1128/AEM.69.8.4628-4638.2003
Article PubMed Central CAS PubMed Google Scholar - Neumann A, Wohlfarth G, Diekert G. Tetrachloroethene dehalogenase from Dehalospirillum multivorans: cloning, sequencing of the encoding genes, and expression of the pceA gene in Escherichia coli. J Bacteriol 1998; 180:4140–4145. PubMed
PubMed Central CAS PubMed Google Scholar - Nijenhuis I, Zinder SH. Characterization of hydrogenase and reductive dehalogenase activities of Dehalococcoides ethenogenes strain 195. Appl Environ Microbiol 2005; 71:1664–1667. PubMed http://dx.doi.org/10.1128/AEM.71.3.1664-1667.2005
Article PubMed Central CAS PubMed Google Scholar - Suyama A, Yamashita M, Yoshino S, Furukawa K. Molecular characterization of the PceA reductive dehalogenase of Desulfitobacterium sp. strain Y51. J Bacteriol 2002; 184:3419–3425. PubMed http://dx.doi.org/10.1128/JB.184.13.3419-3425.2002
Article PubMed Central CAS PubMed Google Scholar - Thibodeau J, Gauthier A, Duguay M, Villemur R, Lepine F, Juteau P, Beaudet R. Purification, cloning, and sequencing of a 3,5-dichlorophenol reductive dehalogenase from Desulfitobacterium frappieri PCP-1. Appl Environ Microbiol 2004; 70:4532–4537. PubMed http://dx.doi.org/10.1128/AEM.70.8.4532-4537.2004
Article PubMed Central CAS PubMed Google Scholar - Stanley NR, Palmer T, Berks BC. The twin arginine consensus motif of Tat signal peptides is involved in Sec-independent protein targeting in Escherichia coli. J Biol Chem 2000; 275:11591–11596. PubMed http://dx.doi.org/10.1074/jbc.275.16.11591
Article CAS PubMed Google Scholar - Sargent F, Berks BC, Palmer T. Pathfinders and trailblazers: a prokaryotic targeting system for transport of folded proteins. FEMS Microbiol Lett 2006; 254:198–207. PubMed http://dx.doi.org/10.1111/j.1574-6968.2005.00049.x
Article CAS PubMed Google Scholar - Bruschi M, Guerlesquin F. Structure, function and evolution of bacterial ferredoxins. FEMS Microbiol Rev 1988; 4:155–175. PubMed
Article CAS PubMed Google Scholar - Krajmalnik-Brown R, Sung Y, Ritalahti KM, Saunders FM, Löffler FE. Environmental distribution of the trichloroethene reductive dehalogenase gene (tceA) suggests lateral gene transfer among Dehalococcoides. FEMS Microbiol Ecol 2007; 59:206–214. PubMed http://dx.doi.org/10.1111/j.1574-6941.2006.00243.x
Article CAS PubMed Google Scholar - Miller E, Wohlfarth G, Diekert G. Studies on tetrachloroethene respiration in Dehalospirillum multivorans. Arch Microbiol 1996; 166:379–387. PubMed http://dx.doi.org/10.1007/BF01682983
Article CAS PubMed Google Scholar
Acknowledgements
The work conducted by the U.S. Department of Energy Joint Genome Institute was supported by the Office of Science of the U.S. Department of Energy under Contract No. DE-AC02-05CH11231. The authors gratefully acknowledge Xiao Ying for assistance with microscopy.
Author information
Authors and Affiliations
- Bioscience Division, Los Alamos National Laboratory, Los Alamos, New Mexico, USA
Shivakumara Siddaramappa, Jean F. Challacombe, Susana F. Delano, Lance D. Green, Hajnalka Daligault, David Bruce, Chris Detter, Roxanne Tapia, Shunsheng Han & Lynne Goodwin - DOE Joint Genome Institute, Walnut Creek, California, USA
James Han, Tanja Woyke, Sam Pitluck, Len Pennacchio, Matt Nolan, Nikos C. Kyrpides, Galina Ovchinnikova & Alla Lapidus - Oak Ridge National Laboratory, Oak Ridge, Tennessee, USA
Miriam Land, Yun-Juan Chang & Loren Hauser - Lawrence Berkeley National Laboratory, Berkeley, California, USA
Alla Lapidus - Louisiana State University, Baton Rouge, Louisiana, USA
Jun Yan, Kimberly S. Bowman & William M. Moe - Department of Life Sciences, University of Coimbra, Coimbra, Portugal
Milton S. da Costa - University of Alaska Anchorage, Anchorage, Alaska, USA
Fred A. Rainey
Authors
- Shivakumara Siddaramappa
You can also search for this author inPubMed Google Scholar - Jean F. Challacombe
You can also search for this author inPubMed Google Scholar - Susana F. Delano
You can also search for this author inPubMed Google Scholar - Lance D. Green
You can also search for this author inPubMed Google Scholar - Hajnalka Daligault
You can also search for this author inPubMed Google Scholar - David Bruce
You can also search for this author inPubMed Google Scholar - Chris Detter
You can also search for this author inPubMed Google Scholar - Roxanne Tapia
You can also search for this author inPubMed Google Scholar - Shunsheng Han
You can also search for this author inPubMed Google Scholar - Lynne Goodwin
You can also search for this author inPubMed Google Scholar - James Han
You can also search for this author inPubMed Google Scholar - Tanja Woyke
You can also search for this author inPubMed Google Scholar - Sam Pitluck
You can also search for this author inPubMed Google Scholar - Len Pennacchio
You can also search for this author inPubMed Google Scholar - Matt Nolan
You can also search for this author inPubMed Google Scholar - Miriam Land
You can also search for this author inPubMed Google Scholar - Yun-Juan Chang
You can also search for this author inPubMed Google Scholar - Nikos C. Kyrpides
You can also search for this author inPubMed Google Scholar - Galina Ovchinnikova
You can also search for this author inPubMed Google Scholar - Loren Hauser
You can also search for this author inPubMed Google Scholar - Alla Lapidus
You can also search for this author inPubMed Google Scholar - Jun Yan
You can also search for this author inPubMed Google Scholar - Kimberly S. Bowman
You can also search for this author inPubMed Google Scholar - Milton S. da Costa
You can also search for this author inPubMed Google Scholar - Fred A. Rainey
You can also search for this author inPubMed Google Scholar - William M. Moe
You can also search for this author inPubMed Google Scholar
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article
Cite this article
Siddaramappa, S., Challacombe, J.F., Delano, S.F. et al. Complete genome sequence of Dehalogenimonas lykanthroporepellens type strain (BL-DC-9T) and comparison to “_Dehalococcoides_” strains.Stand in Genomic Sci 6, 251–264 (2012). https://doi.org/10.4056/sigs.2806097
- Published: 25 May 2012
- Issue Date: March 2012
- DOI: https://doi.org/10.4056/sigs.2806097