HSP90_evolutionのブログ (original) (raw)
2024 年10月9日
前回紹介した論文に引き続き、Cellに掲載された論文で、デンマークの2つの研究グループと都医学研の正井久雄さんの研究グループが共同で行った研究結果である。読んでみると、前の論文より分かりやすいので、こちらを先に紹介すべきであった。重複するところもあるが、そのまま紹介する。
文献19:Charlton, S. J. et al. Cell 187: 5029-5047 (2024)
The fork protection complex promotes partial histone recycling and epigenetic memory
Replisomeの構造的解析によれば、Mrc1 (ヒトではCLASPIN), Tof1(ヒトではTimeless; fission yeastではSwi1), Csm3(ヒトではTipin、fission yeastではSwi3)よりなるfork protection complex (FPC) がreplication forksを安定化させている(図1)。Fission yeast FPC mutantsはheterochromatin欠損変異(centromeres, telomeres, mating-type regionなどの異常)として得られた。Replisome はnucleosome assembly factors, histone modification enzymesやrecycling of modified parental histonesのrecruitを仲介しており、parental and newly synthesized histonesの子細胞のDNA鎖への配分も支配している。著者たちは、fission yeastのgeneticsとゲノム解析によって、epigenetic inheritanceにおけるFPCの役割を解明しようとした。
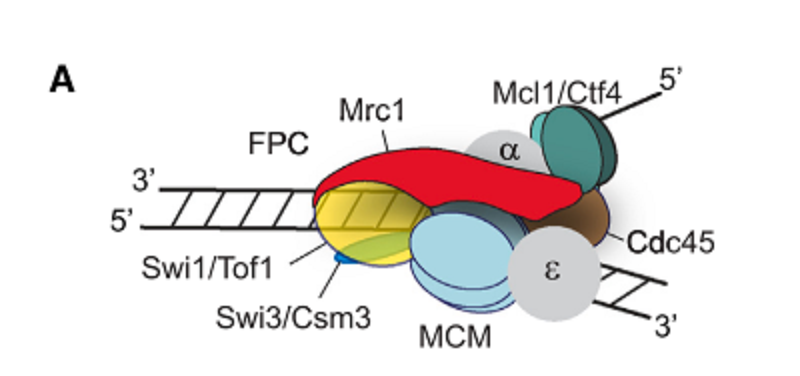
図1.文献19より引用。
Heterochromatin (HC) の維持にはFPCが必要である。
Fission yeastのmating type regionのHCを実験系として使った。cenH elementのHCをRNAiによって樹立し、Atf1-binding sitesからdomain boundaries IR-Lまで全領域に拡張させた。一度HCは構築されると、数百回の細胞分裂にわたって、RNAiと_cenH_の関与なしに維持された(図2)。HC樹立は_cenH_に隣接する部位に挿入したYFP発現変化で調べ、HCの拡張は縁側に挿入した_mCherry_の発現変化でモニターした。
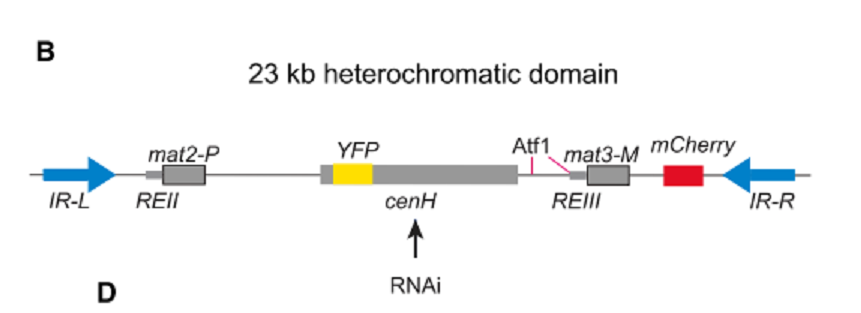
図2.文献19より引用。
Clr4Δ細胞(H3K9me HCを欠損する)に_clr4_ geneを導入することで、de novo HC formationのrateを測定した。HC形成にそれぞれ_swi1_Δ, _swi3_Δ, _mrc1_Δがどう影響するかを見た(図3)。その結果、FPCの欠損はHC構築形成には必要がないことが示された。同時に、FPCに欠損がある場合、継代25代程度続けると、HCは完璧には維持されなかった。Clonalな培養によって、_mCherry_の発現抑制(HCの維持)が、二つの娘細胞の一つで、壊れる場合が観察された(HCのasymmetric伝達)。
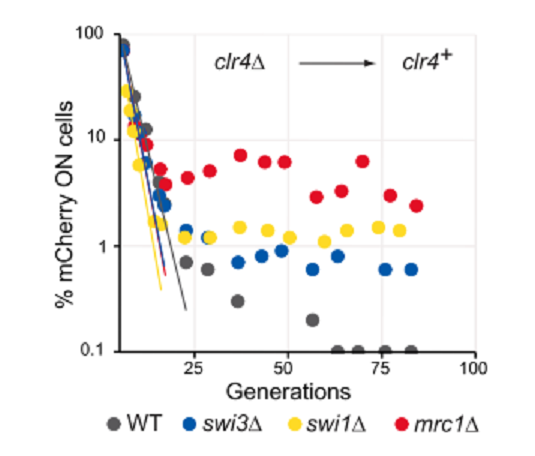
図3.文献19より引用。
Mrc1 のHC維持の機能はcheckpointにおける役割とは異なる
図4のMrc1のdomain構成を参照のこと。Mrc1Δ、ΔHBS (Hsk-bypass segment)、ΔNTHBSはどれもsilencingに欠損を示した(図5B,C)。HBS mutantsはcheckpoint機能があったが、NTHBS mutantsは欠損していた(図5D).したがって、Mrc1のHBS domainがHC維持機能の鍵となっている。さらに、Mrc1とHsk1はどちらもHC gene silencingで互いに独立に機能している、データが示されている。
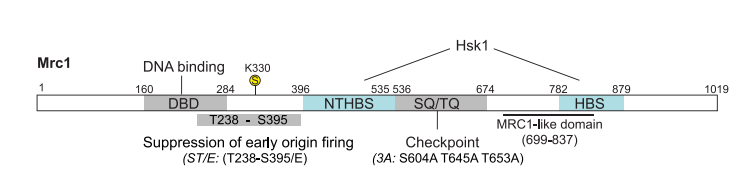
図4.文献19より引用。
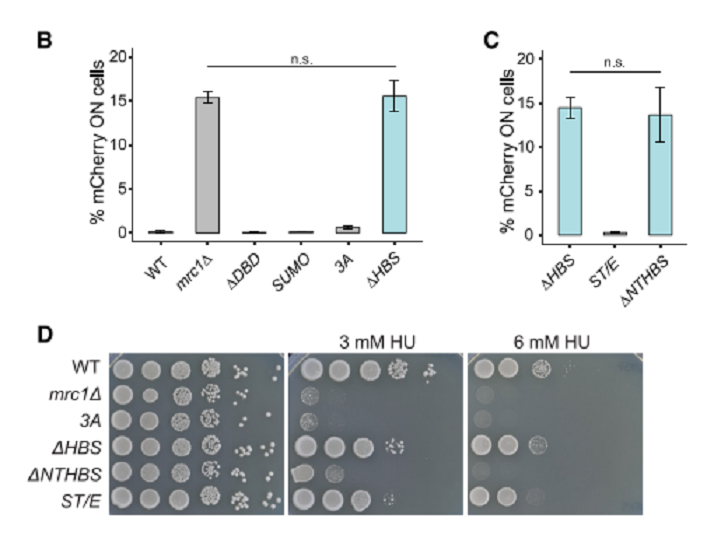
図5.文献19より引用。
HC 維持におけるMrc1とMcm2の共同作業
図6Aに示したように、pombe_と_cerevisiae_は共に、Mrc1 HBSにDDDSDDED (795-801)およびKAF (833-835)配列を有している。これらの配列をAに改変したところ、silencingに欠損を生じたが、checkpoint機能には影響しなかった。Cross-linking mass spectrometry or cryo-EMの結果から推定したreplisome factorsと_cerevisiae Mrc1のcontact pointsは、pombe Mrc1にもあるNTHBSあるいはHBS domainsで、Mcm2とcontactしていた(図6A)。仮に、Mcm2 amino-terminal HBDがparental histone recyclingに関わっているとすれば、Mrc1との共同作業が考えられる。Pombe_のMcm2 HBDのY80 and Y89(他の主でBHに必要とわかっている)をAに痴漢したmutantを作った。Mcm2-2A mutantでは、_mCherry reporterの抑制がなくなった(図6B)。このsilencingの消失はmrc1 mutantsと同じ性質を示した。これらの結果から、Mrc1とMcm2は共同でHC維持に作用していることが示唆された。
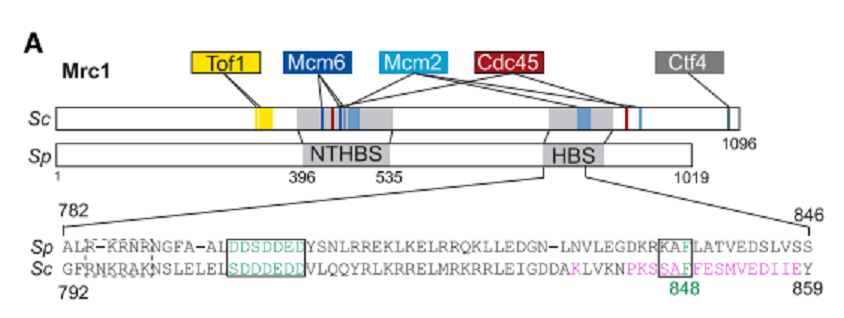
図6A. 文献19より引用。
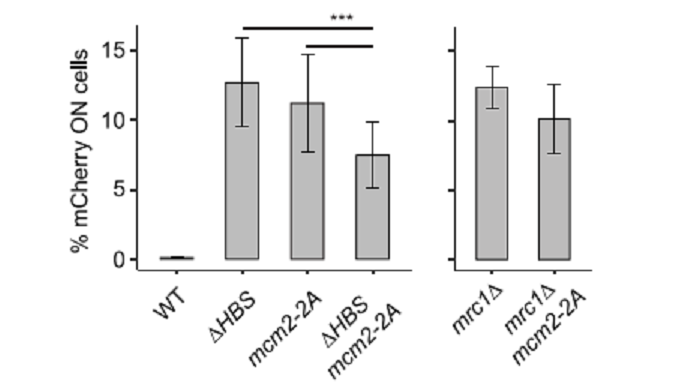
図6B. 文献19より引用。
Mrc1ΔHBSとmcm2-2Aはsub-telomeric regionsのH3K9me2を消失させたが、RNAi activityの部位のH3K9me2はそのままだった(図6C)。このことは、HC構築を支える強いシグナルがMrc1の作用の他にあることを想像させる。そこで、HCの_cenH_ RNAi nucleation siteのdeletionを作っても、HCは壊れなかった。しかし、mrc1ΔHBSを欠損させると、H3K9me2の部分的欠損が現れた。したがって、Mrc1とmcm2 double mutantでH3K9me2が残ったのは、ΔK部位にsilencing elemtntsが残っていたのかもしれない。
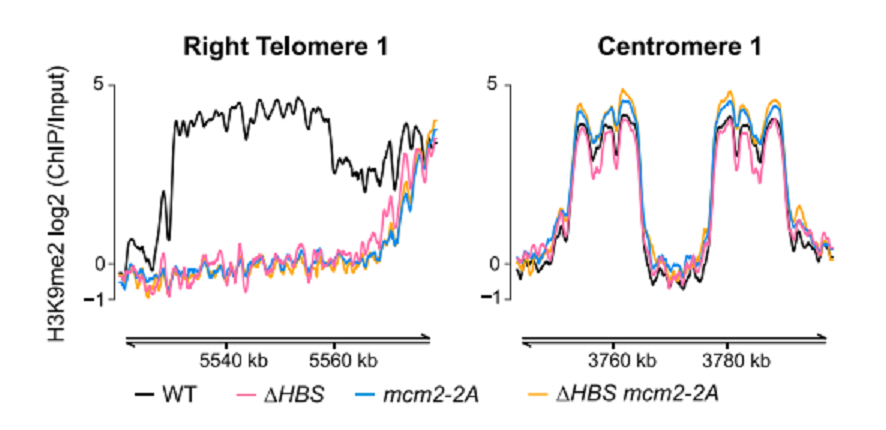
図6C.文献19より引用。
Mrc1のHBSがhistone recyclingで機能しているとすると、Mrc1ΔHBSにGFPをfuseさせ、そこにPob3-GBPを加えて、HBS欠損を補えるかどうかを見た(図7左)。結果は、silencing欠損がある程度補償された(図7右).これらの結果は、Mrc1 HBSがreplication forkにおけるhistone chaperone activityを提供していることを示唆する。
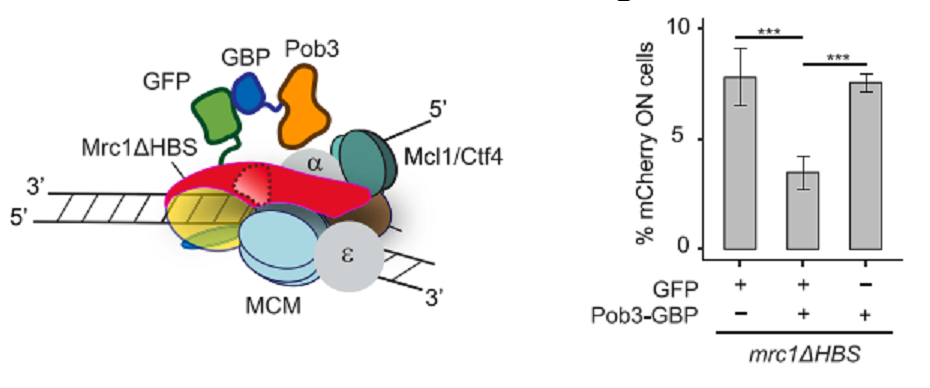
図7.文献19より引用。
Parental histones のlagging strandへのrecycling
DNAが複製したとき、parental histonesとnew histonesがleading and lagging strandsにどう分配されるかを調べた。H3K36me3をparental histonesの印とし、H4K20me0をnew histonesの印とした。Fission yeastでは、cell cycle M-phaseのすぐ後にS-phaseが始まるので、cold-sensitive tubulin mutantを使い、S-phase開始に同調させておき、さらにそのとき、合成されたDNAはEdUでラベルした。histone markersでparental histoneとnew histoneがそれぞれ結合しているDNA断片を取り分けた。さらに、DNA2本鎖を分けて、EdUを含むDNA鎖を分別し、sequencingを行った(図8)。S-phaseのDNA合成開始点はsequencingの結果から同定できる。
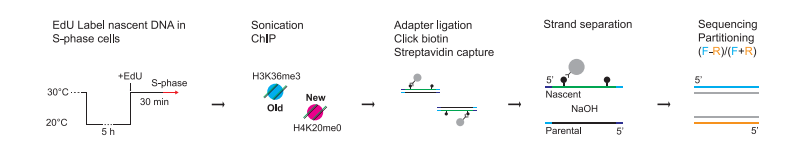
図8.文献19より引用。
図9に示したように、WTでは、parental histoneのleading strandとlagging strandへの結合はsymmetricalであるが、mrc1ΔBSやmcm2-2Aの変異株では、leading strandに偏っていた。当然、new histoneは逆になった。結論として、Mrc1はMcm2と同様に、parental histonesをlagging strandに振り分ける働きをして、WTにおけるhistones分配のバランスをとっている。
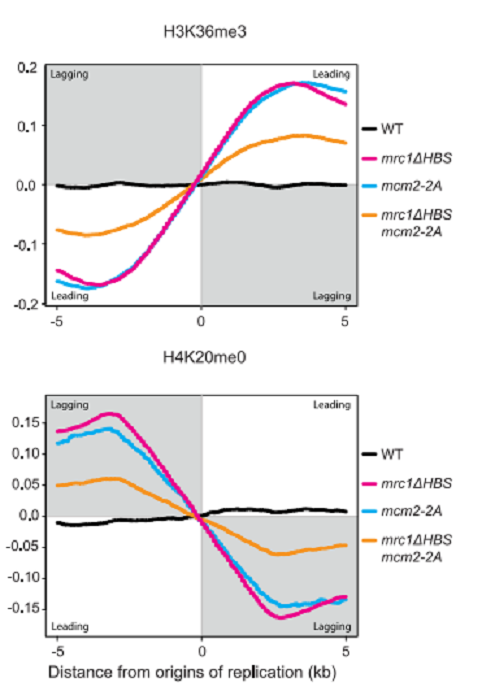
図9.文献19より引用。
構造的観点からみたMrc1のhistone recyclingでの役割
Mrc1は、場合によってはMcm2と共に、histonesに結合し、他の多くの因子と共同でhistoneのco-chaperoning機能を発揮しているであろう。既に、FACTとAsf1は、replisome componentsのMCM2やPolαと共に、histone co-chaperoningを行っていることが報告されている。これらのことから、soluble chaperonesがhistonesを、replisome中をhistone-binding interfacesをたどって、しかるべき部位に運ぶと考えられる。AlphaFold構造予測によれば、Mrc1は単独、あるいはMcm2と共に、nucleosome中と同じ構造をとっているH3-H4 tetramerに結合する。Mrc1の710-790(HBSの先の部分を含む:図10参照)の部分がH3-H4 dimerを包み込み、長いα-helixをbridgeとしてもう一つのH3-H4とつながっている(図10A)。この結合様式は、結晶構造解析によって示された、mammalian Mcm2がH3-H4を包み込む様式と同じである。
Mrc1 HBSはMrc1 HBDとMcm2の連結を担っている(KAF motifがMcm2のF204、L208などが形成するhydrophobic grooveに接している(図10A)。DSE motifは両者のflexible connective linkerとして機能している(図10i)。これらのHBSが両者の連結にかかわる構造は、XL-MSやcryo-EMのデータと一致している。H3-H4側からみると、HBSのR784からの配列にH4のC-terminusやdimer中のH3のL1-linkerなどに接している(図10ii)。
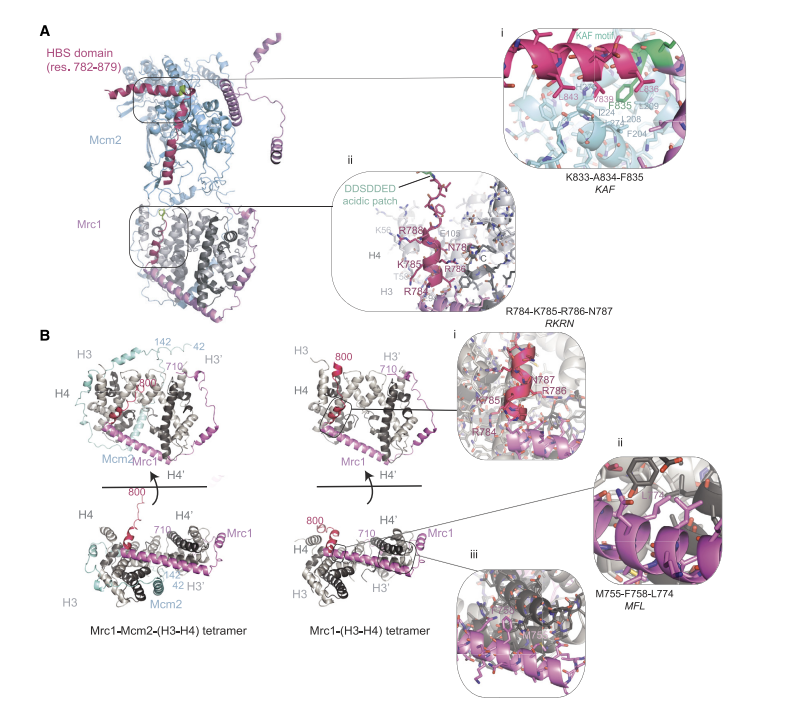
図10.文献19より引用。
Pull-down assay で検証したMrc1とH3-H4の結合
Full-length Mrc1と、HBDは有するがMrc1 binding siteを欠損したMcm2によるpull-down assayを行った(図11)。H3-H4が存在するときにのみ、Mrc1はMcm2 mutant共沈した。すなわち、Mrc1とMcm2 mutantは独立に、しかも同時にH3-H4に結合する。また、Mrc1はH3-H4 dimerよりも、ずっと強くcross-linked H3-H4 tetramerに結合した。図10Bにある、RKRN residueおよびMFL residueは、それぞれlong Mrc1 helixをtwo H3-H4 dimersに結合が予測されたが、それぞれの残基をAlanineに置換したmutantsでは、実際にin vitro H3-H4 dime bindingが弱かった。それらのmutantsは細胞レベルでみた、silencing効果も弱かった(図12)。RKRN mutantでは、parental histonesのleading strandへの振り分けが強くみられた(図13)。しかし、MFL mutantでは、逆に、lagging strandへのparental histonesのほうが優先された(図13)。すなわち、Mrc1はparental histonesのleading and lagging strandの両方で機能している(ただし、後者ではMcm2と共同で)。これらのMrc1の分子機能は、conserved MRC1-like domainのHBD、HBS connectorおよびMcm2-binding regionsにある。
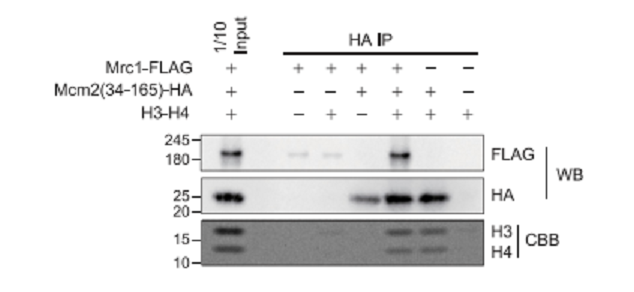
図11.文献19より引用。
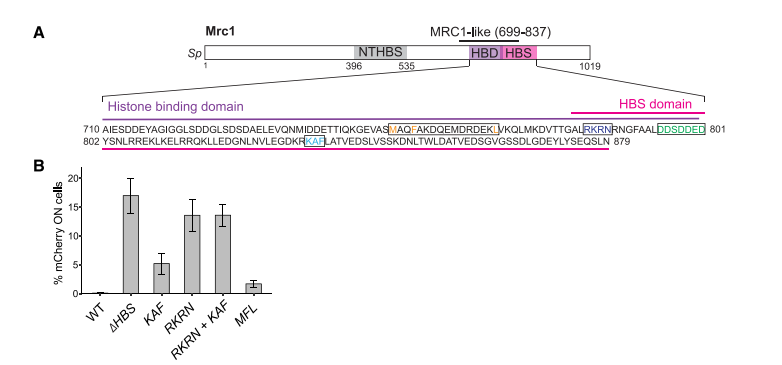
図12.文献19より引用。
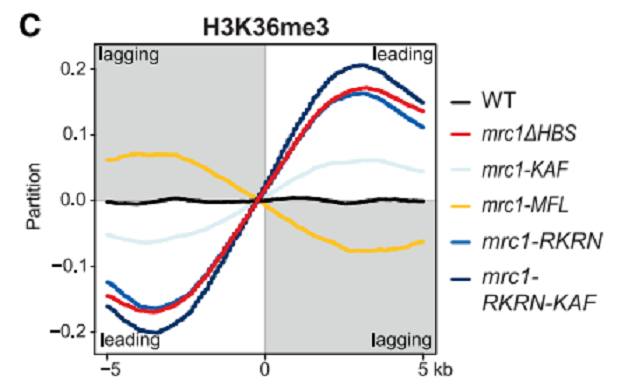
図13. 文献19より引用。
最後に、Mrc1のmammalian homologであるCLASPINも、mouse ESCsにおいて、parental H3-H4のleading strandへのrecyclingについての短い記述がある。
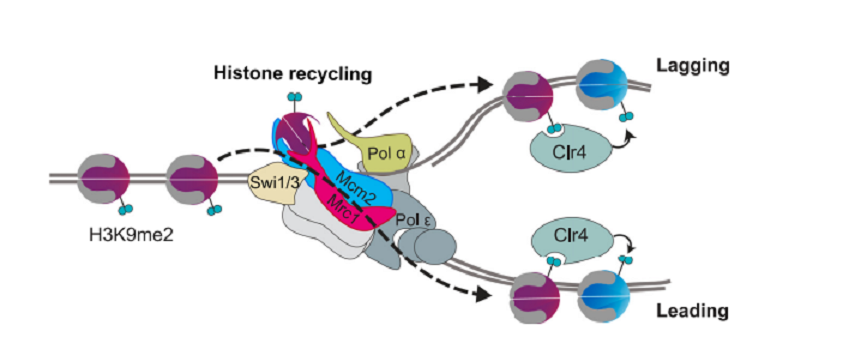
図14. 文献19より引用。
まとめとして、H3K9me2 histonesがreplisome上でleading and lagging strandsに振り分けられるモデルが示されてある(図14)。Lagging strandへ振り分けられるhistoneはMrc1とMcm2の両者に結合している。
2024 年9月29日投稿
ヒストン情報の親細胞から子細胞への伝達
最新のCellに次の論文が掲載されていたので、僕の勉強もかねて、紹介する。本論文では、例えば、1つの幹細胞が同じ性質をもつ2つの子細胞に分裂するとき、親DNAのchromatin構築が子細胞のDNAに移る仕組みを明らかにしようとしている。
文献18: Yu, et al. Cell 187: 5010-5028 (2024), A replisome-associated histone H3-H4 chaperone required for epigenetic inheritance
_Schizosaccharomyces pombe_(分裂酵母)の実験系で、H3K9me3(heterochromatin形成の主役)の親DNA鎖から子DNA鎖への伝達(以後単に伝達とする)において、Cir4/Suv39h methyltransferaseが必須であることが示されている。DNA複製時に、親DNA鎖のH3K9me3の位置を読み取り、子のDNAの同じ位置にコピーを作り、親と同じgene silencingを引き起こす。
Nucleosomeは147bp DNA,とoctameric histone complex (two H2A-H2B dimers and H3-H4 tetramer)からなる。DNA複製の時、親鎖のH3-H4 tetramerはそのまま子鎖(の一方)に移る。H2A-H2BはDNA複製時にrecycleされたりするdynamicな挙動を示す。Cdc45-Mcm2-7-GINS (CMG) replicative helicaseのMcm2 subunit, Pol1 catalytic subunit of the DNA polymeraseα, Dpb3-Dpb4 subunit of DNA polymeraseε, single-strand binding protein complex RPA, replication licensing factor Mcm10などのreplisome componentsがH3-H4に結合している。新たに合成されたDNA鎖に親DNAに結合していたhistonesが正しく対称的に(leading strandとlagging strandの上に)配分結合されるには、これらのreplisome-binding componentsの作用が必要である。これらのcomponentsの変異によって、偏ったhistone配分となることが知られている。これ以外にも、the facilitates chromatin transaction (FACT) complexがreplicationにcoupleしたnucleosome assemblyにかかわっている。
実験系の説明
1.目的:heterochromatin (HC)のectopic domainを誘導形成する。
2.10XtetO-ade6+ reporter geneをeuchromatic locusに挿入した。
3.Bacterial tetracycline repressor (TeTR)をH3K9 methyltransferase Clr4にfuseさせた。TetR-Clr4ΔCDあるいはTetR-Clr4-initiator (TetR-Clr4-I)を使った。
4.TerR-Clr4-IがTetOに結合すると、45kb H3K9me2/3が形成され、ade6 reporter geneがsilencingされる(図1)。
5.細胞をanhydrotetracycline (AHT)存在下に培養すると、TerR-Clr4-Iが遊離し、HCはそのまま次世代にepigenetic inheritanceする。
6.HC構造の形成と維持を区別してみることができ、HC構造の維持に必要な因子をscreeningすることができる(AHT存在下に_Ade6_の発現が抑制されるか、どうかによって)。
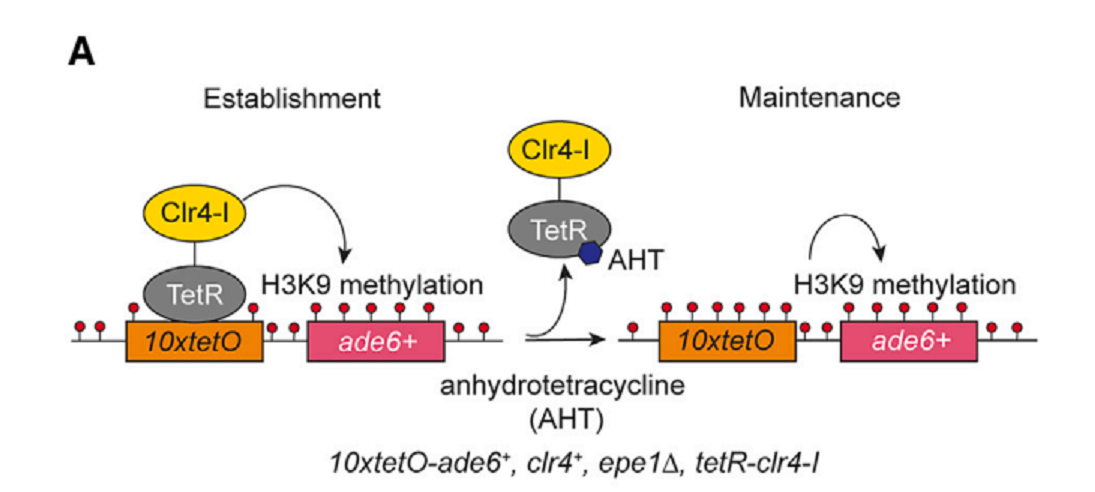
図1.文献18より引用
クェgtHeterochromatin 維持に必要なreplisome components and histone chaperonesの探索
これまでの研究によって、Mac1などのreplisome componentsの変異がgene silencingの欠損をもたらすことが知られていた。上述した実験系を使って、HC構造のestablishmentおよびmaintenanceを区別して調べた結果、Mcm2 (DNA replication licensing factor), Pol1 (DNA polymeraseαcatalytic subunit), Dbp3-Dbp4 (RNA helicase)は構造維持に必要なことが分かった。一方、Rfa3(subunit 3 of replication factor A )やCtf18 (chromosome segregation factor)は、構造維持に必要でない ことが示された。同じようにして、FACT complexを構成する因子のうち、Pob3とFft3 ATPaseはmaintenanceに必要で、Nhp6は必要でないことが示された。
Mrc1 のreplication checkpointとepigenetic inheritanceにおける役割を区別する
Mrc1がcheckpoint signalingのmediatorであることは知られている。Replication proteinsであるSwi1とSwi3はHCの維持に必要でもある。つまり、the replication fork protection complex (FPC)はまるごとHCの維持に必要である(図2)。
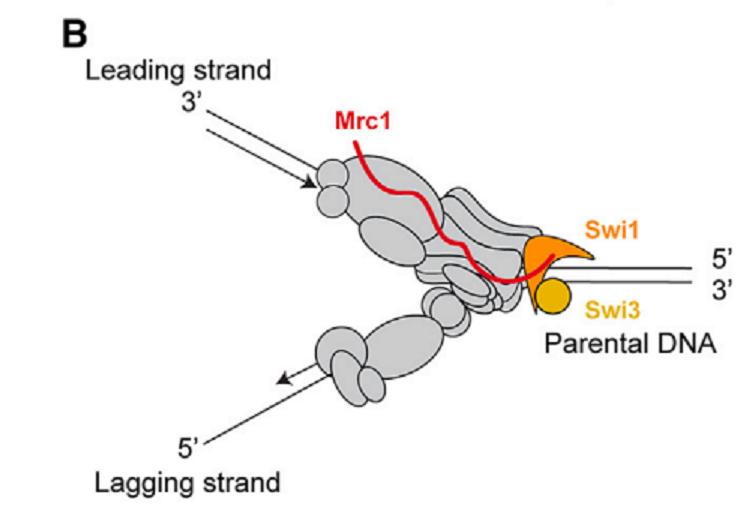
図2.文献18より引用
細胞がreplication stressを受けたとき、Mrc1はstress signalをシグナル下流のcheckpoint effector kinase Cds1に移す(そのserine-glutamine/threonine-glutamine(SQ/TQ)motifsのhyperphosphorylationを通じて)。Cds1のT645, T653とS604がhyperphosphorylation sitesとして同定されている。Mrc1のT or SをAに置換すると、_mrc1_Δと同様にhydroxyureaに感受性となったが、HCの維持は行われた。したがって、replication checkpointの欠損は、HCの維持できないことの原因ではない。
Immunoprecipitation coupled with mass spectrometry (IP-MS)の結果によると、Mrc1はreplisome componentsに結合しているが、replisome componentsはC-terminal phosphodegronを欠損するMrc1 mutant proteinには結合しない。また、Mrc1はreplisome-independentlyにFACTに結合する。その他の結果を総合すると、Mrc1は親鎖のhistonesの移行にかかわっている可能性が浮かび上がる。
Histone H3-H4 tetramer のMrc1/CLASPINへの結合
著者たちは、template-free mode of AlphaFold-Multimerを利用して、Mrc1にhistone-binding regionがあるかどうか検討した。Template-freeとは、既存の構造データベースにある類似の構造を参照しないで複合体の構造予測をする手法のこと。AlphaFoldは構造予測に深層学習モデルをとりいれている構造予測の手法。図3に示したように、Mrc1の701-839に6個のαhelicesがある。予測された構造は図4のように、3つのα1-α3のhelicesはhistone H3.1-H4 tetramerの支柱のように囲み、α4-α6のhelicesは別の位置に想定されたが、5つの予測モデルが同列に出されたように信頼度がやや低い。Mrc1のα1とα3はそれぞれH3-H4 dimersに結合しており、α2はH4 subunitsに結合している。このMrc1の結合によって、H3-H4 tetramerがDNA複製時にも、安定してDNAに結合している。
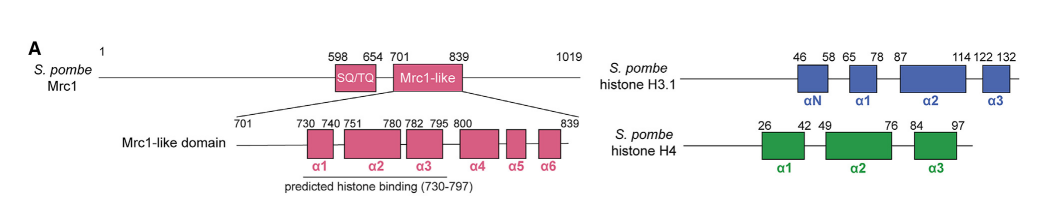
図3.文献18より引用。
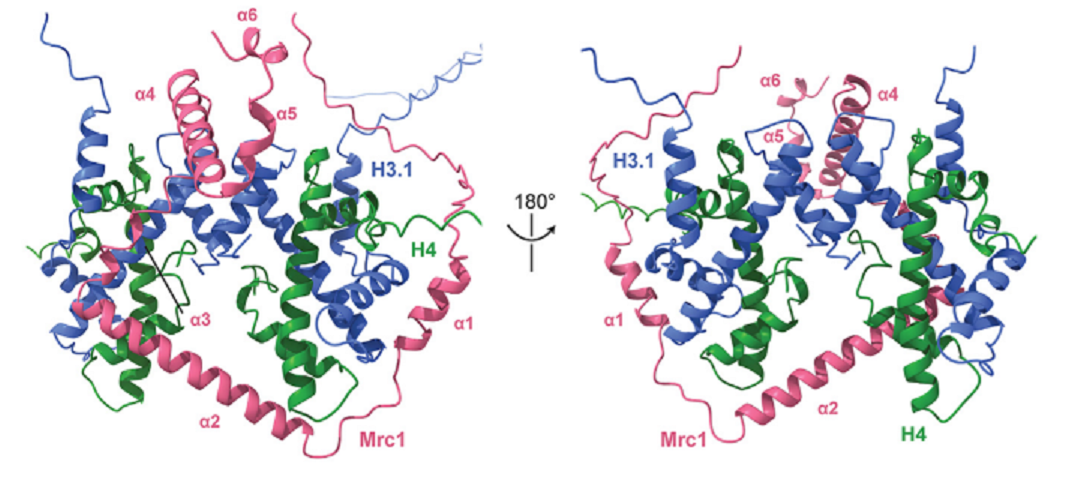
図4.文献18より引用。
AlphaFold で予測したMrc1 histone-binding domainの実験的検証
Glutathione S-transferase (GST)-tagged Mrc1 fragmentsとhistone H3-H4の結合をin vitro pull-down assayで調べた。図5に示したように、Mrc1のaa 651-900 fragmentにH3-H4は結合していた。Mrc1 fragmentsはH2A-H2Bにはほとんど結合しなかった。S. cerevisiae Mrc1およびhuman CLASPIN (checkpoint factor)も、それぞれH3-H4に結合した。
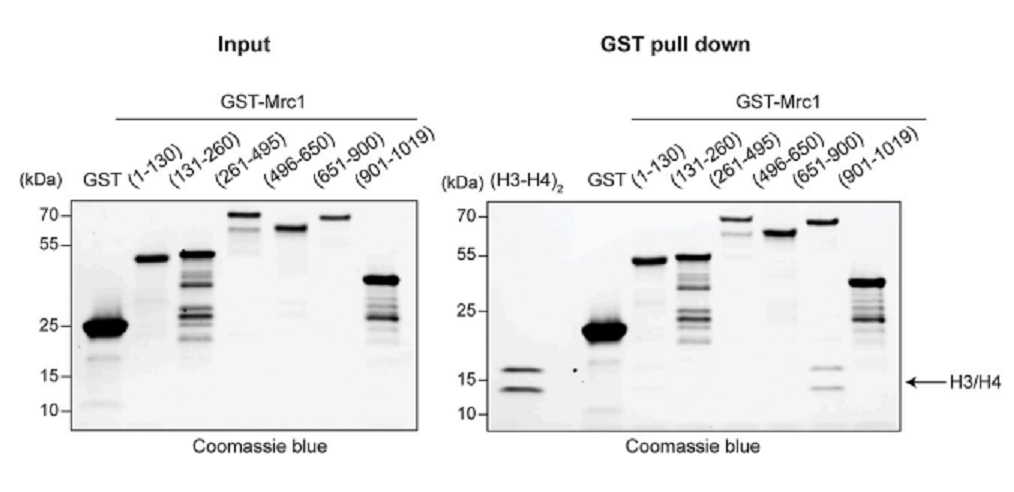
図5.文献18より引用。
GST-tagを外したMrc1 (651-900)と(H3-H4)2は、安定した複合体としてsize exclusion chromatographyで単離できた。このH3-H4 tetramerに結合するMrc1-like domainを以後Mrc1-HBDと称する。AlphaFoldによって予想された構造では、Mrc1-HBDα2 helixの中央にある酸性アミノ酸残基E763とE767が2つのH4にcontactしている(図6)。Mrc1-HBDαhelixの両側にあるhydrophobic amino acids(図6の黄色の部分)は、2つのH4のhydrophobic領域と相互作用している。実際に、Mrc1-HBDのhydrophobic pocketsを構成すると予想されたM755, F758, L774に変異を入れると、GST pull down assayによる結合が見られなくなった。同様に、mrc1-E763R, _D767K_変異を導入すると、H3-H4への結合が失われた。HBDを除いたMrc1を発現している細胞では、HCの維持が認められなかった(replication check-point signalingは正常だった)。このことは、_S. cerevisiae_でも確認できた。
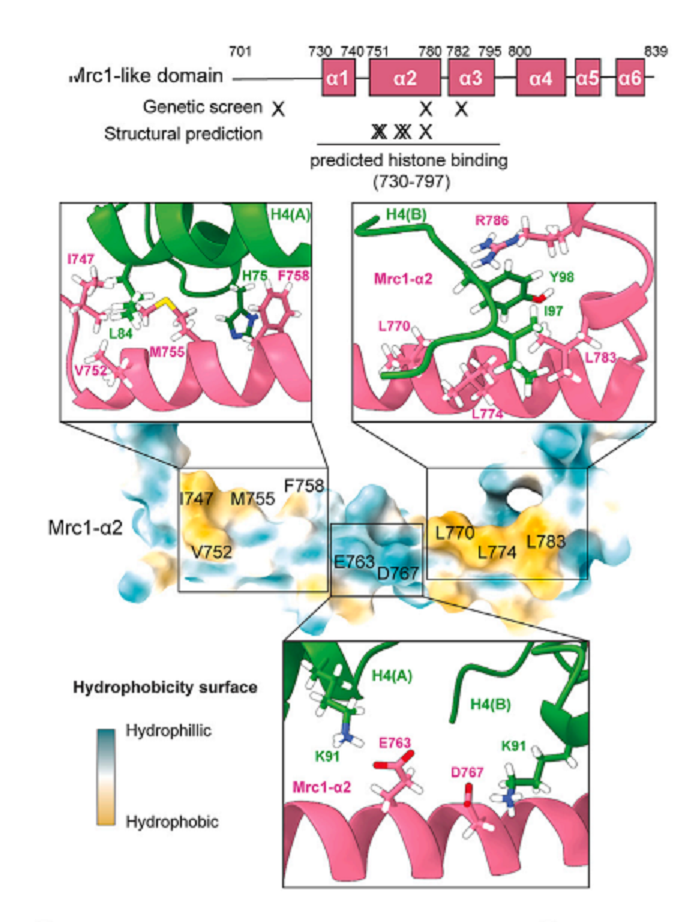
図6.文献18より引用。
DNA 複製中にheterochromatin構造を維持するのにMrc1が必要である
実験は_S. pombe_と_S. cerevisiae_の両方を使った。_Cerevisiae_の細胞をzymolase digestionし、MNase(Ca+-dependent endonucleaseで、proteinsと結合していないDNAを選択的に切断する)で処理し、以後定法で分画した(protein-associated nascent DNA, eSPANと略す)。_Cerevisiae_のwild typeでは、replication origin regionsで、parental histone (H3K4me3をmarkerとして)とnew histone(H3K56acをmarkerとして)が複製した両DNA鎖に均等に配分された。しかし、_mrc1_Δでは、lagging strandへのparental histoneの移行が弱かった。
Mrc1は多くのreplisome componentsに結合し、Mrc1-like domain(図2参照)は直接histoneと相互作用しない部位を有する。そこで、eSPAN実験によって、mrc1 mutantsを使って、replisomeと相互作用に影響を与えず、histone bindingに欠損を示すものを探した。すると、_cerevisiae_では_mrc1_-Δα2、_pombe_で_mrc 3-A_がその性質を示した。他の結果を併せて考慮すると、Mrc1-HBDのmutationsはhistoneの親鎖から子鎖へのsymmetrical transferには影響を与えなかった。
Replisome 上のFACT binding sites
FACT (facilitates chromatin transcription)はchromatin DNAをtemplateにしてin vitro transcriptionを行うのに必要な因子である。Mrc1はreplisome-independentにFACTに結合する。また、Mcm2はFACTと共同してhistoneに結合する。著者たちは、Mrc1と他のhistone-binding proteinsはFACTが結合しているhistonesのco-chaperoneとして働くのではないかとの仮説を立てた。Pairwise AlphaFold-Multimerによって、FACT subunitsとreplisome componentsの相互作用を予想した。IP-MSの結果と同様に、その予想通り、Mrc1にtwo FACT binding domains (FBDs)を示した。さらに、FACT構成成分Spt16とFPC subunitのSwi1のinteraction interfacesが示された。また、Swi1-Mcm2とSwi1-Mrc1 interactionsも示唆された。
以上のことから、Mrc1/CLASPINはFACTや他のreplisome componentsとchaperone networkを構成し、親鎖のhistone H3-H4 tetramersを新たに合成された子鎖に移動させる仕組みの一部が明らかになった(図7にモデルを示した)。ただし、replisomeの前方から、新たに合成された子鎖までは、かなり距離があるので、解明されるべき仕組みが残っている。
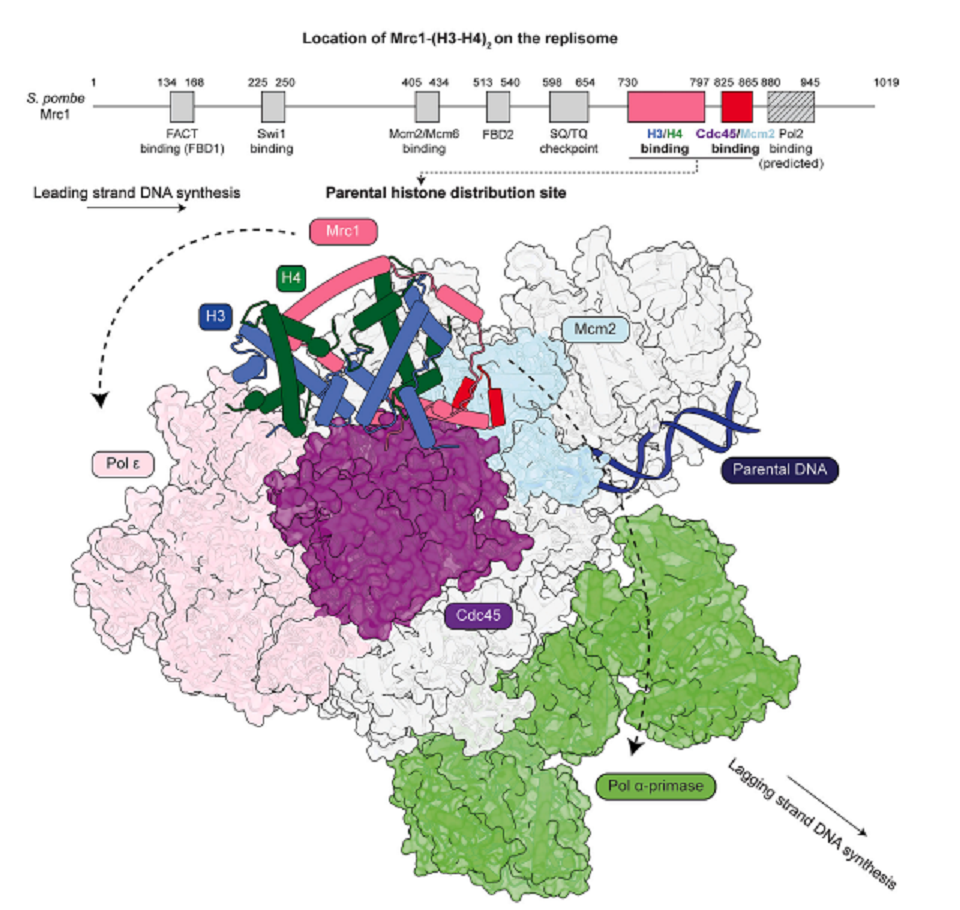
図7.文献18より引用。
2024 年9月17日(火)投稿
料理のこと:ジェノベーゼ・スパゲッティ
わが家では、スパゲッティというと、カルボナーラやボロニアを差し置いて、ジェノベーゼを一番よく食べている。その理由は、この何十年、春から秋まで、バジルを鉢やプランターで育てているからだ。若枝を切って、コップの水に差しておけば、2、3週間程度で根が出てきて、簡単にいくらでも増やせる。
材料は、バジル、ニンニク、松の実、パルミジャーノ・レッジアーノ、オリーブ油と簡単だ。松の実とパルミジャーノがやや高価なのが欠点だが、美味しさには代えられないので、しかたがない。バジルは使う直前に、簡単に水で洗う。先に洗っておくと、すぐに黒くなってしまうので注意。生の松の実(僕は、富沢商店の通販で買っているが、100gで1,000円程度)は乾煎りする(電子レンジでも可)。40gパルミジャーノはすりおろす。60gのバジル葉、ニンニク2かけ、30g松の実をフードプロセッサーに入れ、攪拌する。オリーブ油40gを足してさらに攪拌する。最後に、パルミジャーノを加え、軽く攪拌して味をみて、塩で味を調整する。料理の本では、これを一晩冷蔵庫で寝かせると、味が深くなるとしているが、わが家では、すぐにフレッシュなのを食べる。食べる直前にスパゲッティの上に、チーズを少量すりおろす。スパゲッティは、わが家は老人二人で150g程度なので、ソースが余る。冷蔵庫に貯蔵し、翌朝、トーストにつけると美味しい。
僕は、1995年6月にジェノバに学会で行ったが、ホテルのランチで食べたジェノベーゼには、スパゲッティとジャガイモが入っていた。僕も、メークイーンを入れてみたが、これも美味しい。

写真1.スイート・バジル

2024 年9月15日
第三部 近代世界システム
第二章 産業資本
1.商人資本と産業資本
商品交換様式Cがドミナントであるような社会構成体は、産業資本主義とともにあらわれた。交易や市場の拡大は資本主義経済の必要条件ではあるが、十分条件ではない。例えば、世界市場に向けられた商品生産は、東ヨーロッパでは「再版農奴制」を、また、ラテン・アメリカでは新たな奴隷制をもたらした。商品交換様式Cが交換様式BやAの抵抗を越えるためには、ある変化が生じなければならない。では、それは何か。
産業資本主義はそれまでの資本主義とどこが違うのか。この問題を最初に扱ったのはアダム・スミスである。彼は、商人資本は安く買って高く売る差額から利潤を得るのに対し、産業資本は生産性の向上によって利潤を得ると主張した。ウェーバーは、産業資本主義の根底に、商品資本主義における利益追求や消費欲望を断念し、代わって勤勉な労働のエートスを見ようとした。これに対し、産業資本主義を、根本的に商人資本主義の延長であると、ヴェルナー・ゾンバルトはウェーバーに反対した。彼は、産業資本主義を商人資本主義の延長であるとみなし、資本主義的発展の契機を、禁欲よりも奢侈を求める欲望に見た。消費社会になり、ゾンバルトの考えは高く評価されるようになった。
しかし、以上のような見方は、どれも産業資本主義の一面しか見ていない。柄谷は、産業資本主義を両面(生産と消費)から明らかにしたのは、やはりマルクスであるとする。マルクスはスミスのような立場に立っているとみなされているが、彼が古典派と異なるのは、流通過程に焦点を当てたところにある。マルクスは、資本主義が交換様式Cから生まれたという認識に基づいて考え、産業資本が交換による差額から利益を得る点で、商人資本と変わらないと考えた。本書のP146に書いてあるように、産業資本もM-C-Mの範式の中で利益を得る。しかし、この式の中に不等価交換はない。では、等価交換からどうして利潤が得られるのか。この題は異なる価値体系の間の商品交換を想定することで解決できる。商人がある物をやすい場所で買って高いところで売れば、それぞれ等価交換でありながら、剰余価値を得られる。スミスはピンの製造を例にとって、商人資本が生産に関与する場合を論じた。一方、産業資本も生産過程における技術革新からだけでなく、より安い原料と労働力、さらに消費者を求めて「遠隔地」に赴く。それゆえ、商人資本と産業資本の差異は、単に流通過程や生産過程を見るだけでは明記できない。
2.労働力商品
マルクスは流通過程を重視した重商主義者と生産過程を重視した古典派のどちらをも批判した。彼は、産業資本における剰余価値が、単なる流通過程でもなく、単なる生産過程でもないようなところで得られると考えた。つまり、Mで商品Cを買い、売るときにM+ΔMとして剰余を出すが、ΔMを生む商品は使用することが生産過程であるような商品、つまり労働力なのである。産業資本の価値増殖過程は、M-C‥P‥C‘-M’という公式で示される。
マルクスは産業プロレタリアを「二重の意味で自由な」人々として見た。第一に、かれらは、さまざまな封建的拘束にとらわれず、自由に労働力を売ることができる。第二に、かれらは労働力以外に売るものをもっていない。つまり、生産手段(土地)から自由である。この二種類の自由は不可分離である。第一の「自由」は、労働力の売買についての契約(雇用契約)にのみ縛られるが、それ以外の自由である。第二の「自由」は、普通の意味での「自由」ではない。プロレタリアは生産手段には一切かかわりがない(その意味で自由)ので、生産物である生活物質(商品)を、労働力を売ったお金で、買わなければならない。小作農民が生産物の大部分を献納しても、一部を自給自足に回すことができるのと、大違いである。産業資本主義経済において、労働者の消費は、労働力を再生産するのであるから、資本の蓄積過程の一環でもある。労働者が自ら作った商品を買い、そこに生じる差額(剰余価値)によって、産業資本は増大する。
柄谷は書いている。プロレタリアという語には、どうしても貧窮者というイメージがつきまとう。今日、ホワイト・カラーと名付けられる人たちは、紛れもなく賃労働者であるにもかかわらず、自身をプロレタリアと考えない。プロレタリアは貧しい肉体労働者だという固定観念があるからだ。したがって、柄谷は、本書でプロレタリアという語の使用を控え、賃労働者と呼ぶことにする。
3.産業資本の自己増殖
資本は資本一般あるいは総資本として見る場合と、個別の資本として見る場合があり、この区別は重要である。本書には、産業資本の特性は、労働者が資本の下で自らが作ったものを買い戻すシステムにある、と書いた。しかし、これは総資本と総労働についてしか該当しない。個別の労働者は、自分の作ったものより、他の労働者が作った物を多く買う。
マルクスからの引用(略記):どの資本家も自分の労働者の賃金を制限したい、つまり消費能力を少なくしたいと思っている。一方、他の資本家の労働者が自分の商品の大きな消費者であることを望んでいる。ここでは、労働者が消費者および交換価値措定者として資本に相対するので、労働者としての規定性が消し去られている。
総資本と個別資本の違いは明白である。危機の際しては、個々の資本家の意思に反して、総資本があらわれる。それも「国家」というかたちであらわれる。例えば、1930年代の大不況において、国家=総資本は、ケインズ主義(公共投資)あるいはフォーディズム(フォード主義:生産性の向上と労働者の賃金と購買力の上昇)のような、個別の資本がとれない政策をとった。しかし、この違いは、資本主義の本質、つまり資本の自己増殖、あるいは剰余価値の実現については変わらない。総資本は総労働に対し等価交換を行い、それでもなお剰余価値を生むのである。
スミスやリカードは、個々の労働者は、資本家が組織した分業と協業を通じてなしとげた生産の全成果を要求することはできないとした。その増加分(利潤)はそれを考案し組織した資本家が受け取るべきと考えた。しかし、リカード派の社会主義者やプルードンは、その増加分が「剰余価値」であり、本来労働者に帰属すべきものであると主張した。つまり、搾取が行われているというわけだ。マルクスもこのような見方を受け継いでいる。マルクスは労働時間の延長や労働強化によって得られる剰余価値(註:これはもはや等価交換ではない)とよび、技術開発=生産性の向上によってもたらされる剰余価値を「相対的剰余価値」と呼んだ。資本論の「絶対的剰余価値」に関する記述はよく知られているが、むしろ大事なのは「相対的剰余価値」である。ここにこそ、産業資本の精髄があるからである。しかし、生産性向上で増えた生産物はだれが消費するのか。生産した労働者の賃金では買えない。つまり閉じた生産・価値体系の中だけでは、成り立たない。そこで、外国の市場を拡大するか、共同体の中に新たな消費者=労働者(プロレタリア)を参入させなくてはならない。資本が蓄積を続けるためには、絶えず新たなプロレタリアが必要である。
今回は、ここまでにしておく。産業資本主義の原理を扱ったおり、本書の記述だけでは、不十分な印象を受けた(僕にとっては)。以後も産業資本主義についての記述が続いているので、期待したい。
次項は、4.産業資本主義の起源、についてである。
2024 年9月10日 投稿
Ghalambor, C. K., et al., Non-adaptive plasticity potentiates rapid adaptive evolution of gene expression. Nature 525: 372 (2015), (文献19)の紹介
同じgenotypeを有する個体群が、異なる環境要因によって、表現型(発生過程の変化によるもの、あるいは個体の行動や学習によるもの、など)が違ってくることを、 plasticity(可塑性)と言う。しかし、この表現型のplasticityと表現型の進化の関係は、不明のままである。表現型のplasticityはnon-heritable variationなので、この問題は、正当には扱われてこなかった。
特定の地域や集団だけに限定してみても、ある生物が環境変化に出会ったとき、様々なphenotypesの変化を示す。その変化が、the local phenotypic optimum(局所的に、一定の集団が最適な生育をする表現型)に向かっていればadaptiveといい、逆方向になっていればnon-adaptiveという。別の言い方をすれば、plasticityが自然選択に有利な方向であれば、adaptiveとする。しかし、adaptive plasticityはadaptive evolutionに向けての自然選択の力を弱める、とする考えもある。逆に、前回取り上げた論文(文献18)やWaddingtonやLindquistたちの研究でみてきたgenetic assimilationは、adaptive plasticityの集積と言える。これまでの、phenotypic plasticityの研究は実験室内で行われてきた。この論文の著者たちは、自然の環境の中でどうなっているかを、調べてみた。
High-predator(HP)強捕食者(例えば、pike cichlid)の脅威にさらされながら生育しているTrinidadian guppiesを研究対象に選んだ。Trinidad島の図1のような地形の川を実験場所とした。下側のHigh Predationは捕食者のcichlidとguppyが一緒に棲息している地域、上流のLow Predation (LP)はcichlidが棲息していない地域。HPに住んでいたguppiesをcichlidの棲息しない地域に移したら、どう変化が生じるかを調べた。それぞれ、38匹の妊娠中の♀と38匹の♂を、上流の二か所LPに放流した(Intro 1 and Intro 2)。1年後(3-4 guppy generations)、放流した魚の子孫を回収した。Low Predation地域から天然のLP魚を集めた。これは、ずっと昔に、HPからLPに移って進化したphenotypeを有すると考えられる。

図1.論文19より引用。
もう一度述べると、下記の4つの集団を使った。
1.Cichlidの棲んでいた地域からcichlicのいない地域に移した集団の1(Intro 1)
2.同上の2(Intro 2)
3.Cichlidの棲んでいる地域の集団(HP)
4.Cichlidの棲んでいない地域の集団(LP)
それぞれの地域の川から採取したこれらのguppiesを実験室内(同一条件)で2世代飼育した。それぞれのグループの生後24hのsiblings個体を二分し、一方をpredator cuesとなる化学物質に曝露させ、もう一方を曝露なしとした。Predator cuesはcichlidのpredator kairomonesとguppiesの警戒ホルモンを含んでいる。
4つのグループの魚の脳について、37,493のmRNAsを調べた(high-throughput RNA sequencingによって)。結果を、Multivariate Between-Group Principal Components Analysis (MBPCA)(グループ間の差異を解析する多変量解析手法)を利用して解析した(図2)。図の2つの軸が全体の変動の74.5%を説明できた。PC1(主成分1:全体の変動の44.4%)によって、上記4のグループを1~3のグループから分けることができた。このことは、LPが他のグループから長時間かけて分化したことを反映している。PC2(主成分2:全体の変動の30.1%)は、HPを他の3グループから区別しているので、LP環境への急速な進化的分化それも適応的な変化をとらえていると言える。

文献19より引用。
グループ1と2に共通に認められ、さらにグループ4にも認められるtranscriptsを、general linear statistical modelsを適用し、データの確実性を検証した。その結果、135 transcripts(concordantly differentially expressed, CDE)がこの条件を満たしていることを見出した。つまり、predatorがいない地域に適合している状態で発現しているtranscriptsである(因果関係は不明)。
そこで、グループ3(HP)の魚をpredatorのいない領域に移したとき、CDEのtranscriptsがどう変化するか(グループ3の魚のplasticity)を調べた。著者たちは、CDE transcriptsが、predatorのいない地域に適合している(進化して?)変化とは、逆の方向に変化していたとの結論を得た(図3)。図2では、135 CDE transcriptsが、グループ3の魚をpredator cuesのない状態に移したときに変化した量を横軸に、グループ1と2において発現している量を縦軸に、プロットした。明らかに、HPの魚のplasticity(量的変化)の方向とLPに適合して進化した魚のtranscriptsの量変化は逆の方向を示している(135 CDEの内120、89%がその傾向を示した)。つまり、HPの魚はpredator cuesのない状態に対し、非適応的なplasticityを示した。論文では、具体的データとしてuridine phosphorylase 2の場合を、参考図として示している(図4)。

図3.文献19より引用。

図4.文献19より引用。
以上の結果は、plasticityは急速な適応的進化する力を付与する、ことを示唆している。それは、plasticityがadaptiveだからではなく、むしろ逆にnon-adaptive(強い選択圧が働くときに)だから、と考える。もし、自然選択がnon-adaptive plasticityを示したtranscriptsに働くとすれば、グループ1と2の魚のpredator cuesに対するplasticityは減少しているはずである。この予見は、実験的に裏付けられた。この論文に書いた結果は、non-adaptive plasticityが新しい環境に対するadaptive evolutionを獲得する上で、重要な役割を担っていることを示唆している。
240905 投稿
Evolution of a Polyphenism by Genetic Accomodation (文献18)
同じ遺伝子セット(genome)を持つ生物個体が、環境の違いによって異なる表現型を示す事象をpolyphenismsという。要するに、epigeneticsの問題である。例えば、ハチやアリのような社会性生物のカースト制、サバクトビバッタの孤立相と群生相、アリマキの有翅型と無翅型、あるいはチョウの春型と夏型、などなどいくらでもある。このような、表現型が転換する刺激に関心をもち、多くの研究がなされてきた。本研究もその一環であるが、実験系が単純なのと、polyphenismsの進化につながる糸口が見えているので、面白い。このブログで論じてきたHsp90関係の研究との接点もはっきりしている。
Manduca sexta(タバコスズメガ)の幼虫の体色は緑である。近縁のM. quinquemaculataの体色は20℃で黒であるが、28℃では緑である。M. sexta(5齢後期幼虫)の体色は安定で、熱ショック42℃、6h処理によっても変化しない。M. sextaには幼虫の体色が黒い変異体がある。その変異はsex-linkedで劣性であり、juvenile hormone(JH)のレベルが低い。このホルモンレベル低下が幼虫表皮のメラニンを増やしている。外部からJHを投与することで、体色黒化を防ぐことができる。
変異体の4齢幼虫は、20℃~28℃で黒色である。4齢幼虫を42℃、6hの熱ショック処理をすると、脱皮した5齢幼虫の体色が変化し、元の黒色から緑色まで様々な体色を示した(図1)。熱ショックによって誘導された多様な表現型を示す幼虫から、2つの系統を作成した。一つは、熱ショック処理によって緑化した幼虫のスズメガの系統A(polyphenic line)、もう一つは、体色変化しなかった黒色幼虫のスズメガの系統B(monophenic line)である。

図1.
それぞれ300の幼虫を使い、世代ごとに熱ショック処理し、60匹の目的の表現型を示す幼虫を選択し、次の世代を作成した。
コントロール実験として、選択しないで飼育した幼虫の系統も作成した。
選択実験を繰り返すと、熱ショックによって誘導される幼虫の体色のスコアは、図2のようになった。すなわち、緑化スコアの大きいlineは世代を重ねると、ますますスコアが増え、黒色のラインはますますスコアが低くなった。つまり、系統Aでは熱ショックに対する応答様式は次第に強くなり次世代に伝達された。系統Bの黒色のラインは、6世代後にほぼ完全に熱ショック応答性(緑化するという)を失った。
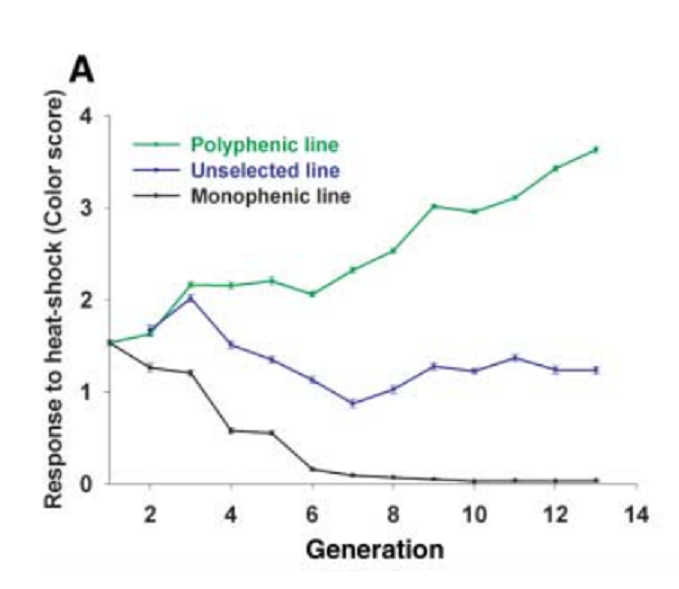
図2.文献18より引用
体色変化のHeat-shockに対する感受する時間とJH-sensitiveの時間は、同じである。黒色変異体にJHを局所投与すると、黒色の表現型を緑色の表現型に変えることができる。Dopa decarboxylase (DDC)はmelanine合成において、dopaをdopamineに変換する酵素であるが、JH-snsitive時間の16時間後に合成される。したがって、熱ショック処理を、melanine合成のこの時間より前に行えば効果を発揮する。
JHあるいはecdysteroidsはmelanine合成の調節にかかわるという報告がある。これらのホルモンはthoraxのprothoracic glands、あるいは頭部の脳とcorpora allata(アラタ体)から分泌される。どちらのホルモンか緑化にかかわるかを調べた。Polyphenic linesの幼虫の腹部体節で結紮を行うと、heat shockを与えた時、結紮より前方の体色は緑になるが、後方は黒色となった(図3A、3B)。Monophenic linesの幼虫を腹部結紮した場合、当然ながら緑化は起こらなかった(図3C)。Polyphenic linesの幼虫の頚部を結紮した場合、heat shockを与えても、緑化はまったく起こらなかった(図3Aの右下)。これらの結果は、脳とアラタ体からの信号が体色の緑化に必要なことを示している。JH analog, methopreneを結紮より後部に塗布すると、当然のことながら、polyphenic linesでもmonophenic linesでも緑色化が起こった(緑色化が起こらなかったのは、JHに対するsensitivityの欠損が原因ではない)。
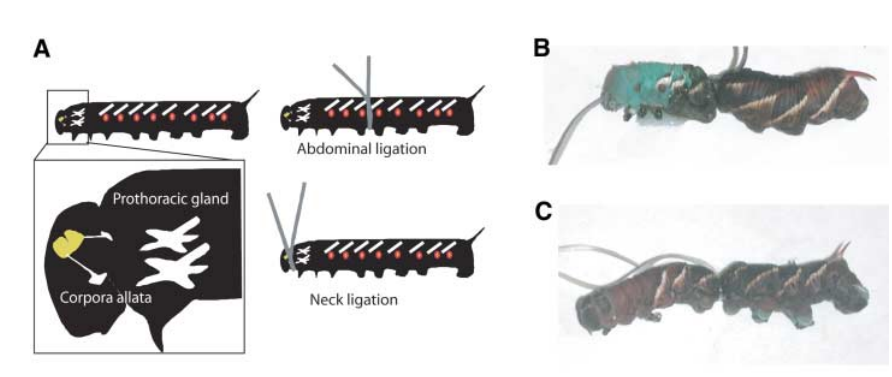
図3.文献18より引用
本研究では、JH合成の調節系の欠損突然変異(monophenic黒化型)を有する個体が、高温処理によって次第に緑化する能力(heat shockを必要とするが)を獲得してゆくプロセスを示した。選択と継代によって、能力が高まるので、いわゆるgenetic accommodationの例である。最終的にはheat shockが不要になる、genetic assimilationとは違う、と著者は強調している。概要を、図4に示した。上から、wild type, black mutant, black mutant由来のpolyphenc line, monophonic lineの結果である。2つのthresholds, T1, T2で、T1より下では常に黒色で、T2を超えると緑化する。T1とT2の間では、intermediate phenotypeを示す。図の山(個体の数)は、low temperature, high temperature, heat-shockによって変化する。
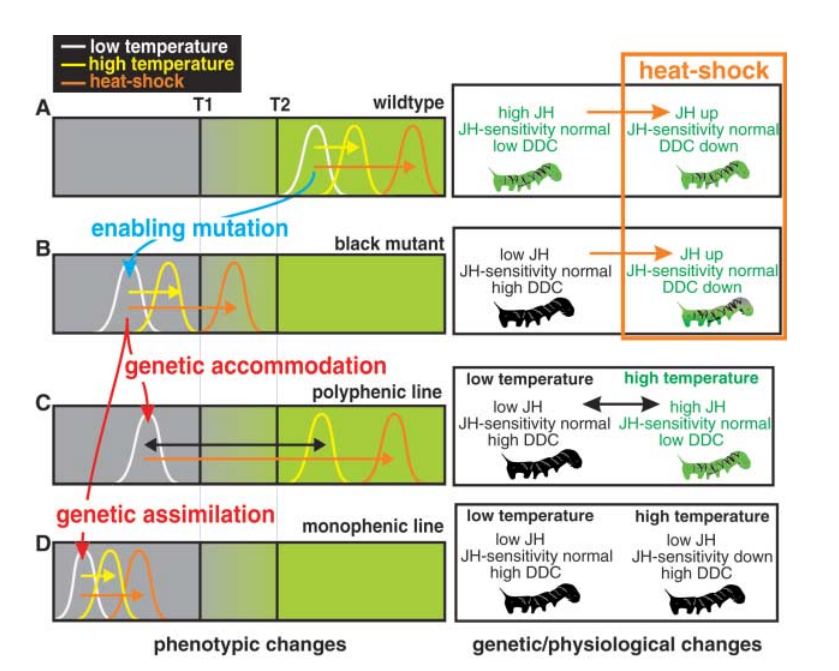
図4.文献18より引用
ブログの著者による意見:すでにこのブログで取り上げたが、epigeneticな変化は世代を超えて蓄積してゆくことが明瞭に示されているので、この論文が示したことは、新しくはない。選択によって、一方向への表現型の変化がadaptiveに蓄積した例として、発表当時は新規性が評価された。
- Suzuki, Y. & Nijhout, H. F., Science 311: 650 (2006)
2024 年8月29日
第三部 近代世界システム
第一章 近代国家
1.絶対主義王権
世界=帝国では商業や交易が発展したが、それは国家によって独占的に管理されたもので、そこでは商品交換の原理は他の交換様式を上回ることはできない(註:いまの中国は?)。世界=経済、すなわち、商品交換の原理が他の交換様式に優越する事態は、国家が一元的な集積性を持つことがない地域、つまり西ヨーロッパにだけ起こった。西ヨーロッパにおいて集権的な国家が始まるのは、絶対主義的王権国家(以後、絶対王権とよぶ)によってである。王は周辺の封建諸侯を制圧し、教会の支配権を奪った。これが可能だったのは、一つには破壊力を有する火器の発明である。火器は旧来の戦力を無効にし、貴族=戦士の身分を無効にした。もう一つの要因は貨幣経済の浸透だった。14世紀のイギリスでは、封建領主は農民から封建的貢納を受け取る代わりに、地代を受け取る地主階級となっていた。王は都市の商工業者と結託して、封建諸侯の諸特権を廃止し、地祖を独占した。権力を奪われた封建諸侯は、国家から租税を分配される宮廷貴族・地方の地主階級となった。こうして、貨幣経済は、絶対王権を支える官僚や常備軍をもたらした。絶対王権国家はアジア的な専制国家(世界=帝国)に似ているが、後者が、交換様式Bが支配的な社会構成体であるのに対し、前者は交換様式Cが支配的であるところである。そのため、アジア的な専制国家が崩壊してもまもなく再建されるのに対し、絶対王政が崩壊するときは、ブルジョア社会になってしまう。
絶対王権国家は世界=帝国の存在しない西ヨーロッパに生まれた。教会、王、封建領主、都市などが拮抗し、かつ相互に依存する状態から、絶対王権国家が出現した。絶対王権は二つの意味で「絶対的」である。まず、一定の地域において、他の封建諸侯を超越した地位に立った。次に、王権の絶対性は、上位の組織や観念(教会や皇帝)を斥けた。その意味で、一定の地域外の王権国家と共存する。こうして、西ヨーロッパに、かってないタイプの集権的な(複数の)国家が出現したのである。主権国家という観念は、主権国家として認められない国ならば、支配されてもしかたがない、ということを含意する。ヨーロッパの植民地支配を支えたのはこの考えである。
西洋列強(複数の主権国家)は、オスマン、清朝、ムガールといった巨大な世界帝国には手をだせないので、それらの帝国の統治形態を非難し、従属している諸民族を解放し主権を与えるかのようにふるまった。その結果、旧世界帝国は解体され、多数の民族国家に分解され、それぞれが主権国家として独立した。つまり、主権国家は西ヨーロッパに始まったとしても、グローバルに主権国家を生み出した。
2.国家と政府
主権国家は、内部での集権化によって生じるとはいえ、本来、外部に対して存在するものである。しかし、絶対王政を倒した市民革命後には、そのような事実が見落とされるようになる。ジョン・ロックは、国家を主権者である市民らの社会契約としてとらえた。国家はその内部だけで考えられている。したがって、国家は主権者である人々の代表である政府に還元されてしまう。国家が何よりも他の国家に対して存在することが見失われる。市民革命に基づく今日のイデオロギーは、国家が何たるかを見逃すことになる。
イギリスのピューリタン革命によって、絶対王政が倒され(1648年)、議会派だったクロムウェルが独裁体制をしき、さらにそれが倒されて王政復古(1666年)となった。この後、名誉革命があり、立憲君主制が確立された(1689年)。この激動の時代に関して、ロックとトマス・ホッブスの考えが述べられているが、省略する。柄谷は、主権という考えは国内だけで考えられるものではなく、主権はまず外に対して存在するものだ、と断定している。したがって、絶対王政が倒されても、他の国家に対す主権の性格はなにも変わらない。
3.国家と資本
まず、国民主権と国家の主権の違いについて述べてある。国家の本質は、なによりも他の国家に対し存在するところにある。市民革命以後の社会契約論では、国家の意思とは国民の意思であり、それは選挙を通して政府によって代行されると考えられている。ところが、国家は政府とは別のものであり、国民の意思から独立した意思をもっている。このことは、戦争のような例外状況において露出する。このことは、絶対王政や近代以前の国家においては、明瞭に可視的であった。これが国民国家以後に見えなくなった。国家の意思は、国家機構である常備軍と官僚によって実行される。国民主権の下であろうと、国家はそれ自身のために(国民のために、ではなく)存続しようとするのである。もう一つ、国民国家によって曖昧にされたが、絶対王政において明瞭にことは、資本=国家、すなわち資本と国家の結合である。市民革命以後のブルジョア国家においては、国家は市民社会の階級的利害が政治的に表現される場とみなされる。国家それ自体、能動的な主体であるとは考えられない。だが、絶対王政にこそ、国家の自律性あるいは資本=国家の本質がみえる。
資本と国家の結合は、次の二点において明瞭である。一つは、国債の発行である。絶対主義王権は、この「魔法の杖」(マルクス)によって、いつでも税収入を先取りすることができた。もう一つは、保護主義的政策である。イギリスの産業資本の発展も国家の保護によって可能であったが、イギリスに後れをとった後発資本主義国で、国家による産業資本主義化がとられるのは当然のことである。国家は資本主義経済において、単なる上部構造ではなく、不可欠な要素なのである。
国家は産業資本主義に必要なインフラを整備するが、国家が行うことの中で、より重要なのは、産業プロレタリアートの育成である。これは、規律をもち、勤勉で、新たな多様な仕事にすばやく適応できる能力をもつ人々である。彼らは、農民のように自給自足することなく、賃労働で得た金で、生産物を買う消費者である。資本はこのような産業プロレタリア(労働力商品)を生産することができない。それを行うのが国家なのだ。具体的にいえば、学校教育であり、徴兵制による軍隊である(徴兵制は軍事力よりも、産業プロレタリア養成に貢献する)。
市民革命以後、官僚は、議会を通して表現・決定された国民の意思を実行する「公僕」であると考えられるようになった。しかし、実情がそうでないことは誰でも知っている。ヘーゲルによれば、議会は、人々の意見によって国家の政策を決めていく場ではなく、官吏たちによる判断を人々に知らせ、まるで彼ら自身が決めたことであるかのように思わせる場なのである。
20世紀において、国家がケインズ主義的な経済介入をとるようになったことが注目される。しかし、国家が経済に介入しなかった時期は一つもないというべきである。例えば、19世紀に経済的な自由主義とよばれたものは、イギリス国家の「経済政策」であって、それはその体制を守るための巨大な軍事予算と課税にもとづいていた。後発資本主義国家(フランス、ドイツ、日本など)において、国家の経済への介入は自明のことであった。国家が資本主義経済を発展させたのである。それを担ったのが官僚機構である。
国家をブルジョア階級の支配のための暴力装置としてみる一般的なマルクス主義者に対し、アントニオ・グラムシは、暴力的な権力と、被支配者が自発的に服従するようにさせるヘゲモニーを区別した。後者の装置は、家族、学校、教会、メディアなどである。しかし、この見方も、国家をその内部だけで見る点では、国家が他の国家に対して存在するという位相が抜けている点で、同等である。国家の自立性は、それが他の国家に対して存在するという位相においてのみ見出されるのである。
4.マルクスの国家論
社会主義革命は旧来の国家機構を廃棄しようとした。しかし、それはただちに外からの干渉を招くので、革命の防衛のために旧来の軍・官僚機構に依存せざるをえない。かくして、旧来の国家機構が保存され再強化される。たとえば、国家の側から見れば、ロシア革命は、旧ロシア帝国が国民国家に分解することを阻止して、新たな世界=帝国を再建することに貢献した、ことになる。
マルクスは資本主義について深い考察を行ったが、国家については不十分だった。「資本論」に国家が事実上抜けていたことは、マルクス主義者に影響した。しかし、「ルイ・ボナパルトのブリューメル18日」では、国家機構(官僚装置)が一つの階級として存在することを見落としてはいない。マルクスは、資本、賃労働、地代というカテゴリーに入らない諸階級、特に小農(分割地農民)の果たした役割を見落とさなかった。彼は、「ブリューメル18日」という夢のような二度の事件を解明する鍵を、1848年の革命がもたらした普通選挙による議会に見出した。マルクスによれば、普通選挙には、代表する者(言説)と代表される者(経済的諸階級)との間に、必然的なつながりはない。ここにこそ、近代国家の議会の特質がある。だからこそ、諸階級が自分たちの本来の代表に背を向け、ボナパルトにかれらの代表を見出したのである。このとき、50万の軍隊と50万の官僚制組織がフランス社会にからみついていた(これは絶対王政の時代に育ったものだが)。さらに、1851年の周期的世界恐慌が重ねて作用したことを、マルクスは指摘している。この例外的状況において、普通選挙の下に、あるいは市場経済の下に隠れていた官僚機構が、言い換えれば「国家」そのものが前景に登場したのである。この国家機構の自立は、ボナパルトが皇帝として議会を越えたときに成立した。マルクスは、ルイ・ボナパルトがあらゆる階級に対し気前よく「贈与」することによって権威を得てゆく過程を描いている。しかし、「贈与」するには、すべての者から「略取」しなければならない。つまり、国家機構による略取―再分配に、贈与―返礼という互酬交換の外見を与えることで、皇帝権力が確立された。
第一次フランス革命(1789年~)の後、権力を握ったのが皇帝ナポレオンであったが、1848年にも繰り返された(二月革命)。この直前に、マルクスとエンゲルスは『共産党宣言』を発表した。しかし、世界が資本家とプロレタリアの二大階級の決戦になるだろうとの予見は、まったくはずれた。マルクスは、経済的な階級的対立が揚棄されれば上部構造である国家は自然に消滅するだろう、との観方をとりつづけた。柄谷は書いている:そして、そのことが後に、社会主義にとって致命的な結果をもたらすことになったと。
5.近代官僚制
絶対主義国家においては、軍と官僚という国家機構が主権者である王の意思を履行していた。ところが、ブルジョア革命以後、国家は、主権者である国民の意思を代行する政府と同じこととなった。しかし、国民主権は虚構にすぎない。事実、危機的な状態においては、絶対主義的な王に似た強力な指導者が、国民の喝采とともに出現する。
ウェーバーは、官僚制を国家による国民の「合法的支配」の一形態とみなした。具体的には、規則による権限の明確化、官職階層制、自由な契約による任命、規律による昇進、専門的訓練、貨幣で支払われる俸給、といったものである。近代においては、官僚制が国家機構だけではなく、私企業においても存在する。というよりも、近代官僚制は資本主義的な経営形態(分業と協業)に基づいて形成されたから、当然のことである。
ライト・ミルズが分析したように、ホワイト・カラーは、貨幣と商品という経済的カテゴリーに基づく階級(class)で言えば、プロレタリアなのだが、実際はブルー・カラーを支配する身分(status)にある。ホワイト・カラーは組織の歯車として働き、位階を上がるためにあくせくしなければならない。これは賃労働制というよりは、官僚制そのものである。労働者階級は、経営陣、正社員、パートタイマーというような位階制の下に分断されている。ここでは、従来の階級闘争は通用しない。この問題は後に述べる(第四部第二章)。
ネオリベラリスト(リバタリアン)は、国家の官僚機構(警察や軍隊を含めて議論する場合もある)を民営化すべきであり、それによって能率が上がり、官僚は縮小されるだろうと主張する。しかし、柄谷は、官僚化によって能率が上がる、というのは欺瞞である、と言う。なぜなら、私企業そのものが既に官僚制的だからだ。また、利潤という計算可能な目的をもたない、あるいは持ちえない領域にかかわる公的官僚に目的合理性(利潤の最大化)を強制することはできない(註:外交、司法など)。商品交換様式Cがどんなに広がっても、国家やネーションが自動的に解消されることはない。なぜなら、それらは商品交換とは別の交換様式に根差しているからである。また、商品交換も国家やネーションを不可欠なものとするからである。
今回はここまでにする。以下、第三部 近代世界システム、第二章 産業資本と続いている。