PGP expression in Cooperia oncophora before and after ivermectin selection (original) (raw)
Introduction
Infections with parasitic gastrointestinal nematodes (GIN) are common among grazing cattle worldwide, and may cause serious economic losses due to decreased growth of their hosts (Sutherland and Leathwick [2011](/article/10.1007/s00436-013-3473-5#ref-CR41 "Sutherland IA, Leathwick DM (2011) Anthelmintic resistance in nematode parasites of cattle: a global issue? Trends Parasitol 27(4):176–181. doi: 10.1016/j.pt.2010.11.008
")). In Sweden, the most important GIN species include _Cooperia oncophora_ (Fig. [1](/article/10.1007/s00436-013-3473-5#Fig1)) and the more pathogenic _Ostertagia ostertagi,_ which usually are present as mixed infections in grazing cattle (Höglund [2010](/article/10.1007/s00436-013-3473-5#ref-CR15 "Höglund J (2010) Parasite surveillance and novel use of anthelmintics in cattle. Acta Vet Scand 52:2–5")). Regular strategic treatment with anthelmintic drugs, as topical or injectable formulations, remains the principal means of control of helminth infections (Prichard et al. [2007](/article/10.1007/s00436-013-3473-5#ref-CR38 "Prichard RK, von Samson-Himmelstjerna G, Blackhall WJ, Geary TG (2007) Foreword: towards markers for anthelmintic resistance in helminths of importance in animal and human health. Parasitology 134(Pt 8):1073–1076. doi:
10.1017/S0031182007000078
")). Lately, reports have shown that a worldwide spread of anthelmintic resistance (AR) has occurred, probably due to an extensive usage of anthelmintics in the cattle livestock industry (Demeler et al. [2009](/article/10.1007/s00436-013-3473-5#ref-CR8 "Demeler J et al (2009) Monitoring the efficacy of ivermectin and albendazole against gastro intestinal nematodes of cattle in Northern Europe. Vet Parasitol 160(1–2):109–115. doi:
10.1016/j.vetpar.2008.10.030
"); Gasbarre et al. [2009](/article/10.1007/s00436-013-3473-5#ref-CR13 "Gasbarre LC, Smith LL, Lichtenfels JR, Pilitt PA (2009) The identification of cattle nematode parasites resistant to multiple classes of anthelmintics in a commercial cattle population in the US. Vet Parasitol 166(3–4):281–285. doi:
10.1016/j.vetpar.2009.08.018
"); Sutherland and Leathwick [2011](/article/10.1007/s00436-013-3473-5#ref-CR41 "Sutherland IA, Leathwick DM (2011) Anthelmintic resistance in nematode parasites of cattle: a global issue? Trends Parasitol 27(4):176–181. doi:
10.1016/j.pt.2010.11.008
")). However, the underlying mechanisms of AR development in cattle nematodes remain essentially unknown. It is understood that drug resistance can arise in four different ways: a change in the molecular target causing failure at the binding site, changes in drug metabolism preventing activation or removing the drug, changes in drug distribution in the target organism, or amplification of target genes that overcome drug action (Wolstenholme et al. [2004](/article/10.1007/s00436-013-3473-5#ref-CR46 "Wolstenholme AJ, Fairweather I, Prichard R, von Samson-Himmelstjerna G, Sangster NC (2004) Drug resistance in veterinary helminths. Trends Parasitol 20(10):469–476. doi:
10.1016/j.pt.2004.07.010
")).Fig. 1
C. oncophora, bursa and spicules of adult male
Ivermectin (IVM) is one of several anthelmintics in the group of macrocyclic lactones (ML), which has been extensively used in the control of parasitic nematodes of livestock since they were introduced in the early 1980s (Omura [2008](/article/10.1007/s00436-013-3473-5#ref-CR34 "Omura S (2008) Ivermectin: 25 years and still going strong. Int J Antimicrob Agents 31(2):91–98. doi: 10.1016/j.ijantimicag.2007.08.023
")). MLs act through irreversible binding to γ-aminobutyric acid- and glutamate-gated chloride (GluCl) channels, causing hyperpolarisation and thus paralysis of pharyngeal and somatic muscle cells, leading to starvation and expulsion of worms from the GI tract (Blackhall et al. [1998b](/article/10.1007/s00436-013-3473-5#ref-CR4 "Blackhall WJ, Pouliot JF, Prichard RK, Beech RN (1998b) Haemonchus contortus: selection at a glutamate-gated chloride channel gene in ivermectin- and moxidectin-selected strains. Exp Parasitol 90(1):42–48. doi:
10.1006/expr.1998.4316
"), [2003](/article/10.1007/s00436-013-3473-5#ref-CR5 "Blackhall WJ, Prichard RK, Beech RN (2003) Selection at a gamma-aminobutyric acid receptor gene in Haemonchus contortus resistant to avermectins/milbemycins. Mol Biochem Parasitol 131(2):137–145"); Martin et al. [1998](/article/10.1007/s00436-013-3473-5#ref-CR30 "Martin RJ, Murray I, Robertson AP, Bjorn H, Sangster N (1998) Anthelmintics and ion-channels: after a puncture, use a patch. Int J Parasitol 28(6):849–862"); Rana and Misra-Bhattacharya [2013](/article/10.1007/s00436-013-3473-5#ref-CR39 "Rana AK, Misra-Bhattacharya S (2013) Current drug targets for helminthic diseases. Parasitol Res 112:1819–1831. doi:
10.1007/s00436-013-3383-6
")). It has been shown for the model organism _Caenorhabitidis elegans_ (Cully et al. [1994](/article/10.1007/s00436-013-3473-5#ref-CR6 "Cully DF et al (1994) Cloning of an avermectin-sensitive glutamate-gated chloride channel from Caenorhabditis elegans. Nature 371(6499):707–711. doi:
10.1038/371707a0
"); Dent et al. [2000](/article/10.1007/s00436-013-3473-5#ref-CR10 "Dent JA, Smith MM, Vassilatis DK, Avery L (2000) The genetics of ivermectin resistance in Caenorhabditis elegans. Proc Natl Acad Sci USA 97(6):2674–2679"); Lynagh and Lynch [2010](/article/10.1007/s00436-013-3473-5#ref-CR28 "Lynagh T, Lynch JW (2010) A glycine residue essential for high ivermectin sensitivity in Cys-loop ion channel receptors. Int J Parasitol 40(13):1477–1481. doi:
10.1016/j.ijpara.2010.07.010
"); McCavera et al. [2007](/article/10.1007/s00436-013-3473-5#ref-CR31 "McCavera S, Walsh TK, Wolstenholme AJ (2007) Nematode ligand-gated chloride channels: an appraisal of their involvement in macrocyclic lactone resistance and prospects for developing molecular markers. Parasitology 134(Pt 8):1111–1121. doi:
10.1017/S0031182007000042
")) as well as for _C. oncophora_ (Njue et al. [2004](/article/10.1007/s00436-013-3473-5#ref-CR33 "Njue AI, Hayashi J, Kinne L, Feng XP, Prichard RK (2004) Mutations in the extracellular domains of glutamate-gated chloride channel alpha3 and beta subunits from ivermectin-resistant Cooperia oncophora affect agonist sensitivity. J Neurochem 89(5):1137–1147. doi:
10.1111/j.1471-4159.2004.02379.x
")) that IVM resistance can be associated with mutations in _GluCl_, resulting in receptor conformation changes and lowered sensitivity to IVM. However, the mechanisms behind IVM resistance are complex, and there are many studies that suggest the involvement of P-glycoproteins (PGP). PGPs are part of a larger superfamily of transmembrane efflux transporters that have been implicated as a primary cause of anthelmintic resistance both in _C. elegans_ and in the abomasal sheep nematode _Haemonchus contortus_ (Blackhall et al. [1998a](/article/10.1007/s00436-013-3473-5#ref-CR3 "Blackhall WJ, Liu HY, Xu M, Prichard RK, Beech RN (1998a) Selection at a P-glycoprotein gene in ivermectin- and moxidectin-selected strains of Haemonchus contortus. Mol Biochem Parasitol 95(2):193–201"); James and Davey [2009](/article/10.1007/s00436-013-3473-5#ref-CR19 "James CE, Davey MW (2009) Increased expression of ABC transport proteins is associated with ivermectin resistance in the model nematode Caenorhabditis elegans. Int J Parasitol 39(2):213–220. doi:
10.1016/j.ijpara.2008.06.009
"); Kerboeuf et al. [2003a](/article/10.1007/s00436-013-3473-5#ref-CR22 "Kerboeuf D, Blackhall W, Kaminsky R, von Samson-Himmelstjerna G (2003a) P-glycoprotein in helminths: function and perspectives for anthelmintic treatment and reversal of resistance. Int J Antimicrob Agents 22(3):332–346")). PGPs, for which IVM is a well-known substrate (Kerboeuf and Guegnard [2011](/article/10.1007/s00436-013-3473-5#ref-CR21 "Kerboeuf D, Guegnard F (2011) Anthelmintics are substrates and activators of nematode P glycoprotein. Antimicrob Agents Chemother 55(5):2224–2232. doi:
10.1128/AAC.01477-10
"); Lespine et al. [2008](/article/10.1007/s00436-013-3473-5#ref-CR25 "Lespine A, Alvinerie M, Vercruysse J, Prichard RK, Geldhof P (2008) ABC transporter modulation: a strategy to enhance the activity of macrocyclic lactone anthelmintics. Trends Parasitol 24(7):293–298. doi:
10.1016/j.pt.2008.03.011
")), are members of the ATP binding cassette (ABC) superfamily of genes coding for molecules involved in active transport of endo- and exogenous hydrophobic molecules (Jones and George [2005](/article/10.1007/s00436-013-3473-5#ref-CR20 "Jones PM, George AM (2005) Multidrug resistance in parasites: ABC transporters, P-glycoproteins and molecular modelling. Int J Parasitol 35(5):555–566. doi:
10.1016/j.ijpara.2005.01.012
")). Dupuy et al. ([2010](/article/10.1007/s00436-013-3473-5#ref-CR12 "Dupuy J, Alvinerie M, Menez C, Lespine A (2010) Interaction of anthelmintic drugs with P-glycoprotein in recombinant LLC-PK1-mdr1a cells. Chem Biol Interact 186(3):280–286. doi:
10.1016/j.cbi.2010.05.013
")) showed that PGPs are involved in the pharmacokinetics of IVM, and recently Kerboeuf and Guegnard ([2011](/article/10.1007/s00436-013-3473-5#ref-CR21 "Kerboeuf D, Guegnard F (2011) Anthelmintics are substrates and activators of nematode P glycoprotein. Antimicrob Agents Chemother 55(5):2224–2232. doi:
10.1128/AAC.01477-10
")) found evidence that MLs activate transport activity in nematode PGP, and suggest that several substituents in the ML structure are involved in modulating the stimulatory effect. Altogether, 14 isoforms of PGP plus 1 pseudogene have been annotated in the _C. elegans_ genome database (WormBase, version: WS234).The majority of ML-resistance studies to date have investigated target site mutations. Some attention has been paid to the role of changes in gene expression, and upregulation in helminth PGP gene expression has been suggested to enhance the parasite’s ability to survive exposure to IVM. The potential effect of this may be an increased membrane transport and thus faster drug elimination. Recently, PGP expression from several variants of PGP in Teladorsagia circumcincta from sheep was investigated in IVM-resistant isolates (Dicker et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR11 "Dicker AJ, Nisbet AJ, Skuce PJ (2011) Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int J Parasitol 41(9):935–942. doi: 10.1016/j.ijpara.2011.03.015
")). One of the PGP, _Tci-Pgp-9_, showed increased expression at the mRNA level and nucleotide sequences also showed high levels of polymorphism, which could play an important role in helminth survival of IVM exposure. Pachnicke ([2009](/article/10.1007/s00436-013-3473-5#ref-CR36 "Pachnicke S (2009) Molecular mechanisms involved in anthelmintic resistance. Dissertation. A thesis submitted for the degree of Doctor of philosophy in the subject of Parasitology. Institute for Parasitology, University of Veterinary Medicine, Hannover")) performed similar studies in a benzimidazole (BZ)-selected _O. ostertagi_ population and found evidence of increased PGP-related signals in eggshells from BZ-selected _O. ostertagi_, but no signs of increased PGP gene expression in adult worms.In the summers of 2006 and 2007, the efficacy of ML (Ivomec inj.®) was investigated in five cattle herds in the Uppsala region of central Sweden (Demeler et al. [2009](/article/10.1007/s00436-013-3473-5#ref-CR8 "Demeler J et al (2009) Monitoring the efficacy of ivermectin and albendazole against gastro intestinal nematodes of cattle in Northern Europe. Vet Parasitol 160(1–2):109–115. doi: 10.1016/j.vetpar.2008.10.030
")). The evaluation was made among first season grazing (FSG) cattle some weeks after turnout, and was conducted using the faecal egg count reduction test (FECRT). It showed unsatisfactory efficacy results, with only one farm reaching acceptable reductions of egg output in 2006\. In 2007, the reduction was insufficient on all farms where animals were treated with IVM. Isolates from two of these farms were collected, and a pen trial was carried out to investigate the effect of injectable IVM under controlled conditions (unpublished data, Areskog et al.). The objectives of the current study were to further characterise the recovered worms in vitro and to compare PGP expression in adult worms before and after treatment.Materials and methods
Parasites
During 2009, an IVM selection trial was carried out where three groups, each of 11 FSG, were experimentally inoculated with mixtures of 40,000 infective third stage (L3) larvae of C. oncophora and O. ostertagi. The study in its entirety will be presented in a separate publication, but in brief, control group A received an equal mixture of L3 from each of two laboratory isolates from Tierärztliche Hochschule, Hannover, originating from the Central Veterinary Laboratory at Weybridge, UK. Both isolates had no history of being refractory to treatment with any anthelmintics in vitro (Demeler et al. [2010](/article/10.1007/s00436-013-3473-5#ref-CR9 "Demeler J, Kuttler U, von Samson-Himmelstjerna G (2010) Adaptation and evaluation of three different in vitro tests for the detection of resistance to anthelmintics in gastro intestinal nematodes of cattle. Vet Parasitol 170(1–2):61–70. doi: 10.1016/j.vetpar.2010.01.032
")). Calves in groups B and C were inoculated with mixed L3 isolated from the Swedish farms showing poor reductions to IVM according to FECRT: 84 % \[95 % confidence interval (CI), 67–95\] and 85 % (62–99) reductions, respectively, at day 7 (Demeler et al. [2009](/article/10.1007/s00436-013-3473-5#ref-CR8 "Demeler J et al (2009) Monitoring the efficacy of ivermectin and albendazole against gastro intestinal nematodes of cattle in Northern Europe. Vet Parasitol 160(1–2):109–115. doi:
10.1016/j.vetpar.2008.10.030
")). All animals were injected with IVM at the standard dose rate of 0.2 mg IVM per kg 35 days post-infection (d.p.i). Four days before IVM treatment (31 d.p.i), one animal per group (A, B and C) was euthanized intravenously with pentobarbital (ex tempore 300 mg/ml, 60 ml/calf). Another two animals per group were euthanized 10 days post-IVM treatment (45 d.p.i). Individual faecal samples were also collected rectally at the day of slaughter. A modified McMaster method (Anonymous [1986](/article/10.1007/s00436-013-3473-5#ref-CR1 "Anonymous (1986) Manual of veterinary parasitological laboratory techniques. Reference book 418. Her Majesty’s Stationary Office, London, p 166")) was used to determine the number of parasite eggs in 3 g of faeces, giving a diagnostic sensitivity of 50 eggs per gram faeces (EPG). Animals were treated, and the trial was performed according to formal institutional ethical approval (C276/8).At slaughter, worms were basically recovered according to Larsson et al. (2007). Briefly, the abomasum and ∼7 m of the proximal small intestine (duodenum, jejunum) were separated, and their content emptied into individual vessels. The mucosal surfaces were washed with tap water, and from each animal, five male and five female adult worms were immediately recovered and stored at –80 °C in individual Eppendorf tubes until analysis. In addition, 20-ml aliquot subsamples were collected and stored at –20 °C for worm counts and additional analysis. Also, the abomasal mucosa was scraped off and digested in a pepside solution for O. ostertagi recovery.
Isolation of RNA
Total worm RNA was isolated, using Macherey-Nagel NucleoSpin RNA II columns with DNAse I, individually from five single male and five female C. oncophora each pre- and post (10 days) IVM treatment, from three different isolates, giving in total 60 worms. The three isolates represented the propagated susceptible control group plus two isolates showing signs of phenotypic clinical ML resistance according to previous FECRT results. Washing buffers and rDNAse buffer were used to remove contaminating DNA, and integrity of the RNA was checked visually following electrophoresis. Eluates were kept frozen at –80 °C for long-term storage. RNA quantification was performed according to the RiboGreen protocol (Invitrogen) using aVictor2 1420 Multilabel counter (Wallac).
Primer design
A conserved 202-bp PGP sequence from C. oncophora (Pachnicke 2009) was identified by ClustalW2 as having 83 % similarity to the PGP-9 of C. elegans. Primers specific to this conserved PGP sequence were then designed in Primer3, generating a 153-bp PCR product. Sequencing of the PCR product was performed, followed by a BLAST search in Wormbase (genome view), which was identified as WBGene00004003.
RT-qPCR
One-tube RT-PCR QuantiTect with SYBR Green (Qiagen) was used to measure PGP gene expression by RT-qPCR using known concentrations of PGP-PCR product as standards and non-template controls as blanks. The primer sequences used were C. elegans Pgp-9 Forward 5′ CGA ACG AGG TGT GCA ACT AA 3′ and C. elegans Pgp-9 Reverse 5′ GCC TTC TCC AAA GCT TCT TGT 3′. The 25-μl RT-PCR reactions contained 12.5 μl 2× QIAGEN Quanti Tect SYBR Green master mix, 0.5 μl Forward primer (10 μM), 0.5 μl reverse primer (10 μM), 0.25 μl Q-RT, 1.25 μl of RNAse-free H2O and 10 μl RNA sample adjusted to contain 2 ng total RNA from each single worm. Thus, PGP expression was normalised to the amount of total RNA in the PCR reaction as previously described by Tyden et al. ([2009](/article/10.1007/s00436-013-3473-5#ref-CR43 "Tyden E, Tallkvist J, Tjalve H, Larsson P (2009) P-glycoprotein in intestines, liver, kidney and lymphocytes in horse. J Vet Pharmacol Ther 32(2):167–176. doi: 10.1111/j.1365-2885.2008.01017.x
")). All samples were run in technical duplicates. To determine the level of expression of PGPs before and after treatment, a standard curve was applied using five concentrations of the purified and sequenced 153 bp PGP-PCR product diluted from 7,000 to 0.7 ag μl−1 in tenfold titrations. The PGP-PCR product was quantified by applying PicoGreen reagent (Invitrogen) according to the instructions of the manufacturer and a Victor2 1420 Multilabel counter (Wallac). To minimise the risk of amplifying genomic DNA, the samples were DNAse treated during RNA isolation, as previously described.RNA samples were run in a Rotor-Gene 3000 (Corbett) and the data analysed using Rotor-Gene 6.1.90 software (Corbett). The RT-qPCR conditions were as follows: RT step, 50 °C for 30 min; Taq-polymerase activation step, 95 °C for 15 min followed by 45 cycles of 94 °C for 60 s, 55 °C for 60 s and 68 °C for 45 s and a final extension step at 68 °C for 7 min. Melting curve analysis was performed for each sample in direct connection to the PCR, to verify that no secondary products were generated. To quantify the relative changes in expression of putative Con-pgp-9 before and after IVM treatment, the 2−Δ_C_T method of Livak and Schmittgen ([2001](/article/10.1007/s00436-013-3473-5#ref-CR26 "Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25(4):402–408. doi: 10.1006/meth.2001.1262
")) was used. In addition, in order to measure absolute _Con-pgp-9_ expression, absolute gene concentrations were also calculated from the standard curve.Amplified fragment length polymorphism
DNA extraction
DNA was prepared from individual worms using a QIAamp DNA Micro Kit (Qiagen) according to the manufacturer’s recommendation. Samples were eluted in 30 μl buffer AE. Treatment with RNaseA was performed post-DNA isolation by adding 60 μg of RNaseA to 30 μl of sample and incubating for 5 min at room temperature to obtain RNA-free samples. The purified DNA was stored at −20 °C until use.
AFLP procedure
In total, 108 adult C. oncophora of both sexes were used for AFLP analysis, but unlike the worms used for gene expression studies, these were recovered from aliquots frozen at −18 °C (Table 1). The AFLP procedure was performed basically as described by Applied Biosystems (ABI) in their Plant Mapping Protocol and in accordance with Höglund et al. ([2004](/article/10.1007/s00436-013-3473-5#ref-CR16 "Höglund J, Engström A, Morrison DA, Mattsson JG (2004) Genetic diversity assessed by amplified fragment length polymorphism analysis of the parasitic nematode Dictyocaulus viviparus the lungworm of cattle. Int J Parasitol 34(4):475–484. doi: 10.1016/j.ijpara.2003.11.007
")). All reagents were supplied in the AFLP® Plant Mapping Kit (ABI), except for the restriction enzymes and the T4 DNA ligase, which were purchased from New England BioLabs. For all worms, the maximum allowed volume, 5.5 μl, of extracted DNA was used. The DNA from each individual worm was digested with 1 unit of MseI and 5 units of EcoRI, and MseI adaptor and EcoRI adaptor were ligated in the same reaction as the digestion. After the restriction–ligation reaction, the mixture was diluted to a total volume of 50 μl with TE0.1 (20 mM Tris–HCl, 0.1 mM EDTA, pH 8.0). From the restriction–ligation reaction, 4 μl of the diluted DNA was mixed with 1 μl of the ABI pre-selective primer pairs (5 μM) and 15 μl of ABI’s Core Mix for pre-selective amplification. The amplified products were diluted to a final volume of 200 μl. The FAM dyed EcoRI-ACA + MseI-CTC primer combination was used for selective amplification. From the selective amplification products, 1.0 μl was mixed with 9.6 μl deionised formamide and 0.4 μl GeneScan-500 size standard (ABI). The mixture was denaturated for 3 min at 95 °C, before being loaded onto the ABI Prism 3100 Genetic Analyzer and analysed using GeneMapper analysis software 4.0.Table 1 EPG data from day of slaughter and estimated numbers of adult worms recovered in 20 ml subsamples at necropsy, from calves before and 10 days after injection with ivermectin (Ivomec®, Merial) at a dose rate of 0.2 mg IVM/kg
Statistical analysis
Data were summarised in Microsoft Excel (2007), whereas statistical analyses and graphs were done in Minitab (version 15). Gene expression was analysed by sex using both log transformed- and original concentrations, and where we compared duplicate mean concentrations, before and after treatment with two sample t tests. In addition, a three-factor ANOVA (GLM) comparing treatment × isolate × sex, with isolate as a random factor, was performed. The significance level was set to p < 0.05.
The estimates of allele frequencies for each isolate used the Bayesian method with informative priors of Zhivotovsky (1999). The calculation of genetic diversity (H j) and population structure (F st) from these estimates followed the procedures of Lynch and Milligan (1994), with 2,000 randomizations used for the statistical tests. These statistics were all computed using the AFLPSurvey program of Vekemans (2002).
Results
Parasites
Faecal egg counts revealed patent infections in all animals at slaughter, both pre-treatment and 10 days post-IVM treatment (Table 1). At necropsy, mainly low to moderate numbers of C. oncophora remained in the small intestines of all IVM-treated animals (Table 1). The worm burden showed large variation within groups and no significant difference (p = 0.510) between groups.
RT-qPCR
The expression of the putative Con-pgp-9 before treatment was compared to the results after treatment for each isolate, and males and females separately. Reaction efficiency of the qPCR was 1.08 in females and 0.97 in males. All samples had concentrations within the range of the standard curve, and female worms had an approximately five times higher gene expression compared to male worms before treatment (Table 2).
Table 2 RT-qPCR results showing concentrations of putative Con-pgp-9 RNA expression (ag/2 ng total RNA) in isolates before and 10 days after selection with injectable ivermectin at a dose rate of 0.2 mg ivermectin/kg body weight
The RT-qPCR results demonstrated that mean putative Con-pgp-9 expression in ML-treated male worms was 70 % higher than in the untreated worms, with mean concentrations of 27 (±25) ag μl−1before treatment and 46.2 (±32) ag μl−1 after treatment, respectively (Table 2). Although mean concentrations and boxplots (Fig. 2) suggested a tendency for increased Con-pgp-9 gene expression after treatment, the statistical analysis showed no significant difference (p = 0.091).
Fig. 2
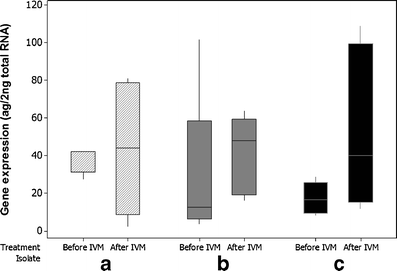
Boxplot of quantitative putative Con-pgp-9 expression, males, all groups, 15 worms before- and 15 worms after selection. Isolate A was propagated IVM susceptible, whereas B and C were two different field isolates showing phenotypic clinical IVM resistance in field trials
In females, mean concentrations of PGP were 172 (±SD of 124) ag μl−1 before treatment and 196 (±250) ag μl−1 after treatment (Table 2). However, as with the male worms, no significant difference (p = 0.747) in concentrations was seen in comparisons between any group pre- and post-selection (Fig. 3).
Fig. 3
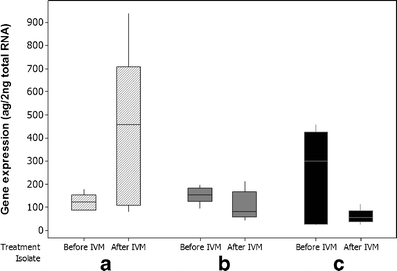
Boxplot of quantitative putative Con-pgp-9 expression, females, all groups, 15 worms before- and 15 worms after selection. Isolate A was propagated IVM susceptible, whereas B and C were two different field isolates showing phenotypic clinical IVM resistance in field trials
The two-sample t test showed large variance within female worm groups. Comparing sex and treatment gave no significance (p = 0.091 and p = 0.747, respectively), but the three-factor ANOVA, with isolate as a random factor, showed that the isolates behaved very differently (p = 0.006). Log transformed concentrations gave similar results (data not shown).
AFLP
AFLP showed no bottleneck effect within the population after treatment, with mean gene diversity (H j) values of 0.158 before treatment and 0.153 after treatment (Table 3). Inbreeding coefficient (F ST) in subpopulations compared to the total population was 0.0112.
Table 3 AFLP results derived from adult worms showing high gene diversity (H j) values ranging between 0.139 and 0.169 in isolates before and 10 days after selection with injectable IVM at a dose rate of 0.2 mg IVM/kg
Discussion
In closely related GI nematodes of sheep, ML resistance has been suggested to be linked to an overexpression of PGPs (Blackhall et al. 1998a; Dicker et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR11 "Dicker AJ, Nisbet AJ, Skuce PJ (2011) Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int J Parasitol 41(9):935–942. doi: 10.1016/j.ijpara.2011.03.015
"); Williamson et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR45 "Williamson SM, Storey B, Howell S, Harper KM, Kaplan RM, Wolstenholme AJ (2011) Candidate anthelmintic resistance-associated gene expression and sequence polymorphisms in a triple-resistant field isolate of Haemonchus contortus. Mol Biochem Parasitol 180(2):99–105. doi:
10.1016/j.molbiopara.2011.09.003
")). Our hypothesis in this study was that changes in expression of efflux pumps such as PGPs would be affected in _C. oncophora_, which survive ML treatment, and/or in isolates with different deworming histories. Increased expression of this molecule has previously been suggested to play a role in drug resistance towards malaria, HIV and cancer, and in nematodes, it has been connected to ML resistance (Gupta and Gollapudi [1993](/article/10.1007/s00436-013-3473-5#ref-CR14 "Gupta S, Gollapudi S (1993) P-glycoprotein (MDR 1 gene product) in cells of the immune system: its possible physiologic role and alteration in aging and human immunodeficiency virus-1 (HIV-1) infection. J Clin Immunol 13(5):289–301"); James and Davey [2009](/article/10.1007/s00436-013-3473-5#ref-CR19 "James CE, Davey MW (2009) Increased expression of ABC transport proteins is associated with ivermectin resistance in the model nematode Caenorhabditis elegans. Int J Parasitol 39(2):213–220. doi:
10.1016/j.ijpara.2008.06.009
"); Loo and Clarke [1999](/article/10.1007/s00436-013-3473-5#ref-CR27 "Loo TW, Clarke DM (1999) Merck Frosst Award Lecture 1998. Molecular dissection of the human multidrug resistance P-glycoprotein. Biochem Cell Biol 77(1):11–23"); Xu et al. [1998](/article/10.1007/s00436-013-3473-5#ref-CR47 "Xu M, Molento M, Blackhall W, Ribeiro P, Beech R, Prichard R (1998) Ivermectin resistance in nematodes may be caused by alteration of P-glycoprotein homolog. Mol Biochem Parasitol 91(2):327–335")). Unlike most studies previously performed, we made an attempt to measure PGP expression levels in individual adult worms of _C. oncophora_, employing replicates reflecting the biological diversity in PGP expression. Our results demonstrate a wide distribution and no statistically significant differences in _Con-pgp-9_ expression pre- and post-IVM treatment.It has previously been reported that several anthelmintics may cause an augmented PGP expression in various nematodes (Pachnicke 2009; Williamson et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR45 "Williamson SM, Storey B, Howell S, Harper KM, Kaplan RM, Wolstenholme AJ (2011) Candidate anthelmintic resistance-associated gene expression and sequence polymorphisms in a triple-resistant field isolate of Haemonchus contortus. Mol Biochem Parasitol 180(2):99–105. doi: 10.1016/j.molbiopara.2011.09.003
"); Dicker et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR11 "Dicker AJ, Nisbet AJ, Skuce PJ (2011) Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int J Parasitol 41(9):935–942. doi:
10.1016/j.ijpara.2011.03.015
")). The reasons for the discrepancy between our results and the ones previously reported are speculative. However, one possible explanation may include analyses of pooled samples missing the biological diversity by performing statistics on technical repeats. Also, use of BZ instead of IVM may influence the PGP expression in a different manner, or worm-specific sensitivity in the upregulation of PGP. Thus, it cannot be excluded that _C. oncophora_ responds differently to anthelmintic treatment than _O. ostertagi_ (Pachnicke [2009](/article/10.1007/s00436-013-3473-5#ref-CR36 "Pachnicke S (2009) Molecular mechanisms involved in anthelmintic resistance. Dissertation. A thesis submitted for the degree of Doctor of philosophy in the subject of Parasitology. Institute for Parasitology, University of Veterinary Medicine, Hannover")), _H. contortus_ (Williamson et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR45 "Williamson SM, Storey B, Howell S, Harper KM, Kaplan RM, Wolstenholme AJ (2011) Candidate anthelmintic resistance-associated gene expression and sequence polymorphisms in a triple-resistant field isolate of Haemonchus contortus. Mol Biochem Parasitol 180(2):99–105. doi:
10.1016/j.molbiopara.2011.09.003
")) and _T. circumcincta_ (Dicker et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR11 "Dicker AJ, Nisbet AJ, Skuce PJ (2011) Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int J Parasitol 41(9):935–942. doi:
10.1016/j.ijpara.2011.03.015
")). It is also often hypothesized that anthelmintic exposure itself induces expression of ABC transporter genes in nematodes, experimental evidence is still scarce (Ardelli and Prichard [2008](/article/10.1007/s00436-013-3473-5#ref-CR2 "Ardelli BF, Prichard RK (2008) Effects of ivermectin and moxidectin on the transcription of genes coding for multidrug resistance associated proteins and behaviour in Caenorhabditis elegans. J Nematol 40:290–298"); James and Davey [2009](/article/10.1007/s00436-013-3473-5#ref-CR19 "James CE, Davey MW (2009) Increased expression of ABC transport proteins is associated with ivermectin resistance in the model nematode Caenorhabditis elegans. Int J Parasitol 39(2):213–220. doi:
10.1016/j.ijpara.2008.06.009
")).The results showed no significant differences before and after ML treatment between the three C. oncophora isolates (A, B and C), each with a different deworming history, possibly due to small sample sizes. However, there was a tendency towards increasing putative Con-pgp-9 expression in male worms (p = 0.091). Female worms seemed to give higher and more variable results (p = 0.747) than male worms, possibly due to the presence of varying amounts of eggs also expressing PGP (Kerboeuf et al. 2003b; Riou et al. [2005](/article/10.1007/s00436-013-3473-5#ref-CR40 "Riou M, Koch C, Delaleu B, Berthon P, Kerboeuf D (2005) Immunolocalisation of an ABC transporter, P-glycoprotein, in the eggshells and cuticles of free-living and parasitic stages of Haemonchus contortus. Parasitol Res 96(3):142–148. doi: 10.1007/s00436-005-1345-3
")). The non-significant statistical results in the comparisons between sex and treatment might be a result of different expression patterns in the investigated isolates. For instance, females in groups B and C actually reduced their mean _Con-pgp-9_ expression after ML treatment. This was in contrast to all the other groups and, hypothetically, it could be explained by a decrease in intrauterine egg production, and thus lower mRNA levels for expression of PGP in egg shells during IVM influences. It has previously been shown that the nematode uterine muscle is one of the most susceptible target organs for MLs (Prichard [2001](/article/10.1007/s00436-013-3473-5#ref-CR37 "Prichard R (2001) Genetic variability following selection of Haemonchus contortus with anthelmintics. Trends Parasitol 17(9):445–453")), which could mean that the drug may temporarily suppress egg laying even though adult worms survive treatment. This would make fecund female worms unsuitable for investigation as it causes difficulties in distinguishing between worm- and egg PGP expression levels. However, the same situation did not seem to occur among females in the lab isolate, group A. The differences between groups, rather than differences before or after treatment, were also confirmed by statistical analysis. Clearly, a larger sample size is required to better ensure the effect of ML on individual worms, but the sampling method is both labor-intensive, expensive and demands euthanizing of animals to collect large number of adults surviving deworming. Sometimes we also had problems to isolate enough RNA from the worms, especially from the males, and the number of surviving worms was limited.Our results therefore can neither support nor exclude that Con-pgp-9 is involved in the increased incidence of resistance to IVM in this cattle nematode, as has recently been reported for T. circumcincta in sheep (Dicker et al. [2011](/article/10.1007/s00436-013-3473-5#ref-CR11 "Dicker AJ, Nisbet AJ, Skuce PJ (2011) Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int J Parasitol 41(9):935–942. doi: 10.1016/j.ijpara.2011.03.015
")). Another factor that may explain the differing results is the time of sampling. Pachnicke ([2009](/article/10.1007/s00436-013-3473-5#ref-CR36 "Pachnicke S (2009) Molecular mechanisms involved in anthelmintic resistance. Dissertation. A thesis submitted for the degree of Doctor of philosophy in the subject of Parasitology. Institute for Parasitology, University of Veterinary Medicine, Hannover")) collected worms 7 days post-treatment, while in other studies, it is unclear when donor animals were sacrificed. In this study, we investigated worms 10 days post-treatment; however, it is not known when would be the best time to measure changes in _Con-Pgp-9_ expression after ML treatment. In a most recently published investigation conducted in parallel to this, De Graef et al. ([2013](/article/10.1007/s00436-013-3473-5#ref-CR7 "De Graef J et al (2013) Gene expression analysis of ABC transporters in a resistant Cooperia oncophora isolate following in vivo and in vitro exposure to macrocyclic lactones. Parasitology: 1–10 doi:
10.1017/S0031182012001849
")) examined effects of IVM treatment on the transcription of _C. oncophora_ PGPs. In agreement with our findings, no significant differences with respect to _Con_\-_Pgp-9_ expression were recorded between adults of susceptible and IVM-resistant Belgian _C. oncophora_ isolates. Neither could any differences be seen between adults from the resistant isolate being ML-exposed or unexposed. However, in the latter adult worms, a 3.1- and 4.7-fold increased expression of _Con-Pgp-11_ was encountered in association with IVM and moxidectin, respectively, after in vivo exposure. Further experiments are thus clearly required to verify the results obtained in the present study, but also to elucidate additional resistance mechanisms that may be involved.When our C. oncophora isolates were analysed with AFLP, we found no evidence of genetic bottlenecking in either of the drug-selected worms, and by comparing their genetic makeup pre- and post-treatment. In veterinary parasitology, AFLP has previously been used mainly to compare the genetic structures of various parasitic nematodes of livestock with different geographic backgrounds (Höglund et al. [2012](/article/10.1007/s00436-013-3473-5#ref-CR18 "Höglund J, Morrison DA, Engström A, Nejsum P, Jansson DS (2012) Population genetic structure of Ascaridia galli re-emerging in non-caged laying hens. Parasit Vectors 5:97. doi: 10.1186/1756-3305-5-97
"), [2006](/article/10.1007/s00436-013-3473-5#ref-CR17 "Höglund J, Morrison DA, Mattsson JG, Engström A (2006) Population genetics of the bovine/cattle lungworm (Dictyocaulus viviparus) based on mtDNA and AFLP marker techniques. Parasitology 133(Pt 1):89–99. doi:
10.1017/S0031182006009991
"); Nejsum et al. [2005](/article/10.1007/s00436-013-3473-5#ref-CR32 "Nejsum P, Frydenberg J, Roepstorff A, Parker ED Jr (2005) Population structure in Ascaris suum (Nematoda) among domestic swine in Denmark as measured by whole genome DNA fingerprinting. Hereditas 142(2005):7–14. doi:
10.1111/j.1601-5223.2005.01864.x
"); Troell et al. [2006](/article/10.1007/s00436-013-3473-5#ref-CR42 "Troell K, Engström A, Morrison DA, Mattsson JG, Höglund J (2006) Global patterns reveal strong population structure in Haemonchus contortus, a nematode parasite of domesticated ruminants. Int J Parasitol 36(12):1305–1316. doi:
10.1016/j.ijpara.2006.06.015
")), but also to compare consecutive stages of increased benzimidazole- and levamisole-resistant isolates during selection (Otsen et al. [2001](/article/10.1007/s00436-013-3473-5#ref-CR35 "Otsen M, Hoekstra R, Plas ME, Buntjer JB, Lenstra JA, Roos MH (2001) Amplified fragment length polymorphism analysis of genetic diversity of Haemonchus contortus during selection for drug resistance. Int J Parasitol 31(10):1138–1143")). Höglund et al. ([2006](/article/10.1007/s00436-013-3473-5#ref-CR17 "Höglund J, Morrison DA, Mattsson JG, Engström A (2006) Population genetics of the bovine/cattle lungworm (Dictyocaulus viviparus) based on mtDNA and AFLP marker techniques. Parasitology 133(Pt 1):89–99. doi:
10.1017/S0031182006009991
")) concluded that most nematodes of domesticated hosts usually have very little genetic difference between populations, and are characterised by strong gene flow and expanding populations. In accordance with Otsen et al. ([2001](/article/10.1007/s00436-013-3473-5#ref-CR35 "Otsen M, Hoekstra R, Plas ME, Buntjer JB, Lenstra JA, Roos MH (2001) Amplified fragment length polymorphism analysis of genetic diversity of Haemonchus contortus during selection for drug resistance. Int J Parasitol 31(10):1138–1143")), we found no significant changes in the genetic diversity of isolates associated with anthelmintic selection. However, in contrast to the high level of genetic variation in _H. contortus_, our obtained _C. oncophora_ inbreeding coefficient, _F_ ST, was low, suggesting no genetic differentiation. This agrees with the results from Höglund et al. ([2006](/article/10.1007/s00436-013-3473-5#ref-CR17 "Höglund J, Morrison DA, Mattsson JG, Engström A (2006) Population genetics of the bovine/cattle lungworm (Dictyocaulus viviparus) based on mtDNA and AFLP marker techniques. Parasitology 133(Pt 1):89–99. doi:
10.1017/S0031182006009991
")) and could mean that, if genetic resistance establishes regionally, then a rapid spread of resistance development would be enabled within the equivalent genetic pool.In conclusion, comparison of putative Con-pgp-9 expression before and after IVM treatment showed no significant difference between the tested groups, but some tendency towards increasing expression in male as compared to female worms. Additional research is required to elucidate whether AR observed in C. oncophora involves increased expression and activity of PGP or whether this is related to some other transmembrane efflux transporter or metabolic enzyme.
References
- Anonymous (1986) Manual of veterinary parasitological laboratory techniques. Reference book 418. Her Majesty’s Stationary Office, London, p 166
Google Scholar - Ardelli BF, Prichard RK (2008) Effects of ivermectin and moxidectin on the transcription of genes coding for multidrug resistance associated proteins and behaviour in Caenorhabditis elegans. J Nematol 40:290–298
CAS Google Scholar - Blackhall WJ, Liu HY, Xu M, Prichard RK, Beech RN (1998a) Selection at a P-glycoprotein gene in ivermectin- and moxidectin-selected strains of Haemonchus contortus. Mol Biochem Parasitol 95(2):193–201
Article PubMed CAS Google Scholar - Blackhall WJ, Pouliot JF, Prichard RK, Beech RN (1998b) Haemonchus contortus: selection at a glutamate-gated chloride channel gene in ivermectin- and moxidectin-selected strains. Exp Parasitol 90(1):42–48. doi:10.1006/expr.1998.4316
Article PubMed CAS Google Scholar - Blackhall WJ, Prichard RK, Beech RN (2003) Selection at a gamma-aminobutyric acid receptor gene in Haemonchus contortus resistant to avermectins/milbemycins. Mol Biochem Parasitol 131(2):137–145
Article PubMed CAS Google Scholar - Cully DF et al (1994) Cloning of an avermectin-sensitive glutamate-gated chloride channel from Caenorhabditis elegans. Nature 371(6499):707–711. doi:10.1038/371707a0
Article PubMed CAS Google Scholar - De Graef J et al (2013) Gene expression analysis of ABC transporters in a resistant Cooperia oncophora isolate following in vivo and in vitro exposure to macrocyclic lactones. Parasitology: 1–10 doi:10.1017/S0031182012001849
- Demeler J et al (2009) Monitoring the efficacy of ivermectin and albendazole against gastro intestinal nematodes of cattle in Northern Europe. Vet Parasitol 160(1–2):109–115. doi:10.1016/j.vetpar.2008.10.030
Article PubMed CAS Google Scholar - Demeler J, Kuttler U, von Samson-Himmelstjerna G (2010) Adaptation and evaluation of three different in vitro tests for the detection of resistance to anthelmintics in gastro intestinal nematodes of cattle. Vet Parasitol 170(1–2):61–70. doi:10.1016/j.vetpar.2010.01.032
Article PubMed CAS Google Scholar - Dent JA, Smith MM, Vassilatis DK, Avery L (2000) The genetics of ivermectin resistance in Caenorhabditis elegans. Proc Natl Acad Sci USA 97(6):2674–2679
Article PubMed CAS Google Scholar - Dicker AJ, Nisbet AJ, Skuce PJ (2011) Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int J Parasitol 41(9):935–942. doi:10.1016/j.ijpara.2011.03.015
Article PubMed CAS Google Scholar - Dupuy J, Alvinerie M, Menez C, Lespine A (2010) Interaction of anthelmintic drugs with P-glycoprotein in recombinant LLC-PK1-mdr1a cells. Chem Biol Interact 186(3):280–286. doi:10.1016/j.cbi.2010.05.013
Article PubMed CAS Google Scholar - Gasbarre LC, Smith LL, Lichtenfels JR, Pilitt PA (2009) The identification of cattle nematode parasites resistant to multiple classes of anthelmintics in a commercial cattle population in the US. Vet Parasitol 166(3–4):281–285. doi:10.1016/j.vetpar.2009.08.018
Article PubMed CAS Google Scholar - Gupta S, Gollapudi S (1993) P-glycoprotein (MDR 1 gene product) in cells of the immune system: its possible physiologic role and alteration in aging and human immunodeficiency virus-1 (HIV-1) infection. J Clin Immunol 13(5):289–301
Article PubMed CAS Google Scholar - Höglund J (2010) Parasite surveillance and novel use of anthelmintics in cattle. Acta Vet Scand 52:2–5
Article Google Scholar - Höglund J, Engström A, Morrison DA, Mattsson JG (2004) Genetic diversity assessed by amplified fragment length polymorphism analysis of the parasitic nematode Dictyocaulus viviparus the lungworm of cattle. Int J Parasitol 34(4):475–484. doi:10.1016/j.ijpara.2003.11.007
Article PubMed Google Scholar - Höglund J, Morrison DA, Mattsson JG, Engström A (2006) Population genetics of the bovine/cattle lungworm (Dictyocaulus viviparus) based on mtDNA and AFLP marker techniques. Parasitology 133(Pt 1):89–99. doi:10.1017/S0031182006009991
Article PubMed Google Scholar - Höglund J, Morrison DA, Engström A, Nejsum P, Jansson DS (2012) Population genetic structure of Ascaridia galli re-emerging in non-caged laying hens. Parasit Vectors 5:97. doi:10.1186/1756-3305-5-97
Article PubMed Google Scholar - James CE, Davey MW (2009) Increased expression of ABC transport proteins is associated with ivermectin resistance in the model nematode Caenorhabditis elegans. Int J Parasitol 39(2):213–220. doi:10.1016/j.ijpara.2008.06.009
Article PubMed CAS Google Scholar - Jones PM, George AM (2005) Multidrug resistance in parasites: ABC transporters, P-glycoproteins and molecular modelling. Int J Parasitol 35(5):555–566. doi:10.1016/j.ijpara.2005.01.012
Article PubMed CAS Google Scholar - Kerboeuf D, Guegnard F (2011) Anthelmintics are substrates and activators of nematode P glycoprotein. Antimicrob Agents Chemother 55(5):2224–2232. doi:10.1128/AAC.01477-10
Article PubMed CAS Google Scholar - Kerboeuf D, Blackhall W, Kaminsky R, von Samson-Himmelstjerna G (2003a) P-glycoprotein in helminths: function and perspectives for anthelmintic treatment and reversal of resistance. Int J Antimicrob Agents 22(3):332–346
Article PubMed CAS Google Scholar - Kerboeuf D, Guegnard F, Vern YL (2003b) Detection of P-glycoprotein-mediated multidrug resistance against anthelmintics in Haemonchus contortus using anti-human mdr1 monoclonal antibodies. Parasitol Res 91:79–85
Article PubMed CAS Google Scholar - Larsson A, Dimander SO, Rydzik A, Uggla A, Waller PJ, Höglund J (2007) A 3-year field evaluation of pasture rotation and supplementary feeding to control parasite infection in first-season grazing cattle: dynamics of pasture infectivity. Vet Parasitol 145(1–2):129–137
Article PubMed CAS Google Scholar - Lespine A, Alvinerie M, Vercruysse J, Prichard RK, Geldhof P (2008) ABC transporter modulation: a strategy to enhance the activity of macrocyclic lactone anthelmintics. Trends Parasitol 24(7):293–298. doi:10.1016/j.pt.2008.03.011
Article PubMed CAS Google Scholar - Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25(4):402–408. doi:10.1006/meth.2001.1262
Article PubMed CAS Google Scholar - Loo TW, Clarke DM (1999) Merck Frosst Award Lecture 1998. Molecular dissection of the human multidrug resistance P-glycoprotein. Biochem Cell Biol 77(1):11–23
Article PubMed CAS Google Scholar - Lynagh T, Lynch JW (2010) A glycine residue essential for high ivermectin sensitivity in Cys-loop ion channel receptors. Int J Parasitol 40(13):1477–1481. doi:10.1016/j.ijpara.2010.07.010
Article PubMed CAS Google Scholar - Lynch M, Milligan BG (1994) Analysis of population genetic structure with RAPD markers. Mol Ecol 3(2):91–99
Article PubMed CAS Google Scholar - Martin RJ, Murray I, Robertson AP, Bjorn H, Sangster N (1998) Anthelmintics and ion-channels: after a puncture, use a patch. Int J Parasitol 28(6):849–862
Article PubMed CAS Google Scholar - McCavera S, Walsh TK, Wolstenholme AJ (2007) Nematode ligand-gated chloride channels: an appraisal of their involvement in macrocyclic lactone resistance and prospects for developing molecular markers. Parasitology 134(Pt 8):1111–1121. doi:10.1017/S0031182007000042
Article PubMed CAS Google Scholar - Nejsum P, Frydenberg J, Roepstorff A, Parker ED Jr (2005) Population structure in Ascaris suum (Nematoda) among domestic swine in Denmark as measured by whole genome DNA fingerprinting. Hereditas 142(2005):7–14. doi:10.1111/j.1601-5223.2005.01864.x
Article PubMed Google Scholar - Njue AI, Hayashi J, Kinne L, Feng XP, Prichard RK (2004) Mutations in the extracellular domains of glutamate-gated chloride channel alpha3 and beta subunits from ivermectin-resistant Cooperia oncophora affect agonist sensitivity. J Neurochem 89(5):1137–1147. doi:10.1111/j.1471-4159.2004.02379.x
Article PubMed CAS Google Scholar - Omura S (2008) Ivermectin: 25 years and still going strong. Int J Antimicrob Agents 31(2):91–98. doi:10.1016/j.ijantimicag.2007.08.023
Article PubMed CAS Google Scholar - Otsen M, Hoekstra R, Plas ME, Buntjer JB, Lenstra JA, Roos MH (2001) Amplified fragment length polymorphism analysis of genetic diversity of Haemonchus contortus during selection for drug resistance. Int J Parasitol 31(10):1138–1143
Article PubMed CAS Google Scholar - Pachnicke S (2009) Molecular mechanisms involved in anthelmintic resistance. Dissertation. A thesis submitted for the degree of Doctor of philosophy in the subject of Parasitology. Institute for Parasitology, University of Veterinary Medicine, Hannover
- Prichard R (2001) Genetic variability following selection of Haemonchus contortus with anthelmintics. Trends Parasitol 17(9):445–453
Article PubMed CAS Google Scholar - Prichard RK, von Samson-Himmelstjerna G, Blackhall WJ, Geary TG (2007) Foreword: towards markers for anthelmintic resistance in helminths of importance in animal and human health. Parasitology 134(Pt 8):1073–1076. doi:10.1017/S0031182007000078
Article PubMed CAS Google Scholar - Rana AK, Misra-Bhattacharya S (2013) Current drug targets for helminthic diseases. Parasitol Res 112:1819–1831. doi:10.1007/s00436-013-3383-6
Article PubMed Google Scholar - Riou M, Koch C, Delaleu B, Berthon P, Kerboeuf D (2005) Immunolocalisation of an ABC transporter, P-glycoprotein, in the eggshells and cuticles of free-living and parasitic stages of Haemonchus contortus. Parasitol Res 96(3):142–148. doi:10.1007/s00436-005-1345-3
Article PubMed Google Scholar - Sutherland IA, Leathwick DM (2011) Anthelmintic resistance in nematode parasites of cattle: a global issue? Trends Parasitol 27(4):176–181. doi:10.1016/j.pt.2010.11.008
Article PubMed Google Scholar - Troell K, Engström A, Morrison DA, Mattsson JG, Höglund J (2006) Global patterns reveal strong population structure in Haemonchus contortus, a nematode parasite of domesticated ruminants. Int J Parasitol 36(12):1305–1316. doi:10.1016/j.ijpara.2006.06.015
Article PubMed Google Scholar - Tyden E, Tallkvist J, Tjalve H, Larsson P (2009) P-glycoprotein in intestines, liver, kidney and lymphocytes in horse. J Vet Pharmacol Ther 32(2):167–176. doi:10.1111/j.1365-2885.2008.01017.x
Article PubMed CAS Google Scholar - Vekemans X (2002) AFLP-SURV version 1.0. A Program for genetic diversity analysis with AFLP (and RAPD) population data. version 1.0. edn. Laboratoire de Génétique et Ecologie Végétale, Université Libre de Bruxelles, Belgium
Google Scholar - Williamson SM, Storey B, Howell S, Harper KM, Kaplan RM, Wolstenholme AJ (2011) Candidate anthelmintic resistance-associated gene expression and sequence polymorphisms in a triple-resistant field isolate of Haemonchus contortus. Mol Biochem Parasitol 180(2):99–105. doi:10.1016/j.molbiopara.2011.09.003
Article PubMed CAS Google Scholar - Wolstenholme AJ, Fairweather I, Prichard R, von Samson-Himmelstjerna G, Sangster NC (2004) Drug resistance in veterinary helminths. Trends Parasitol 20(10):469–476. doi:10.1016/j.pt.2004.07.010
Article PubMed CAS Google Scholar - Xu M, Molento M, Blackhall W, Ribeiro P, Beech R, Prichard R (1998) Ivermectin resistance in nematodes may be caused by alteration of P-glycoprotein homolog. Mol Biochem Parasitol 91(2):327–335
Article PubMed CAS Google Scholar - Zhivotovsky LA (1999) Estimating population structure in diploids with multilocus dominant DNA markers. Mol Ecol 8(6):907–913
Article PubMed CAS Google Scholar