The Cytokine Network of Wallerian Degeneration: Tumor Necrosis Factor-α, Interleukin-1α, and Interleukin-1β (original) (raw)
Abstract
Wallerian degeneration (WD) is the inflammatory response of the nervous system to axonal injury, primarily attributable to the production of cytokines, the mediator molecules of inflammation. We presently document the involvement of the inflammatory cytokines TNFα, interleukin (IL)-1α, and IL-1β in peripheral nerve (PNS) injury in C57/BL/6NHSD (C57/BL) mice that display the normal rapid progression of WD (rapid-WD) and C57/BL/6-WLD/OLA/NHSD mice that display abnormal slow progression of WD (slow-WD). TNFα and IL-1α mRNAs were expressed, whereas TNFα but not IL-1α protein was synthesized in intact PNS of C57/BL mice. TNFα and IL-1α protein synthesis and secretion were rapidly upregulated during rapid-WD in Schwann cells. IL-1β mRNA expression and protein synthesis and secretion were induced sequentially in Schwann cells with a delay after injury. Thereafter, recruited macrophages contributed to the production of TNFα, IL-1α, and IL-1β, which in turn augmented myelin phagocytosis by macrophages. Observations suggest that TNFα and IL-1α are the first cytokines with protein production that is upregulated during rapid-WD. TNFα and IL-1α may initiate, therefore, molecular and cellular events in rapid-WD (e.g., the production of additional cytokines and NGF). TNFα, IL-1α, and IL-1β may further regulate, indirectly, macrophage recruitment, myelin removal, regeneration, and neuropathic pain. In contrast to rapid-WD, the production of TNFα, IL-1α, and IL-1β protein was deficient in slow-WD, although their mRNAs were expressed. mRNA expression and protein production of TNFα, IL-1α, and IL-1β were differentially regulated during rapid-WD and slow-WD, suggesting that mRNA expression, by itself, is no indication of the functional involvement of cytokines in WD.
Keywords: TNFα, IL-1α, IL-1β, Wallerian degeneration, regeneration, pain, Schwann cell, fibroblast, macrophage, myelin, phagocytosis
PNS injury is followed by Wallerian degeneration (WD) (Ramon y Cajal, 1928). WD is significant to future regeneration and to the development of neuropathic pain. This is evident from comparing WD in normal C57/BL/6NHSD (C57/BL) and mutant C57/BL/6-WLD/OLA/NHSD (Wld) mice (for review, see Stoll and Muller, 1999). Cellular and molecular events are efficient and rapid (rapid-WD) in C57/BL mice but deficient and slow (slow-WD) in Wld mice. Myelin, which inhibits regeneration (McKerracher et al., 1994;Mukhopadhyay et al., 1994; Schafer et al., 1996; Bandtlow and Schwab, 2000), is removed rapidly during rapid-WD but slowly during slow-WD (Brown et al., 1991; Reichert et al., 1994; Be'eri et al., 1998). NGF, which promotes growth and survival of some PNS neurons (Bibel and Barde, 2000), is injury induced in PNS of C57/BL but not Wld mice (Brown et al., 1991). Delayed myelin removal and deficient NGF production therefore delay regeneration in Wld mice (Brown et al., 1991; Chen and Bisby, 1993; Schafer et al., 1996). NGF and interleukin (IL)-6 regulate neuropathic pain (Safieh-Garabedian et al., 1995; Woolf et al., 1997; Mendell et al., 1999; Murphy et al., 1999). Deficient production of NGF and IL-6 (Reichert et al., 1996) reduces neuropathic pain in Wld mice (Myers et al., 1996).
We view WD as an injury-induced inflammatory response attributable to the production of cytokines, the mediator molecules of inflammation. The classical view of inflammation suggests a network of cytokine production (Oppenhheim and Feldman, 2001) in which inflammatory cytokines TNFα, IL-1α, and IL-1β first upregulate the production of additional inflammatory cytokines and thereafter the production of anti-inflammatory cytokines. Anti-inflammatory cytokines downregulate production altogether. Consequently, inflammation is turned off. We termed the orchestrated production of cytokines during WD “the cytokine network of Wallerian degeneration” (Rotshenker, 1997; Be'eri et al., 1998).
TNFα, IL-1α, and IL-1β are of particular significance to WD. First, they could initiate the cytokine network of WD as they do in other networks of inflammation (Oppenhheim and Feldman, 2001). Second, they contribute to macrophage recruitment to inflammatory sites through endothelial cell activation and chemokine production (Oppenhheim and Feldman, 2001). Likewise, they could contribute to macrophage recruitment in WD and consequently to macrophage-dependent functions (e.g., myelin removal by phagocytosis). Third, they could indirectly regulate survival and growth of PNS neurons and neuropathic pain through the regulation of NGF production in PNS resident fibroblasts [see above and Lindholm et al. (1987) and Hattori et al. (1993,1994)].
Our present study was aimed at studying the production of TNFα, IL-1α, and IL-1β in intact and injured PNS of C57/BL and Wld mice. Transcription, translation, and secretion of cytokines can be differentially regulated such that mRNA expression does not necessarily indicate protein synthesis and synthesis does not necessarily indicate secretion (Dinarello, 2001b). Therefore, we studied mRNA expression and protein synthesis and secretion of TNFα, IL-1α, and IL-1β and further identified the cellular sources of their production, as we did previously with respect to granulocyte macrophage colony stimulating factor (GM-CSF), IL-6, and IL-10 (Reichert et al., 1996; Saada et al., 1996; Rotshenker, 1997; Be'eri et al., 1998). The use of the same animal models in all our studies provides the essential common grounds for elucidating the cytokine network of WD, the orchestrated production of cytokines (TNFα, IL-1α, IL-1β, GM-CSF, IL-6, and IL-10). It further enables unraveling of the intricate relations between the cytokine network and additional events in WD (e.g., NGF production, macrophage recruitment, myelin removal, regeneration, and neuropathic pain) that were also studied in the same animal models.
MATERIALS AND METHODS
Animals and surgical procedures. The experimental protocol was approved by the authority for research and development of the Hebrew University of Jerusalem. Two strains of mice, 2–4 months old, were used. The normal strain, C57/BL/6NHSD (Harlan Sprague Dawley, Jerusalem, Israel), which displays rapid-WD, and the mutant strain, C57/BL/6-WLD/OLA/NHSD (Harlan Olac, Bicester, UK), which displays abnormal slow-WD. We refer to them as C57/BL and Wld, respectively. All surgical procedures were performed under anesthesia. The sciatic nerve was transected after leaving the pelvis. Freeze-damaged PNS segments were obtained by removing 15 mm PNS segments from intact animals and subjecting them to three cycles of freeze–thaw in distilled water to extinguish all resident cells. Freeze-damaged PNS segments were returned in situ to donor animals. Macrophages, and to a lesser extent fibroblasts, are recruited to the freeze-damaged nerve. Freeze-damaged nerve segments serve as Schwann cell free sources from which recruited macrophages are isolated for cell culture studies.
Cell dissociation and culture. Detailed protocols for obtaining nerve-derived non-neuronal cell cultures are given in previous studies (Reichert et al., 1994, 1996; Be'eri et al., 1998). Schwann cells, fibroblast, and macrophages were distinguished from each other by their distinct morphology and immunocytochemistry. Macrophages are positive to F4/80, complement receptor 3 (CR3)/MAC-1, and Galectin-3/MAC-2, Schwann cells are positive for S-100 and Galectin-3/MAC-2 but negative for F4/80, and fibroblasts are negative to all (for review, see Reichert et al., 1994, 1996; Be'eri et al., 1998).
TNF_α, IL-1_α_, and IL-1β immunocytochemistry in tissues._ PNS segments were fixed in 4% buffered formalin (3 hr), washed in PBS, cryoprotected in 30% sucrose (overnight at 4°C), and sectioned in a cryostat. Sections were fixed in 2% neutral formalin, washed in PBS, incubated in 0.05% Triton X-100 in PBS (10 min), blocked in 10% FCS in PBS (3 hr), incubated (overnight at 4°C) in goat anti-mouse TNFα (4 μg/ml), rabbit anti-human IL-1α (2 μg/ml), or goat anti-mouse IL-1β (4 μg/ml), washed in PBS, incubated in biotinylated donkey anti-goat or anti-rabbit IgG (1 hr), washed in PBS, incubated in FITC-conjugated streptavidin (2.5 μg/ml for 45 min), and washed in PBS. TNFα, IL-1α, and IL-1β immunocytochemistry in isolated non-neuronal cells was performed using the same steps with the following exceptions. Cultures were first fixed in 4% buffered formalin at 37°C, incubated or not in 0.05% Triton X-100, and thereafter treated as tissue sections after Triton X-100. Triton X-100 permeabilizes cells, enabling antibodies (Abs) access to the cytoplasm and thereby detection of cytoplasmic molecules. In the absence of Triton X-100, Abs cannot gain access to the cytoplasm, and thus only cell surface molecules are visualized. Normal rabbit or goat IgG was used as negative control. Because there were variations between batches of antibodies, we have always performed “positive controls” simultaneously and side by side with all experimental samples. Positive controls were 3 d injured PNS for tissue samples and macrophages for other non-neuronal cells. The two positive controls displayed consistent positive immunoreactivity to all three cytokines.
Detailed protocols for F4/80, CR3/MAC-1, Galectin-3/MAC-2, and S-100 immunocytochemistry of non-neuronal cells are given and have been shown in previous studies (Reichert et al., 1994, 1996; Be'eri et al., 1998).
TNF_α, IL-1_α_, and IL-1_β_mRNA detection by RT-PCR._ PNS segments were removed from non-operated and operated mice at various time points after injury and rapidly frozen in liquid nitrogen. Total RNA was isolated from the tissues using an RNA isolation kit according to manufacturer's instructions (Biological Industries, Beit-Haemek, Israel). Equal amounts of total RNA were used to synthesize cDNA.
Primer sequences for the PCR reaction were as follows: IL-1α: 5′-CTCTAGAGCACCATGCTACA-3′ and 5′-TGGAATCCAGGGGAAACACTG-5′; IL-1β: 5′-GCAACTGTTCCTGAACTCA-3′ and 5′-CTCGGAGCCTGTAGTGCAG-3′; TNFα: 5′-GGCAGGTCTACTTTGGAGTC-3′ and 5′-ACATTCGAGGCTCCAGTGAATTCGG-3′; β-actin: 5′-CAGCTTCTTTGCAGCTCCTT-3′ and 5′-TCACCCACATAGGAGTCCTT-3′. The number of cycles were 30 for TNFα and IL-1α and 25 for IL-1β. β-actin reaction was used as a positive control and performed together with each of the cytokines at their specific conditions. PCR amplification without Moloney murine leukemia virus ensured that RNA preparations were not contaminated with genomic DNA. To increase reaction specificity, the _EX-Taq_polymerase was added to each tube just before insertion into the PCR cycler, which was preheated to 94°C. PCR amplification products were separated on ethidium bromide-stained 1.5% agarose gel, visualized by ultraviolet light, and photographed. The photographs were scanned, and densitometric analysis was performed using the Bio-Rad Multi Analyst/PC version 1.1. The amplified bands showed their predicted sizes: IL-1α, 312 bp; IL1-β 364 bp; TNFα 301 bp; and β-actin, 219 bp.
Conditioned medium. Intact and injured PNS segments were cut into 5 mm pieces and washed for 10 min in DMEM/F-12. PNS tissues were incubated in DMEM/F-12 (0.15–0.25 ml per nerve) supplemented by 10% heat-inactivated FCS in a humidified incubator saturated by 5% CO2 at 37° C for 5 hr. Conditioned media were separated from PNS tissues by aspiration, followed by 10 min of centrifugation. Each sample of conditioned medium was produced by four distinct nerves. Media and supplements were obtained from Biological Industries.
PNS extract. Ten 10 mm PNS segments were frozen immediately in liquid nitrogen after removal from mice. Thereafter, the frozen PNS segments were extracted in 1 ml PBS containing a mixture of protease inhibitors (Sigma, Rehovot, Israel) and centrifuged for 10 min. Different dilutions of the supernatant were tested for cytokine content by ELISA.
_Identification and quantification of TNF_α _and IL-1_β by ELISA. We used two-site sandwich ELISA to identify and quantify TNFα and IL-1β in conditioned media and PNS extract, according to manufacturer's instructions (Duo-Set; R & D Systems, Minneapolis, MN). Levels of detection of the two cytokines were between 8 and 16 pg/ml.
ELISA assay to study myelin basic protein–myelin phagocytosis. Detailed protocols are given in previous studies (Reichert et al., 2001; Slobodov et al., 2001). In principle, thioglycollate-elicited macrophages were seeded (50 × 103 cells per well) in 96-well tissue culture plates in quadruplicates. Isolated myelin was added for 1 hr to allow phagocytosis, and excess myelin was washed out. Then, levels of phagocytosed myelin were determined by ELISA. The ELISA method is based on the ability to detect myelin basic protein (MBP) in lysate of macrophages that phagocytosed myelin. Because MBP is unique to the structure of myelin, levels of MBP are proportional to levels of myelin. We have determined that >90% of the MBP associated with macrophage lysate is cytoplasmic, resulting from myelin phagocytosis (Slobodov et al., 2001).
Myelin isolation. Myelin was isolated from brains of C57/BL/6NHSD mice (Norton and Poduslo, 1973; Reichert et al., 2001;Slobodov et al., 2001). The resulting isolated myelin preparation is made of fragments of myelin membrane that form small vesicles. Isolated myelin vesicles retain the fundamental ultrastructural and biochemical characteristics of intact membrane myelin. Therefore, levels of MBP are proportional to the amount of myelin vesicles.
ELISA assay to study cell surface CR3/MAC-1. In each experiment, an individual macrophage population that was obtained from a single mouse was seeded in quadruplicate (50 × 103 per well). Macrophages were fixed (4% formalin at 37°C for 30 min), washed, blocked in 3% BSA (overnight at 4°C), washed, incubated in monoclonal antibody (mAb) M1/70 rat anti-mouse CR3/MAC-1 (2 hr at 37°C), washed, incubated in alkaline phosphatase-conjugated donkey anti-rat IgG, washed, and incubated in 100 μl per well substrate (1 mg/ml _p-_nitrophenyl phosphate sodium in 10% diethanolamine, pH 9.8; Sigma), and the reaction product was read in a Dynatec ELISA reader at 405 nm wavelength.
Antibody sources. Antibody sources are as follows: hybridoma cell lines producing mAb M1/70 rat anti-mouse CR3/MAC-1 (Developmental Studies Hybridoma Bank, Iowa City, IA) and rat anti-mouse F4/80 and Galectin-3/MAC-2 (American Type Culture Collection, Rockville, MD); mAb rat anti-mouse MBP and matched isotype control mAbs (Serotec, Oxford, UK); antibodies anti-S-100 and FITC-conjugated rabbit anti-rat and goat anti-rabbit IgG (Sigma); alkaline phosphatase-conjugated goat anti-rat, FITC-conjugated streptavidin, and goat and rabbit IgG (Jackson ImmunoResearch, West Grove, PS); and goat anti-mouse TNFα and IL-1β and rabbit anti-human IL-1α (Santa Cruz Biotechnology, Santa Cruz, CA).
Statistical analysis. One-way ANOVA and the Bonferroni's multiple comparison test were used. All p values of significance are two tailed. Values given are average ± SEM.
RESULTS
TNFα, IL-1α, and IL-1β mRNAs expression in intact and injured PNS of C57/BL mice undergoing rapid-WD
The expression of TNFα, IL-1α, and IL-1β mRNAs was studied in intact and rapid-WD PNS of C57/BL mice by RT-PCR (Fig.1). Tissues were frozen immediately after removal from animals and used thereafter as sources for the detection of all three mRNAs. The detection of mRNA thus indicates in vivo expression. TNFα and IL-1α mRNAs were detected in intact PNS, whereas IL-1β mRNA was not. TNFα and IL-1α mRNAs were further detected in the distal PNS segments undergoing rapid-WD 5 hr and 1, 3, and 6 d after injury. IL-1β mRNA was detected 5 hr and 1 and 3 d but barely at 6 d after injury. Thus TNFα and IL-1α mRNAs were constitutively expressed in intact PNS of C57/BL mice and during the first 6 d of rapid-WD tested. In contrast, IL-1β mRNA expression was injury induced and most probably transient. Furthermore, TNFα and IL-1α mRNAs display a similar pattern of expression, which differs from that of IL-1β mRNA. It is most probable, therefore, that the expression of TNFα and IL-1α mRNAs, on one hand, and IL-1β mRNA, on the other, is differentially regulated in intact PNS of C57/BL mice and during rapid-WD.
Fig. 1.

TNFα and IL-1α mRNAs but not IL-1β mRNA are detected in intact PNS of C57/BL mice (N). TNFα and IL-1α mRNAs are further detected during the first 6 d of rapid-WD studied. IL-1β mRNA is detected 5 hr and 1 and 3 d but barely at 6 d after injury during rapid-WD. Intact (N) and rapid-WD PNS segments were removed 5 hr (5h) and 1, 3, and 6 d (D-1,D-3, and D-6, respectively) after injury. The same tissues were the source for the detection of TNFα, IL-1α, IL-1β, and β-actin mRNAs by RT-PCR. RT-PCR amplification products were separated on ethidium bromide-stained 1.5% agarose gel, visualized by ultraviolet light, and photographed (top panel). The photographs were scanned, and densitometric analysis was performed. Levels of mRNA of each cytokine at each time point were further calculated as percentage of levels of β-actin mRNA in the same tissue sample (bottom panel).
TNFα, IL-1α, and IL-1β protein production in intact and injured PNS of C57/BL mice undergoing rapid-WD
TNFα (Aggawal et al., 2001) and IL-1α (Dinarello, 2001a) protein exert their biological activity as soluble and cell membrane-associated molecules, whereas IL-1β (Dinarello, 2001b) exerts its biological activity as a soluble molecule only. We tested for synthesis and secretion of TNFα, IL-1α, and IL-1β protein by immunocytochemistry, ELISA, and bioassay. Immunocytochemistry in tissue sections visualizes cytoplasmic TNFα, IL-1α, and IL-1β and membrane-associated TNFα and IL-1α. Because immunocytochemistry was performed in PNS segments that were fixed immediately after removal from animals, positive immunoreactivity indicates the in vivo synthesis and association of cytokines with the producing cells. This was further verified by detecting cytokines, by ELISA, in extracts of PNS segments that were frozen immediately after removal from mice. We further tested for the presence of soluble TNFα, IL-1α, and IL-1β in medium conditioned by PNS segments using ELISA and bioassay. The presence of these cytokines in conditioned medium indicates the secretion of synthesized cytokines. Immunocytochemistry, ELISA, and bioassay use different mAbs and Abs to detect each cytokine protein. Therefore, these are two independent assays that we used to test for the synthesis of each cytokine protein. The resulting positive identifications are thus specific and significant.
TNFα in PNS tissue was studied by immunocytochemistry (Fig.2). Low levels of TNFα immunoreactivity were detected occasionally in intact PNS in a small number of Schwann cells. Higher levels of immunoreactivity to TNFα were detected in rapid-WD PNS segments 5 and 12 hr and 1 and 3 d after injury. The_in vivo_ presence of TNFα was further verified in 1 d injured PNS segments that were frozen immediately after removal from mice and then extracted. TNFα content in the extract, determined by ELISA, was 6.53 pg in 5 mg wet weight tissue. Secreted TNFα was studied further. Intact and rapid-WD PNS segments that were removed at various time points after injury were incubated immediately in medium for 5 hr. Conditioned medium thus obtained was tested for the presence of soluble TNFα by ELISA (Fig.3A). Soluble TNFα was detected in medium conditioned at all time points tested. Thus the onset of secretion was rapid, within 5 hr after injury, and continuous, peaking at 1 d after injury.
Fig. 2.
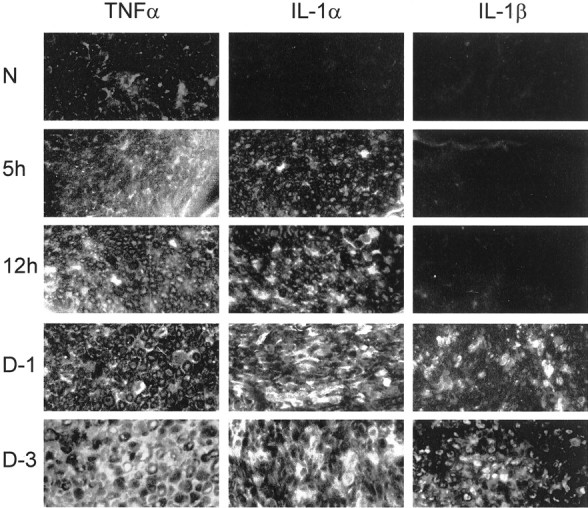
TNFα protein but not IL-1α or IL-1β protein is detected in intact PNS of C57/BL mice (N). TNFα protein is further detected during the first 3 d of rapid-WD studied. IL-1α protein is first detected 5 hr after injury and thereafter during the first 3 d of rapid-WD studied. IL-1β protein is detected 1 and 3 d after injury but not 5 and 12 hr after injury. Cryostat sections of intact (N) and rapid-WD PNS segments that were removed 5 and 12 hr (_5h_and 12h, respectively) and 1 and 3 d (D-1 and D-3, respectively) after injury were used to detect TNFα, IL-1α, and IL-1β protein by immunofluorescence microscopy. Magnification: 250×.
Fig. 3.

TNFα and IL-1β are synthesized and secreted during rapid-WD. The onset of TNFα synthesis and secretion is rapid, within the first 5 hr after PNS injury. The onset of IL-1β synthesis and secretion is delayed, between 5 and 10 hr after PNS injury. PNS segments were removed from non-operated C57/BL mice and used to condition medium for 5 hr (0–5h). Similarly, PNS segments situated distal to transection sites were removed from C57/BL mice 5 hr and 1, 3, 6, and 9 d after injury and used to condition medium for 5 hr (5–10h,D-1, D-3, D-6, and_D-9_, respectively). The same conditioned media were assayed for TNFα and IL-1β content by ELISA, and production levels were calculated (picograms per milligram wet weight of tissue in 5 hr). Bars are the average of three experiments except for time point_D-3_, where six experiments were performed. In each experiment, four different PNS segments were used. Error bars indicate ±1 SEM.
The PNS segments that were incubated in medium for 5 hr were further incubated for an additional 19 hr in fresh medium (5 → 24 hr time period) and conditioned medium assayed for TNFα content by ELISA. Secretion of TNFα was calculated, picograms per milligram wet weight of tissue in 5 hr, to enable comparison with secretion levels during the first 5 hr of incubation (Fig. 3A). Secretion of TNFα by PNS segments removed 1 d after injury averaged 0.18 ± 0.01 pg/mg per 5 hr during the 5 → 24 hr time period compared with 0.49 ± 0.04 pg/mg per 5 hr during the first 5 hr. TNFα was not detected in medium conditioned during the 5 → 24 hr time period by PNS segments removed 0, 3, 6, and 9 d after injury, although it was detected in medium conditioned by the same PNS segments during the first 5 hr of incubation (Fig. 3A). _In vitro_conditions thus impose progressive reduction in secreted TNFα.In vitro imposed progressive reduction, not augmentation, suggests that TNFα secretion during the first 5 hr after removal of injured PNS from animals is most likely a continuation and reflection of cytokine secretion in vivo. This suggestion is supported by the observations that TNFα mRNA and protein are expressed and synthesized in vivo in injured PNS (Figs. 1, 2) (and tissue content by ELISA). Combined, immunocytochemistry and ELISA thus suggest constitutive low levels of TNFα synthesis and transient injury-induced upregulation of TNFα synthesis and secretion.
IL-1α in PNS tissue was studied by immunocytochemistry (Fig. 2). Immunoreactivity to IL-1α was not detected in intact PNS of C57/BL mice. It was detected in rapid-WD PNS segments 5 and 12 hr and 1 and 3 d after injury. We have previously documented the biological activity of soluble IL-1α in medium conditioned within the first 5 hr of rapid-WD and by 7 d rapid-WD PNS segments (Rotshenker et al., 1992). Combined, immunocytochemistry and bioassay suggest no constitutive production but injury-induced rapid onset of synthesis and secretion of soluble IL-1α within 5 hr after injury that continued thereafter. The injury-induced synthesis and secretion of IL-1α protein is further supported by the observations of injury-induced upregulation of IL-1α mRNA expression (Fig. 1). Note, however, that in intact PNS, IL-1α mRNA was detected whereas IL-1α protein was not (see Discussion).
IL-1β in PNS tissue was studied by immunocytochemistry (Fig. 2). Immunoreactivity to IL-1β was not detected in intact PNS of C57/BL mice or in rapid-WD PNS segments 5 and 12 hr after injury. Immunoreactivity to IL-1β was detected in rapid-WD PNS segments 1 and 3 d after injury. The in vivo presence of IL-1β was further verified in 1 d injured PNS segments that were frozen immediately after removal from mice and then extracted. IL-1β content in the PNS extract, determined by ELISA, was 37.69 pg in 5 mg wet weight tissue. We further studied by ELISA the secretion of IL-1β in medium conditioned for 5 hr by PNS segments removed from non-operated mice and at various time points after PNS injury (Fig. 3B). Soluble IL-1β was first detected in medium conditioned 5–10 hr after injury and thereafter, peaking at 1 d after injury.
PNS segments that were incubated in medium for 5 hr were further incubated for an additional 19 hr in fresh medium (5 → 24 hr time period), and conditioned medium was assayed for IL-1β content by ELISA. Secretion of IL-1β was calculated, picograms per milligram wet weight of tissue in 5 hr, to enable comparison with secretion levels during the first 5 hr of incubation (Fig. 3B). Secretion of IL-1β during the 5 → 24 hr time period by PNS segments removed 1 and 3 d after injury averaged, respectively, 0.08 ± 0.007 and 0.28 ± 0.02 pg/mg per 5 hr compared with 0.54 ± 0.11 and 1.72 ± 0.29 pg/mg per 5 hr during the first 5 hr (Fig. 3). IL-1β was not detected in medium conditioned by PNS segments removed from non-operated animals either during the first 5 hr of incubation (Fig. 3) or during the 5 → 24 hr time period. _In vitro_conditions thus impose progressive reduction in secreted IL-1β, as was the case for TNFα (see above). In vitro imposed progressive reduction, not augmentation, suggests that IL-1β secretion during the first 5 hr after removal of injured PNS from animals is most likely a continuation and reflection of cytokine secretion in vivo. Combined, immunocytochemistry and ELISA of conditioned medium suggest no constitutive production of IL-1β and relatively late onset, between 5 and 10 hr after injury, of IL-1β synthesis and secretion that continued thereafter. Injury-induced synthesis and secretion of IL-1β protein is further supported by the detection of injury-induced IL-1β mRNA expression (Fig. 1) and IL-1β protein in injured PNS extract.
Note that low levels of secreted soluble IL-1β were detected by ELISA in medium conditioned during the period of 5–10 hr after injury, whereas IL-1β immunoreactivity was not detected by immunocytochemistry in PNS tissue 12 hr after injury. This discrepancy may be explained by the fact that positive immunoreactivity depends on the accumulation of IL-1β in the cytoplasm to levels detectable by immunocytochemistry. If onset of synthesis is accompanied by rapid secretion, then newly synthesized IL-1β could first reach detectable levels by ELISA in its secreted form in conditioned medium. In contrast, newly synthesized TNFα is first incorporated into the plasma membrane and then released by proteolytic cleavage, and a proportion of newly synthesized IL-1α precursor, which is biologically active, becomes cell membrane associated. It is likely, therefore, that cytoplasmic and membrane-associated TNFα and IL-1α reach detectable levels by immunocytochemistry as fast as the secreted forms reach detectable levels by ELISA and bioassay in conditioned medium.
TNFα, IL-1α, and IL-1β production by Schwann cells, fibroblasts, and macrophages
Schwann cells and fibroblasts compose most of the non-neuronal cell population in intact PNS, whereas macrophages, which are scarce in intact PNS, are recruited in large numbers as of the third day after injury (Perry et al., 1987; Reichert et al., 1994). We established mixed and single-cell type cultures from intact PNS, 3 and 5 d rapid-WD PNS, and 5 d freeze-damaged PNS. Freezing extinguishes all resident non-neuronal cells. Numerous macrophages are recruited into the Schwann cell-free freeze-damaged PNS, which then serve as a source for nerve-derived macrophages. Macrophage cultures were also established from thioglycollate-elicited peritoneal macrophages.
The identification of the non-neuronal cells was based on morphology and immunological markers [reviewed and shown in the same model system in Reichert et al. (1994) and Reichert and Rotshenker (1996, 1999)]: F4/80 is a murine-specific monocyte, macrophage, and microglia marker; Galectin-3/MAC-2 marks in the nervous system-activated myelin phagocytosing macrophages, Schwann cells, and microglia; complement receptor-3 (CR3/MAC-1) is expressed by monocytes, macrophages, and microglia; and S-100 is specific to Schwann cells in peripheral nerves. Macrophages were F4/80, CR3/MAC-1, and Galectin-3/MAC-2 positive. Schwann cells were Galectin-3/MAC-2 and S-100 positive but F4/80 CR3/MAC-1 negative. Schwann cells featured heterogeneous morphology. Some had the “classical” spindle shape, bipolar morphology. Many assumed other shapes that could lead to false identification as macrophages if not for the fact that they were F4/80 and CR3/MAC-1 negative. Fibroblasts were F4/80, CR3/MAC-1, Galectin-3/MAC-2, and S-100 negative.
The cultured non-neuronal cells were studied for TNFα, IL-1α, and IL-1β protein production by immunocytochemistry (Fig. 4) and mRNA expression by RT-PCR (Fig.5). Positive immunoreactivity to TNFα, IL-1α, and IL-1β was detected in Schwann cells that were derived from intact and rapid-WD PNS and in macrophages that were derived from rapid-WD and freeze-damaged PNS. Low levels of immunoreactivity to TNFα, but not to IL-1α or IL-1β, were detected in fibroblasts derived from intact and rapid-WD PNS. Positive immunoreactivity to all cytokines (Fig. 4) was detected after cells were permeabilized by Triton X-100, thereby enabling access of antibodies to the cytoplasm. Immunoreactivity was not detected if Triton X-100 was omitted (data not shown). Immunocytochemistry thus localized cytokines to the cytoplasm and not to the cell surface of cells, as expected from the synthesis of cytokines by these cells. Schwann cells further expressed TNFα, IL-1α, and IL-1β mRNAs (Fig. 5). Thioglycollate-elicited peritoneal macrophages, which express TNFα, IL-1α, and IL-1β, were used as positive controls. They displayed positive immunoreactivity to TNFα, IL-1α, and IL-1β, which were localized to the cytoplasm, and further expressed TNFα, IL-1α, and IL-1β mRNA.
Fig. 4.
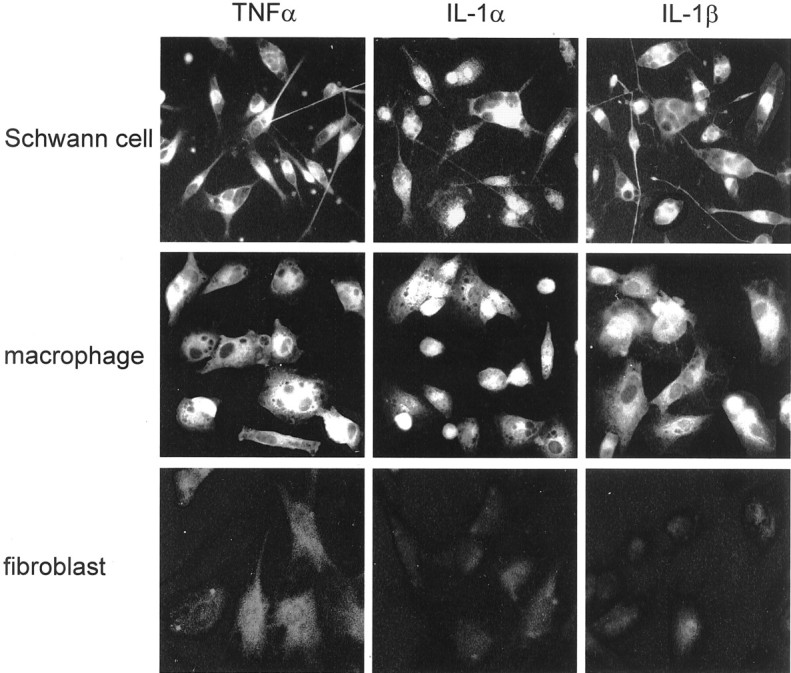
Schwann cells and recruited macrophages produce TNFα, IL-1α, and IL-1β protein. Nerve-derived fibroblasts produce low levels of TNFα but not IL-1α or IL-1β protein. Single-cell type cultures of resident Schwann cells and fibroblasts and recruited macrophages were obtained from intact, rapid-WD, and freeze-damaged PNS of C57/BL mice. The non-neuronal cells were studied for the presence of TNFα, IL-1α, and IL-1β protein by immunofluorescence microscopy. High levels of immunoreactivity for all cytokines were detected in the cytoplasm of Schwann cells and macrophages. In fibroblasts, levels of immunoreactivity to TNFα were slightly above control levels, whereas levels of immunoreactivity to IL-1α and IL-1β were the same as for control. Immunoreactivity of cytokines was localized to the cytoplasm and not cell surfaces because immunoreactivity was detected in cells permeabilized by Triton X-100 but not in non-permeabilized cells (data not shown). Magnification: 400×.
Fig. 5.

TNFα and IL-1α and IL-1β mRNA are detected in Schwann cells and macrophages. Schwann cell cultures were obtained from rapid-WD PNS of C57/BL mice, as in Figure 4, and macrophages were thioglycollate elicited. Schwann cells and macrophages were used as source for the detection of TNFα, IL-1α, IL-1β, and β-actin mRNAs by RT-PCR. RT-PCR amplification products were separated on ethidium bromide-stained 1.5% agarose gel, visualized by ultraviolet light, and photographed. The photographs were scanned, and densitometric analysis was performed. Levels of mRNA of each cytokine were further calculated as percentage of levels of β-actin mRNA in the same cell sample.
TNFα, IL-1α, and IL-1β mRNA expression and protein production in intact and injured PNS of Wld mice undergoing slow-WD
In mutant Wld mice, the PNS segment situated distal to an injury site can be divided into two domains: the injury domain, which is <1 mm in length and situated immediately after the injury site, and the remainder more distally located domain that makes most of the distal PNS segment. The injury domain undergoes rapid-WD, whereas the remainder more distally located domain undergoes slow-WD (Reichert et al., 1994).
The expression of TNFα, IL-1α, and IL-1β mRNAs was studied by RT-PCR in intact and slow-WD Wld mice PNS 3 d after injury. TNFα and IL-1α mRNAs were detected in intact PNS, whereas IL-1β mRNA was not. The mRNAs of all three cytokines were detected in the slow-WD domains 3 d after injury (Fig.6).
Fig. 6.

TNFα and IL-1α mRNAs but not IL-1β mRNA are detected in intact PNS of Wld mice (N). TNFα, IL-1α, and IL-1β mRNAs are detected in slow-WD domains of Wld mice 3 d after injury (D-3). Intact PNS segments and slow-WD PNS domains, 3 d after injury, were removed from Wld mice. The same tissues were the source for the detection of TNFα, IL-1α, IL-1β, and β-actin mRNAs by RT-PCR. RT-PCR amplification products were separated on ethidium bromide-stained 1.5% agarose gel, visualized by ultraviolet light, and photographed. The photographs were scanned, and densitometric analysis was performed. Levels of mRNA of each cytokine at each time point were further calculated as percentage of levels of β-actin mRNA in the same tissue sample.
The production of TNFα, IL-1α, and IL-1β protein was studied by immunocytochemistry in intact and the two domains of injured Wld mice PNS, the injury domains, and the slow-WD domains 3 d after injury. Immunoreactivity to TNFα, IL-1α, and IL-1β was not detected in intact PNS (data not shown). Immunoreactivity to the three cytokines was detected 3 d after injury in the injury domains but not in the slow-WD domains (Fig. 7). It should be noted that each injury domain and mate slow-WD domain, taken from the same injured PNS, were processed simultaneously side by side for immunocytochemistry. The discrepancy between injury domains and slow-WD domains of the same PNS nerve segments is thus significant, suggesting extremely low levels or no production of TNFα, IL-1α, and IL-1β proteins in slow-WD domains.
Fig. 7.

TNFα, IL-1α, and IL-1β protein are detected in injury domains but not slow-WD domains of Wld mice 3 d after injury. The PNS segment, which is situated distal to the injury site, was removed 3 d after injury, and cryostat sections were taken from the injury domain and its mate slow-WD domain. These were then processed simultaneously side by side for the detection of TNFα, IL-1α, and IL-1β protein by immunofluorescence microscopy. Magnification: 250×.
TNFα and IL-1β augment myelin phagocytosis by macrophages
One of the hallmarks of Wallerian degeneration is the removal, by phagocytosis, of myelin that otherwise may inhibit regeneration. We have previously documented the rapid and efficient removal of myelin during rapid-WD by resident Schwann cells and recruited macrophages (Reichert et al., 1994; Be'eri et al., 1998). Complement receptor-3 (CR3/MAC-1) plays a major role in mediating myelin phagocytosis by macrophages (Bruck and Friede, 1990; Mosley and Cuzner, 1996; van der Laan et al., 1996; Reichert et al., 2001; Slobodov et al., 2001). There are conflicting reports regarding the effect of TNFα on myelin phagocytosis by macrophages. In one (Bruck et al., 1992), TNFα inhibited myelin phagocytosis by downregulating cell surface expression of CR3/MAC-1. In others (Smith et al., 1998; Liefner et al., 2000), TNFα had no effect on myelin phagocytosis by macrophages but augmented myelin phagocytosis by microglia.
We studied the effect of TNFα and IL-1β on myelin phagocytosis and cell surface expression of CR3/MAC-1 in macrophages. Thioglycollate-elicited peritoneal macrophages were incubated in medium in the absence or presence of either recombinant mouse TNFα (1 ng/ml) or IL-1β (0.6 ng/ml) for 36 hr. Cultures were then studied for myelin phagocytosis, cell surface levels of CR3/MAC-1, and cell number (Fig.8). In each experiment, an individual macrophage population that was obtained from a single mouse was tested for myelin phagocytosis in quadruplicate. Myelin phagocytosis was quantified by determining levels of MBP in lysate of macrophages by ELISA and further calculated as percentage of phagocytosis by macrophages that were not treated by the cytokines. Myelin phagocytosis was augmented significantly by TNFα (120.85 ± 5.38% of normal;p < 0.01) and by IL-1β (129.17 ± 6.18% of normal; p < 0.001). Duplicate cultures were used to study cell surface levels of CR3/MAC-1 by ELISA. The same cultures were further used to quantify cell number by counting cells in a sample area in the center of wells. Cell surface levels of CR3/MAC-1 and cell number were further calculated as percentage of their values in experiments in which macrophages were not treated by the cytokines. Cell surface levels of CR3/MAC-1 and cell number did not differ from normal. Thus the augmentation of myelin phagocytosis was not the result of either TNFα- or IL-1β-induced upregulation of cell surface levels of CR3/MAC-1 or macrophage proliferation.
Fig. 8.

TNFα and IL-1β augment myelin phagocytosis by macrophages. Thioglycollate-elicited peritoneal macrophages were incubated in the absence or presence of TNFα (1 ng/ml) or IL-1β (0.6 ng/ml) for 36 hr. In each experiment, a macrophage population from a different mouse was studied. Thereafter, myelin phagocytosis, cell surface levels of CR3/MAC-1, and cell number (#) were quantified and further calculated as percentage of normal. Values obtained in the absence of cytokines were defined 100% normal. Bars are the average of six experiments, each performed in quadruplicate. Error bars indicate ±1 SEM.
DICUSSION
We presently document the involvement of TNFα, IL-1α, and IL-1β during normal WD. TNFα and IL-1α are likely to be the first cytokines the protein production of which is upregulated in Schwann cells after PNS injury. The production of IL-1β protein is induced in Schwann cells with a delay after injury. TNFα and IL-1α therefore may play a role in the initiation of the cytokine network of WD. TNFα, IL-1α, and IL-1β can further regulate additional molecular and cellular events in WD. In agreement with and support of this notion is the deficient production of the three cytokines during slow-WD.
Transcription, translation, and secretion of cytokines can be differentially regulated such that mRNA expression does not necessarily indicate protein synthesis and synthesis does not necessarily indicate secretion (Dinarello, 2001b). Therefore, we examined TNFα, IL-1α, and IL-1β mRNA expression (RT-PCR), protein synthesis (immunocytochemistry and ELISA), and protein synthesis and secretion (ELISA and bioassay of conditioned medium) and further identified the cellular sources of their production. The functional involvement of any of the cytokines is indicated by combining positive independent observations on mRNA expression, protein synthesis, and protein secretion and not by just one observation. Each observation complements and verifies the others.
TNFα mRNA and protein are constitutively expressed and synthesized at low levels in Schwann cells that reside in intact PNS of C57/BL mice. Injury induces the rapid upregulation of TNFα mRNA expression and protein synthesis and secretion during rapid-WD. This notion stems from integrating observations on the detection of TNFα mRNA and protein in intact PNS and during rapid-WD, the detection of secreted TNFα within the first 5 hr after PNS injury and thereafter, and the detection of TNFα mRNA and protein in Schwann cells. These findings, along with the fact that Schwann cells are in intimate contact with axons that they ensheathe, suggest that Schwann cells may be the first among the non-neuronal cells to “sense” and respond to axonal injury by secreting already existing constitutively synthesized TNFα protein. Then, Schwann cells further synthesize and secrete TNFα protein using constitutively and newly injury-induced expressed mRNA. Our observations and conclusions agree with and further extend previous findings: injury-induced mRNA expression of TNFα in PNS of mice (La Fleur et al., 1996), constitutive and injury-induced upregulation of TNFα mRNA in rat PNS (Taskinen et al., 2000), and constitutive and injury-induced upregulation of TNFα protein content in rat PNS and Schwann cells (Wagner and Myers, 1996b; George et al., 1999); however, see injury-induced but not constitutive TNFα mRNA expression in rat PNS (Wagner and Myers, 1996b) and injury-induced but not constitutive TNFα protein content in rat PNS (Stoll et al., 1993).
IL-1α mRNA is constitutively expressed but IL-1α protein is not synthesized in intact PNS of C57/BL mice. Injury induces the rapid upregulation of IL-1α mRNA expression and IL-1α protein synthesis and secretion in Schwann cells during rapid-WD. This conclusion is based on integrating the detection of IL-1α mRNA but not protein in intact PNS, the detection of IL-1α mRNA during rapid-WD, the detection of synthesized and secreted IL-1α within the first 5 hr after PNS injury and thereafter (Rotshenker et al., 1992), and the detection of IL-1α mRNA and protein in Schwann cells. The observations in intact PNS that TNFα and IL-1α mRNAs are expressed and TNFα but not IL-1α protein is synthesized suggest that injury-induced IL-1α protein production may lag after TNFα protein production. Our observations agree with and further extend previous findings on constitutive expression of IL-1α mRNA in intact rat PNS (Perry et al., 1987) and injury-induced IL-1α mRNA expression in injured mouse PNS (La Fleur et al., 1996).
IL-1β mRNA is not expressed and IL-1β protein is not synthesized in intact PNS of C57/BL mice. Injury induces the rapid expression of IL-1β mRNA but the delayed synthesis and secretion of IL-1β protein in Schwann cells. This notion arises from integrating the inability to detect IL-1β mRNA and protein in intact PNS, the detection of IL-1β mRNA 5 hr and 1 and 3 d after PNS injury, and the detection of synthesized and secreted IL-1β protein between 5 and 10 hr but not within the first 5 hr of rapid-WD. Constitutive and injury-induced upregulation of IL-1β mRNA expression was reported in rat PNS (Gillen et al., 1998).
Macrophages, which are recruited to the injured nerves as of the third day of WD, may further contribute to TNFα, IL-1α, and IL-1β production.
In contrast to rapid-WD, mRNA expression but not protein production of TNFα, IL-1α, and IL-1β is upregulated during slow-WD in Wld mice. This contrast between efficient versus deficient protein production of TNFα, IL-1α, and IL-1β during rapid-WD and slow-WD, respectively, follows a similar pattern of efficient versus deficient production of GM-CSF, IL-6, and IL-10 that we reported previously in these PNS tissues (Saada et al., 1996; Reichert et al., 1996; Be'eri et al., 1998). This overall efficient versus deficient injury-induced cytokine production points to the inflammatory nature of WD and further suggests that cytokines play significant roles in regulating molecular and cellular events in WD. This notion is supported by a number of observations. GM-CSF, IL-6, and IL-10 are produced efficiently in rapid-WD but not slow-WD, which is in accord with TNFα, IL-1α, and IL-1β inducting GM-CSF, IL-6, and IL-10 production (Aggawal et al., 2001; Dinarello, 2001a,b). Schwann cells are activated during rapid-WD but not slow-WD (Reichert et al., 1994), which is in agreement with the efficient production of GM-CSF that activates Schwann cells during rapid-WD but not slow-WD (Saada et al., 1996). Macrophages are recruited and activated during rapid-WD but not slow-WD (Perry et al., 1987; Brown et al., 1991; Reichert et al., 1994), which is in accord with the role of TNFα, IL-1α, and IL-1β in the recruitment of macrophages to sites of inflammation (Liefner et al., 2000;Oppenhheim and Feldman, 2001; Ousman and David, 2001; Subang and Richardson, 2001). Deficient macrophage recruitment and deficient Schwann cell activation result, in turn, in delayed myelin removal by phagocytosis during slow-WD (Brown et al., 1991; Reichert et al., 1994;Be'eri et al., 1998). NGF production is upregulated during rapid-WD but not slow-WD (Brown et al., 1991), which is in accord with TNFα, IL-1α, and IL-1β inducing NGF production in fibroblasts (Lindholm et al., 1987; Hattori et al., 1993; Hattori et al., 1994). In turn, deficient NGF production combined with delayed myelin removal contributes to delayed regeneration in Wld mice in which slow-WD occurs (Brown et al., 1991; Chen and Bisby, 1993; Schafer et al., 1996). Furthermore, deficient production of NGF and IL-6 during slow-WD contributes to the reduction of neuropathic pain after PNS injury in Wld mice (Safieh-Garabedian et al., 1995; Myers et al., 1996; Wagner and Myers, 1996a; Woolf et al., 1997; Mendell et al., 1999; Murphy et al., 1999).
TNFα, IL-1α, and IL-1β mRNA expression and protein production may be differentially regulated during WD. IL-1α mRNA was detected in intact PNS of C57/BL mice, whereas IL-1α protein was not. IL-1β mRNA was detected within the first 5 hr of rapid-WD, whereas synthesis and secretion of IL-1β protein were not. TNFα, IL-1α, and IL-1β mRNAs were detected during slow-WD in injured PNS of Wld mice, whereas TNFα, IL-1α, and IL-1β protein were not. The inability to detect cytokine protein by immunocytochemistry and ELISA can reflect either no production or extremely low levels of production that are likely to be functionally insignificant. This differential regulation of mRNA expression and protein production suggests that mRNA expression, by itself, is no indication of the functional involvement of TNFα, IL-1α, or IL-1β in WD. Similar observations have been made previously in other cells and tissues (for review, see Dinarello, 2001b).
We have previously studied mRNA expression, the time course of protein synthesis and secretion, and the identity of the non-neuronal cells producing IL-6, GM-CSF, and IL-10 during rapid-WD and slow-WD (Reichert et al., 1996; Saada et al., 1996; Be'eri et al., 1998) as we have presently done for TNFα, IL-1α, and IL-1β. We further studied macrophage recruitment and activation, Schwann cell activation, and myelin removal by phagocytosis. These studies were all performed using the same animal and injury models, which provide the required common grounds for comparison and integration of data. These observations suggest a network of cytokine production that is orchestrated in time and magnitude: the cytokine network of Wallerian degeneration (Fig.9). Timing and magnitude of cytokine production are determined to a large extent by the non-neuronal cell types producing them, the spatial distribution of non-neuronal cells in the PNS tissue, and the timing of macrophage recruitment. In intact PNS, Schwann cells form close contact with axons that they ensheathe, whereas fibroblasts are scattered between nerve fibers. Macrophages, which are scarce in intact PNS, are recruited as of the third day after injury but reach large numbers within the next few days. TNFα and IL-1α are most likely the first inflammatory cytokines to be produced in rapid-WD by Schwann cells, which are the first to sense and respond to axotomy because of their intimate contact with axons (see above). Then follows the production of IL-6 by fibroblasts, within 2 hr after injury, and GM-CSF, within 4 hr after injury (Reichert et al., 1996;Saada et al., 1996). Fibroblasts do not express or produce IL-6 and GM-CSF mRNA or protein in intact PNS. IL-6 and GM-CSF production can be induced by diffusible TNFα and IL-1α produced by Schwann cells (Saada et al., 1996). The onset of IL-1β production by Schwann cells is 5–10 hr after injury. IL-1β can further contribute to IL-6 and GM-CSF production. IL-6, which inhibits TNFα production (Aggawal et al., 2001), may be responsible for the reduction in TNFα production after the first day of rapid-WD. TNFα, IL-1α, and IL-1β produced by Schwann cells can further induce recruited macrophages to produce mostly IL-6 (Reichert et al., 1996) but also TNFα, IL-1α, and IL-1β. TNFα, IL-1α, and IL-1β induce IL-10 production (de Waal Malefyt, 2001). Indeed, the onset of IL-10 production by fibroblasts is rapid (Be'eri et al., 1998), but levels of production are low and insignificant because fibroblasts are poor producers of IL-10. The timing of high levels of IL-10 production, after the fourth day of rapid-WD, is determined by the timing of macrophage recruitment, which produces large amounts of IL-10. IL-10 then downregulates the production of the inflammatory cytokines and itself (Be'eri et al., 1998; de Waal Malefyt, 2001), thereby downregulating WD.
Fig. 9.

The cytokine network of Wallerian degeneration. The cellular elements depicted are a resident Schwann cell ensheathing an axon, a resident fibroblast, and a recruited macrophage.Solid lines represent induction and upregulation, and_dotted lines_ represent downregulation of the production of cytokine protein. Axotomy induces the production of TNFα and IL-1α in resident Schwann cells first. Thereafter, IL-6 and GM-CSF are produced in resident fibroblasts; then follows IL-1β production in resident Schwann cells. Macrophages, which are recruited as of the third day after injury, produce the inflammatory cytokines TNFα, IL-1α, IL-1β, and IL-6 and the anti-inflammatory cytokine IL-10. IL-10 downregulates the production of all inflammatory cytokines and itself in all non-neuronal cells. Not shown in this diagram are low, functionally insignificant levels of IL-10, which are produced by fibroblasts, and the ability of IL-6 to downregulate TNFα production (see Discussion).
TNFα and IL-1α most probably play a critical role in setting the cytokine network in motion under normal conditions (Fig. 9), as they do in other inflammatory conditions and disease states (Oppenhheim and Feldman, 2001). Their deficient injury-induced production in Wld mice thus offers one possible explanation for the deficient production of the additional cytokines as well as for the deficient progression of other molecular and cellular events of WD that depend on these cytokines (see above).
Footnotes
This research was supported by Grant 96-00211 from the U.S.–Israel Binational Science Foundation, Jerusalem, Israel. We acknowledge the collaboration of Dr. E. M. Shooter in this project.
Correspondence should be addressed to Shlomo Rotshenker, Department of Anatomy and Cell Biology, Hebrew University–Hadassah Medical School, P.O. 12272, Jerusalem 91120, Israel. E-mail:ROTSH@md.huji.ac.il.
REFERENCES
- 1.Aggawal BB, Samanta A, Feldman M. TNFα. In: Oppenhheim JJ, Feldman M, editors. Cytokine reference. Academic; New York: 2001. pp. 413–434. [Google Scholar]
- 2.Bandtlow CE, Schwab ME. NI-35/250/nogo-a: a neurite growth inhibitor restricting structural plasticity and regeneration of nerve fibers in the adult vertebrate CNS. Glia. 2000;29:175–181. doi: 10.1002/(sici)1098-1136(20000115)29:2<175::aid-glia11>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- 3.Be'eri H, Reichert F, Saada A, Rotshenker S. The cytokine network of wallerian degeneration: IL-10 and GM-CSF. Eur J Neurosci. 1998;10:2707–2713. [PubMed] [Google Scholar]
- 4.Bibel M, Barde YA. Neurotrophins: key regulators of cell fate and cell shape in the vertebrate nervous system. Genes Dev. 2000;14:2919–2937. doi: 10.1101/gad.841400. [DOI] [PubMed] [Google Scholar]
- 5.Brown MC, Perry VH, Lunn ER, Gordon S, Heumann R. Macrophage dependence of peripheral sensory nerve regeneration: possible involvement of nerve growth factor. Neuron. 1991;6:359–370. doi: 10.1016/0896-6273(91)90245-u. [DOI] [PubMed] [Google Scholar]
- 6.Bruck W, Friede RL. Anti-macrophage CR3 antibody blocks myelin phagocytosis by macrophages in vitro. Acta Neuropathol (Berl) 1990;80:415–418. doi: 10.1007/BF00307696. [DOI] [PubMed] [Google Scholar]
- 7.Bruck W, Bruck Y, Friede RL. TNF-alpha suppresses CR3-mediated myelin removal by macrophages. J Neuroimmunol. 1992;38:9–17. doi: 10.1016/0165-5728(92)90085-y. [DOI] [PubMed] [Google Scholar]
- 8.Chen S, Bisby MA. Impaired motor axon regeneration in the C57BL/Ola mouse. J Comp Neurol. 1993;333:449–454. doi: 10.1002/cne.903330310. [DOI] [PubMed] [Google Scholar]
- 9.de Waal Malefyt R. IL-10. In: Oppenhheim JJ, Feldman M, editors. Cytokine reference. Academic; New York: 2001. pp. 165–185. [Google Scholar]
- 10.Dinarello CA. IL-1α. In: Oppenhheim JJ, Feldman M, editors. Cytokine reference. Academic; New York: 2001a. pp. 307–318. [Google Scholar]
- 11.Dinarello CA. IL-1β. In: Oppenhheim JJ, Feldman M, editors. Cytokine reference. Academic; New York: 2001b. pp. 351–374. [Google Scholar]
- 12.George A, Schmidt C, Weishaupt A, Toyka KV, Sommer C. Serial determination of tumor necrosis factor-alpha content in rat sciatic nerve after chronic constriction injury. Exp Neurol. 1999;160:124–132. doi: 10.1006/exnr.1999.7193. [DOI] [PubMed] [Google Scholar]
- 13.Gillen C, Jander S, Stoll G. Sequential expression of mRNA for proinflammatory cytokines and interleukin-10 in the rat peripheral nervous system: comparison between immune-mediated demyelination and Wallerian degeneration. J Neurosci Res. 1998;51:489–496. doi: 10.1002/(SICI)1097-4547(19980215)51:4<489::AID-JNR8>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 14.Hattori A, Tanaka E, Murase K, Ishida N, Chatani Y, Tsujimoto M, Hayashi K, Kohno M. Tumor necrosis factor stimulates the synthesis and secretion of biologically active nerve growth factor in non-neuronal cells. J Biol Chem. 1993;268:2577–2582. [PubMed] [Google Scholar]
- 15.Hattori A, Iwasaki S, Murase K, Tsujimoto M, Sato M, Hayashi K, Kohno M. Tumor necrosis factor is markedly synergistic with interleukin 1 and interferon-gamma in stimulating the production of nerve growth factor in fibroblasts. FEBS Lett. 1994;340:177–180. doi: 10.1016/0014-5793(94)80132-0. [DOI] [PubMed] [Google Scholar]
- 16.La Fleur M, Underwood JL, Rappolee DA, Werb Z. Basement membrane and repair of injury to peripheral nerve: defining a potential role for macrophages, matrix metalloproteinases, and tissue inhibitor of metalloproteinases-1. J Exp Med. 1996;184:2311–2326. doi: 10.1084/jem.184.6.2311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liefner M, Siebert H, Sachse T, Michel U, Kollias G, Bruck W. The role of TNF-alpha during Wallerian degeneration. J Neuroimmunol. 2000;108:147–152. doi: 10.1016/s0165-5728(00)00262-9. [DOI] [PubMed] [Google Scholar]
- 18.Lindholm D, Heumann R, Meyer M, Thoenen H. Interleukin-1 regulates synthesis of nerve growth factor in non-neuronal cells of rat sciatic nerve. Nature. 1987;330:658–659. doi: 10.1038/330658a0. [DOI] [PubMed] [Google Scholar]
- 19.McKerracher L, David S, Jackson DL, Kottis V, Dunn RJ, Braun PE. Identification of myelin-associated glycoprotein as a major myelin-derived inhibitor of neurite growth. Neuron. 1994;13:805–811. doi: 10.1016/0896-6273(94)90247-x. [DOI] [PubMed] [Google Scholar]
- 20.Mendell LM, Albers KM, Davis BM. Neurotrophins, nociceptors, and pain. Microsc Res Tech. 1999;45:252–261. doi: 10.1002/(SICI)1097-0029(19990515/01)45:4/5<252::AID-JEMT9>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 21.Mosley K, Cuzner ML. Receptor-mediated phagocytosis of myelin by macrophages and microglia: effect of opsonization and receptor blocking agents. Neurochem Res. 1996;21:481–487. doi: 10.1007/BF02527713. [DOI] [PubMed] [Google Scholar]
- 22.Mukhopadhyay G, Doherty P, Walsh FS, Crocker PR, Filbin MT. A novel role for myelin-associated glycoprotein as an inhibitor of axonal regeneration. Neuron. 1994;13:757–767. doi: 10.1016/0896-6273(94)90042-6. [DOI] [PubMed] [Google Scholar]
- 23.Murphy PG, Ramer MS, Borthwick L, Gauldie J, Richardson PM, Bisby MA. Endogenous interleukin-6 contributes to hypersensitivity to cutaneous stimuli and changes in neuropeptides associated with chronic nerve constriction in mice. Eur J Neurosci. 1999;11:2243–2253. doi: 10.1046/j.1460-9568.1999.00641.x. [DOI] [PubMed] [Google Scholar]
- 24.Myers RR, Heckman HM, Rodriguez M. Reduced hyperalgesia in nerve-injured WLD mice: relationship to nerve fiber phagocytosis, axonal degeneration, and regeneration in normal mice. Exp Neurol. 1996;141:94–101. doi: 10.1006/exnr.1996.0142. [DOI] [PubMed] [Google Scholar]
- 25.Norton WT, Poduslo SE. Myelination in rat brain: changes in myelin composition during brain maturation. J Neurochem. 1973;21:759–773. doi: 10.1111/j.1471-4159.1973.tb07520.x. [DOI] [PubMed] [Google Scholar]
- 26.Oppenhheim JJ, Feldman M. Introduction to the role of cytokines in innate and defense and adaptive immunity. In: Oppenhheim JJ, Feldman M, editors. Cytokine reference. Academic; New York: 2001. pp. 3–20. [Google Scholar]
- 27.Ousman SS, David S. MIP-1α, MCP-1, GM-CSF, and TNF-α control the immune cell response that mediates rapid phagocytosis of myelin from the adult mouse spinal cord. J Neurosci. 2001;21:4649–4656. doi: 10.1523/JNEUROSCI.21-13-04649.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Perry VH, Brown MC, Gordon S. The macrophage response to central and peripheral nerve injury. A possible role for macrophages in regeneration. J Exp Med. 1987;165:1218–1223. doi: 10.1084/jem.165.4.1218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ramon y Cajal S. Degeneration and regeneration of the nervous system. Hafner; London: 1928. [Google Scholar]
- 30.Reichert F, Rotshenker S. Deficient activation of microglia during optic nerve degeneration. J Neuroimmunol. 1996;70:153–161. doi: 10.1016/s0165-5728(96)00112-9. [DOI] [PubMed] [Google Scholar]
- 31.Reichert F, Rotshenker S. Galectin-3/MAC-2 in experimental allergic encephalomyelitis. Exp Neurol. 1999;160:508–514. doi: 10.1006/exnr.1999.7229. [DOI] [PubMed] [Google Scholar]
- 32.Reichert F, Saada A, Rotshenker S. Peripheral nerve injury induces Schwann cells to express two macrophage phenotypes: phagocytosis and the galactose-specific lectin MAC-2. J Neurosci. 1994;14:3231–3245. doi: 10.1523/JNEUROSCI.14-05-03231.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Reichert F, Levitzky R, Rotshenker S. Interleukin 6 in intact and injured mouse peripheral nerves. Eur J Neurosci. 1996;8:530–535. doi: 10.1111/j.1460-9568.1996.tb01237.x. [DOI] [PubMed] [Google Scholar]
- 34.Reichert F, Slobodov U, Makranz C, Rotshenker S. Modulation (inhibition and augmentation) of complement receptor-3-mediated myelin phagocytosis. Neurobiol Dis. 2001;8:504–512. doi: 10.1006/nbdi.2001.0383. [DOI] [PubMed] [Google Scholar]
- 35.Rotshenker S. The cytokine network of Wallerian degeneration. Curr Topics Neurochem. 1997;1:147–156. [Google Scholar]
- 36.Rotshenker S, Aamar S, Barak V. Interleukin-1 activity in lesioned peripheral nerve. J Neuroimmunol. 1992;39:75–80. doi: 10.1016/0165-5728(92)90176-l. [DOI] [PubMed] [Google Scholar]
- 37.Saada A, Reichert F, Rotshenker S. Granulocyte macrophage colony stimulating factor produced in lesioned peripheral nerves induces the up-regulation of cell surface expression of MAC-2 by macrophages and Schwann cells. J Cell Biol. 1996;133:159–167. doi: 10.1083/jcb.133.1.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Safieh-Garabedian B, Poole S, Allchorne A, Winter J, Woolf CJ. Contribution of interleukin-1 beta to the inflammation-induced increase in nerve growth factor levels and inflammatory hyperalgesia. Br J Pharmacol. 1995;115:1265–1275. doi: 10.1111/j.1476-5381.1995.tb15035.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Schafer M, Fruttiger M, Montag D, Schachner M, Martini R. Disruption of the gene for the myelin-associated glycoprotein improves axonal regrowth along myelin in C57BL/Wlds mice. Neuron. 1996;16:1107–1113. doi: 10.1016/s0896-6273(00)80137-3. [DOI] [PubMed] [Google Scholar]
- 40.Slobodov U, Reichert F, Mirski R, Rotshenker S. Distinct inflammatory stimuli induce different patterns of myelin phagocytosis and degradation in recruited macrophages. Exp Neurol. 2001;167:401–409. doi: 10.1006/exnr.2000.7559. [DOI] [PubMed] [Google Scholar]
- 41.Smith ME, van der Maesen K, Somera FP. Macrophage and microglial responses to cytokines in vitro: phagocytic activity, proteolytic enzyme release, and free radical production. J Neurosci Res. 1998;54:68–78. doi: 10.1002/(SICI)1097-4547(19981001)54:1<68::AID-JNR8>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 42.Stoll G, Muller HW. Nerve injury, axonal degeneration and neural regeneration: basic insights. Brain Pathol. 1999;9:313–325. doi: 10.1111/j.1750-3639.1999.tb00229.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Stoll G, Jung S, Jander S, van der MP, Hartung HP. Tumor necrosis factor-alpha in immune-mediated demyelination and Wallerian degeneration of the rat peripheral nervous system. J Neuroimmunol. 1993;45:175–182. doi: 10.1016/0165-5728(93)90178-2. [DOI] [PubMed] [Google Scholar]
- 44.Subang MC, Richardson PM. Influence of injury and cytokines on synthesis of monocyte chemoattractant protein-1 mRNA in peripheral nervous tissue. Eur J Neurosci. 2001;13:521–528. doi: 10.1046/j.1460-9568.2001.01425.x. [DOI] [PubMed] [Google Scholar]
- 45.Taskinen HS, Olsson T, Bucht A, Khademi M, Svelander L, Roytta M. Peripheral nerve injury induces endoneurial expression of IFN-gamma, IL- 10 and TNF-alpha mRNA. J Neuroimmunol. 2000;102:17–25. doi: 10.1016/s0165-5728(99)00154-x. [DOI] [PubMed] [Google Scholar]
- 46.van der Laan LJ, Ruuls SR, Weber KS, Lodder IJ, Dopp EA, Dijkstra CD. Macrophage phagocytosis of myelin in vitro determined by flow cytometry: phagocytosis is mediated by CR3 and induces production of tumor necrosis factor-alpha and nitric oxide. J Neuroimmunol. 1996;70:145–152. doi: 10.1016/s0165-5728(96)00110-5. [DOI] [PubMed] [Google Scholar]
- 47.Wagner R, Myers RR. Endoneurial injection of TNF-alpha produces neuropathic pain behaviors. NeuroReport. 1996a;7:2897–2901. doi: 10.1097/00001756-199611250-00018. [DOI] [PubMed] [Google Scholar]
- 48.Wagner R, Myers RR. Schwann cells produce tumor necrosis factor alpha: expression in injured and non-injured nerves. Neuroscience. 1996b;73:625–629. doi: 10.1016/0306-4522(96)00127-3. [DOI] [PubMed] [Google Scholar]
- 49.Woolf CJ, Allchorne A, Safieh-Garabedian B, Poole S. Cytokines, nerve growth factor and inflammatory hyperalgesia: the contribution of tumour necrosis factor alpha. Br J Pharmacol. 1997;121:417–424. doi: 10.1038/sj.bjp.0701148. [DOI] [PMC free article] [PubMed] [Google Scholar]