INCENP Centromere and Spindle Targeting: Identification of Essential Conserved Motifs and Involvement of Heterochromatin Protein HP1 (original) (raw)
Abstract
The inner centromere protein (INCENP) has a modular organization, with domains required for chromosomal and cytoskeletal functions concentrated near the amino and carboxyl termini, respectively. In this study we have identified an autonomous centromere- and midbody-targeting module in the amino-terminal 68 amino acids of INCENP. Within this module, we have identified two evolutionarily conserved amino acid sequence motifs: a 13–amino acid motif that is required for targeting to centromeres and transfer to the spindle, and an 11–amino acid motif that is required for transfer to the spindle by molecules that have targeted previously to the centromere. To begin to understand the mechanisms of INCENP function in mitosis, we have performed a yeast two-hybrid screen for interacting proteins. These and subsequent in vitro binding experiments identify a physical interaction between INCENP and heterochromatin protein HP1Hsα. Surprisingly, this interaction does not appear to be involved in targeting INCENP to the centromeric heterochromatin, but may instead have a role in its transfer from the chromosomes to the anaphase spindle.
Keywords: centromeres, heterochromatin, HP1, INCENP, mitosis
Centromeres act at a number of levels to direct chromosome segregation in mitosis and meiosis (Pluta et al., 1995). The kinetochore is responsible for many centromere functions, including microtubule binding, motor activity (Gorbsky et al., 1987; Nicklas, 1989), and cell cycle signaling (Campbell and Gorbsky, 1995; Rieder et al., 1995; Chen et al., 1996; Taylor and McKeon, 1997). The centromeric heterochromatin beneath the kinetochore appears to have a role in sister chromatid cohesion and also as a marshaling area for chromosome passenger proteins such as inner centromere protein (INCENP)1 and TD-60. Why the passenger proteins first concentrate at centromeres before transferring to the central spindle and cleavage furrow during ana/telophase is not known. However, one of these proteins, INCENP, only appears to be able to participate in nonchromosomal events late in mitosis if it has first targeted to centromeres at metaphase, thereby revealing a functional link between the centromere and cytoskeletal events such as cytokinesis (Eckley et al., 1997; Mackay et al., 1998).
INCENP was originally discovered in a mAb screen for proteins that were tightly associated with the chicken mitotic chromosome scaffold (Cooke et al., 1987). The antigen identified by this antibody was a protein doublet of 135/155 kD. Subsequent studies revealed that INCENP appears to accumulate progressively in the central domain of the centromere during prometaphase and early metaphase (Earnshaw and Rattner, 1989; Earnshaw and Cooke, 1991). By late metaphase it leaves the chromosomes and becomes concentrated on linear tracks that transect the metaphase plate. At anaphase onset, these tracks are revealed as the stem body matrix material that coats overlapping antiparallel microtubules of the central spindle (Earnshaw and Cooke, 1991). Later in anaphase, a portion of the INCENP protein localizes to the inner surface of the plasma membrane at the site of presumptive cleavage furrow formation (Earnshaw and Cooke, 1991). INCENP is a very early marker for the presumptive furrow, concentrating at the equatorial cortex before myosin II and radixin (Eckley et al., 1997). During cytokinesis in heterokaryons, INCENP is found at both normal and ectopic “Rappaport” cleavage furrows that form between asters that are not linked by a central spindle bearing chromosomes (Rappaport, 1996; Eckley et al., 1997; Savoian et al., 1999).
Molecular cloning revealed INCENP to be a novel protein whose principal distinguishing features were a region predicted to form a coiled–coil and a number of possible phosphorylation sites (Mackay et al., 1993). INCENP is phosphorylated during mitosis by p34cdc2:cyclin B in Xenopus extracts (Stukenberg et al., 1997), although the functional consequences of this remain unknown. Deletion analysis revealed that the protein apparently has a modular organization, with the amino-terminal portion directing a number of chromosome-associated functions and the carboxyl-terminal portion being involved in interactions with the cytoskeleton (Mackay et al., 1993). Deletion of a portion of the amino terminus blocked the ability of INCENP to target to centromeres (Mackay et al., 1998), and prevented its transfer from the chromosomes to the spindle at anaphase (Mackay et al., 1993). Deletions of the carboxyl-terminal region of the protein interfere with INCENP–cytoskeletal interactions, including the ability to associate with microtubules (Mackay et al., 1993).
Two dominant-negative forms of INCENP have been shown to disrupt mitotic events in transfected cells. Expression of a CENP-B:INCENP chimeric protein, which remained tethered to centromeres throughout the cell cycle, resulted in a failure of cells to complete cytokinesis: daughters remained in pairs connected by a partly constricted intercellular bridge with a prominent midbody (Eckley et al., 1997). Expression of an amino-terminal half molecule (INCENP1-405) disrupted events both early and late in mitosis. This mutant interfered with the completion of prometaphase chromosome alignment and also with the completion of cytokinesis (Mackay et al., 1998). Thus, although INCENP function is as yet poorly understood, present data suggest that the protein plays a role in both chromosomal and cytoskeletal events during mitosis.
The aim of this study was to begin to characterize the mechanism of INCENP action in mitosis. We have focused on the amino-terminal chromosomal function region of INCENP. We start by identifying a 68–amino acid (aa) region of the protein that directs the movement of the protein to centromeres and subsequently to the spindle midzone. Within this region we identify short (13- and 11-aa) conserved functional motifs that are required for these movements. We identify an interaction between INCENP and heterochromatin proteins of the HP1 class, and present evidence suggesting that although interactions with HP1 are not essential for INCENP movements later in mitosis, the interaction of INCENP with HP1 may be part of a priming event that occurs at centromeres and enables INCENP to perform its cytoskeletal functions during the closing stages of mitosis.
Materials and Methods
Buffers and Reagents
Unless otherwise stated, all chemicals were obtained from Sigma (Dorset, UK) and all restriction and modifying enzymes used for DNA cloning were obtained from New England Biolabs (Beverly, MA). TEN buffer is 10 mM Tris-HCl, pH 7.7, 2 mM EDTA, 50 mM NaCl; KB buffer is 10 mM Tris-HCl, pH 7.7, 150 mM NaCl, 0.1% BSA; TE buffer is 10 mM Tris-HCl, pH 8.0, 1 mM EDTA; RSB buffer is 10 mM Tris-HCl, pH 7.4, 10 mM NaCl, 5 mM MgCl2; and glutathione _S_-transferase (GST) binding buffer is TEN buffer containing 0.05% Nonidet P-40.
Mapping INCENP Targeting Domains Using Green Fluorescent Protein (GFP)
Aequoria victoria GFP (clone TU 65, provided by M. Chalfie, Columbia University, New York, NY; Chalfie et al., 1994) was cloned into the Kpn and EcoRI sites within the multiple cloning site of the pECE vector (Ellis et al., 1986) downstream of INCENP1-839 cDNA, previously cloned into the BglII site (Mackay et al., 1993). This generated an intermediate construct, INCENP1-839:GFPi, which contained stop codons at the 3′ ends of both the INCENP and GFP cDNAs. INCENP1-839:GFPi was linearized using an AflII site 1212 bp from the start codon in INCENP, blunt-ended with T4 polymerase, then digested with BglII in the pECE multiple cloning site and blunt-ended using Mung bean nuclease. The vector was then ligated to create INCENP1-405:GFP. All GFP chimeras used in these studies contain the S65T point mutation (Heim et al., 1995). Carboxyl-terminal deletions of INCENP were then made by digesting INCENP1-405:GFP with HindIII and KpnI and subsequently with ExoIII nuclease (Promega, Madison, WI) digestion (Henikoff, 1984). In frame INCENP:GFP fusions were sequenced, CsCl purified (Ausubel et al., 1991), and electroporated into either HeLa or LLCPK cells as described previously (Mackay et al., 1993).
Cells were fixed 16 h after transfection with 4% paraformaldehyde using standard conditions (Mackay et al., 1993). Microtubules were labeled with the mouse mAb Tu27B (Caceres et al., 1983), a gift from Lester Binder (Northwestern University, Chicago, IL). DNA was visualized by staining with 4,6-diamidino-2-phenylindole (DAPI, 0.5 mg/ml; Calbiochem, Nottingham, UK). Coverslips were mounted in Vectashield (Vector Laboratories, Inc., Burlingame, CA). Images were acquired on a Zeiss Axioplan II epifluorescence microscope equipped with a Princeton Instruments Micromax cooled CCD camera, driven by IP Lab Spectrum software.
We used site-directed mutagenesis to generate a unique restriction site at a position 141 bp (47 aa) from the start of the coding region in INCENP1-405:GFP. This site lies between two highly conserved sequences (aa 32–44 and 52–62). Together with flanking restriction sites, this created a cloning cassette that allowed us to delete each sequence independently or to randomize the amino acids using a mutagenic oligonucleotide in which the amino acid composition of this region was retained, but the amino acids were organized in random order (see Fig. 5 B). Each sequence motif was manipulated separately.
Figure 5.

INCENP residues 52–62 constitute a spindle midzone-targeting motif. (A) INCENP1-405:GFP transfers to the central spindle and to midbodies (also see Fig. 2). Subsequent exonuclease III digests produced INCENP1-308:GFP (B), INCENP1-226: GFP (C), and INCENP1-68:GFP (D). Each of these proteins ultimately targeted the spindle midzone. (F) Randomization of the order of amino acid residues 52–62 in INCENP1-405 (52-62r):GFP specifically abolished this transfer. (E) Randomization of amino acid residues 32–44 in INCENP1-405 (32-44r) also abolished transfer to the midzone as a secondary consequence of the fact that this protein did not target centromeres at metaphase (see Fig. 4 F). In all images, the transfected INCENP:GFP fusion protein is shown in green, microtubules are red, and DNA is blue. The arrow in each panel indicates the position of the midbody. The darkened arrows in E and F represent the lack of transfected INCENP at the midbodies. All images are of pig LLCPK cells transfected with chicken INCENP constructs.
Chromosome spreads were made from transfected cells as described previously (Earnshaw and Migeon, 1985; Mackay et al., 1998). In brief, cells were plated onto poly-lysine–coated coverslips in individual 35-mm dishes and 16 h after transfection were incubated with 6 nM vinblastine for an additional 2 h. Cells were incubated with 3 ml of RSB for 10 min at room temperature. RSB was removed by aspiration and replaced with 500 μl of fresh RSB. Dishes were placed on foam cushions in the bottom of the bucket holders of a Heraeus Megafuge and centrifuged at 4,000 rpm for 10 min at room temperature. Cells were then fixed with 4% paraformaldehyde, permeabilized with 0.1% Triton X-100, and immunostained for centromeres using human autoimmune serum GS, which recognizes CENPs A, B, and C (Earnshaw and Rothfield, 1985) at a dilution of 1:5,000. After immunostaining, DNA was visualized with DAPI. Three-dimensional data sets of selected mitotic cells were collected using a DeltaVision microscope (Applied Precision, Issaquah, WA), based on an Olympus IX-70 inverted microscope with a Photometrics PXL cooled CCD camera. Data sets were deconvolved, projected onto a single plane, and the contrast was adjusted before export as TIFF files to Adobe Photoshop.
Yeast Two-Hybrid Assay
We used vectors and yeast strains described by Brent and co-workers (Gyuris et al., 1993). Full-length INCENP and truncations thereof were cloned from pBluescript in frame into the pEG202 bait vector, tested for activation in the absence of an interactor plasmid, and further tested in the presence of HP1Hsα in pJG4-5 (interactor plasmid) and pSH18-34 (reporter plasmid). pRFHM 1 (encoding a bicoid protein) and pSH17-4 were used as negative and positive activation controls, respectively, as described (Gyuris et al., 1993). An initial screen of 500 recombinant clones from a HeLa cDNA library (Gyuris et al., 1993) led to the isolation of one clone of HP1Hsα and two clones of HP1Hsγ. Two independent yeast colonies from each construct listed in Fig. 1 were tested for galactose-dependent growth in the absence of leucine, blue/white selection, and quantitation in a β-galactosidase liquid assay. For blue/white selection, yeast colonies were plated onto Gal-U-H-W-L plates, overlaid with top agar containing NaPO4, pH 7.0, 2% _N_-lauroylsarcosine sodium salt, 0.04% β-mercaptoethanol, 0.02% X-gal, incubated at 30°C, and checked for blue colonies. β-galactosidase activity of yeast transformants grown in liquid culture was determined using _o_-nitrophenyl β-d-galactopyranoside (ONPG) as a substrate as described (Reynolds and Lubland, 1989). All plasmids used for the two-hybrid screen were obtained from the R. Brent laboratory (Harvard Medical School, Boston, MA).
Figure 1.
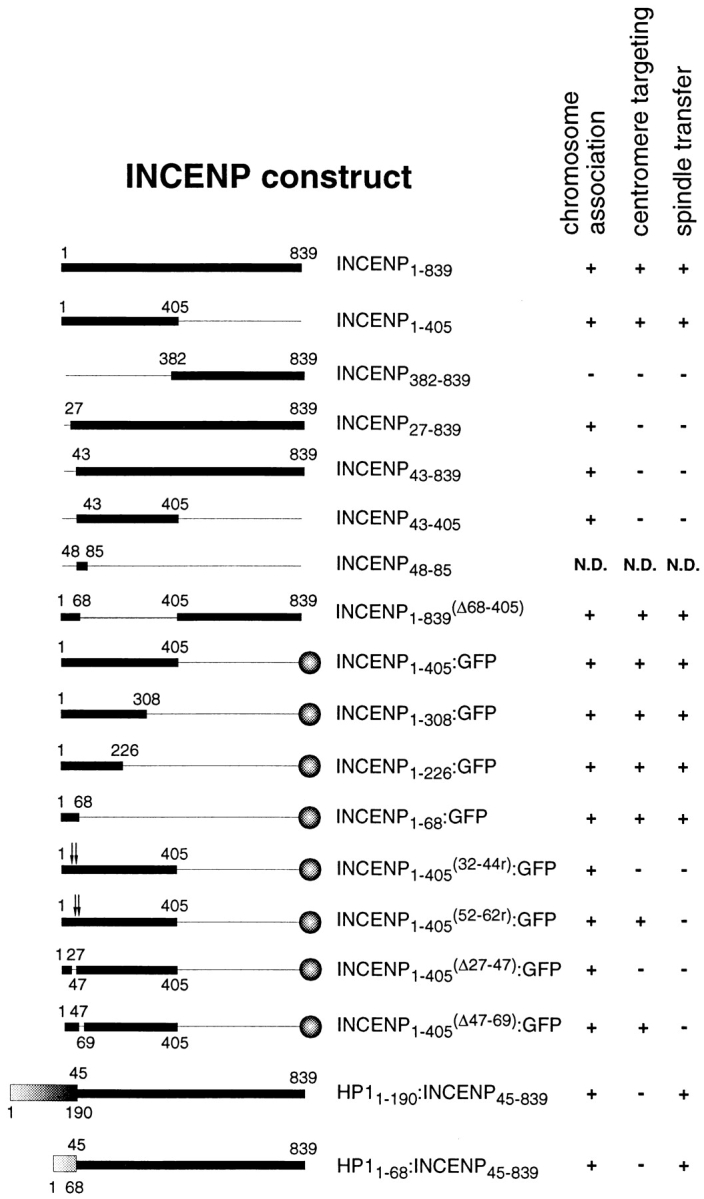
INCENP constructs used in this study. The numbers correspond to amino acids and the relevant behavior of the proteins during mitosis is summarized at the right. The thick and thin lines indicate protein sequences that were expressed or deleted, respectively. The balls at the right indicate carboxyl-terminal fusions with A. victoria GFP. The shaded boxes (bottom left) represent the chromo domain (aa 1–68) or full-length HP1Hsα which was fused in frame to INCENP45-839. In INCENP1-405 (32-46r):GFP and INCENP1-405 (52-62r):GFP the amino acid sequences between the arrows were randomized. These sequences are shown in Fig. 5. Due to cloning constraints, INCENP amino-terminal deletion constructs in the pEG202 vector used for two-hybrid analysis (Fig. 7) begin expression of the protein at aa 45 rather than 43. For simplicity, these constructs have been omitted here.
Cloning GST Chimeras
PCR was used to generate GST:HP1Hsα 1-68, GST:HP1Hsα 68-111, and GST: HP1Hsα 110-190 by designing sequence specific primers with unique restriction sites at the 5′ end. These products were subcloned in-frame into the pGex vector (Pharmacia, Herts, UK) and fully sequenced. GST:HP1Hsα 1-110 and GST:HP1Hsα 68-190 were both subcloned using standard methods. GST:HP1Hsα 1-190 was digested with BamHI and EcoRV to release the GST:HP1Hsα 1-110 insert and this fragment was subcloned into an empty pGex 2T vector previously digested with EcoRI, blunt-ended using Mung bean nuclease, and then digested with BamHI. GST:HP1Hsα 68-190 was generated by ApoI digestion of GST:HP1Hsα 1-190 to release the insert and the fragment was subcloned into an EcoRI-digested and phosphatased pGex 1 vector. All cloning junctions were sequenced to confirm that open reading frames were maintained.
Binding Assay Using In Vitro Translated Proteins
Full-length INCENP1-839 was subcloned into pBluescript (Stratagene, Cambridge, UK) using standard methods and then truncated by restriction enzyme digestion to create INCENP1-405, INCENP43-839, and INCENP43-405 as described (Mackay et al., 1993; Mackay et al., 1998). cDNAs were then transcribed and translated using the TNT T3 system (Promega) in the presence of [35S]methionine (ICN, Thame, UK) for 1 h at 30°C. Labeled proteins were diluted to 500 μl in GST binding buffer and concentrated by centrifugation using a 10,000 molecular weight cutoff filter (Millipore Corp., Bedford, MA) for 15 min at 4°C (Heraeus, Brentwood, UK) followed by two additional washes with binding buffer.
HP1Hsα cDNAs (Saunders et al., 1993) were subcloned (see above), transformed into Escherichia coli CAG456 cells, and expression induced with 0.5 mM IPTG for 5 h, 30°C. GST:HP1Hsα chimeras were purified by repeated rounds of fast freeze and thaw, followed by the addition of 4 mg/ml egg white lysozyme, sonication, and centrifugation at 4,000 rpm, 10 min at 4°C in a Heraeus Megafuge. Translated INCENP protein (20 μl) was added to 150 μl GST binding buffer, incubated with glutathione agarose conjugated to 3–5 mg of GST alone or GST:HP1Hsα chimeras, and incubated on a rotating mixer for 1 h at 4°C. After incubation, samples were washed three times with GST binding buffer. Bound fractions were solubilized by boiling in 1× sample buffer for 3 min and analyzed on a 12% SDS-polyacrylamide gel followed by Coomassie blue staining and autoradiography. Synthesis was confirmed before each experiment.
INCENP1-405:GFP and INCENP1-68:GFP were created in the pECE vector as described above and subcloned into pBluescript (Stratagene) for in vitro translation. These proteins were tested as above with full-length HP1Hsα. GFP and an empty vector were used as controls.
Expression of INCENP That Is Tethered to HP1
Primer T3 and sequence specific PCR primers (Oswel, Southampton, UK) with a 3′ SacII site were used to generate the full-length HP1 (5′GCAGGAATTCTCCGCGGGGCTCTTTGCTGT3′) or the chromo domain alone (5′GCAGGAATTCTCCGCGGGCATAAATTCAGA3′). The PCR products and the CENP-B1-158:INCENP45-839 vector (Eckley et al., 1997) were digested with BglII and SacII and the fragments were subcloned using standard methods. The resultant constructs were purified by CsCl density gradients (Ausubel et al., 1991) and transfected into HeLa cells as described previously (Mackay et al., 1993) onto duplicate coverslips. Cells were fixed 16 h after transfection, for 5 min at −20°C in methanol. Immunofluorescence was performed as described above, using a chicken-specific anti-INCENP antibody (RabC) (Mackay et al., 1993) at a dilution of 1:500. Both full-length INCENP and INCENP43-839 transfectants were used as controls.
Immunoblotting of Transfected Proteins
HeLa cells were transfected with constructs encoding INCENP1-839, INCENP43-839, or HP1Hsα 1-68:INCENP45-839 as described and plated onto one 100-mm petri dish containing a 16-mm coverslip. The coverslip was removed 16 h after transfection, the cells were fixed for 5 min at −20°C in methanol, and stained for INCENP (RabC) and DNA (DAPI) as described above. The transfection efficiency was calculated by dividing the number of transfectants by the total number of cells stained with DAPI. Five fields were counted per sample for a total of ∼100 cells. The cells remaining in the petri dish were trypsinized, washed twice with PBS, and counted in a hemacytometer. Cell pellets were resuspended in 1× sample buffer, boiled for 3 min and sonicated. Equivalent cell numbers were loaded per lane. Blots were probed using an INCENP antibody (RabC) and visualized using ECL reagents (Amersham, Buckinghamshire, UK).
Functional Analysis of INCENP Lacking the HP1-binding Domain
INCENP1-839 in the pECE vector was digested with Eagl and AflII, blunt-ended using Mung bean nuclease, and the vector was religated to generate INCENP1-839 (Δ69-404). After transfection, immunofluorescence was performed using the INCENP antibody (RabC) as described above. In parallel, INCENP1-839 (Δ69-404) was subcloned into pBluescript, transcribed and translated (Promega), and tested for in vitro binding to GST or GST: HP1Hsα. The samples were resolved by SDS-PAGE and visualized by autoradiography. INCENP1-839 was used as a control.
Results
The Amino-terminal End of INCENP Functions as an Autonomous Centromere-targeting Module
During prophase and early prometaphase, INCENP is distributed along the chromosome arms; however, the protein becomes concentrated at centromeres by metaphase (Earnshaw and Cooke, 1991). Other centromere proteins such as CENPs A–C and E all bind directly to the centromere with no prior binding to the chromosome arms (Earnshaw and Rothfield, 1985). Therefore, we refer to this distinctive pattern of INCENP movements as indirect centromere targeting.
Previous protein truncation studies demonstrated that indirect centromere targeting by INCENP requires the amino-terminal 42-aa residues of the protein (Mackay et al., 1998). To delineate the minimal region of the protein required to carry out this novel movement, we generated a series of progressively shorter amino-terminal INCENP fragments, each containing GFP fused at its carboxyl terminus. The localization of these proteins on metaphase chromosomes was confirmed in both cycling and metaphase-arrested cells. We began by confirming that an INCENP:GFP chimeric protein underwent indirect centromere targeting (Fig. 2). INCENP1-405:GFP was distributed all along the chromosomes at prophase, becoming concentrated at centromeres during metaphase. This protein could transfer to the spindle at anaphase, albeit less efficiently than wild-type INCENP (Fig. 2 D; see also Earnshaw and Cooke, 1991; Mackay et al., 1998). Observation of living cells (data not shown) revealed that the targeting of INCENP1-405:GFP to centromeres is not due to loss of the noncentromeric INCENP from chromosomes, but instead represents an accumulation of the protein at centromeres.
Figure 2.
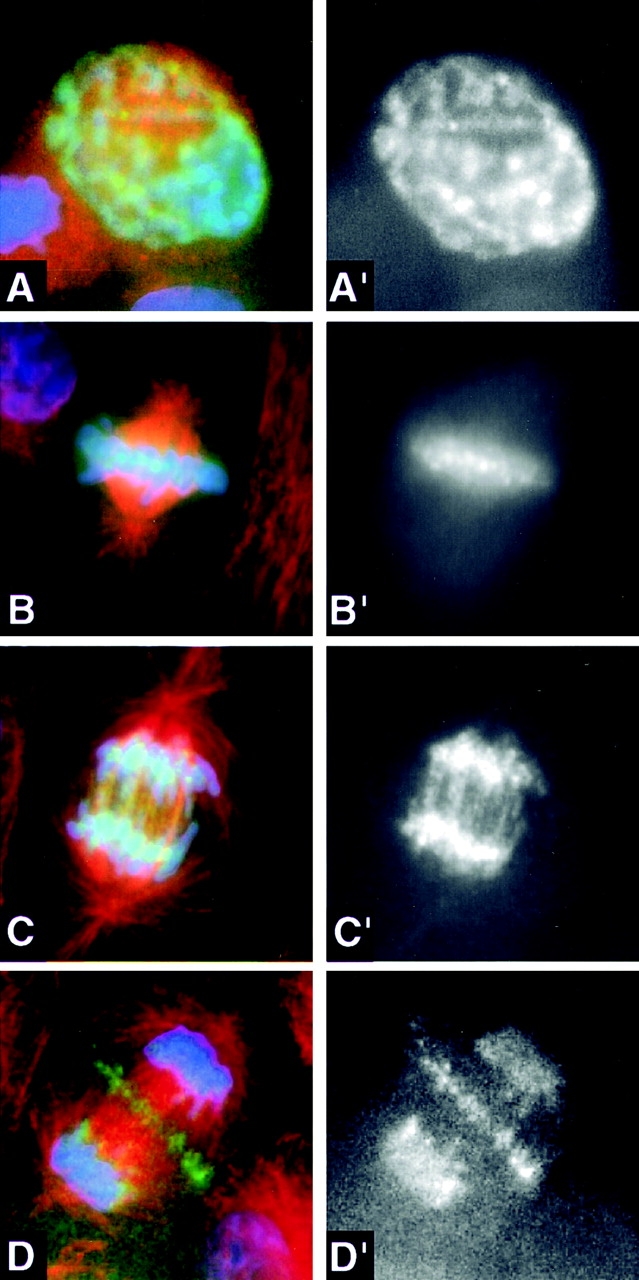
INCENP1-405:GFP is released from centromeres back onto the chromosome arms at anaphase onset, but still transfers from chromosomes to the spindle midzone. Transfected LLCPK cells expressing INCENP1-405:GFP (green) were stained for microtubules (red) and DNA (blue). The panels to the right show the INCENP pattern separately in black and white. (A and A′) Prophase. INCENP is distributed along the chromosome arms. (B and B′) Metaphase. INCENP is concentrated in spots at centromeres. (C and C′) Early anaphase. INCENP1-405:GFP is released from centromeres back onto the chromosome arms. (D and D′) Late anaphase. INCENP1-405:GFP gradually transfers from chromosomes to the spindle midzone. By late telophase, all detectable INCENP1-405:GFP has transferred to the midbody (see Fig. 6 A).
INCENP1-405:GFP (Fig. 3 A′) and the progressively smaller truncated molecules, INCENP1-308:GFP and INCENP1-226: GFP, targeted to centromeres in mitosis (Fig. 3, B′ and C′). Ultimately, we found that a polypeptide containing only the amino-terminal 68 aa of INCENP was sufficient to target GFP to centromeres (Fig. 3 D′). In control experiments, transfected GFP that was not fused to other proteins localized diffusely in the cytoplasm during interphase and was never found associated with the chromatin (Rizzuto et al., 1995; and data not shown). Thus, INCENP contains a relatively compact module at its amino terminus that is necessary and sufficient to target the protein to centromeres during the early stages of mitosis.
Figure 3.

Amino acid residues 32–44 are required for centromere targeting by INCENP. HeLa cells expressing various INCENP constructs fused to GFP (green) were arrested in mitosis with vinblastine, stained for centromeres with an autoimmune serum (red) and for DNA with DAPI (gray), and visualized by three-dimensional deconvolution microscopy (Agard et al., 1989). Transfected INCENP1-405:GFP, INCENP1-308:GFP, INCENP1-226: GFP, INCENP1-68:GFP, and INCENP1-405 (52-62r):GFP all concentrate at the inner centromere. However, INCENP1-405 (32-44r):GFP fails to localize at centromeres, but instead paints the entire chromosome. In INCENP1-405 (52-62r):GFP and INCENP1-405 (32-44r):GFP, the amino acid sequence indicated was randomized using a mutagenic oligonucleotide.
Further analysis of the distribution of the amino-terminal INCENP:GFP fusion proteins in cycling mitotic cells revealed a surprising result. Every truncated INCENP molecule capable of targeting to centromeres was also able to transfer to the anaphase spindle and ultimately to the midbody (see below). Thus, the minimal centromere-targeting module also functions as a midbody-targeting module.
Identification of a Conserved 13-aa Motif That Is Essential for Centromere Targeting
The observation that chicken INCENP targets normally to centromeres in mammalian cells (Mackay et al., 1998) suggests that the mechanism of centromere targeting by INCENP is highly conserved. Therefore, we looked for regions of amino acid conservation within the INCENP minimal centromere-targeting module. Comparison of the first 68 aa of the chicken, Xenopus, and mouse INCENP proteins revealed two highly conserved regions corresponding to aa 32–44 (69% identity) and 52–62 (100% identity) of the chicken INCENP polypeptide (Fig. 4 A).
Figure 4.

The INCENP minimal centromere-targeting module (residues 1–68) contains two highly conserved amino acid sequence motifs required for centromere targeting and spindle transfer. (A) Sequence alignment between the amino-terminal 68-aa residues of chicken, Xenopus, and mouse INCENP. Amino acids that are conserved in all three species are shown in bold type in the line labeled consensus, with similar residues indicated by a black dot. There is 69% sequence identity for residues 32–44 and 100% sequence identity for residues 52–62. (B) Schematic representation of the amino-terminal region of chicken INCENP. The restriction sites used to make deletion and mutant constructs are shown above the bar. The hatched region indicates the centromere-targeting motif. The filled region represents the spindle transfer motif. The boxed sequences below the bar correspond to the wild-type chicken sequences for these motifs. Below these are shown the mutagenized sequences, arrived at by randomization of the amino acid sequences shown in the boxes.
Two approaches were taken to examine the involvement of the conserved region spanning aa 32–44 in indirect centromere targeting by INCENP. First, we deleted this region of the protein. The mutated protein, INCENP 1-405 (Δ27-47):GFP, bound to chromosome arms, but failed to target to centromeres (data not shown). Second, we used a mutagenic oligonucleotide to randomize the amino acid sequence as shown in Fig. 4 B. This also abolished centromere targeting: INCENP1-405 (32-44r):GFP “painted” the chromosome arms (Fig. 3 F). Therefore, the conserved motif encompassing aa 32–44 is essential for targeting INCENP to centromeres during mitosis. To determine whether aa 32–44 are sufficient to target INCENP to centromeres, we prepared a deletion construct (INCENP27-839) encoding a protein missing aa 1–26. This protein failed to target centromeres (data not shown), behaving in all respects like INCENP45-839 (Mackay et al., 1993). This suggests that sequences in the first 26 aa of INCENP are also essential for centromere targeting, although effects of the deletion on protein structure cannot be excluded.
Identification of a Highly Conserved 11-aa Motif Required for Midbody Targeting by INCENP
We next examined the role of the highly conserved sequence (aa 52–62) in centromere targeting. Deletion of aa 47–69 (Fig. 1) had no effect on centromere targeting. In agreement with this result, randomization of the conserved 11-aa sequence as shown in Fig. 4 B also had no effect on indirect centromere targeting: INCENP1-405 (52-62r): GFP located normally to centromeres (Fig. 3 E′). Thus, INCENP residues 52–62 are not involved in targeting the protein to centromeres.
Instead, INCENP residues 52–62 appear to be involved in targeting INCENP to the midbody. Although INCENP 1-405 (52-62r):GFP was unimpaired in its ability to target to the centromere (Fig. 3 E′), the protein was subsequently unable to target to the central spindle or midbody (Fig. 5 F). In telophase cells, INCENP1-405 (52-62r):GFP colocalized with the chromatin as seen by the overlap of blue and green. These data reveal for the first time that INCENP movements to centromeres and to the spindle involve distinct peptide motifs and perhaps different ligands.
In other studies, INCENP molecules in which the sequence of aa 32–44 had been randomized not only failed to target to centromeres, but also remained associated with chromatin throughout mitosis and did not transfer to the anaphase spindle (Fig. 5 E). Although we cannot formally exclude that aa 32–44 encodes a separate determinant required for midbody targeting, the simplest interpretation of this result is that, as suggested by our earlier observations with amino-terminal deletion proteins INCENP43-839, INCENP43-405 (Mackay et al., 1998) and INCENP27-839, INCENP must first target to centromeres if it is to transfer to the midbody later in mitosis.
Identification of Cellular Proteins That Interact with the Amino-terminal Region of INCENP
Given the essential role played by the amino-terminal region of INCENP in centromere targeting and spindle transfer, we predicted that this region might make important contacts with other chromosomal proteins. To identify such interacting factors, we performed a yeast two-hybrid screen of a HeLa cell cDNA library using INCENP 1-405 fused to the LexA DNA-binding domain as bait (Gyuris et al., 1993). An initial screen led to the isolation of seven clones that grew on the appropriate media and were blue in the presence of β-galactosidase. None of the seven proteins interacted with control bait proteins, including the LexA DNA-binding domain alone, bicoid, and GFP alone (Fig. 6 A and data not shown). In addition, all seven interacted both with INCENP1-405:GFP and with full-length INCENP1-839. Subsequent DNA sequence analysis identified three classes of putative INCENP-interacting proteins: known heterochromatin proteins (three clones), β-tubulin (one clone), and novel polypeptides (three clones). The remainder of this report is concerned with our analysis of the interaction of INCENP with members of the first class, which comprised one isolate of human heterochromatin protein 1 alpha, HP1Hsα (Saunders et al., 1993), and two independent isolates of HP1Hsγ (Ye and Worman, 1996).
Figure 6.

INCENP interacts with HP1Hsα in a yeast two-hybrid assay. Various INCENP deletion constructs were subcloned into the pEG202 vector (Gyuris et al., 1993), and expressed as “bait” in S. cerevisiae. These cells were cotransformed with the pSH18-34 reporter plasmid together with the pJG4-5 vector encoding full-length HP1Hsα. (A) Two clones from each transformation were grown in liquid culture and duplicate samples from each were tested for β-galactosidase activity using ONPG as substrate. Each bait construct is listed at the left of the graph and the relative units of activity are indicated on the x-axis. The numbers to the right of the error bar indicate the calculated activity and the standard error measurements (n = 4). (B) Identification of the region of INCENP that interacts with HP1. Various INCENP constructs in pEG202 were tested for interaction with HP1Hsα using the blue/white overlay assay. +, positive interaction between the INCENP bait and HP1 (specific growth on selective media [Gal-U-H-W-L] and activation of the β-galactosidase gene). −, lack of interaction between the INCENP bait and HP1 (failure of cells to grow on selective media).
We performed a number of control experiments using a quantitative liquid assay with ONPG as the substrate in order to better characterize the two-hybrid interaction between HP1Hsα and INCENP (Fig. 6 A). Neither INCENP 382-839, which fails to associate with chromosomes in vivo (Mackay et al., 1998), nor INCENP48-85, which is encoded by a single conserved exon in the human INCENP sequence (Eckley, D.M., and W.C. Earnshaw, unpublished observations) and which contains the spindle targeting motif (residues 52–62), interacted with HP1Hsα in the two-hybrid assay (Fig. 6 A). Further studies using a series of truncated INCENP molecules localized the HP1-binding site to a region between INCENP residues 135 and 270, and possibly between residues 135 and 200 (Fig. 6 B).
We next used an in vitro binding assay to confirm that HP1Hsα and INCENP are capable of direct biochemical interaction. GST-tagged HP1Hsα protein was expressed in E. coli, bound to glutathione agarose, and incubated with [35S]methionine-labeled INCENP proteins that had been translated in vitro using reticulocyte lysates. Under these conditions, GST:HP1Hsα bound selectively to INCENP1-839 (Fig. 7, lane 2), INCENP1-405 (lane 4), and INCENP1-405: GFP (lane 12). In control experiments, these INCENP molecules showed no interaction with GST alone (Fig. 7, lanes 1, 3, and 11). The interaction between INCENP and HP1 in vitro was stable at physiological ionic strength, but was disrupted by higher salt concentrations (data not shown).
Figure 7.

HP1Hsα interacts specifically with INCENP in vitro. Either GST (odd-numbered lanes) or an HP1Hsα:GST fusion protein (even-numbered lanes) expressed in E. coli was bound to glutathione agarose and incubated with in vitro translation reactions containing various INCENP derivatives. The lanes show as follows: INCENP1-839 (lanes 1 and 2); INCENP43-405 (lanes 3 and 4); INCENP43-839 (lanes 5 and 6); INCENP43-405 (lanes 7 and 8); GFP control (lanes 9 and 10); INCENP1-405:GFP (lanes 11 and 12); and INCENP1-68:GFP (lanes 13 and 14). Both panels show autoradiographs.
HP1 May Promote the Release of INCENP from the Chromosomes
Because of the well known association of HP1Hsα with centromeres (Nicol and Jeppesen, 1994; Wreggett et al., 1994), we had initially expected that the INCENP:HP1Hsα interaction would be involved in targeting INCENP to centromeres. However, further experiments revealed that this is unlikely. INCENP1-68:GFP, which defines the minimal INCENP centromere-targeting module (Fig. 3 D), failed to interact with HP1Hsα in the two-hybrid and in vitro binding assays (Fig. 6 and Fig. 7, lanes 13 and 14). In addition, both INCENP43-839 and INCENP43-405, which associate with chromosomes but do not target to centromeres, showed significant interaction with HP1Hsα (Fig. 6 and Fig. 7, lanes 6 and 8).
Although the two-hybrid and in vitro binding analyses suggest that HP1 is not involved in targeting INCENP to centromeres, it is possible that the INCENP:HP1 interaction at centromeres might be required for some other aspect of INCENP function in mitosis. To attempt to identify such a role of HP1 binding in INCENP function, we created a deletion construct, INCENP1-839 (Δ69-404), from which the HP1-binding site was deleted. This protein failed to interact with HP1 in the in vitro binding assay (Fig. 8, A and B), yet it showed surprisingly normal behavior in vivo: it was nuclear in interphase, centromeric at metaphase, and located to the spindle midzone and midbody at ana/telophase (Fig. 8, C–F). Thus, HP1 cannot be solely responsible for directing any of the obvious aspects of INCENP behavior, at least for INCENP molecules that are capable of targeting to centromeres.
Figure 8.
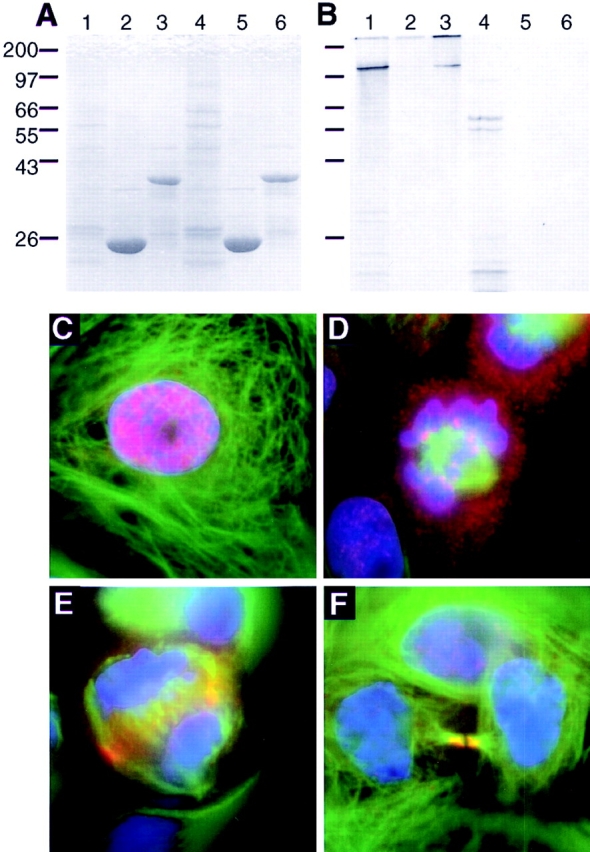
INCENP lacking the HP1-binding domain moves normally in mitosis. (A and B) Either GST (lanes 2 and 5) or GST: HP1Hsα (lanes 3 and 6) immobilized on glutathione agarose was incubated with in vitro translated INCENP1-839 (lanes 1–3) or INCENP1-839 (Δ69-404) (lanes 4–6). Lanes 1 and 4 show the input of translated protein used in this experiment. A shows the Coomassie blue–stained gel, while B is the autoradiograph. (C–F) Transfected LLCPK cells were stained for INCENP1-839 (Δ69-404) (red), tubulin (green), and DNA (blue). The pattern of INCENP movements shown here is identical to that seen with the full-length INCENP.
Therefore, we were surprised when a second experiment revealed that the interaction between INCENP and HP1 might after all be one component of a redundant system promoting the release of INCENP from chromosomes and targeting to the spindle midzone. Initially to examine the potential role of HP1 in centromere targeting by INCENP, we constructed two chimeric cDNAs that encoded proteins in which either the chromo domain (residues 1–68 of HP1Hsα) or full-length HP1Hsα was fused to the amino terminus of INCENP45-839, a form of INCENP that we have shown previously to be unable to target to centromeres or to transfer to the spindle (Mackay et al., 1998). In a previous study, we found that a similar chimeric protein consisting of a fusion between the centromere-binding domain of CENP-B and INCENP45-839 could target to centromeres, where it remained bound throughout the cell cycle (Eckley et al., 1997).
Analysis of transfected cells expressing either HP1Hsα: INCENP45-839 (data not shown) or HP1Hsα 1-68:INCENP 45-839 (Fig. 9 D′) during mitosis revealed a striking result. Both chimeric polypeptides associated with mitotic chromosomes, but were unable to target to centromeres. However, despite this inability to target to centromeres, both chimeric proteins transferred efficiently to the spindle in anaphase (Fig. 9 D′′), whereas control protein INCENP 45-839 did not. Immunoblots of transfected cells confirmed that polypeptides of the predicted size could be observed, and that these and the control proteins accumulated to roughly equal amounts in vivo. This result suggests that the INCENP:HP1 interaction may be one component of a redundant mechanism promoting the transfer of INCENP to the spindle at anaphase.
Figure 9.

Fusion of INCENP to the chromo domain of HP1Hsα bypasses the requirement for INCENP to target to centromeres before transfer to the spindle midzone. (A) Immunoblot showing the protein expression levels of transfected INCENP1-839 (lane B), INCENP43-839 (lane C), and HP1Hsα 1-68:INCENP45-839 (lane D). Equivalent cell numbers were loaded in each lane. Cells transfected with vector alone (lane v) were used as a control. The upper band seen in all lanes is a cross-reactive protein that acts as a loading control. (B′ and B′′) Full-length INCENP1-839 targeted to centromeres at metaphase (B′) and to the midbody in telophase (B′′). (C′ and C′′) INCENP43-839 does not target to centromeres at metaphase (C′) and remains on the chromosomes throughout mitosis (C′′). (D′ and D′′) A chimeric molecule in which the chromo domain of HP1Hsα (aa 1–68) was fused to INCENP45-839 does not target to centromeres at metaphase (D′), but does target to the midbody in telophase (D′′). Identical results were obtained with a chimera containing the full-length HP1Hsα fused to INCENP45-839 (not shown). The figure shows transfected HeLa cells stained for INCENP (red), microtubules (green), and DNA (blue). The arrows indicate the position of the midbody in the telophase cells.
INCENP Interacts with HP1 in a Novel Manner
Sequence analysis and functional experiments in vivo reveal that HP1 has a modular domain organization. The amino- and carboxyl-terminal portions of the molecule have been termed the chromo domain and the chromo shadow domain, respectively (Paro and Hogness, 1991; Aasland and Stewart, 1995). Both the Drosophila melanogaster HP1 chromo domain and chromo shadow domains could target the euchromatic protein Polycomb to heterochromatin when swapped for the Polycomb chromo domain (Platero et al., 1995), although only the chromo shadow domain could target to heterochromatin on its own (Powers and Eissenberg, 1993). Recent studies have identified a number of cellular proteins that interact with HP1s (α, β, and γ) (reviewed in Cavalli and Paro, 1998; Lamond and Earnshaw, 1998). Most of these interactions occur via recognition of the chromo shadow domain. Occasionally, the interaction requires the additional presence of the central so-called hinge region.
To gain further insight into the nature of the interactions between INCENP and HP1, we performed in vitro binding experiments (as described above) using a series of truncated HP1 proteins (Fig. 10 A) and full-length INCENP. These studies mapped the region in HP1Hsα that interacts with INCENP to the hinge region separating the chromo and chromo shadow domains (Fig. 10). Deletion constructs expressing either the chromo domain (HP1Hsα 1-68), or chromo shadow domain (HP1Hsα 110-190), did not bind to INCENP1-839 (Fig. 10, B and C, lanes 2 and 6). However, when either of these fragments was extended to include the hinge region (Fig. 10, lanes 3 and 5) or when the hinge region (HP1Hsα 68-110) was expressed on its own (Fig. 10, lane 4), specific interactions were seen. This is the first demonstration of interaction of a chromatin protein exclusively with the hinge region of HP1.
Figure 10.

INCENP interacts with the hinge region of HP1Hsα. (A) Schematic diagram of the organization of HP1 together with the various GST-HP1 constructs used in this experiment. In each case, the number at the right represents the gel lanes in which the particular construct was used. The various GST:HP1 constructs were expressed in E. coli, isolated on glutathione agarose, incubated with in vitro translation reactions containing 35S-labeled full-length INCENP1-839, and subjected to SDS-PAGE. This gel was first stained with Coomassie blue (B) and subsequently exposed to film (C). The gel lanes are as follows: i, total in vitro translation reaction; C, GST alone; 1, HP1Hsα 1-190; 2, HP1Hsα 1-68; 3, HP1Hsα 1-110; 4, HP1Hsα 64-111; 5, HP1Hsα 64-190; and 6, HP1Hsα 110-190.
Discussion
A Novel Centromere-targeting Module Directs Movement of INCENP from Noncentromeric Sites to Centromeres
Unlike other centromeric proteins, including CENPs A–C, E, and F, Mad2, and Bub1, which target directly to the centromere/kinetochore (Earnshaw and Rothfield, 1985; Yen et al., 1991; Casiano et al., 1993; Rattner et al., 1993; Zhu et al., 1995; Chen et al., 1996; Taylor and McKeon, 1997), INCENP binds first to noncentromeric sites along the chromosome arms, then moves to centromeres only during prometaphase/metaphase. We refer to this behavior as indirect centromere targeting.
Indirect centromere targeting by INCENP is directed by an autonomous module located in INCENP residues 1–68 (Fig. 11) and including the conserved sequence WLEEIHEEAARMF (residues 32–44, Fig. 11, motif C). This sequence is not related to previously determined centromere-targeting motifs (Pluta et al., 1992; Yang et al., 1996). An INCENP molecule in which the 13-aa centromere-targeting motif was mutagenized (INCENP1-405 (32-44r): GFP), in addition to being defective in centromere targeting, was also defective in transfer to the anaphase spindle and midbody. This supports our previous suggestion (Mackay et al., 1998) that INCENP targeting to centromeres is an obligatory event upstream of its transfer to the spindle in ana/telophase.
Figure 11.

Functional map of INCENP from this and previous studies. The Cen module (residues 1–68) is the minimum region giving centromere targeting. Within this, the 13-aa motif required for centromere targeting (motif C) and the 11-aa motif required for spindle transfer (motif S) are shown. The HP1-binding domain includes residues 135–270, the minimum region of INCENP shown to interact with HP1. The chromosome-binding and microtubule-binding regions were identified in previous studies (Mackay et al., 1993; Mackay et al., 1998).
During this analysis we unexpectedly discovered that the minimal centromere-targeting module also functions as an autonomous spindle midzone/midbody-targeting unit during ana/telophase. Site-directed mutagenesis studies revealed that a conserved sequence motif spanning residues 52–62 (PELMPKTPSQK), which was not required for indirect centromere targeting, was essential for spindle/midbody targeting by INCENP1-405:GFP (Fig. 11, motif S). However, in addition to this amino-terminal motif, INCENP must also have a second spindle transfer determinant elsewhere, between residues 405 and 839: full-length INCENP1-839 carrying the (52-62r) mutation also transfers to the spindle during anaphase (data not shown). However, both INCENP1-405:GFP and INCENP1-839 (52-62r) transferred to the central spindle abnormally late, in mid-anaphase, and with reduced efficiency (some mutant INCENP typically remained associated with the chromosomes, Fig. 2 and data not shown). Thus, although either the amino-terminal or carboxyl-terminal motif is sufficient to target centromere-associated INCENP to the midbody, the presence of both motifs appears to be essential for timely, complete, and efficient transfer to occur. These observations reveal that INCENP uses redundant mechanisms to target to the spindle midzone, a theme that is emphasized in our studies of the interaction of INCENP with HP1 (see below).
Centromere Targeting Primes INCENP for Later Spindle Transfer in Ana/Telophase
INCENP1-405:GFP is released from the centromeres back onto the chromosome arms at anaphase onset, yet still transfers to the spindle during mid-anaphase (Fig. 2). A similar release of INCENP back onto the chromosome arms occurs in rare cells in which INCENP transfer to the spindle is spontaneously delayed (Earnshaw, W.C., unpublished observations). However, in our various studies, in observations of many thousands of cells, we have never seen a late anaphase/telophase cell in which transfer of endogenous INCENP to the spindle had failed to occur. These observations reveal a paradox. Why do INCENP 43-405, INCENP43-839, and INCENP27-839, all of which occupy similar locations on the chromosome arms during anaphase and which contain an intact spindle/midbody-targeting motif (aa 52–62), fail to transfer to the central spindle at any time during ana/telophase? The most obvious difference between these molecules is that like endogenous INCENP, INCENP1-405:GFP had previously targeted centromeres during metaphase, whereas the other mutant proteins had not.
We suggest that INCENP may undergo some sort of priming event at centromeres that renders it capable of targeting to the spindle midzone and midbody even if it leaves the centromeres before its exit from the chromosome. This priming event could be a posttranslational modification such as phosphorylation by a centromere-associated kinase such as active ERK (Shapiro et al., 1998; Zecevic et al., 1998) and/or an interaction with a centromeric protein that promotes INCENP release from the chromosomes at anaphase. To explore this second possibility, we have screened for proteins with which INCENP might interact at centromeres.
Specific Interactions of INCENP with Heterochromatin Proteins
A yeast two-hybrid screen revealed a specific interaction between INCENP and heterochromatin proteins HP1Hsα and HP1Hsγ. HP1s are thought to be structural adapter proteins involved in the assembly of chromatin complexes (reviewed in Lamond and Earnshaw, 1998). HP1s interact with a number of proteins including HP1 itself (Ye et al., 1996), the lamin B receptor (Ye and Worman, 1996), Orc1 and Orc2 (Pak et al., 1997), and SP100, a major component of PML nuclear bodies (Lehming et al., 1998; Seeler et al., 1998). The INCENP:HP1 interaction, which was confirmed in in vitro binding studies, involves residues 135–270 of INCENP and the central, so-called hinge region of HP1. Although inclusion of the hinge region has been reported to increase the binding of some proteins in vitro to HP1 (Ye et al., 1996), interaction solely with this sequence has not been reported previously. It is possible that INCENP binding to HP1 might leave the chromo and chromo shadow domains available for further interactions and possible ternary complex formation.
For the interaction between HP1Hsα and INCENP to be functionally significant, then the two proteins should colocalize in vivo, at least during some portion of the mitotic cycle. In fact, indirect immunofluorescence on metaphase chromosome spreads has revealed that both INCENP and HP1Hsα are distributed throughout the central domain of the mammalian centromere (Nicol and Jeppesen, 1994; Wreggett et al., 1994; Mackay et al., 1998), consistent with an association between the two proteins in vivo.
The observed interaction between HP1Hsα and INCENP suggests two possibilities. First, binding to HP1 might be responsible for targeting INCENP to centromeres. This model is effectively ruled out by our two-hybrid, in vitro binding and in vivo transfection studies. Second, interaction with HP1 could be a component of the priming event that occurs at centromeres and which is required for INCENP function during late mitosis. The interaction of HP1 with INCENP cannot be solely responsible for priming the transfer of INCENP to the spindle, since two proteins that do not bind HP1 (INCENP1-68 and INCENP1-839 [Δ69-404]) can transfer to the spindle and one that does bind HP1 (INCENP1-839 [52-62r]) does not transfer. However, both of the proteins that do transfer also target to centromeres and could have received a second priming event there.
Other results suggest that the INCENP:HP1 interaction could have a role in spindle transfer. Thus, fusion of either the chromo domain (residues 1–68 of HP1Hsα) (Aasland and Stewart, 1995) or full-length HP1Hsα to INCENP truncation mutant INCENP45-839 could bypass the need for the protein to target to centromeres. Both chimeric proteins transferred efficiently to the spindle during anaphase (Fig. 9), despite the fact that neither was able to target to centromeres. Although results obtained with such chimeric fusions must be interpreted with caution, the fact that two different constructs both restored the spindle transfer function is consistent with the possibility that INCENP interaction with HP1 is one component of a complex priming event that INCENP normally undergoes at centromeres. Thus, these results suggest an unexpected model in which the heterochromatin protein HP1 may promote INCENP exit from the chromosomes during anaphase.
The experiments reported here have begun to characterize the mechanisms of INCENP movements in mitosis. The identification of discrete motifs required for centromere targeting and spindle transfer will be particularly useful in future screens to identify the relevant ligands that interact with INCENP at these disparate cellular locations. Evidence obtained thus far suggests that INCENP targeting to centromeres may be relatively straightforward, requiring a compact module located within the first 68 aa, but that transfer of the protein to the spindle during anaphase may be more complex. Spindle/midbody transfer is contributed to by sequences at both ends of the protein, and appears to involve substantial functional redundancy. The characterization of the INCENP:HP1 interaction and the suggestion that this interaction might be one factor involved in transfer of INCENP to the spindle have begun to reveal the mechanism by which the various movements of INCENP are regulated.
Acknowledgments
We thank Roger Brent for the gift of the materials necessary for the two-hybrid screen, Lester Binder for sanctioning the gift of TU27b mAb, and Martie Chalfie for the gift of GFP. We thank our colleagues Yasuhisa Adachi, Richard Adams, Jeremy Brown, Jeff Craig, Hiro Ohkura, Paola Vagnarelli, and Sally Wheatley for their helpful comments on the manuscript.
Abbreviations used in this paper
aa
amino acids
DAPI
4,6-diamidino-2-phenylindole
GFP
green fluorescent protein
GST
glutathione _S_-transferase
HP1Hsα and HP1Hsγ
human heterochromatin protein α and γ
INCENP
inner centromere protein
ONPG
_o_-nitrophenyl β-d-galactopyranoside
Footnotes
These experiments were supported by a grant from the Wellcome Trust, of which W.C. Earnshaw is a Principal Research Fellow.
Alexandra M. Ainsztein's present address is Laboratory of Molecular Embryology, NICHD, Building 18, Room 106, 18 Library Dr., MSC-5431, Bethesda, MD 20892-5431. Alastair M. Mackay's present address is Osiris Therapeutics, Inc., 2001 Aliceana St., Baltimore, MD 21231-2001.
References
- Aasland R, Stewart AF. The chromo shadow domain, a second chromo domain in heterochromatin-binding protein 1, HP1. Nucleic Acids Res. 1995;23:3163–3173. doi: 10.1093/nar/23.16.3168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agard DA, Hiraoka Y, Shaw PJ, Sedat JW. Fluorescence microscopy in three dimensions. Methods Cell Biol. 1989;30:353–377. doi: 10.1016/s0091-679x(08)60986-3. [DOI] [PubMed] [Google Scholar]
- Ausubel, F.M., R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl. 1991. Current Protocols in Molecular Biology. John Wiley and Sons, New York.
- Caceres A, Binder LI, Payne MR, Bender P, Rebhun LI, Steward O. Differential subcellular localization of tubulin and the microtubule-associated protein MAP-2 in brain tissue as revealed by immunocytochemistry with monoclonal hybridoma antibodies. J Neurosci. 1983;4:394–410. doi: 10.1523/JNEUROSCI.04-02-00394.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell MS, Gorbsky GJ. Microinjection of mitotic cells with the 3F3/2 antiphosphoepitope antibody delays the onset of anaphase. J Cell Biol. 1995;129:1195–1204. doi: 10.1083/jcb.129.5.1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casiano CA, Landberg G, Ochs RL, Tan EM. Autoantibodies to a novel cell cycle-regulated protein that accumulates in the nuclear matrix during S phase and is localized in the kinetochores and spindle midzone during mitosis. J Cell Sci. 1993;106:1045–1056. doi: 10.1242/jcs.106.4.1045. [DOI] [PubMed] [Google Scholar]
- Cavalli G, Paro R. Chromo-domain proteins: linking chromatin structure to epigenetic regulation. Curr Opin Cell Biol. 1998;10:354–360. doi: 10.1016/s0955-0674(98)80011-2. [DOI] [PubMed] [Google Scholar]
- Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC. Green fluorescent protein as a marker for gene expression. Science. 1994;263:802–805. doi: 10.1126/science.8303295. [DOI] [PubMed] [Google Scholar]
- Chen RH, Waters JC, Salmon ED, Murray AW. Association of spindle assembly checkpoint component XMAD2 with unattached kinetochores. Science. 1996;274:242–246. doi: 10.1126/science.274.5285.242. [DOI] [PubMed] [Google Scholar]
- Cooke CA, Heck MMS, Earnshaw WC. The INCENP antigens: movement from the inner centromere to the midbody during mitosis. J Cell Biol. 1987;105:2053–2067. doi: 10.1083/jcb.105.5.2053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Earnshaw WC, Cooke CA. Analysis of the distribution of the INCENPs throughout mitosis reveals the existence of three distinct substages of metaphase and early events in cleavage furrow formation. J Cell Sci. 1991;98:443–461. doi: 10.1242/jcs.98.4.443. [DOI] [PubMed] [Google Scholar]
- Earnshaw WC, Migeon B. A family of centromere proteins is absent from the latent centromere of a stable isodicentric chromosome. Chromosoma (Berl) 1985;92:290–296. doi: 10.1007/BF00329812. [DOI] [PubMed] [Google Scholar]
- Earnshaw, W.C., and J.B. Rattner. 1989. A map of the centromere (primary constriction) in vertebrate chromosomes at metaphase. In Aneuploidy: Mechanisms of Origin. B. Vig and M. Resnick, editors. Alan R. Liss, New York. 33–42. [PubMed]
- Earnshaw WC, Rothfield N. Identification of a family of human centromere proteins using autoimmune sera from patients with scleroderma. Chromosoma (Berl) 1985;91:313–321. doi: 10.1007/BF00328227. [DOI] [PubMed] [Google Scholar]
- Eckley DM, Ainsztein AM, Mackay A, Goldberg IG, Earnshaw WC. Chromosomal proteins and cytokinesis: patterns of cleavage furrow formation and inner-centromere protein positioning in mitotic heterokaryons and mid-anaphase cells. J Cell Biol. 1997;136:1169–1183. doi: 10.1083/jcb.136.6.1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis L, Clauser E, Morgan DO, Edery M, Roth RA, Rutter WJ. Replacement of insulin receptor tyrosine residues 1162 and 1163 compromises insulin-stimulated kinase activity and uptake of 2-deoxyglucose. Cell. 1986;45:721–732. doi: 10.1016/0092-8674(86)90786-5. [DOI] [PubMed] [Google Scholar]
- Gorbsky GJ, Sammak PJ, Borisy GG. Chromosomes move poleward in anaphase along stationary microtubules that coordinately disassemble from their kinetochore ends. J Cell Biol. 1987;104:9–18. doi: 10.1083/jcb.104.1.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gyuris J, Golemis E, Chertkov H, Brent R. Cdi1, a human G1 and S phase protein phosphatase that associates with Cdk2. Cell. 1993;75:791–803. doi: 10.1016/0092-8674(93)90498-f. [DOI] [PubMed] [Google Scholar]
- Heim R, Cibutt AB, Tsien RY. Improved green fluorescence. Nature. 1995;273:663–664. doi: 10.1038/373663b0. [DOI] [PubMed] [Google Scholar]
- Henikoff S. Unidirectional digestion with exonuclease III creates targeted breakpoints for DNA sequencing. Gene. 1984;28:351–359. doi: 10.1016/0378-1119(84)90153-7. [DOI] [PubMed] [Google Scholar]
- Lamond AI, Earnshaw WC. Structure and function in the nucleus. Science. 1998;280:547–553. doi: 10.1126/science.280.5363.547. [DOI] [PubMed] [Google Scholar]
- Lehming N, Le Saux A, Schuller J, Ptashne M. Chromatin components as part of a putative transcriptional repressing complex. Proc Natl Acad Sci USA. 1998;95:7322–7326. doi: 10.1073/pnas.95.13.7322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay AM, Ainsztein A, Eckley DM, Earnshaw WC. A dominant mutant of inner centromere protein (INCENP), a chromosomal protein, disrupts prometaphase congression and cytokinesis. J Cell Biol. 1998;140:991–1002. doi: 10.1083/jcb.140.5.991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay AM, Eckley DM, Chue C, Earnshaw WC. Molecular analysis of the INCENPs (inner centromere proteins): separate domains are required for association with microtubules during interphase and with the central spindle during anaphase. J Cell Biol. 1993;123:373–385. doi: 10.1083/jcb.123.2.373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicklas RB. The motor for poleward chromosome movement in anaphase is in or near the kinetochore. J Cell Biol. 1989;109:2245–2255. doi: 10.1083/jcb.109.5.2245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicol L, Jeppesen P. Human autoimmune sera recognize a conserved 26 kD protein associated with mammalian heterochromatin that is homologous to heterochromatin protein 1 of Drosophila. . Chromosome Res. 1994;2:245–253. doi: 10.1007/BF01553325. [DOI] [PubMed] [Google Scholar]
- Pak DTS, Pflumm M, Chesnokov I, Huang DW, Kellum R, Marr J, Romanowski P, Botchan MR. Association of the origin recognition complex with heterochromatin and HP1 in higher eukaryotes. Cell. 1997;91:311–323. doi: 10.1016/s0092-8674(00)80415-8. [DOI] [PubMed] [Google Scholar]
- Paro R, Hogness D. The Polycomb protein shares a homologous domain with a heterochromatin-associated protein of Drosophila. . Proc Natl Acad Sci USA. 1991;88:263–267. doi: 10.1073/pnas.88.1.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Platero JS, Hartnett T, Eissenberg JC. Functional analysis of the chromo domain of HP1. EMBO (Eur Mol Biol Organ) J. 1995;14:3977–3986. doi: 10.1002/j.1460-2075.1995.tb00069.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pluta AF, Saitoh N, Goldberg I, Earnshaw WC. Identification of a subdomain of CENP-B that is necessary and sufficient for targeting to the human centromere. J Cell Biol. 1992;116:1081–1093. doi: 10.1083/jcb.116.5.1081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pluta AF, Mackay AM, Ainsztein AM, Goldberg IG, Earnshaw WC. The centromere: hub of chromosomal activities. Science. 1995;270:1591–1594. doi: 10.1126/science.270.5242.1591. [DOI] [PubMed] [Google Scholar]
- Powers JA, Eissenberg JC. Overlapping domains of the heterochromatin-associated protein HP1 mediate nuclear localization and heterochromatin binding. J Cell Biol. 1993;120:291–299. doi: 10.1083/jcb.120.2.291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rappaport, R. 1996. Cytokinesis in Animal Cells. Developmental and Cell Biology Series. Cambridge University Press, Cambridge.
- Rattner JB, Rao A, Fritzler MJ, Valencia DW, Yen TJ. CENP-F is a ∼400 kD kinetochore protein that exhibits a cell-cycle dependent localization. Cell Motil Cytoskel. 1993;26:214–226. doi: 10.1002/cm.970260305. [DOI] [PubMed] [Google Scholar]
- Reynolds, A., and V. Lubland. 1989. Yeast vectors and assays for expression of cloned genes. In Current Protocols in Molecular Biology. F.A. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl, editors. John Wiley and Sons, New York. 13.6.1–13.6.4. [DOI] [PubMed]
- Rieder CL, Cole RW, Khodjakov A, Sluder G. The checkpoint delaying anaphase in response to chromosome malorientation is mediated by an inhibitory signal produced by unattached kinetochores. J Cell Biol. 1995;130:941–948. doi: 10.1083/jcb.130.4.941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rizzuto R, Brini M, Pizzo P, Murgia M, Pozzan T. Chimeric green fluorescent protein as a tool for visualizing subcellular organelles in living cells. Curr Biol. 1995;5:635–642. doi: 10.1016/s0960-9822(95)00128-x. [DOI] [PubMed] [Google Scholar]
- Saunders WS, Chue C, Goebl M, Craig C, Clark RF, Powers JA, Eissenberg JC, Elgin SC, Rothfield NF, Earnshaw WC. Molecular cloning of a human homologue of Drosophila heterochromatin protein HP1 using anti-centromere autoantibodies with anti-chromospecificity. J Cell Sci. 1993;104:573–582. doi: 10.1242/jcs.104.2.573. [DOI] [PubMed] [Google Scholar]
- Savoian, M., W.C. Earnshaw, A. Khodjakov, and C.L. Rieder. 1999. Cleavage furrows formed between centrosomes in fused PtK1 cells that lack an intervening spindle and chromosomes contain microtubule bundles, INCENP and CHO1 but not CENP-E. Mol. Biol. Cell. In press. [DOI] [PMC free article] [PubMed]
- Seeler J-S, Marchio A, Sitterlin D, Transy C, Dejean A. Interaction of SP100 with HP1 proteins: a link between the promyelocytic leukemia-associated nuclear bodies and the chromatin compartment. Proc Natl Acad Sci USA. 1998;95:7316–7321. doi: 10.1073/pnas.95.13.7316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shapiro PS, Vaisberg E, Hunt AJ, Tolwinski NS, Whalen AM, McIntosh JR, Ahn NG. Activation of the MKK/ERK pathway during somatic cell mitosis: direct interactions of active ERK with kinetochores and regulation of the mitotic 3F3/2 phosphoantigen. J Cell Biol. 1998;142:1533–1545. doi: 10.1083/jcb.142.6.1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stukenberg PT, Lustig KD, McGarry TJ, King RW, Kuang J, Kirschner MW. Systematic identification of mitotic phosphoproteins. Curr Biol. 1997;7:338–348. doi: 10.1016/s0960-9822(06)00157-6. [DOI] [PubMed] [Google Scholar]
- Taylor SS, McKeon F. Kinetochore localization of murine Bub1 is required for normal mitotic timing and checkpoint response to spindle damage. Cell. 1997;89:727–735. doi: 10.1016/s0092-8674(00)80255-x. [DOI] [PubMed] [Google Scholar]
- Wreggett KA, Hill F, James PS, Hutchinsa A, Butcher GW, Singh PB. A mammalian homologue of Drosophilaheterochromatin protein 1 (HP1) is a component of constitutive heterochromatin. Cytogenet Cell Genet. 1994;66:99–103. doi: 10.1159/000133676. [DOI] [PubMed] [Google Scholar]
- Yang CH, Tomkiel J, Saitoh H, Johnson DH, Earnshaw WC. Identification of overlapping DNA-binding and centromere-targeting domains in the human kinetochore protein CENP-C. Mol Cell Biol. 1996;16:3576–3586. doi: 10.1128/mcb.16.7.3576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye Q, Worman HJ. Interaction between an integral protein of the nuclear envelope and human chromodomain proteins homologous to DrosophilaHP1. J Biol Chem. 1996;271:14653–14656. doi: 10.1074/jbc.271.25.14653. [DOI] [PubMed] [Google Scholar]
- Ye Q, Callebaut I, Pezhman A, Courvalin J-C, Worman HJ. Domain-specific interactions of human HP1-type chromodomain proteins and inner nuclear membrane protein LBR. J Biol Chem. 1996;272:14983–14989. doi: 10.1074/jbc.272.23.14983. [DOI] [PubMed] [Google Scholar]
- Yen TJ, Compton DA, Earnshaw WC, Cleveland DW. CENP-E, a human centromere associated protein released from chromosomes at the onset of anaphase. EMBO (Eur Mol Biol Organ) J. 1991;10:1245–1254. doi: 10.1002/j.1460-2075.1991.tb08066.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zecevic M, Catling AD, Eblen ST, Renzi L, Hittle JC, Yen TJ, Gorbsky GJ, Weber MJ. Active MAP kinase in mitosis: localization at kinetochores and association with the motor protein CENP-E. J Cell Biol. 1998;142:1547–1558. doi: 10.1083/jcb.142.6.1547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu X, Chang K-H, He D, Mancini MA, Brinkley BR, Lee WH. The C-terminus of mitosin is essential for its nuclear localization, centromere/kinetochore targeting and dimerization. J Biol Chem. 1995;270:1–6. doi: 10.1074/jbc.270.33.19545. [DOI] [PubMed] [Google Scholar]