Immunity and immunopathology to viruses: what decides the outcome? (original) (raw)
Key Points
- Humans are constantly exposed to numerous viruses but the consequences of infection are different in different individuals.
- The outcome of host–viral interactions depend on the dose and route of infection, viral virulence properties, as well as several host factors that mainly involve innate and adaptive immunity.
- Host factors that influence the outcome of viral infection include genetic factors, such as polymorphism in MHC alleles, mutations in genes encoding innate receptors, cytokines, chemokine receptors, age, the nature of endogenous and persistent infections and pre-exposure to other infections.
- The host has numerous anti-inflammatory activities that limit the extent of tissue damage caused by infections.
- Successful pathogens such as HIV, hepatitis B virus (HBV), HCV and herpesviruses persist either as chronic or latent infections in the host with or without causing immediate ill effects; however, they may have lethal consequences when the host is immunocompromised.
- Viruses that cause chronic infection influence immune cells to produce predominantly anti-inflammatory cytokines, such as interleukin-10 (IL-10) and transforming growth factor-β (TGFβ), and they upregulate inhibitory receptors on effector T cells giving them an exhaustion phenotype.
Abstract
Many viruses infect humans and most are controlled satisfactorily by the immune system with limited damage to host tissues. Some viruses, however, do cause overt damage to the host, either in isolated cases or as a reaction that commonly occurs after infection. The outcome is influenced by properties of the infecting virus, the circumstances of infection and several factors controlled by the host. In this Review, we focus on host factors that influence the outcome of viral infection, including genetic susceptibility, the age of the host when infected, the dose and route of infection, the induction of anti-inflammatory cells and proteins, as well as the presence of concurrent infections and past exposure to cross-reactive agents.
Similar content being viewed by others
Main
Humans may be infected by and suffer clinical consequences from numerous different viruses, and in most instances the infection is resolved with or without tissue damage. Reinfection is usually subclinical, and for many viruses we have effective vaccines; classic examples include measles, mumps, rubella, rotavirus and varicella zoster viruses. Other viruses, such as HIV, hepatitis C virus (HCV), hepatitis B virus (HBV) and some herpesviruses, can cause substantial tissue damage in some or all individuals they infect, and lesions can become chronic. These viruses usually have one or more properties that allow them to diminish the efficacy of host adaptive or innate immunity, and we lack effective vaccines against most of these agents. Infection with viruses such as influenza virus and respiratory syncytial virus (RSV) has a variable outcome. Most individuals may suffer mild or subclinical infection, but others experience severe disease that can be lethal. Of particular interest are agents such as West Nile virus, dengue virus and poliovirus that can cause severe disease, but only in a small minority of infected people. Finally some viruses, such as coxsackie B virus, human T lymphotropic virus, Epstein–Barr virus (EBV) and possibly rubella virus, are thought to act as triggering agents for autoimmune diseases and cancer in genetically susceptible individuals. The topic of virus-associated complications has been reviewed by others1,2 and will not be discussed here.
In this Review, we discuss our current understanding of the circumstances of infection and host-controlled factors that could explain why an infection can be resolved with minimal impact or cause substantial tissue damage. Understanding such issues could prove to be useful in the future for the control and perhaps prevention of tissue-damaging virus infection.
Tissue damage caused by the immune system
Almost all virus infections cause the recruitment and activation of inflammatory cell types — particularly macrophages and, in some infections, neutrophils — that in turn release a range of molecules that induce tissue damage or malfunction. These include cytotoxic cytokines, cationic proteins, lipid mediators, metalloproteinases and components of the oxygen burst. The reactive oxygen species that accumulate in the mitochondria may further contribute to tissue damage3. Both innate and adaptive immune signalling events are involved in mediating tissue damage.
Contribution by innate immune responses. Invading viruses and their replicative intermediates can be recognized by several innate immune receptors expressed either at the host cell surface or within cells. Many types of innate immune receptor can participate in the immune response but, in virus infections, the most studied are the Toll-like receptors (TLRs), retinoic acid-inducible gene I (RIG-I; also known as DDX58) and NOD-like receptors (NLRs). Virus infections usually activate the endosomal TLRs (TLR3, TLR7, TLR8 and TLR9) that recognize viral nucleic acids and double-stranded RNA intermediates4. The cytoplasmic RIG-I-like receptors recognize viral genomic RNA or RNA encoded by genomic DNA, whereas the NLRs recognize viral DNA genomes (reviewed in Ref. 5). In general, activation of many of these receptors causes the production of pro-inflammatory cytokines and interferons (IFNs), as well as signals that recruit and activate cells involved in inflammation and the induction of adaptive immunity. The pattern of innate immune events induced after the entry of virus may dictate the outcome of infection. Many viruses that persist trigger innate cells such as dendritic cells (DCs), natural killer (NK) cells and macrophages to produce anti-inflammatory molecules such as interleukin-10 (IL-10) and transforming growth factor-β (TGFβ). For example, DCs from lymphocytic choriomeningitis virus (LCMV)-infected mice produce high levels of IL-10 (Ref. 6), and IL-10 is produced by monocytes from individuals infected with HIV, HCV or HBV7,8,9. Following RSV infection, the interaction between the virus and lung plasmacytoid DCs (pDCs) is crucial, as the removal of pDCs before infection favours an immunopathological reaction in the lungs10. A damaging response to a virus infection is more likely to occur with viruses that can interfere with one or more innate defences. Some examples of viruses that have this effect and the innate defence mechanism that is diminished are listed in Table 1.
Table 1 Virus and host features that favour tissue damage
Contribution by adaptive immune responses. Once adaptive immune effector cells are generated, these can contribute to tissue damage. T cells, for example, can directly destroy virus-infected cells or release cytokines, such as tumour necrosis factor (TNF), that damage cells. With some non-cytopathic virus infections, such as HCV and HBV, destruction of infected cells by CD8+ effector T cells is the main cause of damage to the liver11,12 (Box 1; Fig. 1). Responses to infected cells by different types of CD4+ T cell orchestrate a tissue-damaging inflammatory reaction and these become chronic against persistent viruses. Most often, the cell subsets involved are T helper 1 (TH1) cells, but TH17 cells may contribute to inflammatory responses during HIV, HCV and influenza virus infections13,14,15. In such TH17 cell-driven responses, neutrophils are recruited and become a major source of tissue-damaging molecules. TH2 cells are rarely associated with inflammatory responses during viral infections, but a TH2 cell response can occur during severe lung responses to RSV infection16.
Figure 1: Immunity or immunopathology following viral infection.
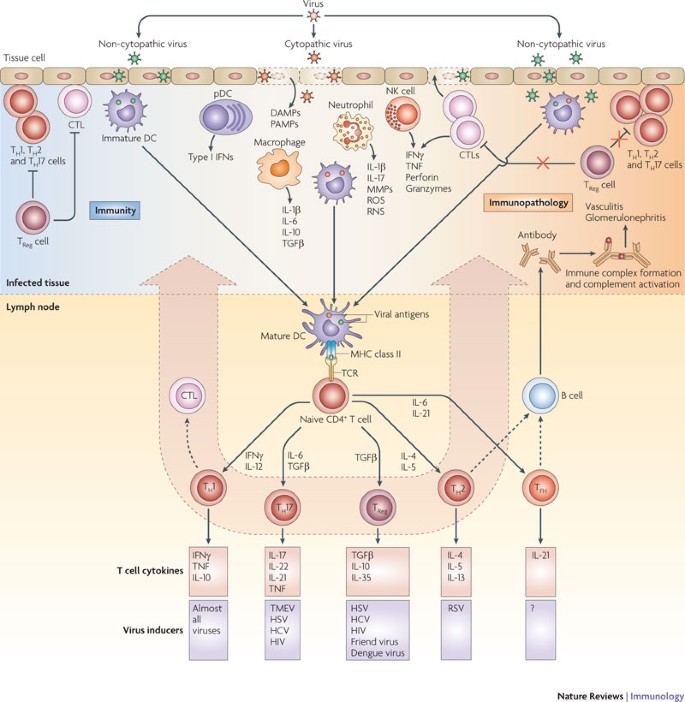
Following entry into host cells, viruses (cytopathic or non-cytopathic) replicate at the site of infection. Cytopathic viruses kill infected cells, causing the release of cellular contents, including proteases and lysosomal enzymes, which digest the extracellular matrix and create an inflammatory milieu. Neutrophils that are rapidly recruited to the site of infection release inflammatory mediators. Innate cells recognize viral replication intermediates and secrete pro-inflammatory cytokines, which, in addition to helping to clear the virus, contribute to tissue damage. Viral antigens are taken up by antigen-presenting cells and carried to local draining lymph nodes. Depending on the cytokine milieu created in the draining lymph node, different types of T helper (TH) cell responses are induced. Primed CD8+ cytotoxic T lymphocytes (CTLs) migrate to the site of infection and kill virally infected cells, thereby contributing to tissue damage. After migrating to the site of infection, TH cells also contribute to the tissue damage. In conditions in which the control of aggressive TH cells and CTLs by regulatory T (TReg) cells is impaired and other inhibitory pathways fail to curtail them, tissue damage is the main consequence of viral infection. TH cells also provide help to B cells to secrete antibodies, which form immune complexes that are deposited in certain tissues such as the glomeruli of the kidneys and blood vessels to cause immune complex-mediated disease. DAMP, danger-associated molecular pattern; DC, dendritic cell; HBV, hepatitis B virus; HCV, hepatitis C virus; HSV, herpes simplex virus; IFN, interferon; IL, interleukin; MMP, matrix metalloproteinase; NK, natural killer; PAMP, pathogen-associated molecular pattern; pDC, plasmacytoid DC; RNS, reactive nitrogen species; ROS, reactive oxygen species; RSV, respiratory syncytial virus; TCR, T cell receptor; TFH, T follicular helper; TGFβ, transforming growth factor-β; TMEV, Theiler's murine encephalomyelitis virus; TNF, tumour necrosis factor.
Antibody responses to viruses may also contribute to tissue damage. This occurs when antibody binds to an infected cell, activates complement and causes an inflammatory reaction. Alternatively, antibody-mediated inflammatory reactions involve toxicity following engagement of IgG with Fc receptors on inflammatory cells, which causes inflammatory mediator release17, or following deposition of viral antigen–antibody complexes in capillary beds, leading to activation of the complement cascade. Lesions formed by immune complex deposition can occur when viruses persist and poorly neutralizing IgG is produced; lesions include nephritis, polyarteritis and arthritis. Immune complex lesions were first reported in LCMV infection but have also been reported in chronic HCV and HBV infection, as well as in idiotypic IgA nephropathy associated with HIV infection18,19,20,21. Viruses such as RSV express antigens that may induce an IgE response and type I hypersensitivity might partially account for lung lesions in some children infected with RSV22.
Despite these possible occurrences, tissue damage is modest in most cases of viral infections; however, the level of tissue damage can vary between individuals infected with the same virus. In the following section, we describe the host responses that function to minimize tissue damage.
Host factors that limit tissue damage
The host can use many countermeasures to limit tissue damage after virus infection. These tissue-protective events are more effective against some viruses and some circumstances of infection than others, accounting for the different pattern of response observed among individuals. The countermeasures include the production of cytokines such as IL-10 and TGFβ that have anti-inflammatory activity, other host-derived anti-inflammatory mediators such as resolvins and galectins, the activity of cell subsets that inhibit other cells from mediating inflammatory events, as well as the induction of molecules on effector cells that result in the loss of effector functions (Fig. 2).
Figure 2: Inhibitory mechanisms to limit tissue damage caused by T cells.
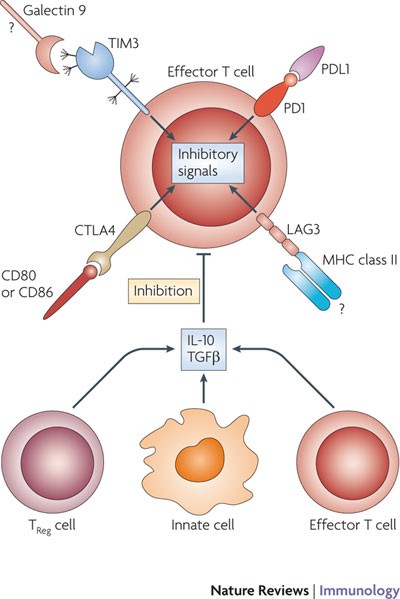
Effector T cells upregulate inhibitory receptors such as programmed cell death 1 (PD1), T cell immunoglobulin domain and mucin domain protein 3 (TIM3), lymphocyte activation gene 3 (LAG3) and cytotoxic T lymphocyte antigen 4 (CTLA4) (and others such as adenosine receptors (not shown)) on their surface. Ligation of these receptors with PDL1, galectin 9, MHC class II molecules and CD80 or CD86, respectively, delivers inhibitory signals to the effector T cells and controls their inflammatory activity and subsequent tissue damage. In addition, activated regulatory T (TReg) cells, specialized innate cells or highly polarized effector T cells that can produce anti-inflammatory cytokines inhibit effector T cell responses. Inadequate control exerted by these pathways under some circumstances therefore results in uncontrolled T cell activation and proliferation causing excessive tissue damage. Question marks indicate interactions for which extensive in vivo studies have not been carried out. IL-10; interleukin-10; TGFβ, transforming growth factor-β.
Anti-inflammatory cytokines. The two best-known cytokines that inhibit inflammatory reactions are IL-10 and TGFβ. Many cell types can produce IL-10, including subsets of activated DCs, macrophages (when infected with some viruses), activated regulatory T (TReg) cells, B cells and some subsets of NK cells following stimulation with TLR ligands23. Although some viruses, such as EBV and cytomegalovirus (CMV), produce an IL-10-like molecule that functions in vitro in a similar manner to host IL-10, it is still not clear if this viral IL-10 has any role during viral infections in vivo24,25. Host-derived IL-10 can block pro-inflammatory cytokine and chemokine production and MHC class II expression, and it can also interfere with many signalling pathways that result in pro-inflammatory cytokine production. For example, nuclear factor-κB (NF-κB) signalling is inhibited by IL-10-mediated induction of p50 and p105 NF-κB subunits, one of which (p50) binds to the stimulatory subunit p65, sequestering it in the cytoplasm and making it unavailable for binding to the promoters of IL-6 and CC-chemokine ligand 3 (also known as MIP1α) for their transcription (reviewed in Ref. 26). IL-10 also suppresses type I IFN-induced tyrosine phosphorylation of signal transducer and activator of transcription 1 (STAT1) and also induces the expression of suppressor of cytokine signalling 3 (SOCS3) by macrophages and neutrophils, which impairs their inflammatory activity.
The extent of IL-10 production during an infection could influence the amount of tissue damage that occurs. Thus, if the IL-10 response is absent, owing to genetic mutation, or is artificially suppressed by antibodies specific for IL-10 or its receptor, inflammatory reactions to infectious agents may be exaggerated. For example, mice lacking an IL-10 response develop more severe inflammatory reactions to ocular infection with herpes simplex virus (HSV) than do normal controls27. IL-10 may be of particular importance to constrain the severity of inflammatory reactions caused by chronic infections, and its anti-inflammatory function has been shown during infection with viruses such as HCV and HIV9,28,29.
Recently, IL-10 production by virus-specific effector T cells during the acute response to influenza virus was shown to be responsible for minimizing the severity of pulmonary lesions in mice30. The responding CD8+ T cells showed plasticity and gained the ability to produce IL-10 in addition to their main effector product, IFNγ. Inhibiting the response to IL-10 with antibody specific for the IL-10 receptor resulted in more severe and sometimes fatal virus-induced lung damage. IL-10 production by effector CD8+ T cells and the IL-10-induced suppression of IL-2 production by these cells were also shown in HIV-infected patients31. Suppression of the effector T cell IL-10 response by lethal strains of influenza virus, such as the 1918 H1N1 strain and the H5N1 strain, is thought to contribute to their virulence32. In some instances excessive IL-10 production during a virus infection may inhibit a protective effector T cell response and favour viral persistence6. This can happen in some circumstances with LCMV infection and may also occur during HIV infection6,9.
The TGFβ superfamily of cytokines has a similar anti-inflammatory role to IL-10, although the effects of TGFβ are more complex. These cytokines have a wide range of activities that include both anti-inflammatory and pro-inflammatory effects with the outcome depending on the concentration of TGFβ available and some other factors33. Moreover, TGFβ superfamily members exist in a latent, inactive form and must be cleaved before they can bind to receptors and mediate their effects on cells. Nevertheless, the extent of TGFβ production during virus infection could influence whether the response becomes overtly tissue damaging. Accordingly, TGFβ inhibits several functions of T cells, including proliferation, differentiation into effector T cells and some effector functions such as cytotoxicity34. The inflammatory activities of CD8+ T cells, TH1 cells and TH17 cells, as well as the production of inflammatory products by recruited cells, are all inhibited by TGFβ34.
Most of our knowledge of the role of TGFβ in microbial pathogenesis concerns non-viral pathogens35,36. However, some viruses do cause an increase in TGFβ levels, and other viruses express proteins that can cleave and activate TGFβ. For example, influenza virus neuraminidase can activate TGFβ, and the extent to which this happens could influence virulence37. Among the viruses that cause increased TGFβ production are chronic infections by HBV and HCV. A nonstructural protein of HCV (NS4) was shown to be responsible for TGFβ induction and it seems that the magnitude of the TGFβ response, with IL-10 production, could determine if HCV infection is effectively cleared or becomes chronic14. Acute infection with reovirus may also activate TGFβ signalling to an extent that correlates with damage to the central nervous system38. Of note, virus-specific CD8+ T cells isolated from HCV-infected individuals produce TGFβ that suppresses virus-specific T cell responses39. Moreover, blockade of TGFβ enhanced the in vitro activity of T cells39. Furthermore, a recent report attributed an intrinsic role of TGFβ signalling in effector T cells to explain their diminished survival and effector functions during chronic LCMV infection40.
IL-10 and TGFβ are not the only cytokines that can limit damage caused by inflammatory reactions. Recently, for example, IL-17 — a cytokine that is normally associated with the promotion of tissue damage — was shown to have an anti-inflammatory role by suppressing TH1 cell-mediated inflammatory effects41. The suppressive effect of IL-17 was noted initially with autoimmune lesions but was recently also observed during infection with Theiler's murine encephalomyelitis virus in mice42.
Other anti-inflammatory molecules. Several other natural host products can also participate in the control and resolution of inflammatory reactions (Fig. 3). These include the galectins, resolvins and protectins. These molecules contribute to the resolution of inflammatory lesions in several non-infection-related lesions43. Members of the galectin family might also help to constrain inflammatory reactions43. Galectin 9, for example, binds to the T cell immunoglobulin domain and mucin domain protein 3 (TIM3; also known as HAVCR2) on activated effector T cells and causes apoptosis, and at the same time it expands the TReg cell response44,45,46; both effects minimize tissue damage. It is possible that variations in galectin concentrations between individuals could explain why the outcome in some individuals is rapid and protective but in others it is prolonged and causes tissue damage. Currently, there is no evidence for this hypothesis; however, galectin levels are different between HIV- and HCV-infected individuals and controls, and in HCV infection galectin levels may correlate with viral loads47,48.
Figure 3: Balance between pro-inflammatory and anti-inflammatory mechanisms may decide the outcome of viral infection.
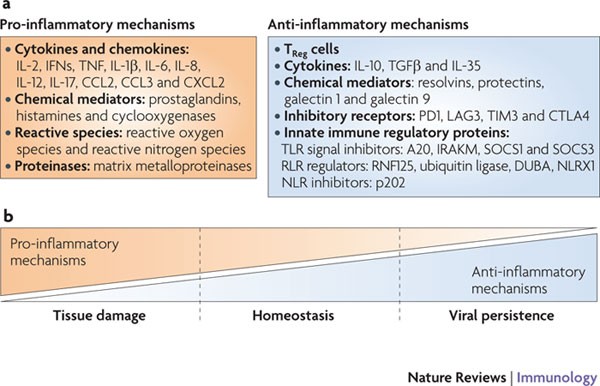
a | Pro-inflammatory and anti-inflammatory mechanisms induced after viral infection. b | The balance between immunity and immunopathology following viral infection might depend on the levels of anti-inflammatory and pro-inflammatory mechanisms. A balanced combination of pro-inflammatory and anti-inflammatory mechanisms would facilitate viral clearance and immunity to reinfection, with minimal damage to host tissues. An excess of pro-inflammatory mechanisms would ensure viral clearance but causes tissue damage. If anti-inflammatory mechanisms outweigh pro-inflammatory mechanisms, the pathogen could persist in the host as a subclinical infection, an opportunist or a tissue-damaging agent. CCL2, CC-chemokine ligand 2; CTLA4, cytotoxic T lymphocyte antigen 4; CXCL2, CXC-chemokine ligand 2; DUBA, deubiquitinase enzyme A; IFN, interferon; IL, interleukin; IRAKM, IL-1R-associated kinase M; LAG3, lymphocyte activation gene 3; NLRX1, NOD-like receptor X1; PD1, programmed cell death 1; RNF125, ring finger containing domain 125; RLR, RIG-I-like receptor; SOCS, suppressor of cytokine signalling; TGFβ, transforming growth factor-β; TIM3, T cell immunoglobulin domain and mucin domain protein 3; TLR, Toll-like receptor; TNF, tumour necrosis factor; TReg, regulatory T.
T Reg cells. Another mechanism of counteracting excessive tissue damage following virus infection is the induction, activation or expansion of several types of TReg cell, the main activity of which is to inhibit the function of other cell types. The best-studied TReg cells are CD4+ T cells that express the transcription factor forkhead box P3 (FOXP3)49. Such TReg cells could influence the outcome of infection, particularly those that are chronic in nature50. In model systems in which the activity of TReg cells can be inhibited, tissue-damaging immunopathological reactions to some viruses are increased. This was observed with the HSV ocular immunopathology model in mice51. Similar tissue-protective effects of TReg cells were noted in a Friend retrovirus model, as well as in mouse models of RSV and West Nile virus52,53. In the HSV ocular immunopathology model, recent observations showed the therapeutic value of increasing the proportion of FOXP3+ TReg cells to limit virus-induced immunopathological reactions54,55. This was achieved by administering reagents, such as galectin 9 and the fungal metabolite FTY720, that could cause some conventional CD4+ T cells to convert in vivo into FOXP3+ CD4+ T cells that have regulatory functions45,55. In several chronic human virus infections, TReg cells are thought to influence the extent of tissue damage, but this view remains debatable and is difficult to evaluate in natural disease situations56. Some of the most convincing evidence that TReg cells are beneficial in a human chronic viral infection came from observations of the disease outcome in patients accidentally infected with HCV. In this example, it was shown that a favourable outcome was more likely to occur in those individuals that made the highest IL-10-producing TReg cell responses57.
The role of inhibitory receptors. An additional mechanism that could affect the pattern of events that follow virus infection is the signalling of inhibitory receptors on effector cells of both innate and adaptive immune systems. Such events may favour tissue damage over infection control. Several negative signalling systems that affect innate immune responses to virus infections have been noted, and many of these function by terminating NF-κB signalling, which curtails pro-inflammatory cytokine and chemokine production58 (Box 2).
The protective function of effector T cells may also be compromised if they express inhibitory receptors and engage their ligands. The circumstances that result in upregulation of inhibitory receptors by effector T cells is not fully understood at a mechanistic level, but the effect usually becomes evident during chronic infections, especially those that involve high levels of persistent antigen59. Some of the well-characterized receptors that can be induced in chronic infection include programmed cell death 1 (PD1) and the IL-10 receptor. These molecules are expressed at higher levels by CD8+ T cells during chronic infection with LCMV in mice6,60. Their engagement with their respective ligands impairs effector T cell function, resulting in a so-called exhaustion phenotype. This exhaustion phenotype for effector T cells has since been observed in other chronic infections, including HIV, HCV and HBV61,62,63. Some recent studies have suggested the existence of a correlation between the PD1–PD1 ligand 1 (PDL1) pathway and the production of IL-10. Thus, triggering of PD1 by PDL1 induced high levels of IL-10 production in monocytes that in turn inhibited the function of CD4+ T cells64. The IL-10-mediated inhibition of CD4+ T cell effector function was also shown in chronic LCMV infection in mice6. Exhausted T cells may express additional inhibitory receptors that include lymphocyte activation gene 3 (LAG3) and TIM3, with distribution varying in different locations59. Ligation of inhibitory receptors on exhausted T cells diminishes their protective function, allowing viruses to persist and cause more tissue damage. In fact, the TNF-producing ability of exhausted T cells that contributes to tissue damage is one of the last functions to wane59. It is not clear how infection causes inhibitory receptor upregulation but in some instances a viral component has been implicated (for example, the HCV core protein) and in other cases virus-induced IFNs63,65 or TLR ligands expressed by viruses are thought to have a role66.
The discovery of the exhaustion phenomenon provides an enticing opportunity for the development of new approaches for improving immunity and controlling the severity of chronic viral infections. Accordingly, blockade of the signals that are responsible for the exhausted state can result in partial recovery of immune function and more effective infection control6,60. Initial observations focused on blockade of PD1 or PDL1, as well as the IL-10 receptor, using monoclonal antibodies, but blockade of several inhibitory signals was even more efficacious than blockade of a single component67,68. Thus, inhibition of either the PD1–PDL1 interaction and the IL-10 receptor or the PD1–PDL1 interaction and LAG3 provided the best approach to achieve immune recovery and viral control after chronic LCMV infection in mice. Furthermore, when both blockade of the exhaustion pathways and therapeutic vaccinations were carried out, control of chronic LCMV infection was greater than either approach alone69,70. It is likely, therefore, that similar strategies might prove to be useful for the control of human chronic infections.
Factors that favour tissue damage
In this Review, we explore the question of why virus infection has varied outcomes in different individuals infected with the same virus. This issue is particularly difficult to address in the context of sporadic diseases that cause overt disease in only a small minority of infected persons. The classic example of this is paralytic polio, which affects less than 1% of individuals infected with poliovirus71. Reasons for susceptibility to poliovirus-associated disease remain unexplained but one favoured hypothesis is that the involvement of the central nervous system (CNS) is associated with a defect in type I IFN production, allowing robust poliovirus replication in the periphery and spread of the virus to the CNS71. Another example is herpes simplex encephalitis that occurs in adults. This is a rare, often lethal, debilitating disease caused by infection with HSV-1 (Ref. 72). The disease usually occurs following reactivation of the host's resident latent virus, although primary infection can occasionally result in encephalitis and is a common outcome in seronegative neonates infected with HSV-2. Lesions of herpes simplex encephalitis in adults are, in part, immune mediated, but why they only occur in an unfortunate few is not known. Primary genetic susceptibility factors have not been implicated nor has the emergence of a neurotropic mutant, although recently a minority of cases of herpes simplex encephalitis in children were associated with polymorphism in the genes encoding TLR3 or UNC93B (which is involved in TLR signalling)73,74. In the following section we describe the situations that favour a tissue-damaging response over immune control (Table 1).
Age of infection. Whether a virus causes severe tissue damage often depends on the age at which infection occurs. For example, intrauterine infections may result in severe tissue damage, as happens following rubella virus and CMV infections in humans75. In general, it is the young and the elderly who suffer the most severe consequences of infection, as shown by increased morbidity and mortality among these age groups following infection with seasonal influenza virus76. Increased susceptibility of the young, especially neonates, has been attributed to immature responsiveness of the immune system, particularly components of innate immunity. For example, RSV is usually the first pathogen that a human infant encounters and clinical signs of RSV infection are common by 2–3 months of age77. Premature infants are particularly prone to develop severe lesions in the respiratory tract following RSV infection and these account for many of the 3–5% of childhood RSV infections that require hospitalization77,78. This high susceptibility of infants to RSV-induced immunopathology is mainly explained by an inadequate type I IFN response together with a failure to activate subsets of DCs that induce CD8+ T cell- and TH1 cell-mediated protective responses79. Instead, the DCs that are stimulated tend to produce IL-10 and TGFβ and induce the generation of FOXP3+ TReg cells80. Studies of RSV infection using animal model systems indicate that the main mediators of the lung pathology are T cells, but there is still debate as to which subsets are primarily involved. Some studies indicate a pathological role for TNF-producing CD8+ T cells, whereas other studies advocate that lesions are associated with TH2 cell-dominated responses16 (reviewed in Ref. 78). Because childhood infection with RSV is a notable health problem, against which there is no effective vaccine, understanding at a mechanistic level why some individuals develop severe lesions to RSV is an important issue.
Aged individuals may also suffer more problems than younger individuals following primary or secondary infection with some viruses. For example, re-exposure of aged individuals to RSV can cause lesions similar to those that occur in infants81. In addition, aged individuals may develop lesions following reactivation of latent infections that had been successfully controlled by their immune system for decades. The best example of this is shingles, which is characterized by painful inflammatory skin lesions that occur at sites innervated by a sensory ganglion in which varicella zoster virus has been reactivated from latency82. It is not clear why this happens in only a few of the many ganglia that contain latent virus. It is possible that reactivation of latent virus occurs often but is successfully controlled by T cell immunity83. Presumably this T cell activity can fail at some sites but not others; further study is required to understand how this happens.
Understanding senescence of the immune system is an active area of research and the topic has been covered in excellent reviews84,85. Studies, mainly done in mice, have revealed that ageing has a greater effect on primary than on memory immune responses84. The T and B cells from aged individuals respond less well to antigens and are compromised in their ability to carry out effector functions compared with responses in young individuals. Moreover, naive T cells from aged compared with young animals undergo less homeostatic proliferation, probably because of competition with memory T cells for growth factors and anatomical space84. An additional effect observed during senescence is that the breadth of the T cell repertoire is reduced, perhaps because the thymus has involuted84. Indeed, compared with young individuals, aged individuals often have increased numbers of CD8+ T cells that recognize latent viral infections, particularly CMV infections — an effect known as memory inflation86. These CMV-specific CD8+ T cells can account for 50% or more of the total CD8+ memory T cell population. The space such cells occupy could diminish the number of lymphocytes available to react to pathogens, perhaps explaining why infections such as influenza virus are more severe in the elderly.
Dose and route of infection. As can readily be shown in experimental systems, the dose and route of transmitted virus can markedly influence the outcome of a virus infection. Minimal doses may be controlled subclinically by innate defences and may be insufficient to induce adaptive immune responses. Massive doses can overwhelm immune defences and cause severe disease and rapid death, in some instances by direct cytotoxic effects of viral components. Doses between these extremes can have a variable outcome from undetectable infection to tissue-damaging lesions. This issue has received surprisingly little formal study with virus infections, but it is a common practice for those who study viral pathogenesis to choose optimal doses of infection to evaluate their concepts. The dose and route of infection is expected to influence how successful the virus will be in gaining access to susceptible target cells and being transported by different types of DC to lymphoid tissues. Thus, it is clear from studies with protein antigens that the magnitude and quality of immune responses induced is influenced by the type of DCs that engage antigen87. With replicating agents such as viruses, dose effects might depend on the type of virus, the rates of replication and the virus-intrinsic properties such as expression of ligands for host innate immune receptors. For example, the dose of lentiviruses seems to have little effect, perhaps because potent infection is usually established by a single (founder) virion from the large numbers of virions (up to 106) that are transmitted to the host88. Experimental studies of simian immunodeficiency virus (SIV) infection of macaques also show little or no effect of dose on the outcome of infection89. By contrast, for cytopathic viruses, the dose of infection can influence the response pattern90. It is curious that for influenza virus, the dose of infection of mice also affects the range of cells that become infected, as well as the balance of the immune response that results91. At high doses of virus, DCs and alveolar epithelial cells become infected. Moreover, the infected DCs deliver apoptotic signals to CD8+ T cells, which are normally necessary for resolving infection. At lower doses of virus, DC infection does not occur and the protective CD8+ T cell response is not compromised.
Recently, an interesting and unexpected effect of infection dose was observed in chimpanzees infected with HBV92. HBV is a non-cytopathic virus, and both control of the virus and the development of immunopathological hepatic lesions are mediated by CD8+ T cells. Over a wide dose range (104–108 virions), the outcome of infection was similar, being controlled successfully 6–8 weeks after infection with minimal hepatitis. However, after administering a large dose (>108) of virus, 100% of hepatocytes became infected, the virus reached high levels for at least 16 weeks and the animals developed chronic active hepatitis. Unexpectedly, animals infected with a very low dose (100 virons or less) had a comparable immunopathological outcome to those infected with a large dose. Reasons for this are still uncertain, but it is possible that at low doses the virus can go undetected by the immune system and fail to induce priming of CD4+ T cells, which are needed to provide help to mount a protective CD8+ T cell response. Furthermore, if animals were depleted of CD4+ T cells and given the medium dose of virus they developed chronic active hepatitis. How these observations relate to HBV infection of humans needs evaluation.
Dose of infection might explain the variable outcome observed following infection by insect-transmitted flaviviruses. The flavivirus West Nile virus is becoming a common infection in the Western hemisphere but most infected individuals do not suffer any clinical consequences. However some individuals develop a life-threatening meningoencephalitis. In such individuals, the virus crosses the blood–brain barrier and lesions develop as a consequence of a viral antigen-specific T cell-mediated immunopathological reaction to infected cells93. One hypothesis explaining the pathological outcome is that it occurs in circumstances in which the virus replicates rapidly and exceeds the host's ability to constrain it, despite the induction of a neutralizing antibody response and numbers of functional T cells that would otherwise be protective93. This scenario is more likely to occur in those exposed to many mosquito bites (providing a high-dose infection), especially if the infected person is young, aged or has some degree of immunodeficiency. Curiously, some flaviviruses, including West Nile virus, stimulate TLR3 (which recognizes double-stranded DNA) and induce TNF production, and this may increase the permeability of the blood–brain barrier and allow entry of the virus to the CNS94.
The route of infection can also affect the extent of tissue damage that ensues. For example, with HSV infection of humans, oral or genital infection usually results in lesions that resolve without long-term damage. However, infection of the eye can result in chronic inflammatory lesions that cause blindness95. Similarly, mice infected with corona virus or Theiler's murine encephalomyelitis virus can develop immunopathological lesions in the CNS, but this response only occurs if the virus is given intracerebrally or intranasally and fails to occur after systemic infection96,97. The meningitis that adult mice develop after LCMV infection occurs only if the virus is administered directly into the cerebrospinal fluid and is not evident if administered by other routes98.
Influence of heterologous immunity. Heterologous immunity is the term used to describe the observation that exposure to one pathogen will generate an immune response against numerous antigenic epitopes derived from that pathogen some of which might cross-react with epitopes derived from other pathogens. Following infection with the second pathogen, the cross-reactive memory cells expand more rapidly and may dominate the overall response. However, this cross-reactive response may be of low avidity and poorly protective but may still be able to mediate tissue damage. These circumstances could explain the occurrence of severe disease (dengue haemorrhagic fever (DHF)) in a small minority of the 50 million people who are infected with dengue virus. DHF is characterized by a high fever, vascular leakage, hypotension, circulatory shock and some bleeding manifestations. It usually occurs in individuals who are already immune to one dengue virus strain and become infected with a heterologous strain. An initial explanation for the DHF syndrome was that it occurred because cross-reactive, but non-neutralizing, antibodies opsonized the virus, facilitating uptake by macrophages99. This was proposed to result in immune activation, abundant production of cytokines and vascular leakage. Other investigators now suggest that T cells are the main orchestrators of the disease and that they cross-react with epitopes primed by exposure to a different viral strain99. The expanded T cell populations are not protective but they can mediate a severe inflammatory reaction that includes the production of vascular endothelial growth factor A (VEGFA), the main mediator of vascular leakage. DHF does not occur as often as might be expected on the basis of this mechanism, which suggests that additional factors are also involved. These include viral virulence, the ability to replicate efficiently and host genetic factors. Clinical and experimental evidence for the proposed pathogenesis of DHF is reviewed in Ref. 100.
Another tissue-damaging human disease that could be explained by the existence of heterologous immunity is infectious mononucleosis. The disease is debilitating and can last for weeks but it only occurs in a minority of usually young adults who have experienced a primary infection with EBV. In patients that develop infectious mononucleosis, there is a 5–20 fold increase their circulating T cells, most of which are viral antigen-specific CD8+ T cells. The pathogenesis of infectious mononucleosis is poorly understood but one provocative hypothesis is that the disease is the consequence of heterologous immunity, with infectious mononucleosis only occurring in individuals with EBV-cross-reactive T cells at the time of infection101. Such cross-reactive T cells are proposed to expand more rapidly than EBV-specific naive T cells induced following infection. However, the cross-reactive cells that dominate the response have low affinity for the virus antigen-expressing cells and cannot adequately control the infection, thereby setting the stage for chronic immunopathology. Supporting this concept, cross-reactivity between the influenza virus matrix protein and the EBV BMLF1 protein has been noted102. Experimental evidence that heterologous immunity can account for immunopathological responses in some but not all animals was provided by Kim and colleagues103. Other examples of cross-reactivity between different pathogens that influence disease patterns are reviewed in Ref. 104.
Host genetics and the 'virome'. Variation in clinical responses of individuals to virus infections is influenced by the host genotype. Virulent infectious agents are assumed to have helped to shape our genome and are responsible for the extreme polymorphisms of many loci involved in MHC antigen processing and presentation105. There is evidence that the outcome of the virus infection is affected by the HLA alleles expressed106, but the influence is usually modest. Thus, resistance to pathogens, including viruses, is directed by several genes that act at different stages of the virus–host interaction. For example, at least 250 genes are estimated to affect the outcome of infection by HIV107. As a result, the absence or malfunction of a single gene would probably have a negative or minimal effect on the outcome of a virus infection. Exceptions to this generalization include the primary immunodeficiencies that arise from rare monogenic defects and lead to increased susceptibility to various virus infections108. Mutations of genes encoding viral receptors and co-receptors can also influence disease susceptibility. For example, individuals with a homozygous 32 base pair deletion in CC-chemokine receptor 5 (CCR5, the entry co-receptor for X5 HIV) have greater resistance to HIV than heterozygous individuals109. Another example is that non-secretors of ABO blood groups are refractory to diarrhoea caused by Norwalk virus110. Mutations in genes that encode proteins involved in innate defence, such as the TLRs, can affect the clinical expression of some infections, but so far little has been reported for viruses. One example is a mutation in UNC93B, which encodes a transmembrane protein involved in TLR signalling111. Another is a loss of function mutation of TLR3 (Ref. 74). Both defects result in defective cytokine production, especially IFNα and IFNβ, in response to infection with HSV-1. Affected children develop herpes simplex encephalitis74,112 but, curiously, not other problems with HSV infection or other viruses.
A more common scenario that accounts for more severe disease following virus infection is polymorphisms in a few or many genes. Investigations on this topic have focused on HIV (reviewed in Refs 106, 113) but most changes detected so far have been minor, probably because several genes affect resistance. Genetic factors have not yet been shown to explain the finding that ∼1% of HIV-infected patients control their infection long term without treatment114.
In addition to host genetics affecting the pattern of disease, other sources of genetic material in the host could influence the outcome of infection. These are the endogenous retrovirus elements, some of which are transcriptionally active, that are estimated to contribute to 8–9% of total human DNA115. The other sources are exogenous viruses that establish persistent infections and are mainly not retroviruses. According to a recent review, every individual may harbour several (8–12) different chronic asymptomatic viral infections that lie undetected59. Together these endogenous and exogenous viruses have been referred to as the 'virome'59. Already it is known that endogenous retroviruses could help to shape the T cell repertoire, deleting some specificities and expanding others116. Resident exogenous viruses might also influence the T cell repertoire and the activity status of the innate immune system, causing, for example, the production of cytokines and altered antigen presentation efficiency of DCs, but this needs to be formally shown. These effects will influence the balance of the response by individuals to exogenous agents. With technological advances in detecting both host gene polymorphisms and the virome, we anticipate that the influence of these factors on the outcome of virus infection will soon be better understood. This knowledge might provide clues to customize successful preventative and therapeutic approaches for viral infection and associated immunopathology.
Conclusions
Whether a virus infection results in severe, sometimes prolonged, lesions or is resolved with minimal bystander tissue damage depends on numerous factors. Some viruses (for example, HIV and HCV) have intrinsic properties that make immune control difficult, and attempts by the host immune system to achieve control results in notable tissue damage. Other infectious agents (for example, many herpesviruses) are successfully controlled in most individuals, but tissue damage occurs in those individuals that have predisposing genetic or acquired problems affecting one or more components of innate or adaptive immune system. Finally, some infections that are normally well controlled can cause extensive tissue damage under unusual circumstances. These might relate to the dose or route of exposure, the age of infection, host genetics and priming with cross-reacting viruses or co-infection with other agents.
References
- Munz, C., Lunemann, J. D., Getts, M. T. & Miller, S. D. Antiviral immune responses: triggers of or triggered by autoimmunity? Nature Rev. Immunol. 9, 246–258 (2009).
Google Scholar - de Martel, C. & Franceschi, S. Infections and cancer: established associations and new hypotheses. Crit. Rev. Oncol. Hematol. 70, 183–194 (2009).
PubMed Google Scholar - Tal, M. C. et al. Absence of autophagy results in reactive oxygen species-dependent amplification of RLR signaling. Proc. Natl Acad. Sci. USA 106, 2770–2775 (2009).
CAS PubMed PubMed Central Google Scholar - Pichlmair, A. & Reis e Sousa, C. Innate recognition of viruses. Immunity 27, 370–383 (2007).
CAS PubMed Google Scholar - Iwasaki, A. & Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nature Immunol. 5, 987–995 (2004).
CAS Google Scholar - Brooks, D. G. et al. Interleukin-10 determines viral clearance or persistence in vivo. Nature Med. 12, 1301–1309 (2006). This paper implicated the role of IL-10 in the persistence of LCMV infection of mice and showed the therapeutic value of its neutralization in achieving viral control.
CAS PubMed Google Scholar - Brady, M. T., MacDonald, A. J., Rowan, A. G. & Mills, K. H. Hepatitis C virus non-structural protein 4 suppresses Th1 responses by stimulating IL-10 production from monocytes. Eur. J. Immunol. 33, 3448–3457 (2003).
CAS PubMed Google Scholar - Hyodo, N., Nakamura, I. & Imawari, M. Hepatitis B core antigen stimulates interleukin-10 secretion by both T cells and monocytes from peripheral blood of patients with chronic hepatitis B virus infection. Clin. Exp. Immunol. 135, 462–466 (2004).
CAS PubMed PubMed Central Google Scholar - Brockman, M. A. et al. IL-10 is up-regulated in multiple cell types during viremic HIV infection and reversibly inhibits virus-specific T cells. Blood 114, 346–356 (2009).
CAS PubMed PubMed Central Google Scholar - Smit, J. J., Rudd, B. D. & Lukacs, N. W. Plasmacytoid dendritic cells inhibit pulmonary immunopathology and promote clearance of respiratory syncytial virus. J. Exp. Med. 203, 1153–1159 (2006).
CAS Google Scholar - Guidotti, L. G. et al. Viral clearance without destruction of infected cells during acute HBV infection. Science 284, 825–829 (1999).
CAS PubMed Google Scholar - Rehermann, B. Hepatitis C virus versus innate and adaptive immune responses: a tale of coevolution and coexistence. J. Clin. Invest. 119, 1745–1754 (2009).
CAS PubMed PubMed Central Google Scholar - Favre, D. et al. Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathog. 5, e1000295 (2009). This paper showed that T H 17 cells are induced after SIV infection and that the balance of T H 17 and T Reg cells is a crucial determinant in the progression of disease in pigtailed macaques but not in African green monkeys, in which T H 17 cells were progressively depleted by the virus.
PubMed PubMed Central Google Scholar - Rowan, A. G. et al. Hepatitis C virus-specific Th17 cells are suppressed by virus-induced TGF-β. J. Immunol. 181, 4485–4494 (2008). This is the first study to show that viral antigen-specific T H 17 cells are induced in HCV-infected individuals and that viral protein (NS4)-induced TGFβ can inhibit the activity of T H 17 cells.
CAS PubMed Google Scholar - Bermejo-Martin, J. F. et al. Th1 and Th17 hypercytokinemia as early host response signature in severe pandemic influenza. Crit. Care 13, R201 (2009).
PubMed PubMed Central Google Scholar - Culley, F. J., Pennycook, A. M., Tregoning, J. S., Hussell, T. & Openshaw, P. J. Differential chemokine expression following respiratory virus infection reflects Th1- or Th2-biased immunopathology. J. Virol. 80, 4521–4527 (2006).
CAS PubMed PubMed Central Google Scholar - Ravetch, J. In vivo veritas: the surprising roles of Fc receptors in immunity. Nature Immunol. 11, 183–185 (2010).
CAS Google Scholar - Buchmeier, M. J. & Oldstone, M. B. Virus-induced immune complex disease: identification of specific viral antigens and antibodies deposited in complexes during chronic lymphocytic choriomeningitis virus infection. J. Immunol. 120, 1297–1304 (1978). This is the first report in which immune complex deposits were measured using sensitive immunofluorescence and radioimmunoprecipitation in the tissue sites after a viral infection.
CAS PubMed Google Scholar - Nowoslawski, A., Krawczynski, K., Nazarewicz, T. & Slusarczyk, J. Immunopathological aspects of hepatitis type B. Am. J. Med. Sci. 270, 229–239 (1975).
CAS PubMed Google Scholar - Johnson, R. J. et al. Membranoproliferative glomerulonephritis associated with hepatitis C virus infection. N. Engl. J. Med. 328, 465–470 (1993).
CAS PubMed Google Scholar - Kimmel, P. L. et al. Brief report: idiotypic IgA nephropathy in patients with human immunodeficiency virus infection. N. Engl. J. Med. 327, 702–706 (1992).
CAS PubMed Google Scholar - Dakhama, A. et al. Virus-specific IgE enhances airway responsiveness on reinfection with respiratory syncytial virus in newborn mice. J. Allergy Clin. Immunol. 123, 138–145 (2009).
CAS PubMed Google Scholar - Couper, K. N., Blount, D. G. & Riley, E. M. IL-10: the master regulator of immunity to infection. J. Immunol. 180, 5771–5777 (2008).
CAS PubMed Google Scholar - Vieira, P. et al. Isolation and expression of human cytokine synthesis inhibitory factor cDNA clones: homology to Epstein-Barr virus open reading frame BCRFI. Proc. Natl Acad. Sci. USA 88, 1172–1176 (1991).
CAS PubMed PubMed Central Google Scholar - Kotenko, S. V., Saccani, S., Izotova, L. S., Mirochnitchenko, O. V. & Pestka, S. Human cytomegalovirus harbors its own unique IL-10 homolog (cmvIL-10). Proc. Natl Acad. Sci. USA 97, 1695–1700 (2000).
CAS PubMed PubMed Central Google Scholar - Moore, K. W., de Waal Malefyt, R., Coffman, R. L. & O'Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 19, 683–765 (2001).
CAS PubMed Google Scholar - Sarangi, P. P., Sehrawat, S., Suvas, S. & Rouse, B. T. IL-10 and natural regulatory T cells: two independent anti-inflammatory mechanisms in herpes simplex virus-induced ocular immunopathology. J. Immunol. 180, 6297–6306 (2008).
CAS PubMed Google Scholar - Mangia, A. et al. IL-10 haplotypes as possible predictors of spontaneous clearance of HCV infection. Cytokine 25, 103–109 (2004).
CAS PubMed Google Scholar - Naicker, D. D. et al. Interleukin-10 promoter polymorphisms influence HIV-1 susceptibility and primary HIV-1 pathogenesis. J. Infect. Dis. 200, 448–452 (2009).
CAS PubMed Google Scholar - Sun, J., Madan, R., Karp, C. L. & Braciale, T. J. Effector T cells control lung inflammation during acute influenza virus infection by producing IL-10. Nature Med. 15, 277–284 (2009). This study showed that by making IL-10, effector CD8+ T cells limit the extent of pulmonary tissue damage.
CAS PubMed Google Scholar - Elrefaei, M. et al. HIV-specific IL-10-positive CD8+ T cells suppress cytolysis and IL-2 production by CD8+ T cells. J. Immunol. 178, 3265–3271 (2007).
CAS PubMed Google Scholar - Kobasa, D. et al. Aberrant innate immune response in lethal infection of macaques with the 1918 influenza virus. Nature 445, 319–323 (2007).
CAS PubMed Google Scholar - Li, M. O. & Flavell, R. A. Contextual regulation of inflammation: a duet by transforming growth factor-β and interleukin-10. Immunity 28, 468–476 (2008).
PubMed Google Scholar - Li, M. O. & Flavell, R. A. TGF-β: a master of all T cell trades. Cell 134, 392–404 (2008).
CAS PubMed PubMed Central Google Scholar - Aung, H., Wu, M., Johnson, J. L., Hirsch, C. S. & Toossi, Z. Bioactivation of latent transforming growth factor β1 by Mycobacterium tuberculosis in human mononuclear phagocytes. Scand. J. Immunol. 61, 558–565 (2005).
CAS PubMed Google Scholar - Omer, F. M., de Souza, J. B., Corran, P. H., Sultan, A. A. & Riley, E. M. Activation of transforming growth factor β by malaria parasite-derived metalloproteinases and a thrombospondin-like molecule. J. Exp. Med. 198, 1817–1827 (2003).
CAS PubMed PubMed Central Google Scholar - Schultz-Cherry, S. & Hinshaw, V. S. Influenza virus neuraminidase activates latent transforming growth factor β. J. Virol. 70, 8624–8629 (1996).
CAS PubMed PubMed Central Google Scholar - Beckham, J. D., Tuttle, K. & Tyler, K. L. Reovirus activates transforming growth factor β and bone morphogenetic protein signaling pathways in the central nervous system that contribute to neuronal survival following infection. J. Virol. 83, 5035–5045 (2009).
CAS PubMed PubMed Central Google Scholar - Alatrakchi, N. et al. Hepatitis C virus (HCV)-specific CD8+ cells produce transforming growth factor β that can suppress HCV-specific T-cell responses. J. Virol. 81, 5882–5892 (2007).
CAS PubMed PubMed Central Google Scholar - Tinoco, R., Alcalde, V., Yang, Y., Sauer, K. & Zuniga, E. I. Cell-intrinsic transforming growth factor-β signaling mediates virus-specific CD8+ T cell deletion and viral persistence in vivo. Immunity 31, 145–157 (2009). This report showed that TGFβ functions intrinsically to limit CD8+T cell responses to viral infection.
CAS PubMed PubMed Central Google Scholar - O'Connor, W. Jr et al. A protective function for interleukin 17A in T cell-mediated intestinal inflammation. Nature Immunol. 10, 603–609 (2009).
CAS Google Scholar - Hou, W., Kang, H. S. & Kim, B. S. Th17 cells enhance viral persistence and inhibit T cell cytotoxicity in a model of chronic virus infection. J. Exp. Med. 206, 313–328 (2009).
CAS PubMed PubMed Central Google Scholar - Rabinovich, G. A. & Toscano, M. A. Turning 'sweet' on immunity: galectin-glycan interactions in immune tolerance and inflammation. Nature Rev. Immunol. 9, 338–352 (2009).
CAS Google Scholar - Zhu, C. et al. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nature Immunol. 6, 1245–1252 (2005).
CAS Google Scholar - Sehrawat, S., Suryawanshi, A., Hirashima, M. & Rouse, B. T. Role of Tim-3/galectin-9 inhibitory interaction in viral-induced immunopathology: shifting the balance toward regulators. J. Immunol. 182, 3191–3201 (2009). This paper showed that galectin 9 could promote FOXP3+ T Reg cell responses and that ligation of TIM3 with galectin 9 induces apoptosis of effector T cells but not T Reg cells.
CAS PubMed Google Scholar - Sehrawat, S., Reddy, P. B. J., Rajasagi, N., Suryawanshi, A., Hirashima, M. & Rouse, B. T. Galectin-9/TIM-3 interaction regulates virus-specific primary and memory CD8+ T cell response. PLoS Pathog. 6, e1000882 (2010).
PubMed PubMed Central Google Scholar - Chagan-Yasutan, H. et al. Persistent elevation of plasma osteopontin levels in HIV patients despite highly active antiretroviral therapy. Tohoku J. Exp. Med. 218, 285–92 (2009).
CAS PubMed Google Scholar - Mengshol, J. A. et al. A crucial role for kupffer cell-derived galectin-9 in regulation of T cell immunity in hepatitis C infection. PLoS ONE 5, e9504 (2010).
PubMed PubMed Central Google Scholar - Sakaguchi, S. Regulatory T cells: key controllers of immunologic self-tolerance. Cell 101, 455–458 (2000).
CAS PubMed Google Scholar - Belkaid, Y. & Tarbell, K. Regulatory T cells in the control of host-microorganism interactions. Annu. Rev. Immunol. 27, 551–589 (2009).
CAS PubMed Google Scholar - Suvas, S., Azkur, A. K., Kim, B. S., Kumaraguru, U. & Rouse, B. T. CD4+CD25+ regulatory T cells control the severity of viral immunoinflammatory lesions. J. Immunol. 172, 4123–4132 (2004).
CAS PubMed Google Scholar - Ruckwardt, T. J., Bonaparte, K. L., Nason, M. C. & Graham, B. S. Regulatory T cells promote early influx of CD8+ T cells in the lungs of respiratory syncytial virus-infected mice and diminish immunodominance disparities. J. Virol. 83, 3019–3028 (2009).
CAS PubMed PubMed Central Google Scholar - Lanteri, M. C. et al. Tregs control the development of symptomatic West Nile virus infection in humans and mice. J. Clin. Invest. 119, 3266–3277 (2009).
CAS PubMed PubMed Central Google Scholar - Sehrawat, S., Suvas, S., Sarangi, P. P., Suryawanshi, A. & Rouse, B. T. _In vitro_-generated antigen-specific CD4+ CD25+ Foxp3+ regulatory T cells control the severity of herpes simplex virus-induced ocular immunoinflammatory lesions. J. Virol. 82, 6838–6851 (2008).
CAS PubMed PubMed Central Google Scholar - Sehrawat, S. & Rouse, B. T. Anti-inflammatory effects of FTY720 against viral-induced immunopathology: role of drug-induced conversion of T cells to become Foxp3+ regulators. J. Immunol. 180, 7636–7647 (2008).
CAS PubMed Google Scholar - Rouse, B. T., Sarangi, P. P. & Suvas, S. Regulatory T cells in virus infections. Immunol. Rev. 212, 272–286 (2006).
CAS PubMed Google Scholar - MacDonald, A. J. et al. CD4 T helper type 1 and regulatory T cells induced against the same epitopes on the core protein in hepatitis C virus-infected persons. J. Infect. Dis. 185, 720–727 (2002). This report showed that induction of T regulatory 1 cells in patients infected with HCV negatively correlated with the extent of liver damage.
CAS PubMed Google Scholar - Liew, F. Y., Xu, D., Brint, E. K. & O'Neill, L. A. Negative regulation of Toll-like receptor-mediated immune responses. Nature Rev. Immunol. 5, 446–458 (2005).
CAS Google Scholar - Virgin, H. W., Wherry, E. J. & Ahmed, R. Redefining chronic viral infection. Cell 138, 30–50 (2009).
CAS PubMed Google Scholar - Barber, D. L. et al. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 439, 682–687 (2006). This seminal paper showed that the functional T cell exhaustion caused by a chronic viral infection could be reversed using antibody specific for the inhibitory molecule PD1.
CAS PubMed Google Scholar - Day, C. L. et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 443, 350–354 (2006). This report showed that antigen-specific CD8+ T cells isolated from patients with HIV expressed higher levels of the inhibitory molecule PD1 and that the exhaustion could be reversed, at least ex vivo , by using blocking antibody.
CAS PubMed Google Scholar - Radziewicz, H. et al. Liver-infiltrating lymphocytes in chronic human hepatitis C virus infection display an exhausted phenotype with high levels of PD-1 and low levels of CD127 expression. J. Virol. 81, 2545–2553 (2007).
CAS PubMed Google Scholar - Maier, H., Isogawa, M., Freeman, G. J. & Chisari, F. V. PD-1:PD-L1 interactions contribute to the functional suppression of virus-specific CD8+ T lymphocytes in the liver. J. Immunol. 178, 2714–2720 (2007).
CAS PubMed Google Scholar - Said, E. A. et al. Programmed death-1-induced interleukin-10 production by monocytes impairs CD4+ T cell activation during HIV infection. Nature Med. 16, 452–459 (2010).
CAS PubMed Google Scholar - Yao, Z. Q., King, E., Prayther, D., Yin, D. & Moorman, J. T cell dysfunction by hepatitis C virus core protein involves PD-1/PDL-1 signaling. Viral Immunol. 20, 276–287 (2007).
CAS PubMed Google Scholar - Boasso, A. et al. PDL-1 upregulation on monocytes and T cells by HIV via type I interferon: restricted expression of type I interferon receptor by CCR5-expressing leukocytes. Clin. Immunol. 129, 132–144 (2008).
CAS PubMed PubMed Central Google Scholar - Brooks, D. G. et al. IL-10 and PD-L1 operate through distinct pathways to suppress T-cell activity during persistent viral infection. Proc. Natl Acad. Sci. USA 105, 20428–20433 (2008).
CAS PubMed PubMed Central Google Scholar - Blackburn, S. D. et al. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nature Immunol. 10, 29–37 (2009).
CAS Google Scholar - Brooks, D. G., Lee, A. M., Elsaesser, H., McGavern, D. B. & Oldstone, M. B. IL-10 blockade facilitates DNA vaccine-induced T cell responses and enhances clearance of persistent virus infection. J. Exp. Med. 205, 533–541 (2008).
CAS PubMed PubMed Central Google Scholar - Ha, S. J. et al. Enhancing therapeutic vaccination by blocking PD-1-mediated inhibitory signals during chronic infection. J. Exp. Med. 205, 543–555 (2008).
CAS PubMed PubMed Central Google Scholar - Racaniello, V. R. One hundred years of poliovirus pathogenesis. Virology 344, 9–16 (2006).
CAS PubMed Google Scholar - Whitley, R. J. Herpes Simplex Viruses (eds Knipe, D. M & Howley, P. M.) (Lippincott Williams & Wilkins, New York, 2001).
Google Scholar - Tabeta, K. et al. The Unc93b1 mutation 3d disrupts exogenous antigen presentation and signaling via Toll-like receptors 3, 7 and 9. Nature Immunol. 7, 156–164 (2006).
CAS Google Scholar - Zhang, S. Y. et al. TLR3 deficiency in patients with herpes simplex encephalitis. Science 317, 1522–1527 (2007).
CAS PubMed Google Scholar - McIntosh, E. D. Paediatric infections: prevention of transmission and disease — implications for adults. Vaccine 23, 2087–2089 (2005).
PubMed Google Scholar - Rothberg, M. B., Haessler, S. D. & Brown, R. B. Complications of viral influenza. Am. J. Med. 121, 258–264 (2008).
PubMed PubMed Central Google Scholar - Tregoning, J. S. & Schwarze, J. Respiratory viral infections in infants: causes, clinical symptoms, virology, and immunology. Clin. Microbiol Rev. 23, 74–98 (2010).
CAS PubMed PubMed Central Google Scholar - Collins, P. L. & Graham, B. S. Viral and host factors in human respiratory syncytial virus pathogenesis. J. Virol. 82, 2040–2055 (2008).
CAS PubMed Google Scholar - Spann, K. M., Tran, K. C., Chi, B., Rabin, R. L. & Collins, P. L. Suppression of the induction of α, β, and λ interferons by the NS1 and NS2 proteins of human respiratory syncytial virus in human epithelial cells and macrophages. J. Virol. 78, 4363–4369 (2004).
CAS PubMed PubMed Central Google Scholar - Smit, J. J. et al. The balance between plasmacytoid DC versus conventional DC determines pulmonary immunity to virus infections. PLoS ONE 3, e1720 (2008).
PubMed PubMed Central Google Scholar - Culley, F. J., Pollott, J. & Openshaw, P. J. Age at first viral infection determines the pattern of T cell-mediated disease during reinfection in adulthood. J. Exp. Med. 196, 1381–1386 (2002). The influence of age at first exposure to a viral infection on the susceptibility of the same infection later in the life was shown in a mouse model of RSV infection.
CAS PubMed PubMed Central Google Scholar - Whitley, R. J. A 70-year-old woman with shingles: review of herpes zoster. JAMA 302, 73–80 (2009).
CAS PubMed Google Scholar - Rouse, B. T. & Kaistha, S. D. A tale of 2 alpha-herpesviruses: lessons for vaccinologists. Clin. Infect. Dis. 42, 810–817 (2006).
PubMed Google Scholar - Nikolich-Zugich, J. Ageing and life-long maintenance of T-cell subsets in the face of latent persistent infections. Nature Rev. Immunol. 8, 512–522 (2008).
CAS Google Scholar - Maue, A. C. et al. T-cell immunosenescence: lessons learned from mouse models of aging. Trends Immunol. 30, 301–305 (2009).
CAS PubMed PubMed Central Google Scholar - Snyder, C. M. et al. Memory inflation during chronic viral infection is maintained by continuous production of short-lived, functional T cells. Immunity 29, 650–659 (2008).
CAS PubMed PubMed Central Google Scholar - Schakel, K. Dendritic cells — why can they help and hurt us. Exp. Dermatol. 18, 264–273 (2009).
PubMed Google Scholar - Haaland, R. E. et al. Inflammatory genital infections mitigate a severe genetic bottleneck in heterosexual transmission of subtype A and C HIV-1. PLoS Pathog. 5, e1000274 (2009).
PubMed PubMed Central Google Scholar - McDermott, A. B. et al. Repeated low-dose mucosal simian immunodeficiency virus SIVmac239 challenge results in the same viral and immunological kinetics as high-dose challenge: a model for the evaluation of vaccine efficacy in nonhuman primates. J. Virol. 78, 3140–3144 (2004).
CAS PubMed PubMed Central Google Scholar - Oh, S., McCaffery, J. M. & Eichelberger, M. C. Dose-dependent changes in influenza virus-infected dendritic cells result in increased allogeneic T-cell proliferation at low, but not high, doses of virus. J. Virol. 74, 5460–5469 (2000).
CAS PubMed PubMed Central Google Scholar - Legge, K. L. & Braciale, T. J. Lymph node dendritic cells control CD8+ T cell responses through regulated FasL expression. Immunity 23, 649–659 (2005).
CAS PubMed Google Scholar - Asabe, S. et al. The size of the viral inoculum contributes to the outcome of hepatitis B virus infection. J. Virol. 83, 9652–9662 (2009). This study investigated the influence of dose of infecting HBV on the pathogenesis of liver disease and clearly showed that very high or very low doses of infection led to severe liver damage.
CAS PubMed PubMed Central Google Scholar - King, N. J. et al. Immunopathology of flavivirus infections. Immunol. Cell Biol. 85, 33–42 (2007).
CAS PubMed Google Scholar - Wang, T. et al. Toll-like receptor 3 mediates West Nile virus entry into the brain causing lethal encephalitis. Nature Med. 10, 1366–1373 (2004).
CAS PubMed Google Scholar - Koelle, D. M. & Corey, L. Herpes simplex: insights on pathogenesis and possible vaccines. Annu. Rev. Med. 59, 381–395 (2008).
CAS PubMed Google Scholar - Weiner, L. P. Pathogenesis of demyelination induced by a mouse hepatitis. Arch. Neurol. 28, 298–303 (1973).
CAS PubMed Google Scholar - Fazakerley, J. K. & Walker, R. Virus demyelination. J. Neurovirol. 9, 148–164 (2003).
CAS PubMed PubMed Central Google Scholar - Cole, G. A., Nathanson, N. & Prendergast, R. A. Requirement for θ-bearing cells in lymphocytic choriomeningitis virus-induced central nervous system disease. Nature 238, 335–337 (1972).
CAS PubMed Google Scholar - Halstead, S. B. Dengue. Lancet 370, 1644–1652 (2007).
PubMed Google Scholar - Mathew, A. & Rothman, A. L. Understanding the contribution of cellular immunity to dengue disease pathogenesis. Immunol. Rev. 225, 300–313 (2008).
CAS PubMed Google Scholar - Hadinoto, V. et al. On the dynamics of acute EBV infection and the pathogenesis of infectious mononucleosis. Blood 111, 1420–1427 (2008).
CAS PubMed PubMed Central Google Scholar - Clute, S. C. et al. Cross-reactive influenza virus-specific CD8+ T cells contribute to lymphoproliferation in Epstein-Barr virus-associated infectious mononucleosis. J. Clin. Invest. 115, 3602–3612 (2005). This study showed that major contributors to infectious mononucleosis are cross-reactive T cells specific for a previously encountered virus.
CAS PubMed PubMed Central Google Scholar - Kim, S. K. et al. Private specificities of CD8 T cell responses control patterns of heterologous immunity. J. Exp. Med. 201, 523–533 (2005).
CAS PubMed PubMed Central Google Scholar - Welsh, R. M. & Fujinami, R. S. Pathogenic epitopes, heterologous immunity and vaccine design. Nature Rev. Microbiol. 5, 555–563 (2007).
CAS Google Scholar - Segal, S. & Hill, A. V. Genetic susceptibility to infectious disease. Trends Microbiol. 11, 445–448 (2003).
CAS PubMed Google Scholar - Goulder, P. J. & Watkins, D. I. Impact of MHC class I diversity on immune control of immunodeficiency virus replication. Nature Rev. Immunol. 8, 619–630 (2008).
CAS Google Scholar - Brass, A. L. et al. Identification of host proteins required for HIV infection through a functional genomic screen. Science 319, 921–926 (2008).
CAS PubMed Google Scholar - Good, R. A. & Hansen, M. A. Primary immunodeficiency diseases. Adv. Exp. Med. Biol. 73, 155–178 (1976).
PubMed Google Scholar - Liu, R. et al. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell 86, 367–377 (1996).
CAS PubMed Google Scholar - Hill, A. V. Aspects of genetic susceptibility to human infectious diseases. Annu. Rev. Genet. 40, 469–486 (2006).
CAS PubMed Google Scholar - Casrouge, A. et al. Herpes simplex virus encephalitis in human UNC-93B deficiency. Science 314, 308–312 (2006).
CAS PubMed Google Scholar - Zhang, S. Y. et al. Inborn errors of interferon (IFN)-mediated immunity in humans: insights into the respective roles of IFN-α/β, IFN-γ, and IFN-λ in host defense. Immunol. Rev. 226, 29–40 (2008).
CAS PubMed Google Scholar - Kaur, G. & Mehra, N. Genetic determinants of HIV-1 infection and progression to AIDS: susceptibility to HIV infection. Tissue Antigens 73, 289–301 (2009).
CAS PubMed PubMed Central Google Scholar - Hubert, J. B. et al. Natural history of serum HIV-1 RNA levels in 330 patients with a known date of infection. The SEROCO Study Group. AIDS 14, 123–131 (2000). This study showed that some patients with HIV can control the virus for a long time without the need for antiretroviral therapy.
CAS PubMed Google Scholar - Seifarth, W. et al. Comprehensive analysis of human endogenous retrovirus transcriptional activity in human tissues with a retrovirus-specific microarray. J. Virol. 79, 341–352 (2005).
CAS PubMed PubMed Central Google Scholar - Lower, R., Lower, J. & Kurth, R. The viruses in all of us: characteristics and biological significance of human endogenous retrovirus sequences. Proc. Natl Acad. Sci. USA 93, 5177–5184 (1996).
CAS PubMed PubMed Central Google Scholar - Wilkins, C. & Gale, M. Jr. Recognition of viruses by cytoplasmic sensors. Curr. Opin. Immunol. 22, 41–47 (2001).
Google Scholar - York, I. A. et al. A cytosolic herpes simplex virus protein inhibits antigen presentation to CD8+ T lymphocytes. Cell 77, 525–535 (1994).
CAS PubMed Google Scholar - Ahn, K. et al. Human cytomegalovirus inhibits antigen presentation by a sequential multistep process. Proc. Natl Acad. Sci. USA 93, 10990–10995 (1996).
CAS PubMed PubMed Central Google Scholar - Gilbert, M. J., Riddell, S. R., Plachter, B. & Greenberg, P. D. Cytomegalovirus selectively blocks antigen processing and presentation of its immediate-early gene product. Nature 383, 720–722 (1996).
CAS PubMed Google Scholar - Levitskaya, J., Sharipo, A., Leonchiks, A., Ciechanover, A. & Masucci, M. G. Inhibition of ubiquitin/proteasome-dependent protein degradation by the Gly-Ala repeat domain of the Epstein-Barr virus nuclear antigen 1. Proc. Natl Acad. Sci. USA 94, 12616–12621 (1997).
CAS PubMed PubMed Central Google Scholar - Koppelman, B., Neefjes, J. J., de Vries, J. E. & de Waal Malefyt, R. Interleukin-10 down-regulates MHC class II αβ peptide complexes at the plasma membrane of monocytes by affecting arrival and recycling. Immunity 7, 861–871 (1997).
CAS PubMed Google Scholar - Greenberg, M. E. et al. Co-localization of HIV-1 Nef with the AP-2 adaptor protein complex correlates with Nef-induced CD4 down-regulation. EMBO J. 16, 6964–6976 (1997).
CAS PubMed PubMed Central Google Scholar - Holmes, E. C. Evolutionary history and phylogeography of human viruses. Annu. Rev. Microbiol. 62, 307–328 (2008).
CAS PubMed Google Scholar - Devergne, O., Birkenbach, M. & Kieff, E. Epstein–Barr virus-induced gene 3 and the p35 subunit of interleukin 12 form a novel heterodimeric hematopoietin. Proc. Natl Acad. Sci. USA 94, 12041–12046 (1997).
CAS PubMed PubMed Central Google Scholar - Moskophidis, D., Lechner, F., Pircher, H. & Zinkernagel, R. M. Virus persistence in acutely infected immunocompetent mice by exhaustion of antiviral cytotoxic effector T cells. Nature 362, 758–761 (1993).
CAS PubMed Google Scholar - Periwal, S. B. & Cebra, J. J. Respiratory mucosal immunization with reovirus serotype 1/L stimulates virus-specific humoral and cellular immune responses, including double-positive (CD4+/CD8+) T cells. J. Virol. 73, 7633–7640 (1999).
CAS PubMed Google Scholar - Fulton, J. R., Smith, J., Cunningham, C. & Cuff, C. F. Influence of the route of infection on development of T-cell receptor β-chain repertoires of reovirus-specific cytotoxic T lymphocytes. J. Virol. 78, 1582–1590 (2004).
CAS PubMed PubMed Central Google Scholar - Le Goffic, R. et al. Detrimental contribution of the Toll-like receptor (TLR)3 to influenza A virus-induced acute pneumonia. PLoS Pathog. 2, e53 (2006).
PubMed Google Scholar - Bochud, P. Y., Magaret, A. S., Koelle, D. M., Aderem, A. & Wald, A. Polymorphisms in TLR2 are associated with increased viral shedding and lesional rate in patients with genital herpes simplex virus type 2 infection. J. Infect. Dis. 196, 505–509 (2007).
CAS PubMed Google Scholar - Almarri, A. & Batchelor, J. R. HLA and hepatitis B infection. Lancet 344, 1194–1195 (1994).
CAS PubMed Google Scholar - Gao, X. et al. AIDS restriction HLA allotypes target distinct intervals of HIV-1 pathogenesis. Nature Med. 11, 1290–1292 (2005).
CAS PubMed Google Scholar - Fellay, J. et al. A whole-genome association study of major determinants for host control of HIV-1. Science 317, 944–947 (2007).
CAS PubMed PubMed Central Google Scholar - Fanning, L. J. et al. HLA class II genes determine the natural variance of hepatitis C viral load. Hepatology 33, 224–230 (2001).
CAS PubMed Google Scholar - Thomas, D. L. et al. Genetic variation in IL28B and spontaneous clearance of hepatitis C virus. Nature 461, 798–801 (2009).
CAS PubMed PubMed Central Google Scholar - Ge, D. et al. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature 461, 399–401 (2009).
CAS PubMed Google Scholar - Knapp, S. et al. Polymorphisms in interferon-induced genes and the outcome of hepatitis C virus infection: roles of MxA, OAS-1 and PKR. Genes Immun. 4, 411–419 (2003).
CAS PubMed Google Scholar - Monto, A. S. Epidemiology of influenza. Vaccine 26, D45–D48 (2008).
PubMed Google Scholar - Peebles, R. S. Jr & Graham, B. S. Pathogenesis of respiratory syncytial virus infection in the murine model. Proc. Am. Thorac Soc. 2, 110–115 (2005).
CAS PubMed PubMed Central Google Scholar - Lemke, G. & Rothlin, C. V. Immunobiology of the TAM receptors. Nature Rev. Immunol. 8, 327–336 (2008).
CAS Google Scholar
Acknowledgements
We thank D. Masopust and M. Sangster for valuable comments. The work was supported by US National Institutes of Health grants RO1 AI 106336501 and RO1 EY 05093.
Author information
Authors and Affiliations
- Department of Pathobiology, College of Veterinary Medicine, University of Tennessee, Knoxville, 37996-0845, Tennessee, USA
Barry T. Rouse - Whitehead Institute of Biomedical Research, 9 Cambridge center, Cambridge, 02142, Massachusetts, USA
Sharvan Sehrawat
Authors
- Barry T. Rouse
You can also search for this author inPubMed Google Scholar - Sharvan Sehrawat
You can also search for this author inPubMed Google Scholar
Corresponding author
Correspondence toBarry T. Rouse.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Related links
Related links
FURTHER INFORMATION
Glossary
Plasmacytoid DC
A dendritic cell (DC) subset with a morphology that resembles that of a plasmablast. Plasmacytoid DCs produce large amounts of type I interferons in response to viral infection.
Resolvins
Resolution-phase interaction products that are made by the host from eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) and that have anti-inflammatory properties.
Galectins
A family of lectin proteins that bind a wide variety of glycoproteins and glycolipids containing β-galactosides through their carbohydrate-recognition domain. They have extracellular and intracellular functions, including the regulation of apoptosis, RAS signalling, cell adhesion and angiogenesis.
Regulatory T (TReg) cells
A subset of T cells that control the activity of effector T cells under inflammatory or steady state conditions.
Protectins
A family of compounds that are derived from DHA and that are characterized by a conjugated triene-containing structure. They have been shown to regulate the influx of neutrophils at inflammatory sites.
Exhaustion
Impaired ability of effector T cells to carry out their functions such as cytotoxicity and cytokine secretion owing to chronic stimulation by antigen.
Latent infection
A dormant infection with a microorganism that persists in the body for a long period of time, but it can be reactivated under certain conditions, such as immunosuppression.
Shingles
A viral disease characterized by painful, blistering skin rashes due to infection or recurrence of infection with varicella zoster virus.
Innate receptors
The receptors that are present either on the surface or in the cytoplasm of innate immune cells that recognize microbial surface patterns or their replicative products and induce cytokine production.
Dengue haemorrhagic fever
A viral disease transmitted by the bites of Aedes egypti mosquitos that carry dengue virus and is characterized by headache, fever, rash and evidence of haemorrhage in the body.
Infectious mononucleosis
A viral disease caused by infection with Epstein–Barr virus most commonly during adolescence and young adulthood. The classical symptoms include fever, sore throat and swollen lymph glands, especially in the neck.
Virome
The total virus-derived genetic material present in the host owing to integrated or persistent exogenous viruses.
Rights and permissions
About this article
Cite this article
Rouse, B., Sehrawat, S. Immunity and immunopathology to viruses: what decides the outcome?.Nat Rev Immunol 10, 514–526 (2010). https://doi.org/10.1038/nri2802
- Issue Date: July 2010
- DOI: https://doi.org/10.1038/nri2802