Syntactic and Thematic Constraint Effects on Blood Oxygenation Level Dependent Signal Correlates of Comprehension of Relative Clauses (original) (raw)
. Author manuscript; available in PMC: 2008 Jun 13.
Published in final edited form as: J Cogn Neurosci. 2008 Apr;20(4):643–656. doi: 10.1162/jocn.2008.20044
Abstract
The effects of plausibility of thematic role assignment and syntactic structure on blood oxygenation level dependent (BOLD) signal were studied using event-related functional magnetic resonance imaging by orthogonally varying syntactic structure (subject- vs. object-extracted relative clauses) and the plausibility of nouns playing thematic roles (constrained vs. unconstrained sentences) in a plausibility judgment task. In plausible sentences, BOLD signal increased for object-compared to subject-extracted clauses in unconstrained sentences in left middle temporal and left inferior frontal areas, for this contrast in constrained sentences in left middle temporal but not left inferior frontal areas, and for constrained subject-extracted sentences compared to unconstrained subject-extracted sentences in the left inferior frontal gyrus and the left dorsolateral prefrontal cortex. We relate these areas of activation to the assignment of the syntactic structure of object-compared to subject-extracted structures and the process of checking which thematic roles activated in the course of processing a sentence are licensed by the syntactic structure of the sentence.
INTRODUCTION
Sentences with relative clauses have been the subject of considerable study in neuroimaging (Waters, Caplan, Alpert, & Stanczak, 2003; Caplan et al., 2002; Cooke et al., 2001; Indefrey, Hagoort, Herzog, Seitz, & Brown, 2001; Caplan, Alpert, Waters, & Olivieri, 2000; Caplan, Alpert, & Waters, 1998, 1999; Just, Carpenter, Keller, Eddy, & Thulborn, 1996; Stromswold, Caplan, Alpert, & Rauch, 1996). Increases in neurovascular responses associated with object- compared to subject-extracted relative clauses have been interpreted as providing evidence regarding the localization of the neural structures that support the memory and computational demands associated with assigning the object-extracted structure. However, recent studies have shown that both behavioral and neurovascular measures of processing are affected by the interaction of syntactic structure and semantic features of the nouns that play thematic roles in these sentences (Caplan, Chen, & Waters, in press; Caplan & Chen, 2006; Chen, West, Waters, & Caplan, 2006). The present study explores the blood oxygenation level dependent (BOLD) signal correlates of assigning the structure and meaning of sentences with relative clauses and thematic roles that are and are not constrained by real-world events.
To introduce the study, we begin with a brief review of psycholinguistic findings that are relevant to the processing of these structures. The most detailed studies are by Traxler, Morris, and Seely (2002). These authors studied sentences with object- and subject-extracted relative clauses that differed with respect to the constraints provided by real-world knowledge regarding likely thematic roles. They presented sentences in which both nouns in the relative clause were animate and thematic roles were unconstrained (1) and sentences in which both nouns in the relative clause were animate and thematic roles were constrained by real-world factors (2).
- Unconstrained
- Subject extracted (Subject–subject: SS):1 The lawyer that irritated the banker filed a hefty lawsuit.
- Object extracted (Subject–object: SO): The lawyer that the banker irritated filed a hefty lawsuit.
- Constrained
- Subject extracted (Subject–subject: SS): The policeman that arrested the thief was known to carry a knife.
- Object extracted (Subject–object: SO): The thief that the policeman arrested was known to carry a knife.
Traxler et al. (2002) found that, for both Sentences (1) and (2), eye fixations were longer on both the relative clause and the main verb in sentences with object-extracted relative clauses than in sentences with subject-extracted relative clauses. The existence of constraints on plausible thematic roles played by the noun phrases in the relative clause did not affect eye fixations on either the relative clause or the main verb in these sentences. However, participants regressed from the main verb more often and fixated the sentence-initial noun phrase and the relative clause longer in these regressions in object-extracted unconstrained sentences (1b) than object-extracted constrained sentences (2b). That is, readers appeared to make no use of the real-world constraints available in sentences such as (2) compared to sentences such as (1) during their initial reading of the sentences, but they found a need to return to the sentence-initial noun phrase and the relative clause for a longer period when the sentences were unconstrained and their syntactic structure was complex (1b). Traxler et al. concluded that readers were less secure in their interpretation of the unconstrained object-extracted sentences and that the regressions to the sentence-initial noun phrase supported a process that checked that the correct (the syntactically licensed) thematic roles had been assigned.
Turning to neuroimaging results, two groups of researchers have examined BOLD signal in the unconstrained sentences presented by Traxler et al. (2002). Just et al. (1996) used the following type of stimuli in a verification task:
- 3a. Object-extracted sentences (SO): The policeman who the guard admired congratulated the officer.
- 3b. Subject-extracted sentences (SS): The policeman who congratulated the officer admired the guard.
- 3c. Conjoined sentences (Conjoined [CON]): The policeman congratulated the officer and admired the guard.
They found progressively larger increases in BOLD signal for these sentence types—(3a) > (3b) > (3c)—bilaterally in the posterior inferior frontal gyri (IFG) and posterior superior temporal gyri, with greater BOLD signal on the left than on the right in both areas. Just et al. attributed the BOLD signal effects of these contrasts to the increased “working memory” load associated with the assignment of the relative clause in object-extracted structures and the use of this syntactic structure to determine thematic roles.2
The object/subject extraction contrast did not produce BOLD signal effects for unconstrained object compared to subject extracted structures in verification tasks reported by Fiebach, Schlesewsky, and Lohmann (2005), Fiebach, Vos, and Friederici (2004), and Fiebach, Schlesewsky, and Friederici (2001). However, Fiebach et al. did find that increasing the distance of the head noun of the relative clause from the verb of the clause (which is clause final in German) produced increases in BOLD signal in the bilateral inferior frontal and superior temporal gyri, and concluded that these areas supported one part of the working memory system that supports the assignment of object-extracted structures—the “storage” costs of assigning syntactic structure.
The differences between the Just et al. and Fiebach et al. studies could have been due to many factors: differences between English and German word order (subject–verb–object; subject–object–verb); the specific structures presented (relative clauses; indirect questions); presentation mode (whole sentence; rapid serial visual presentation); the aspect of the BOLD signal that was analyzed (entire trial; selected TRs during target presentation); the selection of voxels for analysis; the BOLD signal analysis (counts of voxels above a threshold in selected regions of interest [ROIs]; the statistical analysis of all voxels in selected ROIs), or others. Both studies do suggest, however, that the left, and to a lesser extent the right, IFG and superior temporal gyrus (STG) support the working memory system that is required in syntactic processing and sentence interpretation.
In light of the Traxler et al. (2002) results, however, other possibilities must be considered. Traxler et al. concluded that there was a late checking process that occurred only in SO unconstrained sentences, and this checking process might be one source of BOLD signal increase in a contrast of object- and subject-extracted unconstrained structures. To further study the determinants of BOLD signal effects in the object- and subject-extracted contrast, the present study examines the neurovascular response to sentences with constrained and unconstrained object- and subject-extracted relative clauses in a plausibility judgment task.
METHODS
Participants
Sixteen healthy participants from Boston University (12 women, 4 men) were paid for their participation in the study. All subjects gave written informed consent and were screened for MRI compatibility before entry into the scanning room. All subjects were right-handed, with no history of neurological or psychiatric disorders. All subjects spoke English as a native language, and were between 18 and 30 years of age.
Materials
Stimuli consisted of sentences modeled on (and some taken from) Traxler et al.’s (2002) “animate unconstrained” and “animate constrained” sentence types. Examples of plausible stimuli are as mentioned below.
Plausible Sentences
- 4a. Object-extracted unconstrained
The fireman who the deputy called saved the sailor. - 4b. Object-extracted constrained
The thief who the policeman arrested watched the driver. - 4c. Subject-extracted unconstrained
The fireman who called the deputy saved the sailor. - 4d. Subject-extracted constrained
The policeman who arrested the thief watched the driver.
The implausible sentences were derived from the plausible sentences in the following fashion: Approximately half were created by changing the verb of the relative clause to create an implausible proposition (e.g., The policeman that diluted the thief watched the driver), one third by changing the verb of the main clause to create an implausible proposition (e.g., The terrorist that captured the hostage surrounded the demands), and one sixth by changing the object noun of the main clause to violate the animacy constraint of the main verb on that noun (e.g., The lawyer that prosecuted the mobster thanked the bribe). The exact percentage of these types of changes used to create implausible sentences differed somewhat between the plausible constrained and unconstrained sentences.
The inclusion of violations of animacy constraints on thematic roles in the implausible sentences increases the range of plausibility of the stimuli and introduces variety in lexical content, both of which were judged to be valuable in a study that presented a large number of sentences of two syntactic types. It allows participants to discriminate plausible sentences from some implausible sentences on the basis of the presence of inanimate nouns in the implausible sentences, but the number of implausible sentences with inanimate nouns was small and this cannot have been the basis for making most plausibility judgments; moreover, it could not lead to differences in how judgments were made for plausible sentences with object- and subject-extracted relative clauses or for plausible sentences with constrained or unconstrained thematic roles.
We obtained plausibility ratings for plausible and implausible sentence types in individuals who did not participate in the functional magnetic resonance imaging (fMRI) study. Sentences were presented in subject-extracted form and plausibility was rated on a scale of 1 (implausible) to 7 ( plausible). Ten participants rated the entire stimulus sentences, and 10 rated the thematic roles expressed in relative clause alone and the thematic roles created by reversing these noun phrases around the verb. For the sentences as a whole, the main effect of plausibility was significant in a 2 (plausibility: plausible, implausible) × 2 (constraint: constrained, unconstrained) analysis of variance (ANOVA) [_F_1(1, 9) = 120.1, p < .001: mean plausibility rating for the plausible sentences = 5.8 and for the implausible sentences = 2.4]. For the relative clauses, a 2 (“direction:” presented; reversed) × 2 (constraint: constrained, unconstrained) ANOVA for the plausible sentences revealed an interaction of “direction” and constraint [_F_1(1, 9) = 230.1, p < .001]. Tukey’s test (p < .05) showed that, for unconstrained sentences, the presented and reversed sets of thematic roles did not differ (mean rating for presented unconstrained clauses = 6.1; for reversed unconstrained clauses = 5.7); that, for constrained sentences, the presented sets of thematic roles were significantly more plausible than the reversed thematic roles (mean rating for presented constrained clauses = 6.3; for reversed constrained clauses = 3.0); and that the presented clauses did not differ for constrained and unconstrained sentences. The plausibility of the reversed constrained clauses also did not differ from the plausibility of the implausible sentences. Thus, the discrimination of plausible and implausible sentences could not be accomplished on the basis of implausible sentences being less plausible than the transiently activated syntactically unlicensed propositions in any set of the plausible sentences, but required participants to determine whether the syntactically licensed proposition in each sentence was plausible or not, and there were no differences in the plausibility of the constrained and unconstrained sentences that could have affected results.
Other low-level variables that might affect behavioral or BOLD signal responses were also controlled. The words in the constrained and unconstrained sentences did not differ in length or frequency. The degree of semantic association between the nouns in the constrained and the unconstrained sentences was also controlled across the relative clauses in the plausible constrained and unconstrained sentences. University of South Florida Free Association norms, which are based on ~750,000 responses to 5019 stimulus words in 6000 participants, were consulted to obtain these measures for each pair of nouns in each relative clause. The mean forward and backward association strength did not differ for constrained and unconstrained sentences (mean forward constrained = 0.02, unconstrained = 0.02; mean backward constrained = 0.02, unconstrained = 0.01).
Because of these features of the stimuli, differences in participants’ behavioral or BOLD signal responses for plausible constrained and unconstrained object- and subject-extracted sentences in the plausibility judgment task in the fMRI environment are interpreted to be due to an interaction between syntactic structure and the degree of constraint on the thematic roles that NPs can play around verbs in the performance of that task.
Procedure
The task for the participants during the experimental trial was to read the sentence and judge the plausibility of the presented item as quickly and accurately as possible. A plausible sentence was described as a sentence that had a meaning the participant could imagine happening in the real world. RTs and accuracy were recorded.
MR Methods
Stimulus Presentation
Stimuli consisted of 144 sentences (36 plausible constrained, 36 plausible unconstrained, 36 implausible constrained, and 36 implausible constrained), each visually displayed in its entirely on a single line in the center of the screen via computer controlled projection system that delivered a visual stimulus to a projection screen. The participant viewed the screen through a mirror attached to the top of the head coil.
Sentences were presented with PsyScope software (Cohen, MacWhinney, Flatt, & Provost, 1993) on a Macintosh iBook computer. The total trial length for each sentence was 6 sec. Each trial started with a 300-msec fixation cross, followed by a 100-msec interstimulus interval. Then, the sentence was displayed for 5 sec, during which the subjects made a plausibility judgment, which was recorded for accuracy and response time via an fMRI-compatible button box. Responses beyond 5 sec after the initial presentation of the sentence were disregarded (0.7% of trials). A 600-msec fixation followed the sentence. Responses were made with the left hand.
The 144 trials were divided into 8 runs. Fixation trials of 0, 2, 4, 6, 8, 10, 12, or 14 sec were randomly interspersed between each 6-sec trial. Stimuli were presented in a pseudorandomized order that was predetermined by a computer program to maximize efficiency in the deconvolution and estimation of the hemodynamic response (Burock & Dale, 2000; Dale, 1999; Burock, Buckner, Woldorff, Rosen, & Dale, 1998; Dale & Buckner, 1997).
MRI Parameters
Participants were scanned at the Boston University Medical Center on a 3-Tesla Philips Intera scanner. The participants’ heads were firmly secured with padding. One T1-weighted MP-RAGE sequence structural scan was taken, lasting approximate 7.5 min, using a sense head coil (TR = 9.9, TE = 4.6, flip angle = 3.48). Volumes consisted of 137 slices with a slice thickness of 1 mm (matrix = 256, FOV = 256 mm). This was also used to view the positioning of the participant’s head and to graphically prescribe the functional image volumes along the AC–PC plane. Functional image volumes were collected with a T2*-weighted gradient-echo pulse sequence (TR = 2 sec, TE = 35 msec, flip angle = 90°, FOV = 210 mm, 64 × 64 matrix). Each run consisted of 140 volumes; each volume consisted of thirty 3-mm interleaved slices with a gap distance of 0.9 mm between slices. An initial 8 sec (4 TR equivalent) buffer of radio-frequency pulse was run before presenting the stimuli, to ensure maximal signal during the length of the functional run.
Image Processing
Functional images were realigned using the procedure by Friston, Holmes, Poline, Price, and Frith (1996) as implemented in statistical parametric mapping (SPM2; Welcome Department of Imaging Neuroscience). The first image volume for each participant was used to determine parameters for spatial normalization to match the EPI template provided within SPM2. The spatial transformation into standardized Montreal Neurological Institute (MNI) space was determined with a tailored algorithm with both linear and nonlinear components (Friston et al., 1995). The normalized data were smoothed with an 8-mm full-width half-maximum Gaussian filter.
Each condition was modeled with a canonical hemodynamic response in SPM2 (Aguirre, Zarahn, & D’Esposito, 1998). A high-pass filter was incorporated into the model to remove noise associated with low-frequency confounds (e.g., respiratory artifact, scanner drift).
A random effects model was created in which contrasts represented differences between the amplitude of different conditions using t tests at each voxel for each of the 16 subjects. These differences were then entered into a fixed effects model. A correction for multiple comparisons based on the theory of Gaussian fields was employed (Worsley & Friston, 1995; Worsley, 1994), such that reported significance levels are at the voxel level and were all greater than p < .001.
RESULTS
Behavioral Results
The accuracy data are presented in Figure 1. These data were analyzed in 2 (structure: object- vs. subject-extracted) × 2 (plausibility: plausible vs. implausible) × 2 (constraint: constrained vs. unconstrained)3 ANOVAs by subjects and items. There was an effect of constraint [_F_1(1, 15) = 21.0, p < .001; _F_2(1, 312) = 6.9, p < .01]. Errors were more frequent in constrained than in unconstrained plausible sentences but not in implausible sentences. No interactions were significant by both subjects and items.
Figure 1.
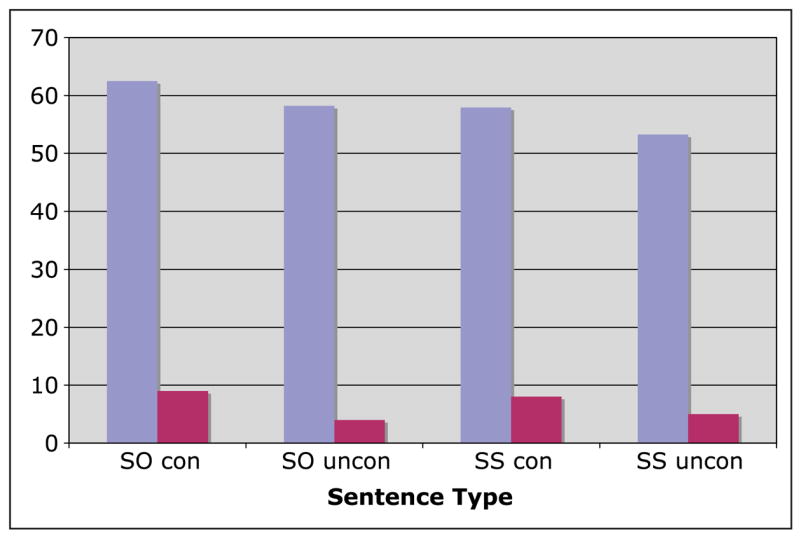
Mean accuracy (percent errors; red bars) and RTs (msec/character; blue bars) for different sentence conditions. SS = subject-extracted clause; SO = object-extracted clause; Con = constrained thematic roles; Uncon = unconstrained thematic roles.
Mean RTs for correct responses are shown in Figure 1. These data were analyzed in 2 (structure: object- vs. subject-extracted) × 2 (plausibility: plausible vs. implausible) × 2 (constraint: constrained vs. unconstrained) ANOVAs by subjects and items. There were effects of all three variables [for sentence type, _F_1(1, 15) = 66.4, p < .001; _F_2(1, 312) = 25.1, p < .001; for constraint, _F_1(1, 15) = 105.0, p < .001; _F_2(1, 312) = 20.8, p < .001; for plausibility, _F_1(1, 15) = 15.9, p < .001; _F_2(1, 312) = 18.3, p < .001]. RTs were longer in SO than in SS sentences, in plausible sentences than in implausible sentences, and in constrained than in unconstrained sentences. No interactions were significant by both subjects and items.
fMRI Results
There were BOLD signal differences for the object/subject extraction contrast for plausible versions of both unconstrained and unconstrained sentences (Table 1; Figure 2). In plausible unconstrained sentences, BOLD signal in object-extracted sentences increased in the left middle and superior temporal gyri (MTG; STG), and in the left IFG. In plausible constrained sentences, it increased in the left MTG and inferior temporal gyrus (ITG) and in the left cuneus and precuneus.
Table 1.
Areas of Activation in Contrasts of Conditions
| Contrast | p | Size | T | Z | MNI | Location | BA |
|---|---|---|---|---|---|---|---|
| Plausible unconstrained sentences, SO–SS | .000 | 1355 | 8.31 | 5.01 | −52 −44 −2 | L MTG | 21 |
| 6.76 | 4.51 | −46 −56 16 | L STG | 22 | |||
| 5.90 | 4.18 | −62 −18 −12 | L MTG | 21 | |||
| .000 | 331 | 6.07 | 4.25 | −38 30 0 | L IFG | 47 | |
| 4.85 | 3.71 | −44 34 −14 | L DLPFC | 11 | |||
| 4.81 | 3.68 | −46 24 −8 | L IFG | 47 | |||
| .002 | 220 | 5.30 | 3.92 | −54 18 28 | L DLPFC | 9 | |
| 4.12 | 3.32 | −46 14 18 | L DLPFC | 9 | |||
| Plausible constrained sentences, SO–SS | .000 | 275 | 7.75 | 4.84 | −24 −76 32 | Precuneus | 19 |
| 4.95 | 3.75 | −28 −84 22 | Cuneus | 18 | |||
| 4.90 | 3.73 | −24 −86 32 | Cuneus | 18 | |||
| .000 | 1323 | 7.48 | 4.76 | −58 −46 2 | L MTG | 22 | |
| 7.29 | 4.69 | −50 −60 12 | L MTG | 22 | |||
| 6.36 | 4.36 | −50 −42 −4 | L ITG | 37 | |||
| Plausible sentences, SS constrained–unconstrained | .000 | 1504 | 7.93 | 4.90 | −56 24 8 | L IFG | 45 |
| 6.08 | 4.25 | −40 36 −14 | L IFG | 46 | |||
| 6.05 | 4.24 | −38 22 −8 | L IFG | 47 | |||
| .000 | 405 | 7.15 | 4.65 | 40 28 −12 | R IFGa | 47 | |
| 5.86 | 4.16 | 56 26 −6 | R IFG | 47 | |||
| .002 | 200 | 5.91 | 4.18 | −54 −62 20 | L MTG | 39 | |
| 5.24 | 3.89 | −54 −72 24 | L MTG | 39 | |||
| 4.64 | 3.60 | −44 −60 18 | L MTG | 39 | |||
| .001 | 227 | 5.86 | 4.16 | −32 20 44 | L PMC | 8 | |
| 5.63 | 4.07 | −42 2 58 | L PMC | 6 | |||
| 4.31 | 3.43 | −30 4 46 | L PMC | 6 |
Figure 2.
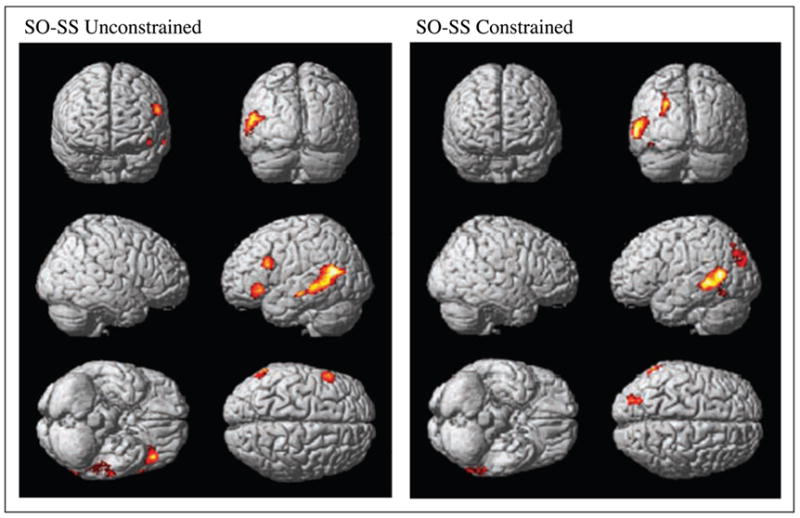
SPM analysis of BOLD signal in contrast of object- and subject-extracted constrained and unconstrained plausible sentences. SO = object-extracted sentences; SS = subject-extracted sentences.
There were increases in BOLD signal for constrained compared to unconstrained plausible SS sentences in the left and right IFG and more anteriorly in the left dorsolateral prefrontal cortex (DLPFC) and premotor cortex (PMC), and in the left MTG (Table 1; Figure 3). There were no areas activated by the contrast of constrained compared to unconstrained plausible SO sentences.
Figure 3.
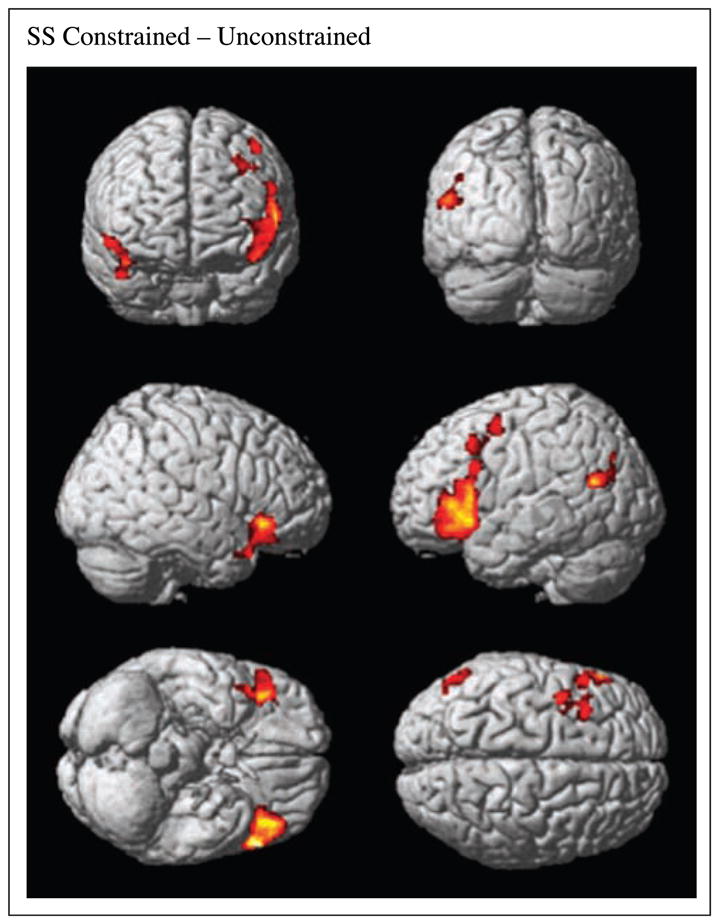
SPM analysis of BOLD signal in contrast of constrained and unconstrained plausible SS sentences.
Voxels that were activated in both the contrast of SO/SS constrained and SO/SS unconstrained plausible sentences in the left temporal and frontal areas were identified using a conjunction analysis and the difference in BOLD signal in those ROIs compared to a prestimulus baseline was measured for each sentence type. The results are shown in Figure 4. These results were analyzed in a 2 (ROI: temporal, frontal) × 2 (structure: SO, SS) × 2 (constraint: constrained, unconstrained) ANOVA, which showed an interaction of ROI, structure, and constraint [F(1, 30) = 4.8, p < .05].
Figure 4.
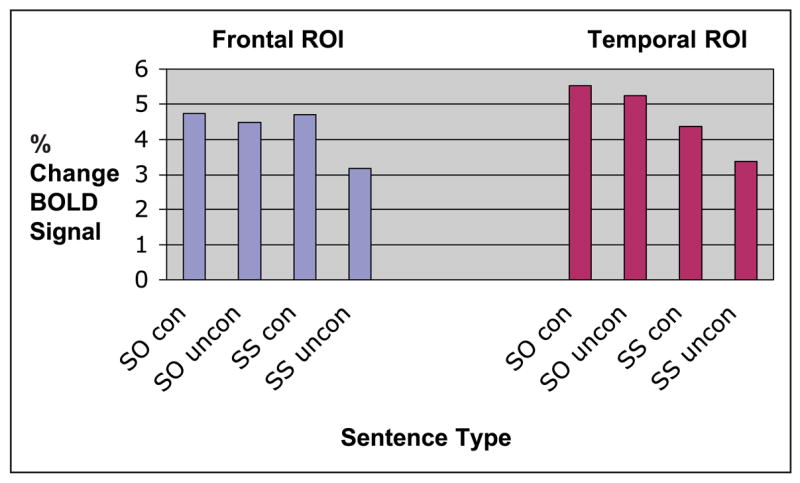
Percent increase in BOLD signal relative to prestimulus baselines in left frontal (blue) and temporal (red) ROIs.
This interaction was explored with Tukey’s tests (set at p < .05). BOLD signal was higher for SO sentences in the temporal ROI than in the frontal ROI in both constrained and unconstrained sentences, and did not differ across the ROIs for either constrained or unconstrained SS sentences. In the temporal ROI, BOLD signal increased for SO compared to SS sentences for both the constrained and unconstrained sentences. BOLD signal also increased for constrained compared to unconstrained SS sentences in the temporal ROI. In the frontal ROI, BOLD signal increased for SO compared to SS sentences in the unconstrained sentences only. This was because of greater BOLD signal in constrained SS sentences, which also did not differ from BOLD signal associated with unconstrained SO sentences. BOLD signal did not differ for constrained and unconstrained SO sentences in either ROI.
DISCUSSION
Behavioral Results
Each of the variables we manipulated—syntactic structure, plausibility, and constraint—resulted in main effects in the RT data. These effects can be attributed to the following cognitive processes.
The effect of syntactic structure is an indication of the processing load imposed by object relativization. There were effects of syntactic structure in both the plausible and implausible sentences, as expected. The lack of an interaction of constraint with sentence type indicates that constraint affected end-of-sentence performance equally in the object- and subject-extracted sentences, as was the case in Traxler et al.’s (2002) question answering data.
The effect of plausibility can be attributed to having to check both propositions to determine that a sentence is plausible but being able to check only one to determine that a sentence is implausible on some trials. If propositions are checked for plausibility incrementally, an implausible proposition will be identified before the end of the sentence in a certain number of cases, leading to overall faster RTs for implausible than for plausible sentences.
The effect of constraint—increased RTs and error rate in constrained compared to unconstrained plausible sentences—might initially seem paradoxical because constrained sentences are intuitively easier to understand, but it is easily explained if participants considered syntactically unlicensed thematic roles as well as those that are actually licensed by the sentence. Participants would have considered syntactically unlicensed thematic roles if they assigned all nouns to all possible thematic role positions around verbs (see Ferreira, 2003, for evidence of such a process). For instance, in a sentence such as The fireman who the deputy called saved the sailor, participants would consider the interpretation that the fireman called the deputy. In plausible constrained sentences (e.g., The thief who the policeman arrested watched the driver), the unlicensed thematic roles (The thief arrested the policeman) are implausible, making the judgment that the presented sentence is plausible more difficult and time-consuming than in plausible unconstrained sentences, where syntactically unlicensed thematic roles were plausible.4
We note that the effect of constraint in plausible sentences works in the opposite direction in a plausibility judgment task, used here, than in question answering or probe verification, used in Fiebach et al. (2001, 2004, 2005), Traxler et al. (2002), and Just et al. (1996). In question answering and probe verification, participants have to remember the thematic roles that were syntactically licensed in a sentence. Because no implausible sentences are presented, when implausible thematic roles are activated by nonsyntactic mechanisms in constrained sentences, they can be assumed to not be syntactically licensed and can be disregarded. The challenge for participants is to determine whether a plausible thematic role is syntactically licensed in unconstrained sentences. This is more difficult in the syntactically more complex SO sentences. In plausibility judgment, because participants have to determine whether the presented sentence is plausible, implausible thematic roles cannot be disregarded, as they are syntactically licensed in implausible sentences. The challenge for participants in plausibility judgment is to determine whether an implausible thematic role is syntactically licensed in constrained sentences.
Because of this, one strategy for accomplishing the plausibility judgment task would be to activate the thematic roles associated with all possible combinations of nouns and verbs in the trial sentence and to determine whether all were plausible; if they were, the “yes” response could be selected. This strategy would not require an initial syntactic analysis: Only if one proposition were implausible would it be necessary to do a syntactic analysis to determine which propositions were licensed by the syntax and which were activated by nonsyntactic combinations of nouns and verb argument positions. The effect of syntax in plausible unconstrained sentences shows that this strategy was not used. This may be because it is not optimally efficient, and/or because the computation of syntactic structure and its use to determine meaning is obligatory when attending to a sentence.
The behavioral data thus point to two aspects of how the task was performed: (1) the syntactic structure of unconstrained sentences was assigned; (2) implausible thematic roles were activated in plausible constrained sentences. The behavioral data are unable to speak to another aspect of how the task was performed—whether participants engaged in the process that Traxler associated with the late regressive eye movements in his study, which he took to consist of checking the thematic roles in the unconstrained object extracted sentences against the syntax of those sentences. There is pressure to undertake this late check in verification and question answering tasks because of the need to retain only syntactically licensed thematic roles. However, this late check may occur in plausibility judgment as well, possibly because it is an intrinsic (obligatory) part of the comprehension process. This issue becomes important in the interpretation of BOLD signal, as we shall discuss below.
fMRI Results
Beginning with the effect of syntactic structure, BOLD signal increased for SO compared to SS sentences for both unconstrained and constrained sentences. Unlike the behavioral data, there were differences between the effects of syntactic structure in unconstrained and constrained sentences. For constrained sentences, BOLD signal increased in the left MTG/ITG and in the left cuneus; for unconstrained sentences, it increased in the left IFG, DLPFC, PMC, and MTG/STG.
A First Model, in Which Both the Left IFG and the Left MTG Support Initial Parsing and Interpretation
The presence of increases in BOLD signal in temporal and frontal areas for the SO/SS contrast in unconstrained sentences suggests that both these brain regions are involved in processes that differ in assigning the syntactic structure of SO and SS sentences and using that structure to determine meaning in these sentence types. The areas in which these processes occur are similar to those activated by similar contrasts in Fiebach et al. (2001, 2004, 2005) and Just et al. (1996); in this study, the temporal activation extends somewhat more inferiorly and the frontal activation somewhat more anteriorly and dorsally, and the activation was solely in the left hemisphere where the activation in the Just et al. and Fiebach et al. studies was greatest. Fiebach et al. and Just et al. suggested that these processes support parsing and interpretation.
The areas activated by the SO/SS contrast in constrained sentences—L MTG/ITG and cuneus5—might initially be interpreted to be ones where real-world constraints and syntactic structure interact to determine the meaning of the sentences. This interpretation is compatible with a finding reported by Grewe et al. (2007). These authors presented transitive sentences with animate subjects in either subject-before-object or object-before-subject order, and manipulated the animacy of the object in an acceptability judgment task. There was increased BOLD signal in the left superior temporal sulcus, left posterior cingulate, and left MTG for sentences with animate compared to sentences with inanimate objects. The authors interpreted these findings as indicating that these areas utilize the fact that an object is likely to be inanimate to aid in the interpretation of a sentence. The present results might suggest that the left MTG may also be involved in the use of real-world information about the relation of particular nouns and verbs to constrain sentence meaning.6 However, this first-pass interpretation cannot withstand closer scrutiny.
The problem is the absence of an effect of syntactic structure in the left IFG in the constrained sentences. If the presence of an effect of syntactic structure in unconstrained sentences in this area indicates that it is involved in processes that support the assignment and interpretation of the object-extracted structure, why would this region not be activated by the SO/SS contrast in constrained sentences?
One possibility is that the absence of an effect of syntactic structure in the left IFG in constrained sentences is due to less reliance on syntactic information in the determination of the meaning of a sentence in constrained than in the unconstrained sentences. As an anonymous reviewer put it, in the unconstrained condition, competition between nouns for thematic roles may require boosting the syntactic signal; conversely, when knowledge of real-world events supports the syntactically determined interpretation in constrained sentences, less syntactic work is required, leading to less of an effect of the SO–SS contrast in the left IFG. There are, however, two problems with this explanation. First, Traxler et al. (2002) did not find any difference in the effect of the SO/SS contrast on eye fixations in constrained and unconstrained sentences until late in the sentence—after the relative clause had been fixated and eye movements were directed toward the main clause. Traxler et al.’s results strongly suggest that syntactic structure is initially assigned and used to determine sentence meaning in the same way in constrained and unconstrained SO and SS sentences. Second, the absence of an effect of the SO/SS contrast in constrained sentences in the left IFG is due to an increase in BOLD signal for SS constrained sentences, not a decrease in BOLD signal for SO constrained sentences.
Traxler et al.’s (2002) results also make it impossible to use the results here to say that the MTG is the locus of an interaction of real-world constraint and syntactic structure in early sentence processing. If there is no interaction between syntactic structure and real-world factors in the early assignment of syntactic structure and its use to determine sentence meaning in SO and SS sentences, there can be no region of the brain that supports such an interaction.
A Second-pass Model That Distinguishes between Initial Parsing and Interpretation and Reanalysis
Pursuing these considerations, a second pass through these data begins with Traxler et al.’s observation that what differs in constrained and unconstrained SO sentences is not the use of real-world information to support a syntactic analysis as it is being developed and interpreted (i.e., not a reduction in the work of the parser in constrained SO sentences) but a late check of the products of the comprehension process against the syntactic analysis in the unconstrained SO sentences. We postulate that this late check occurs in the left IFG.
A model based on this proposal that can account for the results described above is shown in Figure 5. The model incorporates five main operations: (1) activation of thematic roles on the basis of syntactic analysis; (2) activation of thematic roles on the basis of nonsyntactic analysis (combining all nouns and arguments of verbs); (3) evaluating thematic roles for plausibility in semantic memory; (4) checking thematic roles for whether they originated in the syntax; (5) response selection. The model maintains that sets of thematic roles are activated in two ways: as a result of a syntactic analysis, and by considering each noun as playing a thematic role specified in the verb’s argument structure (Ferreira, 2003). Every activated set of thematic roles is checked for its plausibility in semantic memory (Trueswell, Tanenhaus, & Garnsey, 1994). Implausible thematic roles are checked to see if they originated in the syntax or not (Caplan & Waters, 2003). These checks occur incrementally as soon as a set of thematic roles is activated (Boland, Tanenhaus, Garnsey, & Carlson, 1995; Garnsey, Tanenhaus, & Chapman, 1989). An additional syntactic-origin check is performed on plausible sets of thematic roles at the end of SO unconstrained sentences (Traxler et al., 2002).7 Response selection occurs in one of two ways. When all sets of thematic roles are judged to be plausible in the semantic check, the decision is made that a stimulus is plausible. When a set of thematic roles is judged to be implausible in the semantic check, the decision that a stimulus is implausible is made if that set of implausible thematic roles is determined to be syntactically licensed.
Figure 5.

A model of the operations involved in making plausibility judgments about SO and SS constrained and unconstrained sentences. Syntactic operations in A are localized in left middle and superior temporal structures. The operations labeled B that check thematic roles for their plausibility are localized in areas of the brain that are not identified in this study. The operations labeled C that check thematic roles for their origin in the syntax are localized in left inferior and dorsolateral frontal regions.
Table 2 illustrates the steps involved in making plausibility judgments about SO and SS plausible constrained and unconstrained sentences, showing that there is a syntactic check in three of the four sentence types, corresponding to the sentence types that were associated with increased BOLD signal in the left IFG in this study. To illustrate the operation of the model concretely, we will review the operations involved in making the plausibility decision in plausible constrained and plausible unconstrained sentences.
Table 2.
Outline of Operations that Apply in Making Plausibility Judgments about the Sentence Types in This Study
| Sentence Type | Assign Thematic Roles Syntactically | Assign Thematic Roles Combinatorily | Check Thematic Roles Plausibility | Check Implausible Thematic Roles for Origin | Check Plausible Thematic Roles for Origin |
|---|---|---|---|---|---|
| Operation in Relative Clause | |||||
| SO constrained | + | + | + | + | |
| SO unconstrained | + | + | + | ||
| SS constrained | + | + | + | + | |
| SS unconstrained | + | + | + | ||
| Operation in Main Clause | |||||
| SO constrained | + | + | + | ||
| SO unconstrained | + | + | + | + | |
| SS constrained | + | + | + | ||
| SS unconstrained | + | + | + |
In a plausible constrained sentence, two sets of thematic roles are activated for the relative clause, one on the basis of a syntactic analysis of the presented sentence and one on the basis of possible combinations of nouns and verbs. Both sets are checked against semantic memory. One is implausible, and it is checked for its origin. If it had originated from the syntactic analysis, the stimulus would have been implausible and the “implausible” response could have been made. In a plausible constrained sentence, this is not the case (the implausible set of thematic roles will have originated through the “combinatorial” process), and no decision will be made until the second clause is evaluated. The processing of the second clause is the same as for the relative clause: The two sets of thematic roles activated by this clause are checked for plausibility in semantic memory. In this study, in a plausible constrained sentence, both are plausible (the constraint always applies to the relative clause), and the “plausible” response will be selected.
In a plausible unconstrained sentence, two sets of thematic roles are also activated for the relative clause and are checked against semantic memory. Here the processing of unconstrained and constrained sentences differs. Because both sets of thematic roles are plausible in a plausible unconstrained sentence, neither is checked for its origin. However, because the main clause might be implausible, no decision is made until the second clause is evaluated. Two sets of thematic roles are activated by the main clause and checked for plausibility in semantic memory. Both are plausible and, in principle, the “plausible” response can be selected. This occurs in SS sentences; however, as discussed above, we postulate that Traxler et al.’s view that thematic roles are checked for their origin in unconstrained SO sentences is correct for all comprehension tasks, plausibility judgment, as well as question answering and verification.8
The operations in the model can be mapped onto brain areas. We suggest that left temporal structures support the initial assignment of the structure of object-compared to subject-extracted sentences and the use of that structure to determine thematic roles. This leads to the SO/SS effect in both constrained and unconstrained sentences. Checking thematic roles that are activated for their origin in the syntactic analysis is primarily carried out in left frontal areas. The model requires more checks of thematic roles against the syntax in constrained than in unconstrained SS sentences, leading to a BOLD signal effect in the left IFG in the SS constrained/unconstrained contrast. In unconstrained SO sentences, there is an end-of-sentence check of thematic roles against the syntax that is absent in unconstrained subject-extracted sentences. This leads to a BOLD signal effect in the left IFG of the SO/SS contrast in unconstrained sentences.
Other Possible Models
There are other possibilities regarding the source of BOLD signal in this study that we must consider. One concern that we have raised (Caplan et al., in press) is that what are taken to be effects of parsing and interpretive operations are, in fact, effects of the interaction of these operations with performing a task. We believe this is correct, and we have included operations in our model that are directly related to how the task was performed.
Our model makes different predictions about both the behavioral and neurovascular effects of the SO/SS contrast in different tasks. Behaviorally, it predicts that, unlike the case in verification and question answering, there will be differences in eye fixations during processing of the relative clause in constrained and unconstrained sentences in plausibility judgment. It is expected that the syntax check in constrained sentences will lead to longer first-pass fixations on the relative clause and more regressions to the head noun from the relative clause in constrained than in unconstrained sentences in plausibility judgment. This will occur in both SO and SS sentences (and may be greater in SO sentences). Conversely, in verification and question answering, there is no need to ascertain the plausibility of thematic roles (because all sentences are plausible) and implausible thematic roles will not be checked for their origin. We would therefore expect that there would not be greater BOLD signal for constrained than for unconstrained SS sentences in the left IFG/DLPFC. The SO/SS effect for unconstrained sentences in the left IFG reported by Just et al. (1996) is still expected in verification and question answering, as the syntax check occurs in unconstrained SO sentences.
A second concern is that what Page (2006) calls “epiphenomenal” cognitive activity—cognitive activity correlated with operations under study—may be the source of a BOLD signal effect. The leading candidates for such correlated activity in this study are activating and rehearsing phonological representations, which may occur as part of the process of checking thematic roles for their origin. To evaluate this possibility, we need to make a third (and final) pass through the results, developing a slightly more detailed model of the “syntax check” than we have to this point. This more detailed analysis will also allow us to explain other aspects of the BOLD signal effects.
A Third Model, Which Considers the Nature of the Checking Process in More Detail
The regressive eye movements reported by Traxler et al. (2002) strongly suggest that, in SO unconstrained sentences, thematic roles are checked for whether they are syntactically licensed by a reanalysis process, in which readers reconstruct and reinterpret enough of the syntax to assign at least one thematic role in the relative clause. We can identify three major processes that occur during the “syntax check”: reactivation of the input to the parser/interpreter; reanalysis of the syntactic structure of this reactivated input and reassignment of at least one thematic role;9 comparison of this thematic role against the one being checked.
The first of these sets of operations—review of early parts of the sentence in verbatim form—may lead to conversion of orthographic to phonological representations and rehearsal. Both these processes may occur in a checking phase and not an initial parsing stage, because it is at the checking stage that the reader is placing the greatest emphasis on connecting thematic roles to the syntactic analysis; also, we believe that much evidence supports the view that phonologically based short-term memory is not engaged in the initial analysis and interpretation of a sentence but can be involved in subsequent reanalyses of the products of first-pass processing (Caplan & Waters, 1990, 1999). Orthographic-to-phonological conversion and/or subvocal rehearsal may thus be partially responsible for the increased BOLD signal in the left frontal lobe that we are attributing to the “syntax check.” We wish to point out that, on this view, it is not appropriate to think of these processes as “epiphenomenal” in Page’s sense—they are an integral part of the checking operation.
There are, however, several reasons to doubt that phonological conversion and subvocal rehearsal are the sole sources of the increased BOLD signal in the left frontal lobe. We have previously shown that concurrent articulation, which greatly interferes with subvocal rehearsal, does not eliminate neurovascular effects of the object-/subject-extraction contrast in the left IFG (Caplan et al., 2000). Although this does not entail that subvocal rehearsal is not used when it is not suppressed, it does provide strong evidence that processes other than subvocal rehearsal take place in association with syntactic comprehension in the left IFG. Second, the increase in BOLD signal in SS constrained compared to unconstrained sentences occurred in areas that are more superior and rostral than one might expect to be involved in rehearsal. Finally, the checking process occurred in SS sentences, where it is relatively easy to recover the syntax of the sentence, and where rehearsal may be less required.
Thus, some part of the left IFG/DLPFC activation we ascribe to syntactic checking may be due to reanalysis and reinterpretation. This retains a role for the left IFG in parsing and interpretation. However, consideration of the BOLD signal data in the left temporal ROI provides an argument against this possibility. We have accounted for the increased BOLD for SO compared to SS sentences in this area on the basis of its involvement in initial parsing and interpretation, but we must still explain the increased BOLD signal for SS constrained compared to SS unconstrained sentences in this ROI. The entire pattern of BOLD signal in the left temporal ROI can be accounted for if both initial parsing/interpretation and reanalysis and reinterpretation during the syntax check occur in that area, as follows.
If both initial and secondary parsing/interpretive operations take place in the temporal lobe, the initial parsing/interpretive operations will lead to increased BOLD signal for SO compared to SS sentences in both constrained and unconstrained sentences. The parsing/interpretive operations that occur in the syntax check will lead to increased BOLD signal both for constrained and unconstrained SO sentences and for constrained SS sentences. This leads to the highest level of BOLD signal in SO sentences, which are analyzed with some difficulty initially and then again in the syntax check, regardless of whether they are constrained or unconstrained, a lower level of BOLD signal in constrained SS sentences, which are analyzed more easily initially and again in the syntax check, and the lowest level of BOLD signal for unconstrained SS sentences, which are analyzed once, easily, in the initial parse. This corresponds to the BOLD signal pattern in the left temporal ROI, where BOLD signal was greater for both constrained and unconstrained SO than for both constrained and unconstrained SS sentences and greater for constrained than for unconstrained SS sentences. Given that the pattern of BOLD signal in the left temporal ROI can be explained by postulating that parsing/interpretation occurs in that area both initially and during a syntax check, it is most parsimonious to maintain that these operations occur only in this area.
This leaves comparing thematic roles assigned by the syntax check against those in the proposition being checked as the second possible function that is involved in the syntax check that we postulate occurs in the IFG/DLPFC (as discussed above, rehearsal is also a candidate for this function). We will not discuss this comparison process in any detail, but do note that comparing thematic roles assigned by the syntax check against those in the proposition being checked is a different process than comparing thematic roles against propositions in semantic memory. The first of these operations involves comparing representations maintained in a transient memory system, whereas one of the representations being compared in the second type of comparison is maintained in a long-term memory system. In addition, in this study, the thematic role in the proposition being checked will not be in semantic memory because it is implausible. The claim that the left IFG, especially more anterior parts of this area, is involved in comparing the thematic roles of two propositions receives strong support from other results in the literature (Dapretto & Bookheimer, 1999).
Summary and More General Issues
To summarize, the data in this study suggest that the left MTG supports the assignment of syntactic structure and its use to determine thematic roles; more specifically, that it supports those operations in this process that differ in SO and SS sentences. The left IFG supports part of a process of checking thematic roles that are transiently activated during the course of sentence comprehension for their origin in the syntax, either reactivation and maintenance through rehearsal of the verbatim form of the sentence and/or through comparison of a reactivated thematic role with a thematic role being checked for its origin.
The function we have ascribed to the left IFG/DLPFC bears a similarity to a function that has been localized in roughly this area by Thompson-Schill and her colleagues—response selection from a set of alternatives (Novick, Trueswell, & Thompson-Schill, 2005; Kan & Thompson-Schill, 2004). Although the specific operations we have attributed to this region—rehearsal and comparing thematic roles in two propositions—are not themselves part of what is usually considered the response selection process, checking activated implausible thematic roles against the syntactic structure of the sentence is directly related to the response selection requirements of the plausibility task and the control mechanism that triggers these checks may fall within the set of operations that Thompson-Schill and her colleagues would ascribe to this region.
On our interpretation, our results are incompatible with many models of the functional role of the left IFG in syntactic processing in relative clauses that maintain that this area is the site of assignment of syntactic structure in these sentences. Aside from the articles cited in the Introduction, which include papers of our own, these models include the model developed by Friederici (see Grodzinsky & Friederici, 2006, for a summary) that maintains that first-pass assignment of phrase structure occurs in the left IFG, the model developed by Bornkessel and her colleagues (see Bornkessel & Schlesewsky, 2006, for a review) that a variety of features of noun phrases are mapped onto thematic roles during initial comprehension processes in the left IFG, and the model developed by Grodzinsky and his colleagues (Ben-Shachar, Palti, & Grodzinsky, 2004; Ben-Shachar, Hendler, Kahn, Ben-Bashat, & Grodzinsky, 2003; Grodzinsky, 2000) that the left IFG is responsible for operations that relate the head noun of a relative clause to its position in the relative clause (called “A-bar movement” or “the co-indexation of traces” when discussed in terms used in Chomsky’s theory of syntax). It is not that our data suggest that the left IFG is not involved in the processing of relative clauses, but that they suggest that it is involved in processes that occur after the initial structure and meaning have been assigned, not ones that occur while they are being assigned initially.
To conclude, the process of understanding sentences involves assigning thematic roles on the basis of the syntactic structure of a sentence and on the basis of likely combinations of noun phrases with verbs, and involves a scheduled interaction of these operations with checks of thematic roles for plausibility in semantic memory and for the origin of thematic roles in the syntax of the sentence. This article continues the process of identifying the neural areas that support these operations and their interactions.
Acknowledgments
This research was supported by a grant from the NIDCD (DC02146).
Footnotes
1
In this nomenclature, the first term indicates that the relative clause is attached to the subject (S) of the sentence and the second term indicates that the head of the relative clause is related to subject (S) or object (O) of the relative clause.
2
Whether the working memory system that supports syntactic analysis and sentence interpretation is the same as that that supports other verbal tasks is debated (see Caplan et al., 1999; Just & Carpenter, 1992 for discussion, and Traxler, Williams, Blozis, & Morris, 2005, for data on this issue derived from studies of the sentence types used by Just et al., 1996).
3
For implausible sentences, the terms “constrained” and “unconstrained” sentences refer to the plausible sentences on which the implausible sentences were based; as the examples in the tezxt above show, implausible sentences were themselves neither constrained nor unconstrained.
4
The lack of an interaction between constraint and plausibility suggests that there were effects of constraint in the implausible sentences. This would be surprising because implausible sentences were not constrained or unconstrained (see note 3). In fact, the interaction of constraint and plausibility was significant in the item analyses, where there were no differences between constrained and unconstrained implausible sentences. The effect of constraint in the implausible sentences in the subject analysis likely reflects minor differences in the construction of the implausible sentences.
5
The cuneus and the precuneus are not traditionally viewed as being involved in interpretive processing, but there are scattered pieces of evidence suggesting a role for them in some process related to parsing, interpretation, or reanalysis. Caplan et al. (1999) found activation there in the contrast of SO and OS sentences and Caplan et al. (2007) found that lesions in the superior parietal lobe were associated with deficits in syntactic comprehension tasks. However, these data are not easily interpreted. The materials used in Caplan et al. (1999) were those used by Chen et al. (2006), discussed above, that did not separate animacy and syntactic effects. Caplan et al. (2007) found that comprehension deficits associated with these lesions were task-specific. It may be irrelevant, but both these studies used auditory presentation. The ERP P600 component is that is associated with syntactic processes has a scalp distribution suggestive of high parietal sources, though its neural origin is unknown. The P600 has been attributed to reanalysis, but, as the discussion to follow will show, the SO–SS effect found in this study is not easily attributable to reanalysis because it arose in constrained sentences only. We note that the activity in the cuneus and precuneus was very small (% BOLD signal change from baseline ~1.5% for SO and ~.75% for SS sentences, compared to increases of ~5% for SO sentences and ~3–4% for SS sentences in the MTG and IFG areas). In this article, we shall focus on the activation in frontal and temporal areas in which syntactic effects are more often reported, but the possibility that the parietal effects may reflect parsing, interpretation, or reanalysis should be kept in mind.
6
Unlike the Grewe et al. (2007) data, where there was no effect of subject–object word order or an interaction of subject–object word order and noun animacy on BOLD signal in the temporal area, the present result would require that the left MTG supports both syntactic analysis and the use of real-world information to interpret sentences. The difference in the effects of syntactic structure in the two studies may be due to the syntactic structures used—the SO/SS contrast used here differs from that of scrambling used in Grewe et al. in terms of the linguistic operations that differ in the more and less complex structure, and the differences between the more and less complex structure likely impose a greater load on the parser in the present contrast than in Grewe et al.’s.
7
The one feature of this model that is not based on direct evidence is checking thematic roles against the syntax in SO unconstrained sentences in plausibility judgment (as opposed to question answering, where Traxler et al., 2002, provided evidence for its existence). This feature of the model can be tested by examining eye fixation in sentences such as those used here and by Traxler et al.—Sentences (1) and (2)—in a plausibility judgment task.
8
To remind the reader, this late check presumably occurs automatically because readers are less certain of the meaning of these syntactically complex, semantically unconstrained sentences.
9
There are likely to be differences in parsing and interpretation during initial processing and reanalysis. In reanalysis, parsing/interpretation can terminate when a single thematic role is activated, and/or may consist of a more superficial analysis of the input.
References
- Aguirre GK, Zarahn E, D’Esposito M. The inferential impact of global signal covariates in functional neuroimaging analyses. Neuroimage. 1998;8:302–306. doi: 10.1006/nimg.1998.0367. [DOI] [PubMed] [Google Scholar]
- Ben-Shachar M, Hendler T, Kahn I, Ben-Bashat D, Grodzinsky Y. The neural reality of syntactic transformations: Evidence from fMRI. Psychological Science. 2003;14:433–440. doi: 10.1111/1467-9280.01459. [DOI] [PubMed] [Google Scholar]
- Ben-Shachar M, Palti D, Grodzinsky Y. The neural correlates of syntactic movement: Converging evidence from two fMRI experiments. Neuroimage. 2004;21:1320–1336. doi: 10.1016/j.neuroimage.2003.11.027. [DOI] [PubMed] [Google Scholar]
- Boland JE, Tanenhaus MK, Garnsey SM, Carlson GN. Verb argument structure in parsing and interpretation: Evidence from Wh-questions. Journal of Memory and Language. 1995;34:774–806. [Google Scholar]
- Bornkessel I, Schlesewsky M. The extended argument dependency model: A neurocognitive approach to sentence comprehension across languages. Psychological Review. 2006;113:787–821. doi: 10.1037/0033-295X.113.4.787. [DOI] [PubMed] [Google Scholar]
- Burock MA, Buckner RL, Woldorff MG, Rosen BR, Dale AM. Randomized event-related experimental designs allow for extremely rapid presentation rates using functional MRI. NeuroReport. 1998;9:3735–3739. doi: 10.1097/00001756-199811160-00030. [DOI] [PubMed] [Google Scholar]
- Burock MA, Dale AM. Estimation and detection of event-related fMRI signals with temporally correlated noise: A statistically efficient and unbiased approach. Human Brain Mapping. 2000;11:249–260. doi: 10.1002/1097-0193(200012)11:4<249::AID-HBM20>3.0.CO;2-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Alpert N, Waters G. Effects of syntactic structure and propositional number of patterns of regional cerebral blood flow. Journal of Cognitive Neuroscience. 1998;10:541–552. doi: 10.1162/089892998562843. [DOI] [PubMed] [Google Scholar]
- Caplan D, Alpert N, Waters G. PET studies of sentence processing with auditory sentence presentation. Neuroimage. 1999;9:343–351. doi: 10.1006/nimg.1998.0412. [DOI] [PubMed] [Google Scholar]
- Caplan D, Alpert N, Waters G, Olivieri A. Activation of Broca’s area by syntactic processing under conditions of concurrent articulation. Human Brain Mapping. 2000;9:65–71. doi: 10.1002/(SICI)1097-0193(200002)9:2<65::AID-HBM1>3.0.CO;2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Chen E. Using fMRI to discover cognitive operations. Cortex. 2006;42:393–395. doi: 10.1016/s0010-9452(08)70366-6. [DOI] [PubMed] [Google Scholar]
- Caplan D, Chen E, Waters G. Task-dependent and task-independent neurovascular responses to syntactic processing. Cortex. doi: 10.1016/j.cortex.2006.06.005. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Vijayan S, Kuperberg G, West C, Waters G, Greve D, et al. Vascular responses to syntactic processing: Event-related fMRI study of relative clauses. Human Brain Mapping. 2002;15:26–38. doi: 10.1002/hbm.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Waters G. Short-term memory and language comprehension: A critical review of the neuropsychological literature. In: Shallice T, Vallar G, editors. The neuropsychology of short-term memory. Cambridge: Cambridge University Press; 1990. pp. 337–389. [Google Scholar]
- Caplan D, Waters G, Kennedy D, Alpert N, Makris N, Dede G, et al. A study of syntactic processing in aphasia II: Neurological aspects. Brain Language. 2007;101:151–177. doi: 10.1016/j.bandl.2006.06.226. [DOI] [PubMed] [Google Scholar]
- Caplan D, Waters GS. Verbal working memory and sentence comprehension. Behavioral and Brain Sciences. 1999;22:77–126. doi: 10.1017/s0140525x99001788. [DOI] [PubMed] [Google Scholar]
- Caplan D, Waters GS. On-line syntactic processing in aphasia: Studies with auditory moving windows presentation. Brain and Language. 2003;84:222–249. doi: 10.1016/s0093-934x(02)00514-x. [DOI] [PubMed] [Google Scholar]
- Chen E, West WC, Waters G, Caplan D. Determinants of BOLD signal correlates of processing object-extracted relative clauses. Cortex. 2006;42:591–604. doi: 10.1016/s0010-9452(08)70397-6. [DOI] [PubMed] [Google Scholar]
- Cohen JD, MacWhinney B, Flatt M, Provost J. PsyScope: An interactive graphic system for designing and controlling experiments in the psychology laboratory using Macintosh computers. Behavior Research: Methods, Instruments and Computers. 1993;25:257–271. [Google Scholar]
- Cooke A, Zurif EB, DeVita C, Alsop D, Koenig P, Detre J, et al. Neural basis for sentence comprehension: Grammatical and short-term memory components. Human Brain Mapping. 2001;15:80–94. doi: 10.1002/hbm.10006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale AM. Optimal experimental design for event-related fMRI. Human Brain Mapping. 1999;8:109–114. doi: 10.1002/(SICI)1097-0193(1999)8:2/3<109::AID-HBM7>3.0.CO;2-W. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale AM, Buckner RL. Selective averaging of rapidly presented individual trials using fMRI. Human Brain Mapping. 1997;5:329–340. doi: 10.1002/(SICI)1097-0193(1997)5:5<329::AID-HBM1>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- Dapretto M, Bookheimer SY. Form and content: Dissociating syntax and semantics in sentence comprehension. Neuron. 1999;24:427–432. doi: 10.1016/s0896-6273(00)80855-7. [DOI] [PubMed] [Google Scholar]
- Ferreira F. The misinterpretation of noncanonical sentences. Cognitive Psychology. 2003;47:164–203. doi: 10.1016/s0010-0285(03)00005-7. [DOI] [PubMed] [Google Scholar]
- Fiebach CJ, Schlesewsky M, Friederici AD. Syntactic working memory and the establishment of filler-gap dependencies: Insights from ERPs and fMRI. Journal of Psycholinguistic Research. 2001;30:321–338. doi: 10.1023/a:1010447102554. [DOI] [PubMed] [Google Scholar]
- Fiebach CJ, Schlesewsky M, Lohmann G. Revisiting the role of Broca’s area in sentence processing: Syntactic integration versus syntactic working memory. Human Brain Mapping. 2005;24:79–91. doi: 10.1002/hbm.20070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiebach J, Vos SH, Friederici AD. Neural correlates of syntactic ambiguity in sentence comprehension for low and high span readers. Journal of Cognitive Neuroscience. 2004;16:1562–1575. doi: 10.1162/0898929042568479. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Holmes A, Poline JB, Price CJ, Frith CD. Detecting activations in PET and fMRI: Levels of inference and power. Neuroimage. 1996;40:223–235. doi: 10.1006/nimg.1996.0074. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Holmes AP, Worsley KJ, Poline JP, Frith CD, Frackowiak RSJ. Statistical parametric maps in functional imaging: A general approach. Human Brain Mapping. 1995;2:189–210. [Google Scholar]
- Garnsey SM, Tanenhaus MK, Chapman RM. Evoked potentials and the study of sentence comprehension. Journal of Psycholinguistic Research. 1989;18:51–60. doi: 10.1007/BF01069046. [DOI] [PubMed] [Google Scholar]
- Grewe T, Bornkessel I, Zysset S, Wiese R, von Cramon DY, Schlesewsky M. The role of the posterior superior temporal sulcus in the processing of unmarked transitivity. Neuroimage. 2007;35:343–352. doi: 10.1016/j.neuroimage.2006.11.045. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y. The neurology of syntax: Language use without Broca’s area. Behavioral and Brain Sciences. 2000;23:47–117. doi: 10.1017/s0140525x00002399. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y, Friederici A. Neuroimaging of syntax and syntactic processing. Current Opinion in Neurobiology. 2006;16:240–246. doi: 10.1016/j.conb.2006.03.007. [DOI] [PubMed] [Google Scholar]
- Indefrey P, Hagoort P, Herzog H, Seitz R, Brown C. Syntactic processing in the left prefrontal cortex in independent of lexical meaning. Neuroimage. 2001;14:546–555. doi: 10.1006/nimg.2001.0867. [DOI] [PubMed] [Google Scholar]
- Just MA, Carpenter PA. A capacity theory of comprehension: Individual differences in working memory. Psychological Review. 1992;99:122–149. doi: 10.1037/0033-295x.99.1.122. [DOI] [PubMed] [Google Scholar]
- Just MA, Carpenter PA, Keller TA, Eddy WF, Thulborn KR. Brain activation modulated by sentence comprehension. Science. 1996;274:114–116. doi: 10.1126/science.274.5284.114. [DOI] [PubMed] [Google Scholar]
- Kan IP, Thompson-Schill SL. Selection from perceptual and conceptual representations. Cognitive, Affective, & Behavioral Neuroscience. 2004;4:466–482. doi: 10.3758/cabn.4.4.466. [DOI] [PubMed] [Google Scholar]
- Novick JM, Trueswell JC, Thompson-Schill SL. Cognitive control and parsing: Reexamining the role of Broca’s area in sentence comprehension. Cognitive, Affective, & Behavioral Neuroscience. 2005;5:263–281. doi: 10.3758/cabn.5.3.263. [DOI] [PubMed] [Google Scholar]
- Page MP. What can functional imaging tell the experimental psychologist? Cortex. 2006;42:428–443. doi: 10.1016/s0010-9452(08)70375-7. [DOI] [PubMed] [Google Scholar]
- Stromswold K, Caplan D, Alpert N, Rauch S. Localization of syntactic comprehension by positron emission tomography. Brain and Language. 1996;52:452–473. doi: 10.1006/brln.1996.0024. [DOI] [PubMed] [Google Scholar]
- Traxler M, Morris R, Seely R. Processing subject and object relative clauses: Evidence from eye movements. Journal of Memory and Language. 2002;47:69–90. [Google Scholar]
- Traxler MJ, Williams RS, Blozis SA, Morris RK. Working memory, animacy, and verb class in the processing of relative clauses. Journal of Memory and Language. 2005;53:204–224. [Google Scholar]
- Trueswell JC, Tanenhaus MK, Garnsey SM. Semantic influences on parsing: Use of thematic role information in syntactic ambiguity resolution. Journal of Memory and Language. 1994;33:285–318. [Google Scholar]
- Waters G, Caplan D, Alpert N, Stanczak L. Individual differences in rCBF correlates of syntactic processing in sentence comprehension: Effects of working memory and speed of processing. Neuroimage. 2003;19:101–112. doi: 10.1016/s1053-8119(03)00007-7. [DOI] [PubMed] [Google Scholar]
- Worsley KJ. Local maxima and the expected Euler characteristic of excursion sets of X2, F and t fields. Advances in Applied Probability. 1994;26:13–42. [Google Scholar]
- Worsley KJ, Friston KJ. Analysis of fMRI time-series revisited—Again. Neuroimage. 1995;3:173–181. doi: 10.1006/nimg.1995.1023. [DOI] [PubMed] [Google Scholar]