reseptor – Store medisinske leksikon (original) (raw)
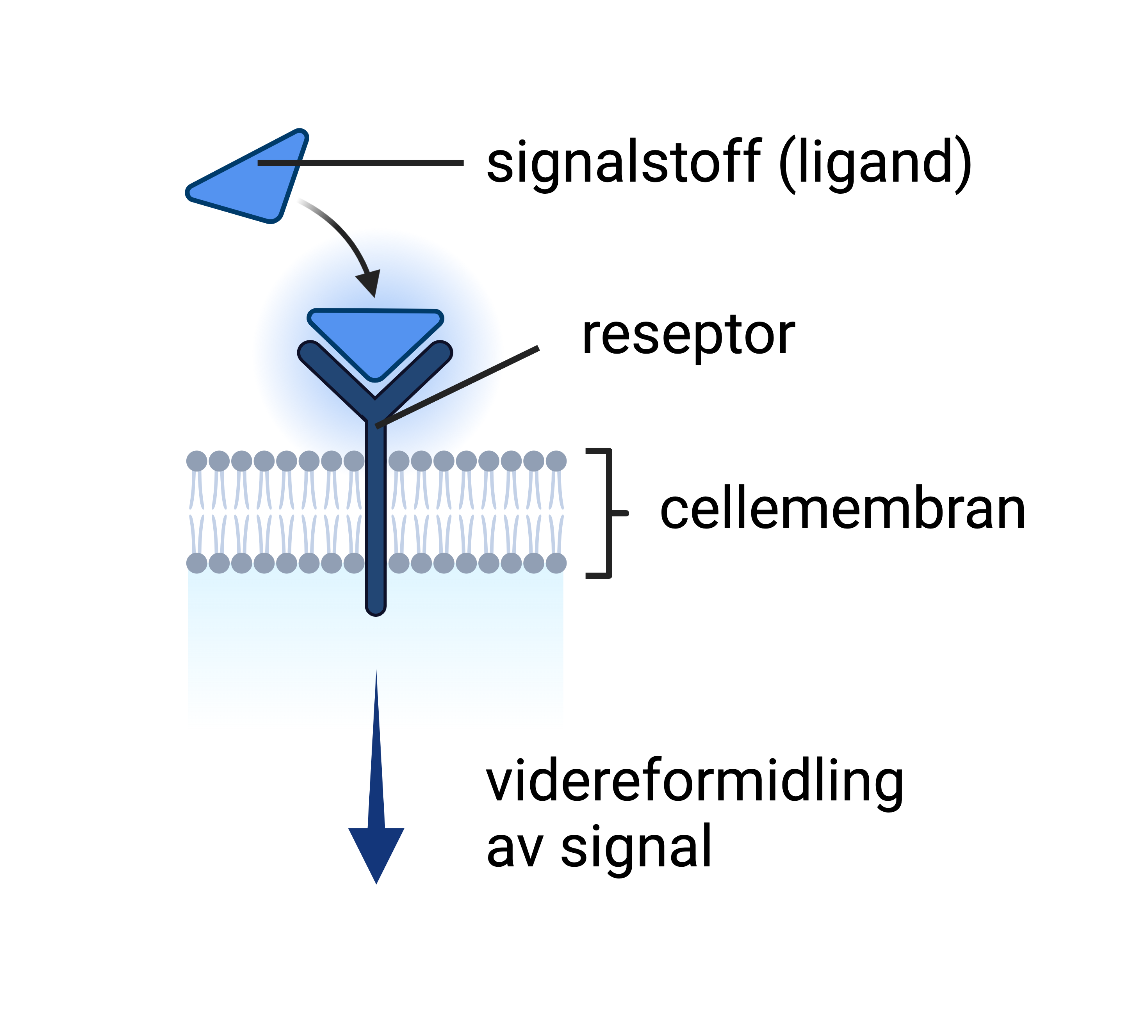
Illustrasjonen viser et signalstoff (ligand) som kommer på plass i bindingssetet på en reseptor. Det finnes egne reseptorer for alle typer signalstoffer, og signalstoffer passer kun i sin respektive reseptor. Når et signalstoff binder til reseptoren vil signalet videreformidles inne i cellen.
En reseptor er et bindingssted for et signalstoff. Reseptorer er lokalisert på utsiden av cellemembranen eller inne i en celle. Reseptorer er bygd opp av ett eller flere proteiner. Signalstoffer er molekyler som skilles ut fra enkelte celler og som kan fremkalle en respons i en målcelle. Eksempler på signalstoffer er hormoner og nevrotransmittere. Slike signalstoffer kalles ofte for ligander. For at en ligand skal ha en effekt på målcellen, er målcellen nødt til å ha en reseptor som gjenkjenner liganden. Det finnes også reseptorer som reagerer på stoffer fra mikroorganismer eller stoffer i omgivelsene våre. Disse reseptorene er som regel på immunceller.
Faktaboks
av latin recipere, ‘ta imot’
Det finnes flere typer kommunikasjon mellom celler. Når en celle skiller ut et signalstoff som den selv responderer på, kalles det autokrin signalering. Dersom en celle sender et signal til en nærliggende celle slik at signalstoffet kan diffundere til målcellen i væsken mellom cellene (ekstracellulærvæsken), kalles det parakrin signalering. Dersom målcellen ligger langt unna cellen som skiller ut signalstoffet, kalles det endokrin signalering. Ofte er det egne spesialiserte kjertler og celler som skiller ut slike endokrine signalstoffer, som for eksempel binyrene eller bukspyttkjertelen som henholdsvis skiller ut adrenalin og insulin. Slike signalstoffer fraktes så med blodbanen til de treffer sine målceller og binder til deres reseptorer.
En celle kan ha mange forskjellige typer reseptorer, og dermed være i stand til å reagere på en rekke forskjellige typer signaler. Hvilke typer reseptorer en celle har avhenger av hva slags type celle det er; en hudcelle og en immuncelle vil ha ulike typer reseptorer.
Oppbygning
En reseptor er bygd opp av ett eller flere proteiner som utgjør en enhet. Hver reseptor har et såkalt bindingssete som er spesifikt for nettopp sitt signalstoff, og kan derfor ikke aktiveres av andre signalstoffer. I enkelte tilfeller passer imidlertid også andre molekyler inn i bindingssetet til reseptoren, og disse kan enten aktivere reseptoren slik som det naturlige signalstoffet, eller de kan hemme reseptoren. En rekke legemidler og giftstoffer fungerer ved at de utkonkurrerer det naturlige signalstoffet og hemmer reseptoren sin funksjon.
Enkelte reseptorer binder sitt respektive signalstoff veldig raskt og med stor tiltrekningskraft, med såkalt høy affinitet. Om et signalstoff har høy eller lav affinitet påvirker hvor sterkt det binder til reseptoren, og hvor mye av signalstoffet som må til for at reseptoren skal bli aktivert. Det trengs lite av et signalstoff med høy affinitet, mens det trengs mye av et signalstoff med lav affinitet for å aktivere en reseptor.
Aktivering og deaktivering
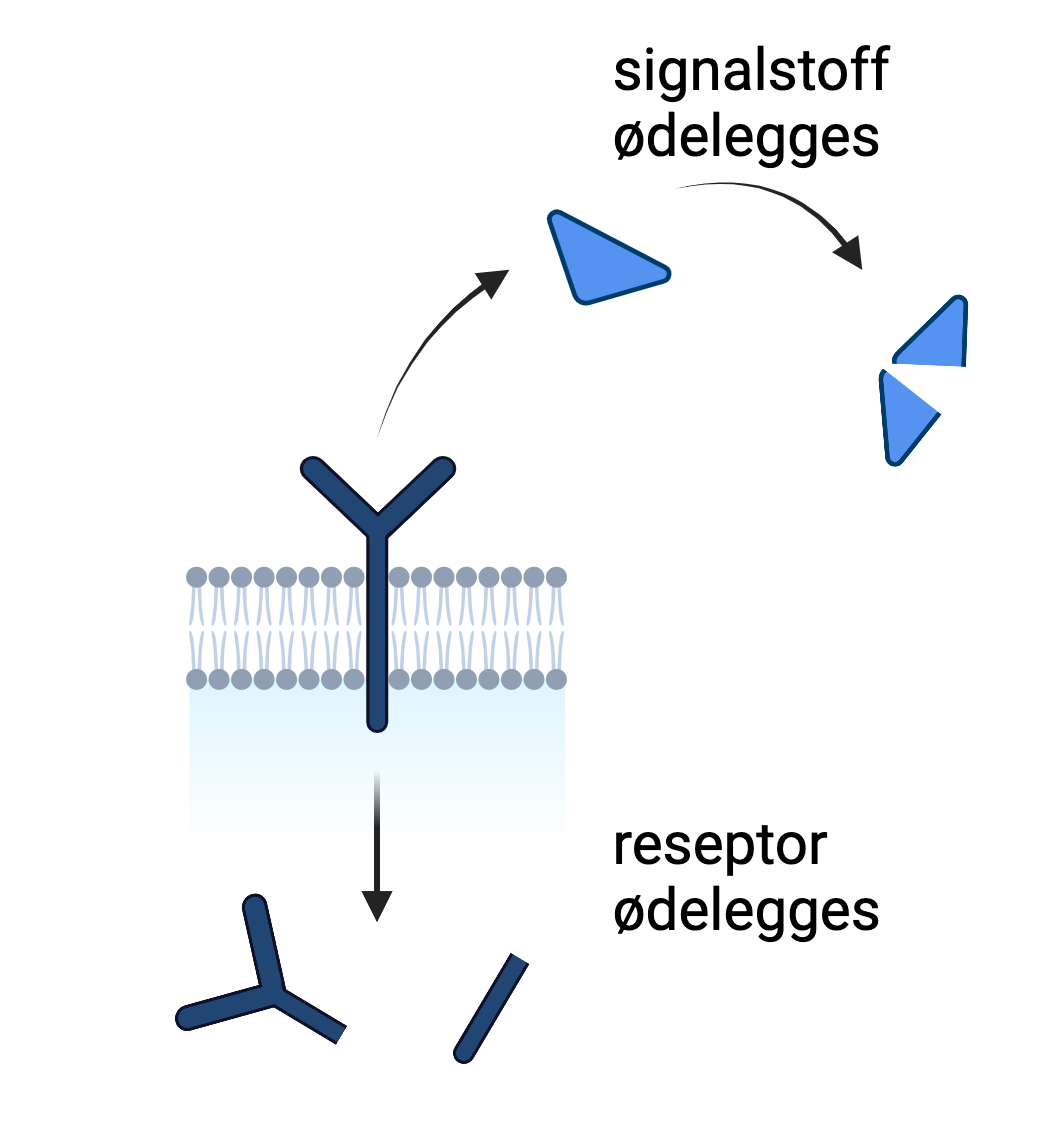
Når et signal skal avsluttes kan for eksempel signalstoffet løsne fra bindingssetet og brytes ned. I noen tilfeller er det reseptoren selv som brytes ned, slik at videre signalering ikke kan finne sted.
Når en reseptor binder sitt signalstoff, vil dette ofte resultere i at reseptoren endrer fysisk form, ofte kalt en konformasjonsendring. Denne formendringen kan tilgjengeliggjøre deler av reseptoren som i sin tur viderefører signalet ved å påvirke andre proteiner eller molekyler inne i cellen. Dermed overføres signalet fra utsiden av cellen, altså fra det ekstracellulære miljøet, til innsiden av cellen, det intracellulære miljøet.
På samme måte som at reseptoren endrer seg ved aktivering, er det like viktig at den endrer seg når signalet tar slutt. Dette kan foregå på flere måter, for eksempel ved at signalstoffet eller reseptoren i seg selv brytes ned, eller ved at andre proteiner påvirker reseptoren slik at den returnerer til sin opprinnelige form. I noen tilfeller tar dette tid, slik at et signalstoff som utløser en respons ikke kan utløse samme respons igjen umiddelbart.
Typer reseptorer
Man skiller gjerne reseptorer ut fra hvordan de fremkaller en respons i målcellen. Dette har også betydning for hastigheten på signaloverføringen.
Metabotrope reseptorer virker langsomt og involverer flere ledd enn motparten, ionotrope reseptorer, som er involvert i svært hurtig signalformidling. Reseptoren for dopamin, som vi blant annet finner i hjernen, er et eksempel på en metabotrop reseptor. Når dopamin binder til en dopaminreseptor, skjer det en rekke endringer i cellen, og mange av disse varer lenge. Nevrotransmitteren acetylkolin kan binde flere typer reseptorer, blant annet nikotinreseptoren som er involvert i muskelsammentrekning. Nikotinreseptoren er en ionotrop reseptorer, og effekten av aktivering er svært hurtig og kortvarig.
Man skiller også mellom membranreseptorer og intracellulære reseptorer. Membranreseptorer er reseptorer som finnes på utsiden av cellen, forbundet med cellemembranen. Slike reseptorer har gjerne en vannavstøtende (hydrofob) del som er festet i, eller som går gjennom cellemembranen. Membranreseptorer aktiveres ved at en korresponderende ligand binder til reseptoren på ekstracellulær side. Den aktiverte reseptoren videreformidler i sin tur signalet intracellulært.
G-proteinkoblede reseptorer
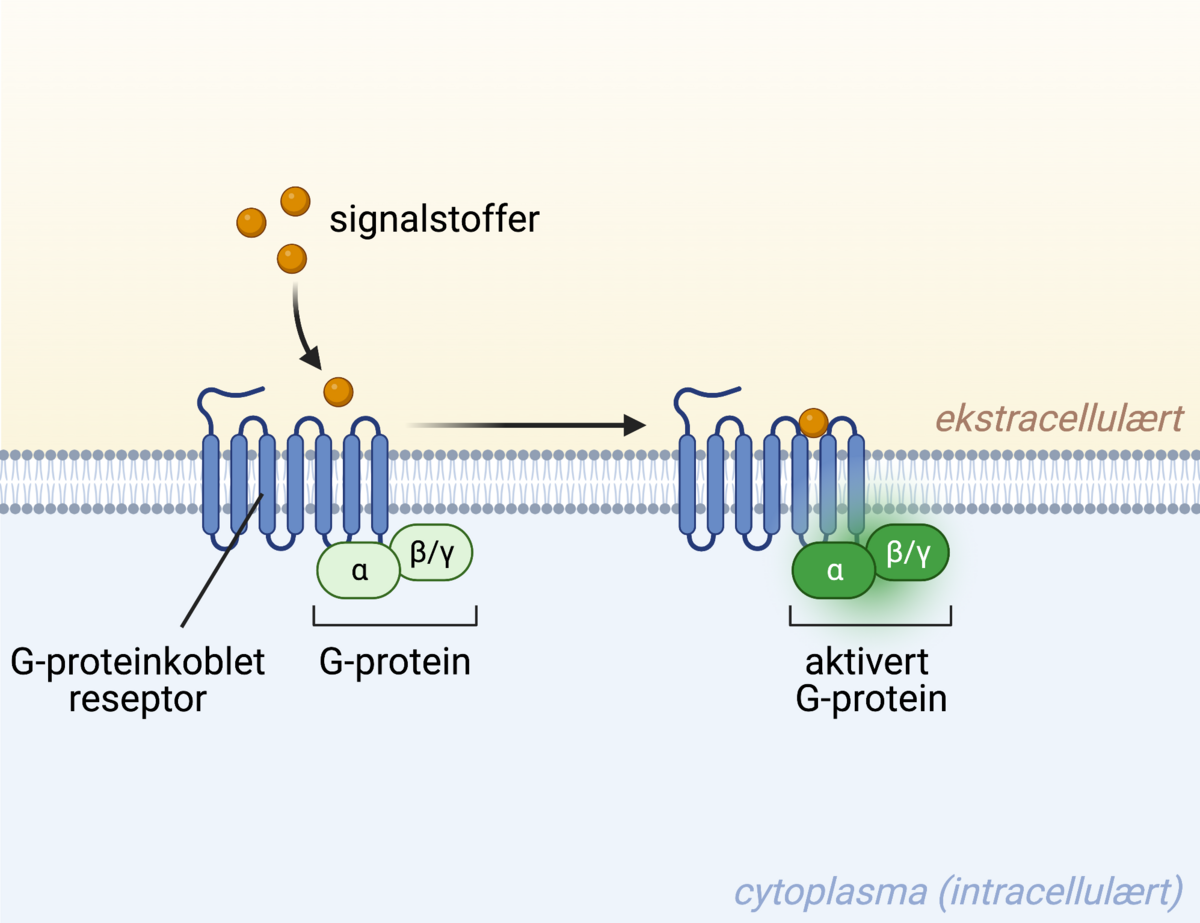
En G-proteinkoblet reseptor går gjennom membranen syv ganger. Signalstoffet kan binde til reseptoren på utsiden av cellen, og dette fører i sin tur til aktivering av G-proteinkomplekset som er bundet til reseptoren på innsiden av cellen.
G-proteinkoblede reseptorer (ofte forkortet GPCR, fra det engelske G-protein coupled receptor) er den største klassen med membranreseptorer, og varierer svært mye i funksjon. Felles for disse reseptorene er at de går gjennom cellemembranen syv ganger, de har altså syv transmembrane domener.
På innsiden av membranen er reseptoren bundet til et såkalt G-protein. Dette proteinet består av flere deler, kjent som subenheter, og en av subenhetene har bundet et lite molekyl kalt GDP (forkortelse for guanosin_di_fosfat) til seg. GDP inneholder to fosfatmolekyler. Ved binding av ligand til reseptoren vil G-proteinet aktiveres og endre form, subenhetene vil gli fra hverandre, og GDP vil byttes ut med GTP (guanosin_tri_fosfat), som inneholder tre fosfatmolekyler. GTP har en høyere energistatus enn GDP.
Det aktiverte G-proteinet kan videre aktivere andre intracellulære proteiner og enzymer, som resulterer i dannelsen av såkalte sekundære budbringere. De kalles sekundære, fordi man ser på liganden som binder til reseptoren som den første (primære) budbringeren. De sekundære budbringerne videreformidler og forsterker signalet.
Signaleringen avsluttes ved at et fosfatmolekyl på GTP spaltes av, og GDP dannes. Subenhetene til G-proteinet vil samles igjen, og hele komplekset vil innta samme form som det hadde før liganden aktiverte reseptoren, og den er klar for å aktiveres igjen.
Enzymkoblede reseptorer
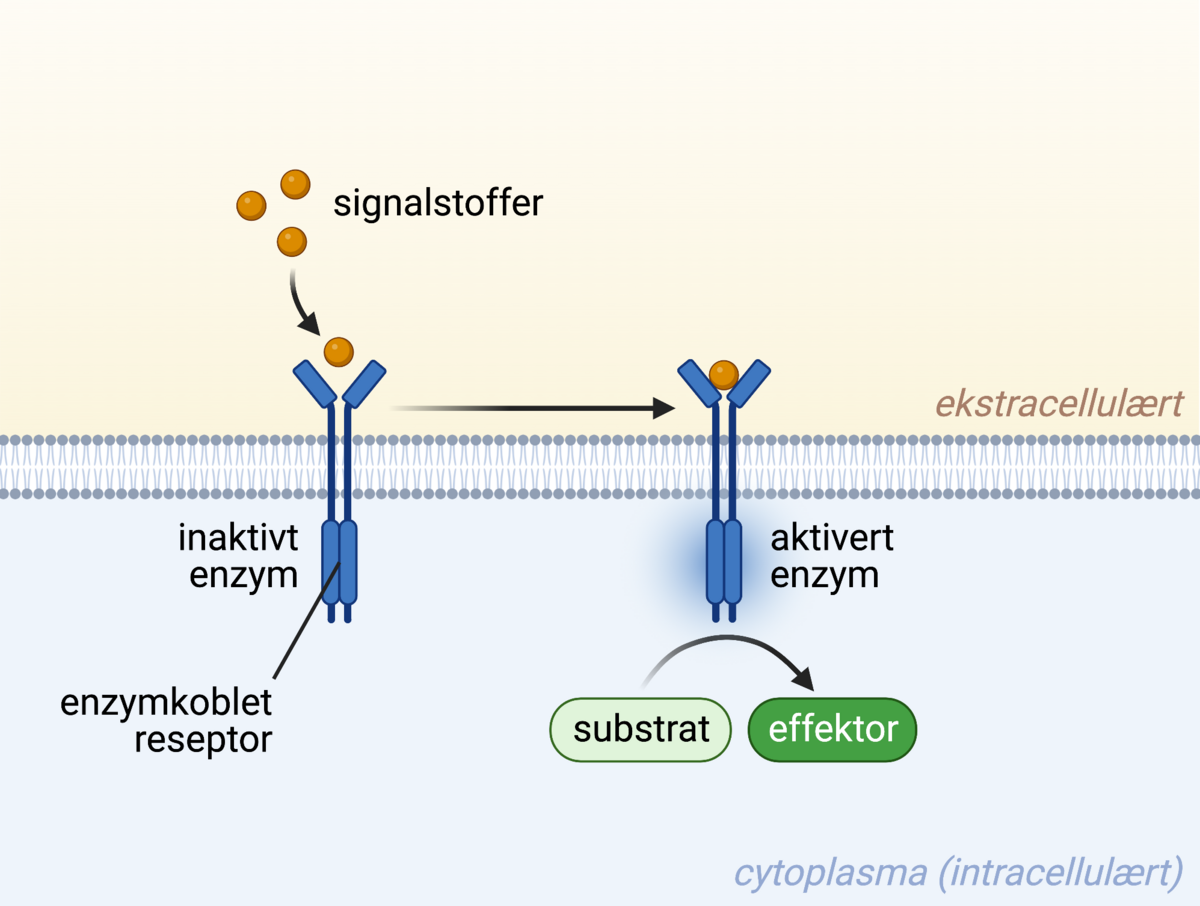
En enzymkoblet reseptor er en type reseptor som har enzymatisk aktivitet på innsiden av cellen. Når et signalstoff binder til reseptoren fører det til aktivering av reseptoren, som i sin tur påvirker andre proteiner innad i cellen, ved å for eksempel feste et fosfatmolekyl på dem.
Enzymkoblede reseptorer er membranreseptorer som enten direkte eller indirekte innehar enzymatisk aktivitet. Enzymatisk aktivitet vil si at de for eksempel kan sette sammen eller spalte molekyler, eller aktivere andre proteiner og enzymer.
Enzymkoblede reseptorer kan ha direkte enzymatisk aktivitet ved at de for eksempel kan fosforylere andre proteiner inne i cellen. Å fosforylere vil si å feste et fosfatmolekyl på et protein, og enzymer som gjør dette kalles kinaser. I de fleste tilfeller vil et protein eller enzym gå over til en aktiv tilstand dersom det blir fosforylert.
Indirekte enzymatisk aktivitet finner sted når reseptoren er forbundet med eller koblet til enzymer inne i cellen. Slike enzymer aktiveres når reseptoren binder en ligand og videreformidler signalet til et enzym inni cellen.
Et eksempel på enzymkoblede reseptorer er reseptor tyrosin kinaser. Disse reseptorene består av flere deler med bindingssete for ligand på utsiden av cellen, og en katalytisk (enzymatisk) del med kinaseaktivitet på innsiden av cellen. Ved binding av ligand vil subenhetene finne sammen, og aktivere hverandre i en prosess kalt kryssfosforylering. Fosforyleringen skjer på aminosyren tyrosin på begge subenhetene, derav navnet tyrosin kinaser. Disse fosforylerte tyrosinene danner i sin tur bindingsseter for andre intracellulære proteiner, som videre kan tilkalle eller aktivere proteiner for å fremkalle en respons.
Ligandstyrte ionekanaler
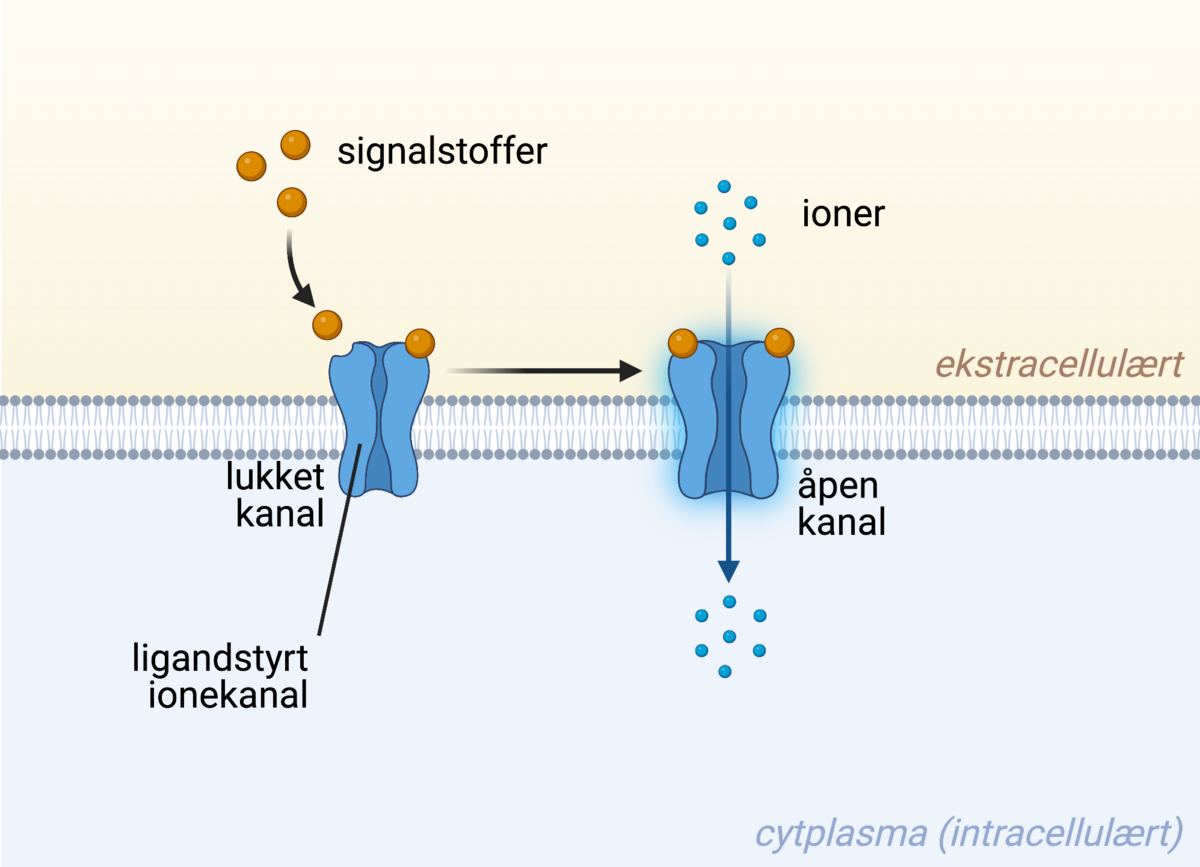
En ligandstyrt ionekanal er en form for reseptor som åpnes og slipper inn (eller ut) ioner dersom den blir aktivert. Dette fører til en rask endring av konsentrasjonen av ioner i cellen.
Ligandstyrte ionekanaler er membranreseptorer som ved binding av ligand åpner en kanal som tillater passasje av spesifikke ioner inn eller ut av cellen. På denne måten kan cellen raskt endre konsentrasjonen av et gitt ion inni cellen som respons til et signal. Slike kanaler kan for eksempel bare slippe gjennom natrium-ioner (Na+) eller bare kalsium-ioner (Ca2+). Hvilken retning ionene passerer avhenger av konsentrasjonen til ionene på innsiden og utsiden av cellen. Ligandstyrte ionekanaler er såkalte ionotrope reseptorer, og har en svært rask virkning på cellen.
Intracellulære reseptorer

Intracellulære reseptorer befinner seg inne i cellen og har ingen direkte tilknytning til cellemembranen. De aktiveres når signalstoffer som kan passere cellemembranen binder til bindingssetet. I de fleste tilfeller fører dette til at reseptor og signalstoff-komplekset beveger seg inn til arvematerialet der det kan påvirke hvilke gener som uttrykkes.
Intracellulære reseptorer er lokalisert inne i cellen. Disse binder gjerne vannavstøtende (hydrofobe) ligander som fritt kan passere cellemembranen, slik som steroidhormonene østrogen og testosteron. I mange tilfeller vil aktivering av slike reseptorer gjøre at de forflytter seg til cellekjernen der de kan regulere et eller flere gener i DNA-et. Dermed kan de også anses for å være transkripsjonsfaktorer.