Recombinase-based conditional and reversible gene regulation via XTR alleles (original) (raw)
Introduction
Identifying causal relationships between gene function and the physiological programmes they control is a central goal of biological research and an unmet challenge in cancer. Although loss-of-function mutations in tumour suppressor genes are the most common type of genetic alteration in cancer, identifying how these genes function within physiologically relevant in vivo settings remains difficult. Restoration of tumour suppressor gene function has the potential to identify relevant programmes of tumour suppression in physiologically diverse settings. This strategy rests on the premise that latent tumour suppressive programmes are poised to react to the reintroduction of tumour suppressor genes, and that the ensuing changes they orchestrate will expose relevant mechanisms of tumour suppression. In addition, experimental restoration of tumour suppressor genes highlights the potential of future therapies aimed at restoring tumour suppressive pathways to treat cancer. Several different approaches have been used to restore endogenous gene expression in vivo and these have identified tumour suppression programmes that are tissue and context specific1,2,3,4,5,6. Despite the power of these approaches, their widespread application to in vivo biological systems has been constrained by multiple technological hurdles and limitations inherent to each method (see Discussion).
Conditional approaches to inactivate gene expression in the mouse commonly rely on expression of specialized site-specific recombinases such as Cre and Flp. These recombinases facilitate deletion of DNA sequences that are flanked by similarly oriented DNA elements called loxP or FRT, respectively. Cre and Flp can also facilitate the reversible inversion of DNA sequences that intervene inversely oriented loxP or FRT sites. However, the inherent reversibility of the inversion reaction is often problematic due to the mosaic pattern of DNA rearrangements that result. To drive reproducible and stable DNA inversions, mutant loxP and FRT sites have been used to facilitate the permanent inversion of DNA sequences containing gene traps, to create conditional alleles in mouse embryonic stem (ES) cells and adult mice7,8,9,10.
Here we developed a synthetic gene switch, XTR, that uses mutant loxP sites to invert a synthetic fluorescent reporter trap element to drive conditional inactivation of endogenous genes. In addition, we engineered the capability to restore accurate endogenous gene expression via Flp-dependent deletion of the synthetic gene switch in a temporally controlled manner. Designed for broad use and compatibility with established recombinase-based tools, XTR alleles provide a powerful method to establish causal relationships between genes and the greater physiological programmes they regulate in specific contexts.
Results
XTR allele design
Our goal was to develop a single unifying allelic system that works in concert with existing genetically engineered mouse models of human cancer using the site-specific DNA recombinases Cre and Flp, and would enable temporally controlled gene inactivation followed by accurate and inducible gene restoration (Fig. 1a). To take advantage of the large array of methods to deliver Cre and the diverse model systems that rely on Cre-driven cancer initiation, we developed a ‘double-floxed’ gene trap that can be stably inverted by Cre, to conditionally inactivate virtually any gene of interest. The gene trap consists of a strong adenoviral 40 splice acceptor (SA), followed by the coding sequence for green fluorescent protein (GFP) and the SV40 polyadenylation transcription termination sequence (Fig. 1b). We chose this DNA element because of its small size, ability to report expression from the endogenous host gene once trapped and its proven ability to recapitulate gene knockout phenotypes in cells11,12,13,14,15,16. To enable stable inversion of the gene trap we oriented inverted pairs of Lox5171 and Lox2722 sites in an alternating manner such that each pair flanked the gene trap9,17. This double-floxed organization results in the permanent inversion of the gene trap after two successive Cre-mediated recombination reactions (Fig. 1b)7,8. In addition, we flanked the entire gene trap with FRT sites to enable deletion of the gene trap by Flp recombinase and restoration of the targeted gene (Fig. 1b). The GFP reporter feature of the XTR system requires in-frame splicing from the upstream exon in instances where XTR is inserted downstream of the translation initiation site of the host gene. To allow general applicability, we developed independent XTR allele targeting vectors with the SA–GFP in each of the three reading frames (Supplementary Fig. 1).
Figure 1: XTR alleles facilitate Cre-mediated inactivation and subsequent Flp-dependent restoration of endogenous genes.
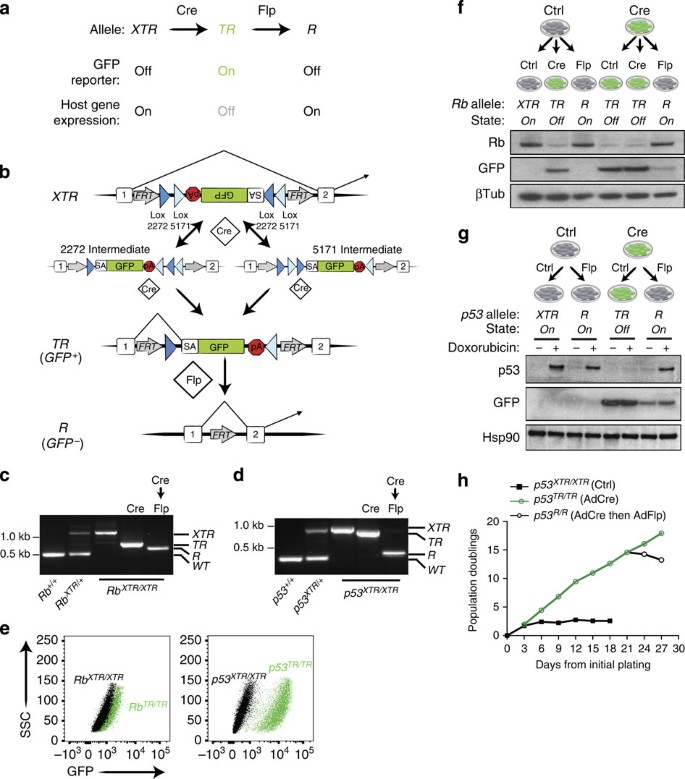
(a) Cre converts XTR alleles to the TR allele, thereby inactivating gene function. Flp restores gene function by conversion to R. (b) Schematic of the XTR allele. Cre drives irreversible inversion of a double-floxed gene trap consisting of a splice acceptor (SA) enhanced GFP complementary DNA and the polyadenylation transcriptional terminator sequence (pA). Inversion can proceed either through sequential action of Cre on Lox2272 sites then Lox5171 sites (2272 intermediate) or Lox5171 then Lox2272 sites (5171 intermediate). Stable inversion accepts splicing from upstream exons in the host gene, reads out GFP expression and then terminates transcription, leading to functional inactivation of the host gene’s expression. Flp drives deletion of the gene trap (SA-GFP-pA), thereby restoring normal splicing of the host gene. AdCre and AdCre followed by AdFlpO treatment is indicated (c,d). PCR-based detection of Rb (c) and p53 (d) XTR, TR, R and wild-type (+) alleles in MEFs of the indicated genotype. (e) Detection of GFP reporter expression from TR alleles in Rb TR/TR and p53 TR/TR MEFs by flow cytometry analysis. Representative of ⩾3 cell lines. (f) Immunoblot analysis of Rb and GFP expression in Rb XTR/XTR MEFs treated sequentially with AdCre and/or AdFlpO as indicated. β-Tubulin is a loading control. (g) Immunoblot analysis of p53 and GFP expression in p53 XTR/XTR MEFs treated sequentially with AdCre and/or AdFlpO as indicated. Hsp90 is a loading control. (h) 3T3 proliferation assay of p53 XTR/XTR MEFs treated sequentially with AdCre (day 3) then AdFlpO (day 21) as indicated. Representative of two p53 XTR/XTR cell lines.
Targeting and validation of XTR alleles at Rb1 and Trp53
To validate this system and generate alleles that will be of widespread use to the cancer biology community, we targeted the XTR cassette to evolutionarily non-conserved regions of the first intron of the Trp53 and Rb1 genes (p53 and Rb hereafter). Correctly, targeted ES cells gave rise to multiple independent mouse lines harbouring XTR alleles (Supplementary Figs 2 and 3). To validate each feature of the XTR system, we established murine embryonic fibroblast (MEF) lines from XTR/XTR, XTR/+ and +/+ littermates for both Rb and p53. MEF lines were sequentially exposed to Cre and then Flp recombinase via adenoviral infection or through the tamoxifen-inducible Cre-ER fusion protein. In both p53 XTR/XTR and Rb XTR/XTR MEFs, Cre efficiently converted the XTR alleles to the TR conformation and Flp subsequently converted the TR alleles to the R conformation (Fig. 1c,d). A key feature of the XTR system is the ability of TR alleles to report host gene expression patterns through GFP expression, while simultaneously eliminating host gene expression. After exposing p53 XTR/XTR or Rb XTR/XTR MEF lines to Cre, these cells expressed GFP (Fig. 1e). As designed, the pattern of GFP expression reports the activity of the respective promoters for each gene: p53 TR/TR cells expressed robust levels of GFP after Cre-mediated recombination, whereas Rb TR/TR cells expressed lower yet detectable levels of GFP. Importantly, the induction of GFP in both p53 TR/TR and Rb _TR/TR_MEFs coincided with a respective loss of p53 and Rb protein expression (Fig. 1f,g). The major goal of the XTR system is to allow a previously inactivated gene to be restored on exposure to Flp recombinase. Infection of p53 TR/TR or Rb TR/TR MEFs with adenoviral FlpO (mammalian codon-optimized Flp) effectively restored p53 and Rb expression to levels indistinguishable from those in p53 XTR/XTR or Rb XTR/XTR MEF lines (Fig. 1f,g).
MEFs from wild-type mice lose proliferative potential after multiple serial passages and enter into a senescent state18. Consistent with their normal regulation of p53, p53 XTR/XTR MEFs ceased to divide after an initial proliferative phase and entered senescence. To determine whether TR alleles phenocopy null alleles, we converted pre-senescent p53 XTR/XTR MEFs to p53 TR/TR MEFs with AdCre. p53 TR/TR MEFs bypassed the proliferative arrest and proliferated indefinitely. Consistent with restoration of p53 in MEFs using other methods, restoration of p53 expression via AdFlpO treatment in immortalized p53 TR/TR MEF cultures completely arrested cell proliferation, demonstrating that sustained inactivation of p53 is required for cellular immortalization of MEFs (Fig. 1h)19. Collectively, these results demonstrate that the XTR allele allows normal p53 regulation, the TR allele is equivalent to null and the R allele restores physiological gene expression.
Rb TR alleles are functionally null
To assess the ability of Rb TR alleles to functionally inactivate Rb, we crossed Meox2-Cre transgenic mice that express Cre in the germline with Rb XTR animals, to generate Rb TR/+ mice20. As Rb is an essential gene, we would not expect live-born Rb TR/TR mice if the TR allele phenocopied a knockout allele21,22. Indeed, out of 51 pups born through these crosses, zero Rb TR/TR pups were generated, whereas both Rb+/+ and Rb TR/+ pups were observed at the expected frequency (Fig. 2a). Analysis of embryonic day 13.5 mice, a time point before the onset of lethality-causing phenotypes, revealed the presence of grossly normal Rb TR/TR embryos that expressed GFP. As anticipated, Rb TR/TR embryos had twofold higher levels of GFP than Rb TR/+ embryos (Fig. 2b). Taken together, these analyses suggest that Rb gene function is lost in Rb TR/TR mice.
Figure 2: Rb TR and p53 TR alleles phenocopy conventional floxed and knockout alleles.
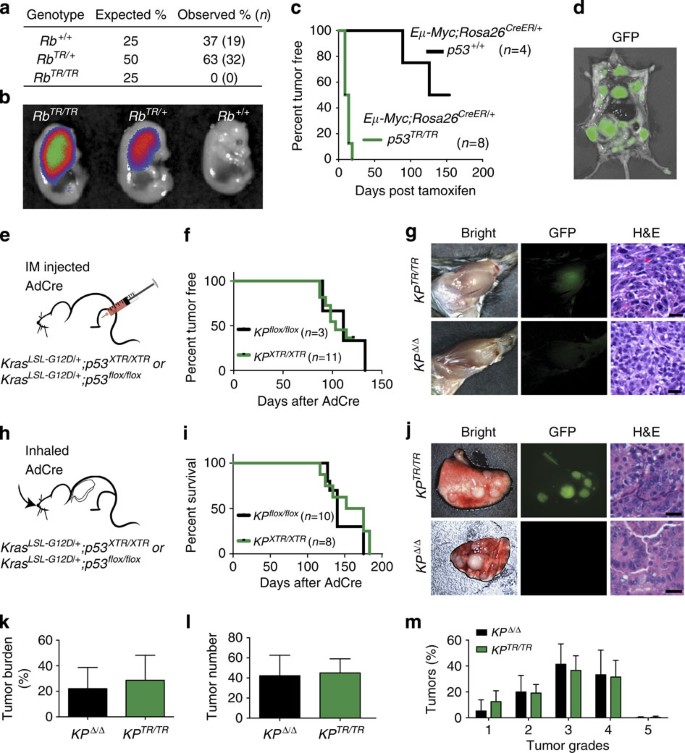
(a) Number of Rb+/+, Rb TR/+ and Rb TR/TR newborn pubs observed in Rb _TR/+_X Rb TR/+ crosses. Percentage and number indicated, χ_2=12.4, df=2, P_=10−4. (b) GFP detection in Rb TR/TR, Rb TR/+ and Rb+/+ embryonic day 13.5 embryos. (c) Kaplan–Meier analysis of lymphoma onset in tamoxifen-treated Eμ-Myc; p53 XTR/XTR ; Rosa26 CreER/+ and Eμ-Myc; p53+/+; Rosa26 _CreER/+_mice; _P_=0.0017, log-rank (Mantel–Cox) test. (d) GFP imaging of lymphoma cells in an Eμ-Myc; p53 XTR/XTR ; Rosa26 CreER/+ mouse after lymphoma onset. (e) Initiation of sarcomas by intramuscular (IM) injection of AdCre into the hindlimb of Kras LSL-G12D/+ ; p53 XTR/XTR(KP XTR/XTR) and Kras LSL-G12D/+ ; p53 flox/flox(KP flox/flox) mice. (f) Kaplan–Meier analysis of sarcoma onset in KP XTR/XTR and KP flox/flox mice. (g) Representative sarcomas from KP XTR/XTR (_n_=11) and KP flox/flox (_n_=3) mice shown by whole-mount bright-field and fluorescent (GFP) microscopy and also (haematoxylin and eosin) staining of histological sections. (h) Initiation of lung adenocarcinoma by inhalation of AdCre in KP XTR/XTR (_n_=8) and KP flox/flox (_n_=10) mice. (i) Kaplan–Meier survival analysis in KP XTR/XTR and KP flox/flox mice after inhalation of AdCre. (j) Representative lungs from KP XTR/XTR and KP flox/flox mice shown by whole-mount bright-field and fluorescent (GFP) microscopy and H&E staining of histological sections. (k,l,m) Comparison of tumour burden (% of lung area), tumour number and tumour grade between KP XTR/XTR and KP _flox/flox_mice. Scale bars, 25 μm.
p53 TR alleles accelerate Myc-driven lymphomagenesis
Loss of p53 expression is causally associated with tumour progression in human cancers and multiple mouse models exist where loss of p53 exacerbates cancer phenotypes. To determine whether XTR alleles could be used to functionally inactivate tumour suppressor genes in diverse tumour models, we crossed the p53 XTR allele to three different well-characterized cancer models. The Eμ-Myc transgenic mouse models Burkitt’s type lymphoma, a B-cell malignancy driven by high levels of Myc expression from the immunoglobulin heavy chain enhancer23. Lymphomagenesis in this model is limited by a p53-dependent apoptotic programme and animals deficient in one or both copies of p53 develop aggressive disease within the first month of life compared with 4–6 months in p53+/+ mice24,25. We exploited the conditional nature of the XTR system, to determine whether somatic recombination of p53 XTR/XTR to p53 TR/TR would inactivate p53 expression and lead to rapid onset of lymphoma in Eμ-Myc transgenic mice. Eμ-Myc; p53 XTR/XTR ;Rosa26 CreER/+ and Eμ-Myc;p53+/+;Rosa26 CreER/+ mice were aged 10–12 weeks and then treated with a single dose of tamoxifen to activate CreER. Conversion of p53 XTR to p53 TR with tamoxifen treatment led to rapid lymphoma formation with all mice developing aggressive B-cell malignancies with a median onset of 11.5 days. In contrast, p53+/+ mice remained tumour free for prolonged periods after tamoxifen treatment (Fig. 2c). Lymphomas that developed in Eμ-Myc;p53 XTR/XTR ;Rosa26 CreER/+ mice treated with tamoxifen expressed GFP, indicating conversion of p53 XTR to p53 TR and activation of the gene trap. GFP signal was easily detected by direct fluorescence microscopy on necropsy, highlighting the potential utility of the XTR system to track and isolate tumour cells (Fig. 2d).
p53 TR alleles phenocopy p53 flox alleles in cancer models
The XTR system was designed to be compatible with cancer models where additional Cre-dependent alleles are required for tumour formation. To assess this potential, we crossed p53 XTR mice to Kras LSL-G12D/+ mice, which harbour a conditional oncogenic Kras G12D allele whose expression is dependent on exposure to Cre26,27. Kras LSL-G12D/+ and traditional p53 flox alleles have been used to model several human tumour types including soft tissue sarcoma and lung adenocarcinoma28,29. In each of these models, Cre-expressing viruses are delivered directly to the site of tumour formation by direct injection into the muscle to generate sarcomas or inhalation to generate lung adenocarcinoma. To determine whether p53 XTR could work in concert with the Kras LSL-G12D/+ allele and phenocopy p53 flox alleles in these tumour models, we initiated tumours in Kras LSL-G12D/+ p53 XTR/XTR(KP XTR/XTR) and Kras LSL-G12D/+ ;p53 flox/flox(KP flox/flox) mice and compared the frequency of tumour initiation and degree of histological progression in models of sarcoma and lung adenocarcinoma (Fig. 2e–m). Intramuscular injection of AdCre initiated sarcomas in KP XTR/XTR and KP _flox/flox_mice with similar penetrance and median time to sarcoma onset (Fig. 2f). KP XTR/XTR and KP flox/flox sarcomas were histologically similar, containing high-grade spindle cell lesions with atypical nuclei and frequent mitotic figures, and consistent with conversion to p53 TR/TR sarcomas arising in KP XTR/XTR mice were GFP positive (Fig. 2g).
Inhalation of AdCre into KP XTR/XTR and KP flox/flox mice resulted in robust lung tumour formation that led to declines in survival with similar rates in each cohort. KP XTR/XTR mice had tumours with bright GFP signal consistent with conversion to p53 TR/TR (Fig. 2j). Tumour number, grade and the extent of tumour burden in KP flox/flox and KP XTR/XTR mice were also indistinguishable (Fig. 2k–m). Taken together, these data demonstrate that XTR is a robust method to inactivate tumour suppressor gene function in diverse tumour types with similar effectiveness to conventional floxed alleles. In addition, the bright GFP signal from the p53 TR/TR allele affords a convenient method to identify, isolate and track tumour cells.
FlpO-ER facilities efficient restoration of p53 TR alleles
Robust methods to temporally regulate Flp activity in vivo are critical to realize the full potential of the XTR system. Based on our success using oestrogen receptor (ER) fusions to Cre recombinase in regulating loxP-directed recombination events in vivo2, we generated KP XTR/XTR ;Rosa26 FlpO-ER/+ mice that ubiquitously express a tamoxifen-inducible FlpO-ER fusion30. KP XTR/XTR ;Rosa26 FlpO-ER/+ MEFs were treated with AdCre to recombine p53 XTR/XTR to p53 TR/TR and monitored by flow cytometry for GFP expression. After 3 days, nearly all of the AdCre-treated cells were GFP positive as expected (Fig. 3a). On replating, we treated cells with 4-hydroxytamoxifen to activate FlpO-ER activity and monitored GFP expression by flow cytometry. Consistent with the long half-life of GFP, at the first passage GFP expression was reduced fivefold in all cells and by the second passage GFP levels had diminished to background levels (Fig. 3a). Consistent with cooperative effects of Kras and p53 mutations to drive transformation, KP TR/TR ;Rosa26 FlpO-ER/+ MEFs rapidly formed tumours after subcutaneous transplantation in nude mice. Importantly, tamoxifen treatment of mice bearing KP TR/TR ;Rosa26 FlpO-ER/+ tumours resulted in rapid loss in growth potential but had no effect on KP flox/flox tumours (Fig. 3b).
Figure 3: Tamoxifen-regulated FlpO-ER efficiently restores p53 TR alleles to p53 R in vitro and in vivo.
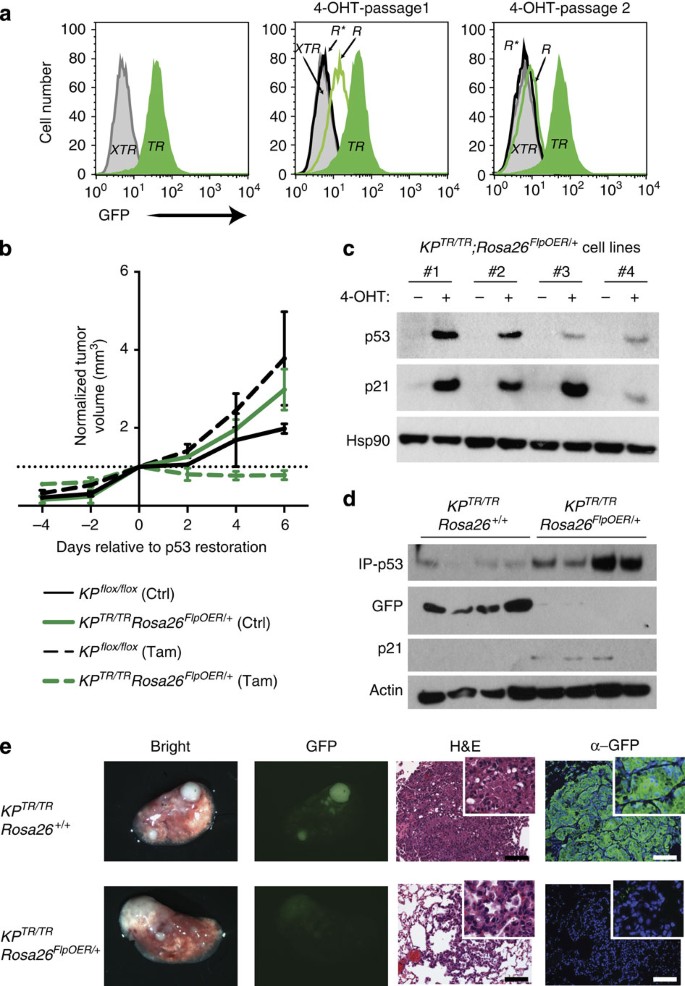
(a) GFP detection by flow cytometry analysis of KP XTR/XTR ; Rosa26 FlpO-ER/+ MEFs. Untreated (p53 XTR/XTR: solid grey), AdCre treated (p53 TR/TR: solid green), tamoxifen-treated cells previously untreated (p53 R*/R*: open black trace) or previously AdCre-treated (p53 R/R: open green trace). R*, direct conversion of p53 XTR to p53 R. (b) Analysis of transformation potential of KP XTR/XTR ; Rosa26 FlpO-ER/+ MEFs after in vitro AdCre treatment followed by subcutaneous engraftment into nude mice. Tumour growth monitored by caliper measurements at the indicated times. Tamoxifen was administered on day 0, 1 and 2 to all mice. Representative of two cell lines (_n_=4). (c) Immunoblot analysis of lysates from KP TR/TR ; Rosa26 FlpO-ER/+ lung tumour-derived cell lines 24 h after addition of 4-hydroxytamoxifen (4-OHT) or vehicle control. (d) Immunoblot analysis of lysates from micro-dissected lung tumours 7 days after tamoxifen delivery. Immune precipitation of p53 was required before western blotting for detection. (e) Microscopic analysis of lung tumours derived from KP XTR/XTR ; Rosa26 FlpO-ER/+ and KP XTR/XTR ; Rosa26+/+ mice 7 days post tamoxifen treatment. From left to right, bright-field and fluorescence stereo microscopy, haematoxylin and eosin staining and immunohistological staining for GFP in representative tumours. Scale bars, 100 μm.
To assess the capabilities of FlpO-ER to regulate tumour suppressor gene restoration in the context of an autochthonous cancer model, we initiated tumours in the lungs of KP XTR/XTR ;Rosa26 FlpO-ER/+ mice by inhalation of AdCre. Initially, we derived cell lines from individual KP TR/TR ;Rosa26 FlpO-ER/+ tumours. In vitro administration of 4-hydroxytamoxifen led to robust induction of p53 and the p53 target gene p21, consistent with FlpO-ER-mediated conversion of p53 TR/TR to p53 R/R(Fig. 3c). Importantly, tamoxifen treatment of KP XTR/XTR ;Rosa26 FlpO-ER/+ mice 13 weeks post tumour initiation also lead to efficient conversion of p53 TR/TR to p53 R/R. Tamoxifen treatment lead to upregulation of p53 and p21, and a concomitant loss of GFP specifically in KP TR/TR ;Rosa26 FlpO-ER/+ but not KP TR/TR ;Rosa26+/+ tumours (Fig. 3d). In addition, direct fluorescence microscopy of harvested lungs or immunohistological analyses of lung tumour sections revealed that KP TR/TR ;Rosa26+/+ control tumours displayed multiple GFP-positive tumours in the lung, suggesting that they maintained p53 TR/TR alleles. In contrast, tamoxifen treatment of KP TR/TR ;Rosa26 FlpO-ER/+ tumour-bearing mice resulted in tumours that had dim or undetectable GFP levels, indicating efficient conversion to p53 R/R alleles (Fig. 3e).
Discussion
Restoration of tumour suppressor gene function in cancer cells in vivo has proven to be a powerful means to identify context-specific programmes of tumour suppression. However, the widespread practice of restoring gene function in established tumours within their natural setting has been greatly limited by previous approaches that are incompatible with specific genes of interest or by strategies that require multiple technically challenging steps to implementation1,2,3,4,5,31.
We and others have used genetically engineered alleles in which a loxP-flanked transcription/translation stop cassette (loxP-STOP-loxP; LSL) is inserted into the first intron of a gene of interest1,2. An LSL allele is a null allele until Cre-mediated recombination deletes the STOP cassette, thus allowing normal expression of the targeted genes. For example, the p53 LSL allele is a functionally null allele of p53 and p53 LSL/LSL mice develop spontaneous lymphomas and sarcomas at the same frequency and rate as p53 KO mice1. Initially, p53 LSL/LSL mice were used to study the consequences of p53 restoration in T-cell lymphomas and soft tissue sarcomas that naturally arise in p53-deficient mice1. Most genetically engineered mouse cancer models rely on Cre to activate or inactivate genes of interest, but because LSL approaches require Cre for gene restoration they are not compatible with these existing models. Because of this major limitation, we made use of a lung cancer model (Kras LA2/+) where Kras G12D is activated spontaneously due to a stochastic recombination event32. These mice develop lung adenocarcinomas with 100% penetrance at an early age. This afforded us the opportunity to generate Kras LA2/+ ;p53 LSL/LSL mice and to restore p53 in these early-to-moderate stage lung cancers2. Performing this restoration in large cohorts of mice was frustrating, owing to the mortality associated with the frequent and rapid development of sarcomas and lymphomas in p53-deficient mice. Extensive ageing of the mice was not possible and we were unable to assess the effects of p53 restoration on the most clinically relevant advanced stages of primary lung tumours and metastases.
The most critical limitation of the LSL system is that LSL alleles cause germline deficiency. Thus, LSL alleles cannot be used to study genes that are required for embryonic development. Unfortunately, the vast majority of tumour suppressor genes are embryonic lethal (for example, Rb1, Pten, Apc, Nf1, Nf2, Ptc, Vhl, Smad4, Atr, Smarca4, Arid1a, Snf5, Nkx2-1, Nkx3-1, Tsc1, Tsc2 and so on), thus leaving very few tumour suppressor genes with which to use this system (for example, p53, Cdkn2a and Atm). This fact, together with its incompatibility with other Cre-based systems, has severely limited the utility of this approach.
Fusion of an ER fragment to proteins may, in some instances, allow for tamoxifen-dependent activity of the fused protein. In the limited cases where this has proven effective, it has been a robust method. A p53 ERTam knock-in allele has been used to model p53 restoration in Myc-induced lymphomas and in Kras _G12D_-induced lung adenocarcinomas3,4. Although, unlike the LSL approach, the ER-fusion alleles are compatible with Cre-based cancer models, ER fusions are still limited to non-essential genes, as mice with homozygous knock-in would still be expected to recapitulate the embryonic lethality of null mice. In addition, not all proteins will tolerate carboxy and/or amino-terminal fusions with the ER and there can be concern of unknown alterations in function of the ER fusion protein. Finally, this approach is limited to proteins that carry out their functions in the nucleus, as the mechanism of induction is based on tamoxifen-induced nuclear translocation of the fusion protein.
Regulatable small hairpin RNA (shRNA) is a very different method that overcomes several of the limitations associated with LSL and ER fusion-based approaches. However, the techniques involved are challenging and, to date, the generation of regulatable RNA interference transgenic mice has been employed by few laboratories. This approach has been used to regulate the expression of three tumour suppressor genes (Apc, Pten and p53) in relevant cancer models5,6,31,33,34. To effectuate potent knockdown of the target gene and recreate phenotypes equivalent to null alleles, multiple specialized techniques, mouse strains and ES cell lines are required35. The success of the regulatable shRNA to recapitulate phenotypes associated with gene loss requires the ability to generate a sufficiently potent shRNA. Screening of dozens of shRNAs is therefore required and specialized protocols to ascertain whether an shRNA is likely to be effective as a single-copy integrated transgene is necessary35,36. Despite this, potent shRNAs have been identified that approximate null alleles in certain experimental systems5,31,37,38.
Off-target effects associated with shRNA expression are also potentially problematic39. High expression of a heterologous shRNA could obscure biological readouts by knocking down the expression of unintended messenger RNA targets or by overwhelming the RNA interference processing machinery to such an extent that naturally expressed micro RNAs are not normally produced40,41,42. Each of these technical issues may have profound consequences on the biology of cells in question that will have an impact on data interpretation.
Currently, regulatable shRNA strategies rely on tet-based systems which limits their functionality within other systems that also utilize tet-regulated transgenes. For example, multiple cancer models rely on tet-inducible oncogenes to drive tumour formation and these models rely on continual expression of the oncogenic driver to maintain the cancer43,44,45,46. Although the regulated shRNA can be easily added into this approach to ascertain the added effect of target gene knockdown, the ability to cleanly determine the effect of target gene restoration is not possible due to the simultaneous loss of oncogene expression on doxycyclin removal.
The simplicity and functionality of the XTR allele system offers several significant advantages over existing strategies, to interrogate gene function in the mouse. Similar to conventional approaches to create conditional alleles in the mouse, XTR integration relies on gene targeting in ES cells. Creation of XTR alleles uses the same methods that are standard protocols in academic and commercial ES cell/transgenic mouse facilities and thus requires no specialized technical hurdles. As outlined in Supplementary Fig. 3, generation of XTR alleles requires a neoXTR allele intermediate that necessitates secondary selection of either ES cell clones or pups that lost the FRT-Neo-FRT. For p53 and Rb, this was achieved by either electroporation with Flp-expressing plasmids or by crossing with germline Flp-expressing mice47. Further use of the XTR system at other loci will be necessary to determine whether locus-dependent effects exist that could limit this strategy. CRISPR-based approaches to generate conditional alleles by directly injecting zygotes with modified XTR vectors lacking the FRT-Neo-FRT cassette could obviate the need for this step48,49.
XTR combines three separate tools into one discrete genetic element to conditionally inactivate a gene of interest, accurately report host gene expression once inactivated and facilitate precise gene restoration in an inducible manner. Bringing these tools together into one strategy offers unparalleled functionality to a single genetically engineered allele. With the increased functionality to mark gene inactivation and report accurate gene expression levels of the targeted gene, as well as the ability to rescue gene function, XTR is positioned to greatly expand current capabilities to interrogate gene function within in vivo systems. Thus far, our experience targeting XTR to p53 and Rb loci suggest a ‘plug and play’ simplicity that abrogates the need for development, testing and optimization that is associated with ER fusion and regulatable shRNA strategies. However, targeting of additional loci will be required to affirm the generalizable nature of the XTR approach with respect to preserving proper host gene regulation and robustness of gene inhibition. Although the XTR system would not be compatible with the few examples of Flp-dependent alleles that exist50,51, its seamless integration into the numerous Cre-based model systems available should facilitate its widespread utility. Similar to most conditional approaches to inactivate gene function in the mouse, XTR alleles require specialized methods or mouse strains to deliver Cre recombinase to specific cell types of interest. Our data suggest that either promoter-specific transgenes or viral based approaches to deliver Cre to tissues of interest is a robust strategy to inactivate genes of interest in the mouse with XTR. Finally, restoration of gene function using XTR alleles requires strategies to regulate Flp activity. As demonstrated, we used a tamoxifen-inducible Rosa26 FlpO-ER allele that is widely expressed and therefore suitable for a broad range of applications using the XTR system30. However, additional strategies to regulate FlpO may augment the utility of the XTR system in specialized scenarios.
Here we have targeted the XTR cassette to two important tumour suppressor genes to address cancer-relevant questions. However, we envision XTR as a powerful approach to investigate gene function in diverse biological settings to gain important insight into mechanisms at the tissue, cellular or molecular level. In addition, XTR alleles have the potential to model therapeutic interventions in disease settings, where temporarily inactivating a putative drug target through the genetic means intrinsic to XTR could predict efficacy or identify unforeseen complications of future therapies. More broadly, the ability to restore gene function using the XTR system offers a major opportunity in conditional genetic methods to facilitate the widespread application of in vivo gene restoration approaches.
Methods
Creation of base targeting vectors
The XTR allele requires targeting to the desired gene loci, to carry out inducible gene trapping via splice acceptance from upstream exons; therefore, pNeoXTR plasmids were created to accept splicing from each of the three reading frames (Supplementary Fig. 1; pNeoXTR f0 (Addgene #69157), pNeoXTR f1 (Addgene #69158) and pNeoXTR f2 (Addgene #69159)). Components were assembled from pL451 (Addgene #22687), pFLIP-FF9 (a gift from Patrick Stern) and pSA-GFP-pA11 (gift from Jan Carette). Alternative reading frames were established by Quickchange method (Agilent) following the manufacturer’s instructions. Full sequence of each targeting plasmid is available at Addgene. Targeting arms were generated by PCR and cloned into base targeting vectors. Amplification of the p53 left arm was accomplished with forward primer 5′- tggcgcgccggatcccagcactactgtggttaag -3′ and reverse primer 5′- ttgcggccgcagaggtttgagtacaaccagggctg -3. Amplification of the p53 right targeting arm was accomplished with forward primer 5′- gtgttaattaacgagtctattgcctttcccagccaac -3′ and reverse primer 5′- ctgcagccaaaggtccagttacagg -3′. Amplification of the Rb left arm was accomplished with forward primer 5′- cacggcgcgccgtcaaacagctatgaccatg -3′ and reverse primer 5′- ccagcgatcgcctcgagaacttaatgatggg -3′. Amplification of the Rb right arm was accomplished with forward primer 5′- gtggttaattaaggctcgag -3′ and reverse primer 5′- gtgttaattaaaaagggatgcaaatagaagg -3′.
ES cell culturing and electroporation
Targeting constructs were linearized and electroporated independently into F1 C57BL/6 × 129S4 hybrid v6.5 ES cells (gift from Rudolph Jaenisch) using standard conditions. Neomycin (300 μg ml−1 G418)-resistant colonies were isolated, expanded and screened by PCR and Southern blotting. Correctly targeted clones were either directly injected into C57BL/6J blastocysts or first electroporated with supercoiled pCAGGS-FLPe (gift from R. Jaenisch) and plated, to generate subclones. ES cell subclones lacking the NeoR cassette after FLP recombination were then injected into C57BL/6J blastocysts. Both strategies yielded several high-percentage chimeras and the establishment of the mouse lines.
Identification of targeted ES clones
PCR was used to screen for probable targeted clones for both p53 and Rb NeoXTR alleles. Reactions spanning the left targeting arm for p53NeoXTR-targeted clones identified positive clones with primers 5′- ttcaagagacggagaaagggcg -3′ and 5′- tggatgtggaatgtgtgcgag -3′. These were subsequently screened via Southern blotting. PCR screening strategies for Rb were not successful and failed to detect even those subsequently identified by Southern blotting. For Southern blotting, genomic DNA was digested overnight at 37 °C with 20 Units of each restriction enzyme. The next day, another 10 Units were added and incubated at 37 °C for an additional 4–6 h. DNA fragments were resolved on 0.7% tris-acetate-EDTA gels at low voltage overnight, stained with ethidium bromide, depurinated, denatured and neutralized before transfer to Hybond XL membranes. Ultraviolet cross-linked DNA was hybridized with radioactive probes suspended in Express Hyb solution (Clontech) as per the manufacturer’s instructions. Radiolabelled probes were generated by random primer methods using PrimeIT II Kits (Agilent) and purified with Quick Spin Columns (Roche). DNA probes were generated by PCR of mouse genomic DNA. Rb 5′-probe was amplified with 5′- attaagttctcgattcctcag -3′ and 5′- tgccaggcggacccgactttg -3′ primers. The p53 3′-probe was amplified with 5′- atagtgggaaccttctgggacg -3′ and 5′- cagtggaggagcacctgtcttatg -3′ primers. The GFP probe was generated using pSA-GFP-pA as a template with 5′- atggtgagcaagggcgagga -3′ and 5′- ttacttgtacagctcgtcca -3′ primers. Genomic DNA from ES cell clones following Flpe electroporation was screened by PCR with the following primers to identify a 528-bp product (Rb XTR) or a 587-bp product (p53 XTR): p53 5′- caactgttctacctcaagagcc -3′ × 5′- taaaaaacctcccacacctcccc -3′ and Rb 5′- tgttgttattgtcagcactag -3′ × 5′- TAAAAAACCTCCCACACCTCCCC -3′.
Germline deletion of Neo R
p53 neoXTR/+ and Rb neoXTR/+ mice were crossed to Rosa26 FlpeR/+ mice. p53 neoXTR/+ ; Rosa26 FlpeR/+ and Rb neoXTR/+ ; Rosa26 FlpeR/+ pups were found to contain all possible alleles after Flp reactions (neoXTR, XTR and R), and were therefore always considered to be mosaic. p53 neoXTR/+ ; Rosa26 FlpeR/+ and Rb neoXTR/+ ; Rosa26 FlpeR/+ mice were subsequently crossed to wild-type mice and pups that lost the Rosa26 FlpeR allele were screened for retention of XTR and loss of Neo R. See also Supplementary Fig. 3.
Generation and analysis of MEFs
MEFs were generated by timed matings from E13.5 to E15.5 embryos. Adenoviral infections to express Cre or FlpO recombinases (purchased from Gene Transfer Vector Core at the University of Iowa) were carried out on sub-confluent cultures using 107–108 viral particles in 6- or 10-cm dishes. Quantification of GFP in MEFs was performed on an LSR II cytometer (BD Biosciences) at the PennFlow Core. Proliferation assays were carried out in 3T3 format counting and replating 3 × 105 cells every 3 days in 6-cm dishes. Genotypes of MEFs were analysed via three-primer PCR reactions using purified genomic DNA as a template to detect XTR, TR, R and WT alleles. Primers used for Trp53 were as follows: (1) 5′- cttggagacatagccacactg -3′, (2) 5′- caactgttctacctcaagagcc -3′ and (3) 5′- cttgaagaagatggtgcg -3′. Primers used for Rb1 were as follows: (1) 5′- tgttgttattgtcagcactag -3′, (2) 5′- ggcagaggcagtaaaacagagagc -3′ and (3) 5′- cttgaagaagatggtgcg -3′.
Immunodetection in cultured cells
Cells were lysed in RIPA buffer and resolved on NuPage BT gels and then transferred to polyvinylidene difluoride (Millipore). Blottings were probed with the following antibodies: p53 (Novocastra IMX25, western, 1:500) or Santa Cruz (FL-393, IP 1 μg), Rb (Santa Cruz, C-15, 1:500), p21 (Santa Cruz, F-5, 1:500), GFP (Cell Signaling Technologies, #2956, 1:1,000), Hsp90 (BD Transduction Labs, Clone 68, 1:1,000), β-Tubulin (Cell Signaling Technologies, #2146 1:5,000), Actin (Sigma, A2066, 1:1,000). Immunoprecipitation was carried out in RIPA buffer with standard techniques using Protein G beads. 4-Hydroxytamoxifen dissolved in ethanol was administered once at the time of cell plating at a final concentration of 500 nM. AdCre or AdFlpO (5 × 107 pfu) was purchased from the University of Iowa Gene Vector Core and administered to adherent cells at the indicated times. Full versions of blots are shown in Supplementary Fig. 4.
Tumour-derived cell lines
Tumours were excised from the lungs of mice using dissecting stereo microscopy, dissociated with Collagenase IV and trypsin for 30–60 min, then quenched with fetal bovine serum, washed in Hank’s balanced salt solution, then cultured in high glucose-containing DMEM supplemented with 10% fetal bovine serum and antibiotics at 37 °C and 5% CO2 until cell line establishment (∼2–4 weeks).
Animal studies and treatments
All animal experiments were carried out under strict compliance with Institutional Animal Care and Use Committee at either MIT or Penn. for Kras LSL-G12D (ref. 26), p53 flox/flox (ref. 52), Rosa26 FlpeR (ref. 47), Rosa26 FlpO-ER (ref. 30) and Meox2-Cre20 mice have been described. Sarcoma and lung adenocarcinoma inductions were carried out as described29,53. Tumour grading was performed at the PennVet Comparative Pathology Core using established tumour-grading schemes28,29. Tamoxifen treatment was performed on 3 consecutive days via oral gavage of 200 μl of a 20 mg ml−1 solution dissolved in 90% sterile corn oil and 10% ethanol.
Histology and microscopy
Fluorescent signals from tumours were imaged with either the IVIS Spectrum (Caliper Life Sciences) or a fluorescence stereo dissection microscope (Leica). Tissues were dissected into 10% Neutral buffered formalin and fixed 16–20 h at 4 °C before being dehydrated in a graded alcohol series. Paraffin-embedded histological sections were produced at the Abramson Family Cancer Research Institute Histopathology Core as 4 μm sections before staining. Immunostaining was carried out after citrate-based antigen retrieval with α-GFP antibody (Abcam ab13970, 1:1,000) and fluorescent secondary detection (Thermo Fisher, α-Chicken Alexa Fluor 594, 1:200). Microscopy was performed on a Leica DMI6000B inverted light and fluorescent microscope.
Additional information
How to cite this article: Robles-Oteiza, C. et al. Recombinase-based conditional and reversible gene regulation via XTR alleles. Nat. Commun. 6:8783 doi: 10.1038/ncomms9783 (2015).
References
- Ventura, A. et al. Restoration of p53 function leads to tumour regression in vivo. Nature 445, 661–665 (2007).
Article CAS PubMed Google Scholar - Feldser, D. M. et al. Stage-specific sensitivity to p53 restoration during lung cancer progression. Nature 468, 572–575 (2010).
Article CAS ADS PubMed PubMed Central Google Scholar - Martins, C. P., Brown-Swigart, L. & Evan, G. I. Modeling the therapeutic efficacy of p53 restoration in tumors. Cell 127, 1323–1334 (2006).
Article CAS PubMed Google Scholar - Junttila, M. R. et al. Selective activation of p53-mediated tumour suppression in high-grade tumours. Nature 468, 567–571 (2010).
Article CAS ADS PubMed PubMed Central Google Scholar - Xue, W. et al. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature 445, 656–660 (2007).
Article CAS PubMed PubMed Central Google Scholar - Miething, C. et al. PTEN action in leukaemia dictated by the tissue microenvironment. Nature 510, 402–406 (2014).
Article CAS ADS PubMed PubMed Central Google Scholar - Schnutgen, F. et al. A directional strategy for monitoring Cre-mediated recombination at the cellular level in the mouse. Nat. Biotechnol. 21, 562–565 (2003).
Article PubMed Google Scholar - Schnutgen, F. et al. Genomewide production of multipurpose alleles for the functional analysis of the mouse genome. Proc. Natl Acad. Sci. USA 102, 7221–7226 (2005).
Article ADS PubMed PubMed Central Google Scholar - Stern, P. et al. A system for Cre-regulated RNA interference in vivo. Proc. Natl Acad. Sci. USA 105, 13895–13900 (2008).
Article CAS ADS PubMed PubMed Central Google Scholar - Economides, A. N. et al. Conditionals by inversion provide a universal method for the generation of conditional alleles. Proc. Natl Acad. Sci. USA 110, E3179–E3188 (2013).
Article CAS PubMed PubMed Central Google Scholar - Carette, J. E. et al. Haploid genetic screens in human cells identify host factors used by pathogens. Science 326, 1231–1235 (2009).
Article CAS ADS PubMed Google Scholar - Carette, J. E. et al. Ebola virus entry requires the cholesterol transporter Niemann-Pick C1. Nature 477, 340–343 (2011).
Article CAS ADS PubMed PubMed Central Google Scholar - Lee, C. C., Carette, J. E., Brummelkamp, T. R. & Ploegh, H. L. A reporter screen in a human haploid cell line identifies CYLD as a constitutive inhibitor of NF-kappaB. PLoS ONE 8, e70339 doi:10.1371/journal.pone.0070339.
Article CAS ADS PubMed PubMed Central Google Scholar - Jae, L. T. et al. Deciphering the glycosylome of dystroglycanopathies using haploid screens for lassa virus entry. Science 340, 479–483 (2013).
Article CAS ADS PubMed PubMed Central Google Scholar - Reiling, J. H. et al. A haploid genetic screen identifies the major facilitator domain containing 2A (MFSD2A) transporter as a key mediator in the response to tunicamycin. Proc. Natl Acad. Sci. USA 108, 11756–11765 (2011).
Article CAS ADS PubMed PubMed Central Google Scholar - Reiling, J. H. et al. A CREB3-ARF4 signalling pathway mediates the response to Golgi stress and susceptibility to pathogens. Nat. Cell Biol. 15, 1473–1485 (2013).
Article CAS PubMed PubMed Central Google Scholar - Chiou, S. H. et al. A conditional system to specifically link disruption of protein-coding function with reporter expression in mice. Cell Rep. 7, 2078–2086 (2014).
Article CAS PubMed PubMed Central Google Scholar - Sherr, C. J. & DePinho, R. A. Cellular senescence: mitotic clock or culture shock? Cell 102, 407–410 (2000).
Article CAS PubMed Google Scholar - Christophorou, M. A. et al. Temporal dissection of p53 function in vitro and in vivo. Nat. Genet. 37, 718–726 (2005).
Article CAS PubMed Google Scholar - Tallquist, M. D. & Soriano, P. Epiblast-restricted Cre expression in MORE mice: a tool to distinguish embryonic vs. extra-embryonic gene function. Genesis 26, 113–115 (2000).
Article CAS PubMed Google Scholar - Jacks, T. et al. Effects of an Rb mutation in the mouse. Nature 359, 295–300 (1992).
Article CAS ADS PubMed Google Scholar - Lee, E. Y. et al. Mice deficient for Rb are nonviable and show defects in neurogenesis and haematopoiesis. Nature 359, 288–294 (1992).
Article CAS ADS PubMed Google Scholar - Adams, J. M. & Cory, S. Myc oncogene activation in B and T lymphoid tumours. Proc. R. Soc. Lond. B. Biol. Sci. 226, 59–72 (1985).
Article CAS ADS PubMed Google Scholar - Schmitt, C. A., McCurrach, M. E., de Stanchina, E., Wallace-Brodeur, R. R. & Lowe, S. W. INK4a/ARF mutations accelerate lymphomagenesis and promote chemoresistance by disabling p53. Genes Dev. 13, 2670–2677 (1999).
Article CAS PubMed PubMed Central Google Scholar - Schmitt, C. A. et al. Dissecting p53 tumor suppressor functions in vivo. Cancer Cell 1, 289–298 (2002).
Article CAS PubMed Google Scholar - Jackson, E. L. et al. Analysis of lung tumor initiation and progression using conditional expression of oncogenic K-ras. Genes Dev. 15, 3243–3248 (2001).
Article CAS PubMed PubMed Central Google Scholar - Tuveson, D. A. et al. Endogenous oncogenic K-ras(G12D) stimulates proliferation and widespread neoplastic and developmental defects. Cancer Cell 5, 375–387 (2004).
Article CAS PubMed Google Scholar - Jackson, E. L. et al. The differential effects of mutant p53 alleles on advanced murine lung cancer. Cancer Res. 65, 10280–10288 (2005).
Article CAS PubMed Google Scholar - Kirsch, D. G. et al. A spatially and temporally restricted mouse model of soft tissue sarcoma. Nat. Med. 13, 992–997 (2007).
Article CAS PubMed Google Scholar - Lao, Z., Raju, G. P., Bai, C. B. & Joyner, A. L. MASTR: a technique for mosaic mutant analysis with spatial and temporal control of recombination using conditional floxed alleles in mice. Cell Rep. 2, 386–396 (2012).
Article CAS PubMed PubMed Central Google Scholar - Premsrirut, P. K. et al. A rapid and scalable system for studying gene function in mice using conditional RNA interference. Cell 145, 145–158 (2011).
Article CAS PubMed PubMed Central Google Scholar - Johnson, L. et al. Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature 410, 1111–1116 (2001).
Article CAS ADS PubMed Google Scholar - Dickins, R. A. et al. Tissue-specific and reversible RNA interference in transgenic mice. Nat. Genet. 39, 914–921 (2007).
Article CAS PubMed PubMed Central Google Scholar - Dow, L. E. et al. Apc restoration promotes cellular differentiation and reestablishes crypt homeostasis in colorectal cancer. Cell 161, 1539–1552 (2015).
Article CAS PubMed PubMed Central Google Scholar - Dow, L. E. et al. A pipeline for the generation of shRNA transgenic mice. Nat. Protoc. 7, 374–393 (2012).
Article CAS PubMed PubMed Central Google Scholar - Fellmann, C. et al. Functional identification of optimized RNAi triggers using a massively parallel sensor assay. Mol. Cell 41, 733–746 (2011).
Article CAS PubMed PubMed Central Google Scholar - Dickins, R. A. et al. Probing tumor phenotypes using stable and regulated synthetic microRNA precursors. Nat. Genet. 37, 1289–1295 (2005).
Article CAS PubMed Google Scholar - Hemann, M. T. et al. An epi-allelic series of p53 hypomorphs created by stable RNAi produces distinct tumor phenotypes in vivo. Nat. Genet. 33, 396–400 (2003).
Article CAS PubMed Google Scholar - Kaelin, W. G. Jr. Molecular biology. Use and abuse of RNAi to study mammalian gene function. Science 337, 421–422 (2012).
Article ADS PubMed PubMed Central Google Scholar - Jackson, A. L. et al. Widespread siRNA “off-target” transcript silencing mediated by seed region sequence complementarity. RNA 12, 1179–1187 (2006).
Article CAS PubMed PubMed Central Google Scholar - Echeverri, C. J. et al. Minimizing the risk of reporting false positives in large-scale RNAi screens. Nat. Methods 3, 777–779 (2006).
Article CAS PubMed Google Scholar - Grimm, D. et al. Fatality in mice due to oversaturation of cellular microRNA/short hairpin RNA pathways. Nature 441, 537–541 (2006).
Article CAS ADS PubMed Google Scholar - Fisher, G. H. et al. Induction and apoptotic regression of lung adenocarcinomas by regulation of a K-Ras transgene in the presence and absence of tumor suppressor genes. Genes Dev. 15, 3249–3262 (2001).
Article CAS PubMed PubMed Central Google Scholar - Politi, K. et al. Lung adenocarcinomas induced in mice by mutant EGF receptors found in human lung cancers respond to a tyrosine kinase inhibitor or to down-regulation of the receptors. Genes Dev. 20, 1496–1510 (2006).
Article CAS PubMed PubMed Central Google Scholar - Ji, H. et al. The impact of human EGFR kinase domain mutations on lung tumorigenesis and in vivo sensitivity to EGFR-targeted therapies. Cancer Cell 9, 485–495 (2006).
Article CAS PubMed Google Scholar - Moody, S. E. et al. Conditional activation of Neu in the mammary epithelium of transgenic mice results in reversible pulmonary metastasis. Cancer Cell 2, 451–461 (2002).
Article CAS PubMed Google Scholar - Farley, F. W., Soriano, P., Steffen, L. S. & Dymecki, S. M. Widespread recombinase expression using FLPeR (flipper) mice. Genesis 28, 106–110 (2000).
Article CAS PubMed Google Scholar - Yang, H. et al. One-step generation of mice carrying reporter and conditional alleles by CRISPR/Cas-mediated genome engineering. Cell 154, 1370–1379 (2013).
Article CAS PubMed PubMed Central Google Scholar - Yang, H., Wang, H. & Jaenisch, R. Generating genetically modified mice using CRISPR/Cas-mediated genome engineering. Nat. Protoc. 9, 1956–1968 (2014).
Article CAS PubMed Google Scholar - Young, N. P. & Jacks, T. Tissue-specific p19Arf regulation dictates the response to oncogenic K-ras. Proc. Natl Acad. Sci. USA 107, 10184–10189 (2010).
Article CAS ADS PubMed PubMed Central Google Scholar - Lee, C. L. et al. Generation of primary tumors with Flp recombinase in FRT-flanked p53 mice. Dis. Model Mech. 5, 397–402 (2012).
Article CAS PubMed Google Scholar - Jonkers, J. et al. Synergistic tumor suppressor activity of BRCA2 and p53 in a conditional mouse model for breast cancer. Nat. Genet. 29, 418–425 (2001).
Article CAS PubMed Google Scholar - DuPage, M., Dooley, A. L. & Jacks, T. Conditional mouse lung cancer models using adenoviral or lentiviral delivery of Cre recombinase. Nat. Protoc. 4, 1064–1072 (2009).
Article CAS PubMed PubMed Central Google Scholar
Acknowledgements
We thank Patrick Stern and Jan Carette for plasmid vectors, and Peter Sandy for insights critical for construction of the XTR system. We are grateful to members of the Swanson Biotechnology Center at MIT, Noranne Enzer for help with ES cell work and Scott Malstrom for help with IVIS imaging. We thank Qian-Chun Yu and Jianping Wang in the Abramson Family Cancer Research Institute Histology Core for help with sample preparations, members of the Flow Cytometry and Cell Sorting Facility at Penn, Elizabeth Buza and Amy Durham in the PennVet Comparative Pathology Core for assistance with tumour histology, Lindsay Lawless and Caroline Kitzmiller for administrative support, and Heidi Feldser for critical reading of the manuscript. This work was funded by grants from the National Cancer Institute; R00-CA158581 (D.M.F.), P30-CA016520 (Penn Abramson Cancer Center), P30-CA14051 (MIT Cancer Center) and R01-CA175336 (M.M.W.). T.J. is supported by the Howard Hughes Medical Institute. M.C. is supported by the AACR-Bayer HealthCare Basic Cancer Research Fellowship. B.L. is supported by the Cell and Molecular Biology Training Grant at the University of Pennsylvania, (T32-GM07229).
Author information
Authors and Affiliations
- Department of Cancer Biology, Abramson Family Cancer Research Institute and Perelman School of Medicine at the University of Pennsylvania, 421 Curie Boulevard, 751 Biomedical Research Building II/III, Philadelphia, 19004, Pennsylvania, USA
Camila Robles-Oteiza, Travis Yates, Michelle Cicchini, Brian Lauderback & David M. Feldser - Koch Institute for Integrative Cancer Research at the Massachusetts Institute of Technology, Cambridge, 02144, Massachusetts, USA
Sarah Taylor, Christopher R. Cashman, Aurora A. Burds & Tyler Jacks - Department of Genetics, Department of Pathology, Stanford University School of Medicine, Stanford, 94305, California, USA
Monte M. Winslow - The Howard Hughes Medical Institute, Chevy Chase, 20815, Maryland, USA
Tyler Jacks
Authors
- Camila Robles-Oteiza
You can also search for this author inPubMed Google Scholar - Sarah Taylor
You can also search for this author inPubMed Google Scholar - Travis Yates
You can also search for this author inPubMed Google Scholar - Michelle Cicchini
You can also search for this author inPubMed Google Scholar - Brian Lauderback
You can also search for this author inPubMed Google Scholar - Christopher R. Cashman
You can also search for this author inPubMed Google Scholar - Aurora A. Burds
You can also search for this author inPubMed Google Scholar - Monte M. Winslow
You can also search for this author inPubMed Google Scholar - Tyler Jacks
You can also search for this author inPubMed Google Scholar - David M. Feldser
You can also search for this author inPubMed Google Scholar
Contributions
D.M.F. conceived the project and designed the XTR system. D.M.F. and C.R.C. cloned XTR targeting vectors. A.A.B. performed ES cell work. D.M.F. and S.E.T. established mouse lines. C.R.O., S.E.T., T.Y., M.C., B.L. and D.M.F. performed experiments. M.M.W. and T.J. provided conceptual advice. C.R.O. and D.M.F. wrote the manuscript with comments from all authors.
Corresponding author
Correspondence toDavid M. Feldser.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article
Cite this article
Robles-Oteiza, C., Taylor, S., Yates, T. et al. Recombinase-based conditional and reversible gene regulation via XTR alleles.Nat Commun 6, 8783 (2015). https://doi.org/10.1038/ncomms9783
- Received: 04 June 2015
- Accepted: 30 September 2015
- Published: 05 November 2015
- DOI: https://doi.org/10.1038/ncomms9783