Understanding the Contribution of Zinc Transporters in the Function of the Early Secretory Pathway (original) (raw)
Author / Affiliation / Email
![]()
Article Menu
/ajax/scifeed/subscribe
Font Type:
Arial Georgia Verdana
Open AccessReview
by
Taiho Kambe
* ,
,
Mayu Matsunaga
and
Taka-aki Takeda
Division of Integrated Life Science, Graduate School of Biostudies, Kyoto University, Kyoto 606-8502, Japan
*
Author to whom correspondence should be addressed.
Submission received: 8 September 2017 /Revised: 12 October 2017 /Accepted: 15 October 2017 /Published: 19 October 2017
Abstract
:
More than one-third of newly synthesized proteins are targeted to the early secretory pathway, which is comprised of the endoplasmic reticulum (ER), Golgi apparatus, and other intermediate compartments. The early secretory pathway plays a key role in controlling the folding, assembly, maturation, modification, trafficking, and degradation of such proteins. A considerable proportion of the secretome requires zinc as an essential factor for its structural and catalytic functions, and recent findings reveal that zinc plays a pivotal role in the function of the early secretory pathway. Hence, a disruption of zinc homeostasis and metabolism involving the early secretory pathway will lead to pathway dysregulation, resulting in various defects, including an exacerbation of homeostatic ER stress. The accumulated evidence indicates that specific members of the family of Zn transporters (ZNTs) and Zrt- and Irt-like proteins (ZIPs), which operate in the early secretory pathway, play indispensable roles in maintaining zinc homeostasis by regulating the influx and efflux of zinc. In this review, the biological functions of these transporters are discussed, focusing on recent aspects of their roles. In particular, we discuss in depth how specific ZNT transporters are employed in the activation of zinc-requiring ectoenzymes. The means by which early secretory pathway functions are controlled by zinc, mediated by specific ZNT and ZIP transporters, are also subjects of this review.
Graphical Abstract
1. Introduction
Zinc is an essential trace element that is required for a large variety of cellular processes [1,2]. Approximately 10% of the eukaryotic proteome requires zinc for cellular activity [3,4], and thus any disturbance in zinc homeostasis can result in disease, including cancer, neuronal degeneration, chronic inflammation, hypertension, osteoarthritis, and age-related macular degeneration. A diverse range of symptoms is also found in cases of zinc deficiency [1,2,5,6,7,8]. The biological functions of zinc can be grouped into three major categories, structural, catalytic, and regulatory. However, the molecular basis of how zinc engages in such diverse functions is still far from being completely understood [1,2].
At the cellular level, zinc plays a pivotal role in the function of a variety of subcellular compartments, one of which is the early secretory pathway constituted by the endoplasmic reticulum (ER), the Golgi apparatus, and other intermediate organelles, such as the ER-Golgi intermediate compartment. Zinc homeostasis in the lumen of these compartments requires a transport system to translocate zinc across biological membranes. In vertebrates, Zn transporters (ZNTs)/Solute carrier family 30 member (SLC30A) and Zrt- and Irt-like proteins (ZIPs)/SLC39A are widely recognized as being critical transporters in zinc metabolism under physiological conditions [1,2,9]. Both of these proteins are clearly important for zinc metabolism involved in early secretory pathways. This review outlines the functions of ZNT and ZIP transporters in the regulation and function of secretory pathways, in particular, the early secretory pathway, focusing on several recent aspects of the molecular processes underlying the ER stress response, as well as the activation of zinc-requiring ectoenzymes. Zinc transporters also play important roles in secretory granules/vesicles that contain high amounts of zinc, such as insulin granules, synaptic vesicles, and secretory vesicles involved in milk secretion; these are discussed in this review for comparison. Further details of these transporters can be found in other comprehensive reviews of zinc transporters [1,10,11,12].
2. Brief Overview of the Properties of ZNT and ZIP Transporters
In mammals, there are nine ZNT and 14 ZIP transporters that play distinct roles in the maintenance of systemic, cellular, and subcellular zinc homeostasis. These transporters act in a cell or tissue-specific manner, and are developmentally regulated [1,2,9] (Figure 1). ZNTs transport zinc from the cytosol into either the lumen of intracellular compartments or the extracellular milieu, whereas ZIPs transport zinc in the opposite direction. Zinc transport by ZNT and ZIP transporters is coordinately controlled through precisely timed increases or decreases in their expression, and by their precise subcellular localization [2,13]. A growing body of evidence has shown that cooperative zinc transport across biological membranes mediated by both transporters contributes to the control of expression, localization, and functional activity of target proteins [1]. The molecular features of ZNT and ZIP transporters have been extensively summarized in other review papers [1,2], and thus only their main features are outlined briefly here.
Based on the three-dimensional structure of the Escherichia coli homolog YiiP, ZNT transporters are predicted to have six transmembrane (TM) helices (TM helices I-VI) [14,15,16,17,18]. ZNT transporters function as zinc/proton exchangers [19,20], and can form homodimers or heterodimers [21,22,23,24]. With respect to the zinc transport mechanism used by YiiP, two models have been proposed, the alternative access mechanism model, in which the TM helices form inward- and outward-facing conformations [17,18], and the allosteric mechanism model, in which cytosolic zinc binding induces a scissor-like movement of the homodimers and interlocks the TM helices at the dimer interface [15,16]. ZNT transporters likely transport zinc using either of the two proposed mechanisms. As has been observed for YiiP, which has an intramembranous zinc-binding site formed by TM helices II and V, ZNT transporters are also thought to have a conserved intramembranous zinc-binding site, which is indispensable for zinc transport activity [19,20,25,26]. The intramembranous zinc-binding site in most ZNT transporters consists of two His and two Asp residues in TM helices II and V [19,27]. Interestingly, ZNT10 has an Asn residue in TM helix II instead of His, which confers the ability to transport manganese [28], as has also been seen for the homologous bacterial protein [29]. The nine ZNT transporters belong to the cation diffusion facilitator (CDF) family of transporters, which are classified into three subgroups, namely Zn-CDF, Zn/Fe-CDF, and Mn-CDF [14,30]. All of the ZNT transporters are classified as being Zn-CDF members (although ZNT10 is a manganese transporter), and, based on their sequence similarities, can be further subdivided into four groups: (i) ZNT1 and ZNT10, (ii) ZNT2, ZNT3, ZNT4, and ZNT8, (iii) ZNT5 and ZNT7, and (iv) ZNT6, [14,31,32] (Figure 2). Of interest to this review, some characteristics of the transporters, such as subcellular localization, are conserved in the members of the same group [32] (Figure 1).
Computational analysis suggested that ZIP transporters have eight TM helices [33], and this was confirmed by the first three-dimensional structure reported for a ZIP transporter homolog in bacteria (Bordetella bronchiseptica) [34]. The structure shows that the ZIP transporter has a novel 3 + 2 + 3 TM architecture with a binuclear metal center, in which two His residues, one each in TM helices IV and V, form two intramembranous zinc binding sites [34]. As in ZNT transporters, ZIP transporters form functional homo- or heterodimeric complexes, which are essential for their zinc transport ability, although no dimer formation was seen in the crystal structure [34,35,36]. ZIP transporters may function as selective electrodiffusion channels [37], or as zinc/bicarbonate symport transporters [38,39,40]. However, their definitive mode of transport has not yet been completely elucidated. Phylogenetic analysis classifies the fourteen ZIP transporter members into four subfamilies, namely ZIP I (ZIP9), ZIP II (ZIP1-ZIP3), LIV-1 (ZIP4-ZIP8, ZIP10, ZIP12-ZIP14), and gufA (ZIP11) [1,41]. In the LIV-1 subfamily, features of their extracellular domains further classify the proteins into four subgroups as follows: (i) ZIP4 and ZIP12, (ii) ZIP8 and ZIP14, (iii) ZIP5, ZIP6, and ZIP10, and (iv) ZIP7 and ZIP13 [42] (Figure 2). Members of the LIV-1 subfamily have an extended extracellular N-terminus, whose structure has been solved only in the case of ZIP4 [42]. The extracellular portion of ZIP4 can form homodimers without the need for TM helices [42], which may facilitate dimerization. Interestingly, in ZIP transporters belonging to subgroup (iii), a prion-like domain is present in the extracellular N-terminal portion proximal to the first TM helix; hence, there is an evolutionary link between these transporters and the prion protein [43]. ZIP8 and ZIP14 in subgroup (ii) have the ability to transport manganese [39,40], because they have a Glu residue in TM helix V rather than a His residue [1,44]. The His residue is therefore involved in metal substrate specificity, because of its contribution to forming intramembranous zinc binding sites [34].
Over the last two decades, the physiological roles of ZNT and ZIP transporters and their involvement in disease pathology have been clarified at the molecular level, as has been described elsewhere [1,2,8,9], and a deeper understanding will likely come in the future.
3. Regulation of Zinc Homeostasis by Zinc Transporters in the Early Secretory Pathway
Approximately one-third of all the cellular proteins in eukaryotes are targeted to the ER, and thus the early secretory pathway [45], in which nascent proteins are folded, assembled, and modified during their trafficking to final destinations. Importantly, a considerable proportion of the secretome requires zinc as a structural and catalytic cofactor. Moreover, resident chaperones require zinc for modulation and potentiation of their functions [46,47,48]. Hence, any disruption of zinc homeostasis in the early secretory pathway can cause and exacerbate ER stress [49,50], and trigger the unfolded protein response (UPR) in cells. Therefore, elaborate regulatory mechanisms are used to control zinc homeostasis in the early secretory pathway. Accumulating evidence clearly shows that both ZNT and ZIP transporters play crucial roles in this process [49,50,51,52,53], and this is summarized in this section.
With the exception of ZNT1 and ZNT10 (which are members of ZNT subgroup (i) described above [28,54]), ZNT transporters are mainly localized to intracellular compartments. Of these, ZNT5, ZNT6, and ZNT7 (members of ZNT subgroups (iii), and (iv) described above) are involved in the early secretory pathway [12,55]. ZNT5 has been shown to be mainly localized to coat protein complex II (COPII) vesicles and the Golgi apparatus [56], whereas ZNT6 is localized to the Golgi apparatus [57], although these ZNTs can also form heterodimers as functional complexes [21,22] (Figure 1). The actual subcellular localization of these heterodimers has however been poorly investigated. ZNT7 is also located in the Golgi apparatus [58], and a recent study indicates that it is also localized to the sarco(endo)plasmic reticulum (S(E)R) [52] (Figure 1). These ZNT transporters are employed as zinc entry routes in the early secretory pathway, suggesting that a lack of them would be potentially to elicit an ER stress response. In fact, it has been clearly shown that cells lacking these ZNT transporters do exhibit an exacerbated ER stress responses [50,59] (Figure 3). A similar exacerbation of ER stress is found in yeast lacking ZNT orthologs [49,60], which highlights the fact that the important role these ZNT transporters play in maintaining zinc homeostasis in the secretory pathway is well-conserved among subgroups. However, there remains the interesting questions of how and where these ZNT transporters transport zinc, and their association with ER stress, because ZNT5, ZNT6, and ZNT7 all appear to be principally localized to the Golgi apparatus [57,58,61]. Recent studies have shown that another ZNT transporter, either ZNT3 or ZNT10, may play a protective role in ER stress-induced toxicities [62,63], although their contributions to the early secretory pathway have not yet been clarified.
In contrast to ZNT transporters, most of which are located in intracellular compartments, most, but not all, ZIP transporters are found on the plasma membrane. Of the 14 ZIP transporters, ZIP7, ZIP9, and ZIP13 (the ZIPI subfamily and the LIV-1 subgroup (iii) described above) are involved in the early secretory pathway [64,65,66,67] (Figure 1), although recent reports also indicate that ZIP9 is also found localized on the plasma membrane where it serves as a membrane androgen receptor [68] and that ZIP13 can also be found in intracellular vesicles [69]. It has also been suggested that ZIP11 can localize to the Golgi apparatus [70], but this has not yet been thoroughly established. Accordingly, this protein is not further discussed here. ZIP7, ZIP9, and ZIP13 are known to release zinc, which is stored in the early secretory pathway, into the cytosol in response to various stimuli, thus contributing to the signaling function of zinc [66,71,72,73]. Importantly, ZIP7, ZIP9, and ZIP13 are also thought to contribute to homeostatic maintenance of the secretory pathway. In this regard, ZIP7, which is located in the ER, plays an indispensable role in the proper regulation of ER function, through the fine-tuning of zinc homeostasis [51,53]. The loss of ZIP7 probably increases zinc levels in the ER, which triggers zinc-dependent aggregation of protein disulfide isomerase, leading to ER stress [53] (Figure 3). This critical functional role of ZIP7 in ER homeostasis contributes to self-renewal processes of intestinal epithelium [51] and appropriate epidermal development [53]. ZIP7 is also involved in the induction of ER stress by mediating the redistribution of zinc into the cytosol from the S(E)R in cardiomyocytes under hyperglycemic conditions [52]. Consistent with the involvement of ZIP7 in ER stress, the yeast ZIP7 homolog, yKE4, has been shown to be involved in ER stress responses [74]. Similarly, Catsup, a Drosophila ZIP7 ortholog, which plays a crucial role in catecholamine synthesis, is also involved in the ER stress response [75,76]. Along with ZIP7, Golgi-localized ZIP9 is also thought to contribute to secretory homeostasis [67], and likewise, ZIP13 contributes directly to zinc homeostasis in the early secretory pathway by mobilizing zinc from the Golgi apparatus, or indirectly by releasing zinc from intracellular vesicles [69]. Recently, although it is localized to the plasma membrane [77,78], ZIP14 has also been shown to play a significant role in the adaptation to ER stress [79,80], suggesting that zinc homeostasis in the early secretory pathway might be indirectly controlled by ZIP transporters that are located in other subcellular regions.
With respect to the direction of zinc transport mediated by ZNT and ZIP transporters, both decreases and increases in zinc levels in the early secretory pathway exacerbate its proper functioning and thus either increases or decreases in zinc levels will result in the homeostatic ER stress response. The molecular basis underlying this phenomenon may be explained by changes in the activity of chaperone proteins that are either positively or negatively regulated by zinc [46,47,48], although this has not yet been completely elucidated.
4. Regulation of Expression of ZNT and ZIP Transporters by ER Stress
Based on the crucial functions of zinc mobilized by ZNT and ZIP transporters in the early secretory pathway, it is easy to imagine that the transcription of ZNT and ZIP genes would be regulated by homeostatic ER stress. In fact, ZNT5 transcription increases in response to inducers of ER stress, and its promoter harbors a UPR element, which serves as the binding site for the transcription factor XBP-1 [50] (Figure 3). ZIP14 transcription is also induced by inducers of ER stress [79,80], and its promoter also has several ER stress response elements, to which the transcription factors ATF6 and ATF4 bind [79,80]. Moreover, the treatment with inducers of ER stress or N,N,N′,N′-tetrakis(2-pyridylmethyl)ethylenediamine, a zinc chelator which also causes ER stress, has been shown to induce the expression of ZIP3, ZIP7, ZIP9, ZIP13, and ZIP14, as well as ZNT3, ZNT6, ZNT7, and ZNT10 [51,62,63,79,80], although the elements responsible for the induction have not yet been identified in the promoter regions of these genes. Several homologues of both types of transporter have been shown to be increased by inducers of ER stress [81]. Thus, the fine-tuning of zinc homeostasis by these zinc transporters in the early secretory pathway, as well as the regulation of their expression triggered by homeostatic ER stress, are important control mechanisms in maintaining homeostasis [12].
5. Importance of ZNT Transporters in the Activation of Ectoenzymes in the Early Secretory Pathway
Zinc-requiring ectoenzymes, which are defined here as secretory, membrane-bound, and organelle-resident enzymes, have attracted considerable attention because they play crucial roles in various physiological functions, and in a number of pathological processes, such as cancer progression, and metastasis [1,55]. Thus, they are regarded as potential therapeutic targets in the treatment of diseases [82,83,84,85,86]. Moreover, the activities of some zinc-requiring ectoenzymes, e.g., alkaline phosphatases (ALP), may be used as clinical markers to reflect systemic zinc status [87,88]. These enzymes are synthesized in the early secretory pathway, at which point they acquire zinc for their activity, before being trafficked to the plasma membrane via the constitutive secretory pathway [12,55]. How zinc is made available to zinc-requiring ectoenzymes is largely unknown, but the importance of ZNT transporters has been partially clarified in the activation of specific enzymes. This section addresses these specific ectoenzyme activation processes in detail.
5.1. ZNT Transporters Involved in Zinc-Requiring Ectoenzyme Activation
Zinc-requiring ectoenzymes likely become active by coordinating with zinc at their active site (i.e., they become metallated) during the secretory process. Zinc coordination is generally achieved by the interaction of zinc with three or four amino acids, including His, Asp, and Glu residues [4,89,90], which must undergo precise regulation for the conversion from the apo- to holo-forms. When compared with cytosolic zinc-requiring enzymes, zinc-requiring ectoenzymes require more complicated and elaborate regulatory processes involving zinc mobilization, because their activation process requires at least two types of zinc transporters that involve two biological membranes. In the first process, zinc transport from the extracellular milieu to the cytosol (i.e., ZIP transporters) occurs, and in the second process zinc transport from the cytosol to the lumen (i.e., ZNT transporters) occurs. Information regarding the identity of the ZIP transporters involved is lacking, while information relating to the ZNT transporters is accumulating. Three ZNT transporters, constituting two independent complexes, have been shown to be indispensable in ectoenzyme metallation; one complex is formed by ZNT5 and ZNT6 as a heterodimer, in which ZNT6 operates as an auxiliary subunit, and the other is formed by ZNT7 homodimers [21,91,92]. Both of these ZNT complexes can specifically activate several zinc-requiring enzymes, such as tissue-nonspecific ALP (TNAP) and placental ALP, as well as autotaxin (ATX) [24,56,93]. These three enzymes have similar active site geometry with a bimetallic core, which consists of two zinc ions, one of which is coordinated by one Asp and two His residues, and the other coordinated by one His and two Asp residues (Table 1), although ALPs and ATX catalyze different enzymatic reactions and have different biological roles. Based on these enzymes, it could be hypothesized that specific regulation mechanisms are operative in this bimetallic core enzyme family during conversion from the apo- to holo-enzymes. Both ZNT complexes, however, can activate other zinc-requiring ectoenzymes, such as matrix metalloproteinase (MMP)-9 and probably MMP-2 [24], and thus, they likely play a critical role in the activation of many zinc-requiring ectoenzymes in the early secretory pathway.
However, some enzymes can be metallated by zinc and become activated through different pathways involving these zinc transporters. For example, carbonic anhydrase IX (CAIX) can acquire zinc via ZNT4 homodimers, in addition to ZNT5-ZNT6 heterodimers and ZNT7 homodimers [24]. ZNT4 homodimers may also be involved in carbonic anhydrase VI maturation [94]. These findings are interesting for two reasons. The first is that ZNT4 homodimers have multifunctional roles depending on their subcellular localization. For example, ZNT4 was originally reported to be localized to late endosomes [95], where it has a role in reducing cytosolic zinc toxicity [25,96], but it has also been shown to be involved in secretory pathways involving the trans-Golgi network, cytosolic vesicles, and probably other secretory vesicles. In addition, it has been shown to be involved in zinc secretion into breast milk in mice [97,98]. The second is that ZNT4 homodimers can become functionally equivalent to ZNT5-ZNT6 heterodimers or ZNT7 homodimers in the activation of specific ectoenzymes, including CAIX, in the early secretory pathway. The involvement of ZNT4 in CAIX activation is specific because ZNT2 expression failed to result in CAIX activation [24].
5.2. Insight into the Activation of TNAP and Other Ectoenzymes by ZNT5-ZNT6 Heterodimers and ZNT7 Homodimers
What affects the metallation of TNAP via ZNT5-ZNT6 heterodimers or ZNT7 homodimers? This question remains to be fully resolved but some important insights have been made to date. First, the number of zinc ions and their coordination manner at the active site does not seem to affect the activation process mediated via ZNT5-ZNT6 heterodimers or ZNT7 homodimers. The ALP protein possesses two zinc ions at the active site (zinc bimetallic core), which are coordinated by His, His, and Asp residues or Asp, Asp, and His residues, as described above [55], whereas MMP-9 has a single zinc ion at the active site, which is coordinated by three His residues (MMP-9 has another zinc ion that acts as a structural component) [55] (Table 1). CAIX, which can be activated by ZNT4 homodimers, ZNT5-ZNT6 heterodimers and ZNT7 homodimers [24], as described above, possesses a single zinc ion coordinated by three His residues at the active site, supporting this notion.
Second, a specific motif may be significantly involved in TNAP activation via ZNT5-ZNT6 heterodimers and ZNT7 homodimers [55,99]. In cells lacking both ZNT complexes, the TNAP protein is destabilized, although it is not destabilized by zinc deficiency [93]. These data show that both ZNT complexes can stabilize the TNAP protein, in addition to supplying it with zinc. In other words, the TNAP activation process can be separated into two steps: the TNAP protein is first stabilized by ZNT5-ZNT6 heterodimers or ZNT7 homodimers in the early secretory pathway and then is metallated by zinc supplied by both ZNT complexes [93] (Figure 4). In this two-step mechanism, the Pro-Pro (PP)-motif in luminal loop 2 of ZNT5 (which corresponds to luminal loop 7, because of the fact that ZNT5 has extra N-terminal TM helices [61]) and ZNT7, which is highly conserved in ZNT5 and ZNT7 homologs across multiple species, is thought to be important [100]. In model structures of ZNT5 and ZNT7, the PP-motif is located just above the intramembranous zinc-binding site in the TM helices [100], suggesting that a unique cooperative mechanism might operate between the intramembranous zinc-binding site and the PP-motif in the activation of TNAP. In contrast, a similar two-step activation mechanism does not seem to operate in the activation of ATX, because the ATX protein is not destabilized in cells lacking both ZNT5-ZNT6 heterodimers or ZNT7 homodimers, and so the PP-motif plays a somewhat minor role [24]. This discrepancy between TNAP and ATX may be explained by differences in the degree of their glycosylation, although this needs to be clarified in future studies.
Znt5 or Znt7 knockout (KO) mice show various phenotypes [101,102,103,104,105], but the association of those phenotypes with the early secretory pathway functions is unclear to date. One exception is the phenotype of osteopenia [101], which may be associated with reduced TNAP activity caused by a lack of ZNT5-ZNT6 heterodimers.
6. Importance of ZNT Transporters in Zinc-Related Regulated Secretory Pathway: After the Early Secretory Pathway
There are a number of cells that accumulate large amounts of zinc in cytoplasmic vesicles/granules (Figure 1). One can think that zinc, which is transported to the early secretory pathway, traffics those vesicles/granules through the secretory pathway and gets accumulated there. However, this is not the case. In fact, specific ZNT transporters are localized to the specific vesicles/granules and perform specific functions. In this section, representative vesicles/granules and the ZNT transporters involved in their function are briefly summarized to emphasize these points. Other aspects of this have been extensively reviewed elsewhere [1,10,11,12].
Insulin granules in pancreatic islet β-cells require high amounts of zinc in order to form insulin-zinc crystals, a process in which ZNT8 plays an indispensable role [106,107,108,109,110]. Nevertheless, a clear and crucial role for ZNT8 in regulating glucose homeostasis is yet to be established (Znt8 KO mice are largely glucose tolerant), and thus the physiological relevance of zinc accumulation in insulin secretory granules remains unclear. Although, an interesting hypothesis is that zinc, which is secreted in concert with insulin, suppresses the insulin clearance in the liver by inhibiting clathrin-dependent insulin endocytosis [110]. Alterations in ZNT8 function are thought to lead to an increase in the risk of type 2 diabetes [111,112], because the R325W polymorphism in ZNT8 is associated with an increased risk of type 2 diabetes [113], and the R-form (i.e., the increased-risk form of ZNT8) likely alters its zinc transport activity [111,112]. In addition, another study has shown that haploinsufficiency of ZNT8 is protective against type 2 diabetes [114]. The relationship between ZNT8 and type 2 diabetes therefore requires further investigation [115]. The transporters ZNT5 and ZNT7 are also relatively highly expressed in pancreatic β-cells, [61,116,117], and both may be associated with β-cell function: loss-of-function of Znt5 is associated with attenuation of the incidence of diabetes and mortality [103], whereas loss-of-function of Znt7 impairs glucose tolerance and reduces glucose-stimulated increases in plasma insulin levels, hepatic glycogen levels, and pancreatic insulin content [104,105,118]. Moreover, loss-of-function of Znt7 results in a markedly-reduced zinc content in β-cells, which is made more profound by the combined loss of function of Znt8 [118]. ZNT5 and ZNT7 may therefore contribute to β-cell function in the early secretory pathway, but not in the insulin granules themselves.
Synaptic vesicles present in a subset of glutamatergic neurons in the hippocampus and neocortex also accumulate high amounts of zinc, which is mediated by ZNT3: Znt3 KO mice lack synaptic zinc [119]. Zinc secreted from the synaptic vesicles into the extracellular space, as a result of ZNT3 activity, acts as a signaling molecule by modulating neuronal transmission and plasticity through binding to multiple ion channels, transporters, and receptors on postsynaptic neurons involved in neurotransmission [120,121,122]. The importance of synaptic zinc has been confirmed in knock-in mice studies, using glycine and N-methyl-d-aspartate receptors, in which the zinc-binding sites in each receptor were mutated [123,124]. A disturbance of synaptic zinc homeostasis or a dysfunction in ZNT3 has been suggested to result in neurodegenerative diseases [125,126,127].
Because zinc is essential for the growth and health of neonates, breast milk contains high amounts of zinc, considerably higher than the levels found in serum. The transporter ZNT2 is responsible for supplying zinc to the breast milk produced by the mammary epithelial cells in humans. Mothers with missense or nonsense mutations in the ZNT2 gene secrete zinc-deficient milk (75–95% reduction), and thus infants exclusively breast-fed by mothers carrying the mutation experience transient neonatal zinc deficiency (TNZD; OMIM 608118) [87,128,129,130,131]. Zinc-deficient milk is produced by mothers with a heterozygous mutation in the ZNT2 gene, and their infants also suffer from TNZD, suggesting that having one active copy of the ZNT2 gene is not sufficient to provide zinc levels in breast milk adequate to support normal infant growth. One report has also suggested the involvement of ZNT5-ZNT6 heterodimers in the pathogenesis of TNZD [132], but there are no reports indicating that low-zinc breast milk can be attributed to mutations in the ZNT4 gene in humans, although Znt4 is involved in low-zinc breast milk in mice [97,98].
Zinc that accumulated in granules/vesicles can be released in response to various stimuli and thereby regulate a number of diverse processes [110,133,134]. This phenomenon can be divided into two classes, the first being zinc secretion into the extracellular environment (e.g., zinc “sparks”) [133,134], as described for synaptic zinc, and the second being zinc release from intracellular stores into the cytosol (e.g., zinc “wave”) [135]. The latter phenomenon is strongly associated with the signaling functions of zinc and thus contributes to driving major signaling pathways [71,72,136]. The function of zinc in signaling has been extensively reviewed [2,41,137].
7. Perspectives
This review focuses on crucial functions of specific ZNT and ZIP zinc transporters in the early secretory pathway. As we have shown, these two classes of zinc transporter are doubtless key molecules required for the proper function of the early secretory pathway. However, there are many unsolved and fundamental questions that remain to be addressed. Specifically, how do both zinc deficiency and elevation in the early secretory pathway cause and exacerbate ER stress? How do those ZNT and ZIP transporters properly regulate zinc metabolism in a spatiotemporal manner in the early secretory pathway? Moreover, how is zinc coordinated in zinc-requiring ectoenzymes in the early secretory pathway? Are zinc chaperones required to facilitate zinc metallation of the large number of nascent proteins found in the early secretory pathway? Even the most fundamental question as to what the actual zinc concentration is in the early secretory pathway has not yet been definitively addressed, because the proposed zinc concentrations in the ER and the Golgi are controversial [138,139]. Moreover, clarification of a functional relationship(s) between the early secretory pathway and constitutive secretory or the regulated secretory pathway is required from the perspective of zinc metabolism. The answers to these questions can help our understanding of zinc in the early secretory pathway, and provide information that should be useful for the treatment of numerous diseases.
Acknowledgments
This work was supported by Grants-in-Aid for Scientific Research (B) from the Japan Society for the Promotion of Science (KAKENHI, Grant No. 15H04501 to Taiho Kambe). Taka-aki Takeda is a Research Fellow (DC1) of the Japan Society for the Promotion of Science.
Author Contributions
Taiho Kambe wrote the paper. Mayu Matsunaga and Taka-aki Takeda edited the manuscript and described the figures.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ALP | Alkaline phosphatase |
|---|---|
| ATX | Autotaxin |
| CAIX | Carbonic anhydrase IX |
| CDF | Cation diffusion facilitator |
| ER | Endoplasmic reticulum |
| KO | Knockout |
| MMP | Matrix metalloproteinase |
| PP | Pro-Pro |
| S(E)R | Sarco(endo)plasmic reticulum |
| SLC | Solute carrier |
| TM | Transmembrane |
| TNAP | Tissue non-specific alkaline phosphatase |
| TNZD | Transient neonatal zinc deficiency |
| UPR | Unfolded protein response |
| ZIP | Zrt- and Irt-like protein |
| ZNT | Zn transporter |
References
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The Physiological, Biochemical, and Molecular Roles of Zinc Transporters in Zinc Homeostasis and Metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Takeda, T.A.; Takagishi, T.; Fukue, K.; Kambe, T.; Fukada, T. Physiological roles of zinc transporters: Molecular and genetic importance in zinc homeostasis. J. Physiol. Sci. 2017, 67, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Zinc through the three domains of life. J. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Li, Y. Coordination dynamics of zinc in proteins. Chem. Rev. 2009, 109, 4682–4707. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Devirgiliis, C.; Zalewski, P.D.; Perozzi, G.; Murgia, C. Zinc fluxes and zinc transporter genes in chronic diseases. Mutat. Res. 2007, 622, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Age-Related Eye Disease Study Research Group. A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E, beta carotene, and zinc for age-related macular degeneration and vision loss: AREDS report No. 8. Arch. Ophthalmol. 2001, 119, 1417–1436. [Google Scholar]
- Kambe, T.; Hashimoto, A.; Fujimoto, S. Current understanding of ZIP and ZnT zinc transporters in human health and diseases. Cell. Mol. Life Sci. 2014, 71, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Lichten, L.A.; Cousins, R.J. Mammalian zinc transporters: Nutritional and physiologic regulation. Annu. Rev. Nutr. 2009, 29, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, S.L.; McCormick, N.H.; Velasquez, V.; Lopez, V. Zinc in specialized secretory tissues: Roles in the pancreas, prostate, and mammary gland. Adv. Nutr. 2011, 2, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Hennigar, S.R.; Kelleher, S.L. Zinc networks: The cell-specific compartmentalization of zinc for specialized functions. Biol. Chem. 2012, 393, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T. An overview of a wide range of functions of ZnT and Zip zinc transporters in the secretory pathway. Biosci. Biotechnol. Biochem. 2011, 75, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T. Regulation of zinc transport. In Encyclopedia of Inorganic and Bioinorganic Chemistry; Culotta, V., Scott, R.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 301–309. [Google Scholar] [CrossRef]
- Kambe, T. Molecular Architecture and Function of ZnT Transporters. Curr. Top. Membr. 2012, 69, 199–220. [Google Scholar] [PubMed]
- Lu, M.; Fu, D. Structure of the zinc transporter YiiP. Science 2007, 317, 1746–1748. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Chai, J.; Fu, D. Structural basis for autoregulation of the zinc transporter YiiP. Nat. Struct. Mol. Biol. 2009, 16, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Coudray, N.; Valvo, S.; Hu, M.; Lasala, R.; Kim, C.; Vink, M.; Zhou, M.; Provasi, D.; Filizola, M.; Tao, J.; et al. Inward-facing conformation of the zinc transporter YiiP revealed by cryoelectron microscopy. Proc. Natl. Acad. Sci. USA 2013, 110, 2140–2145. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chai, J.; Cheng, J.; D’Mello, R.; Chance, M.R.; Fu, D. Visualizing the kinetic power stroke that drives proton-coupled zinc(II) transport. Nature 2014, 512, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Ohana, E.; Hoch, E.; Keasar, C.; Kambe, T.; Yifrach, O.; Hershfinkel, M.; Sekler, I. Identification of the Zn2+ binding site and mode of operation of a mammalian Zn2+ transporter. J. Biol. Chem. 2009, 284, 17677–17686. [Google Scholar] [CrossRef] [PubMed]
- Shusterman, E.; Beharier, O.; Shiri, L.; Zarivach, R.; Etzion, Y.; Campbell, C.R.; Lee, I.H.; Okabayashi, K.; Dinudom, A.; Cook, D.I.; et al. ZnT-1 extrudes zinc from mammalian cells functioning as a Zn(2+)/H(+) exchanger. Metallomics 2014, 6, 1656–1663. [Google Scholar] [CrossRef] [PubMed]
- Fukunaka, A.; Suzuki, T.; Kurokawa, Y.; Yamazaki, T.; Fujiwara, N.; Ishihara, K.; Migaki, H.; Okumura, K.; Masuda, S.; Yamaguchi-Iwai, Y.; et al. Demonstration and characterization of the heterodimerization of ZnT5 and ZnT6 in the early secretory pathway. J. Biol. Chem. 2009, 284, 30798–30806. [Google Scholar] [CrossRef] [PubMed]
- Golan, Y.; Berman, B.; Assaraf, Y.G. Heterodimerization, altered subcellular localization, and function of multiple zinc transporters in viable cells using bimolecular fluorescence complementation. J. Biol. Chem. 2015, 290, 9050–9063. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Feresin, R.G.; Falcon-Perez, J.M.; Salazar, G. Differential Targeting of SLC30A10/ZnT10 Heterodimers to Endolysosomal Compartments Modulates EGF-Induced MEK/ERK1/2 Activity. Traffic 2016, 17, 267–288. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, T.; Kurokawa, Y.; Chiche, J.; Pouyssegur, J.; Sato, H.; Fukuzawa, H.; Nagao, M.; Kambe, T. Dissecting the Process of Activation of Cancer-promoting Zinc-requiring Ectoenzymes by Zinc Metalation Mediated by ZNT Transporters. J. Biol. Chem. 2017, 292, 2159–2173. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Itsumura, N.; Tsuji, T.; Anan, Y.; Tsuji, N.; Ogra, Y.; Kimura, T.; Miyamae, Y.; Masuda, S.; Nagao, M.; et al. Cooperative functions of ZnT1, metallothionein and ZnT4 in the cytoplasm are required for full activation of tnap in the early secretory pathway. PLoS ONE 2013, 8, e77445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golan, Y.; Itsumura, N.; Glaser, F.; Berman, B.; Kambe, T.; Assaraf, Y.G. Molecular Basis of Transient Neonatal Zinc Deficiency: Novel ZNT2 Mutations Disrupting ZINC Binding and Permeation. J. Biol. Chem. 2016, 291, 13546–13559. [Google Scholar] [CrossRef] [PubMed]
- Hoch, E.; Lin, W.; Chai, J.; Hershfinkel, M.; Fu, D.; Sekler, I. Histidine pairing at the metal transport site of mammalian ZnT transporters controls Zn2+ over Cd2+ selectivity. Proc. Natl. Acad. Sci. USA 2012, 109, 7202–7207. [Google Scholar] [CrossRef] [PubMed]
- Nishito, Y.; Tsuji, N.; Fujishiro, H.; Takeda, T.; Yamazaki, T.; Teranishi, F.; Okazaki, F.; Matsunaga, A.; Tuschl, K.; Rao, R.; et al. Direct comparison of manganese detoxification/efflux proteins and molecular characterization of ZnT10 as a manganese transporter. J. Biol. Chem. 2016, 291, 14773–14787. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.E.; Giedroc, D.P. Functional Determinants of Metal Ion Transport and Selectivity in Paralogous Cation Diffusion Facilitator Transporters CzcD and MntE in Streptococcus pneumoniae. J. Bacteriol. 2016, 198, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Gustin, J.L.; Zanis, M.J.; Salt, D.E. Structure and evolution of the plant cation diffusion facilitator family of ion transporters. BMC Evol. Biol. 2011, 11, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambe, T.; Suzuki, T.; Nagao, M.; Yamaguchi-Iwai, Y. Sequence similarity and functional relationship among eukaryotic ZIP and CDF transporters. Genom. Proteom. Bioinform. 2006, 4, 1–9. [Google Scholar] [CrossRef]
- Antala, S.; Ovchinnikov, S.; Kamisetty, H.; Baker, D.; Dempski, R.E. Computation and Functional Studies Provide a Model for the Structure of the Zinc Transporter hZIP4. J. Biol. Chem. 2015, 290, 17796–17805. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, J.; Fellner, M.; Zhang, C.; Sui, D.; Hu, J. Crystal structures of a ZIP zinc transporter reveal a binuclear metal center in the transport pathway. Sci. Adv. 2017, 3, e1700344. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Fukada, T.; Hosaka, T.; Yamasaki, S.; Ohashi, W.; Hojyo, S.; Miyai, T.; Nishida, K.; Yokoyama, S.; Hirano, T. Biochemical characterization of human ZIP13 protein: A homo-dimerized zinc transporter involved in the spondylocheiro dysplastic Ehlers-Danlos syndrome. J. Biol. Chem. 2011, 286, 40255–40265. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Muraina, I.A.; Brethour, D.; Schmitt-Ulms, G.; Nimmanon, T.; Ziliotto, S.; Kille, P.; Hogstrand, C. Zinc transporter ZIP10 forms a heteromer with ZIP6 which regulates embryonic development and cell migration. Biochem. J. 2016, 473, 2531–2544. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chai, J.; Love, J.; Fu, D. Selective electrodiffusion of zinc ions in a Zrt-, Irt-like protein, ZIPB. J. Biol. Chem. 2010, 285, 39013–39020. [Google Scholar] [CrossRef] [PubMed]
- Gaither, L.A.; Eide, D.J. Functional expression of the human hZIP2 zinc transporter. J. Biol. Chem. 2000, 275, 5560–5564. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Girijashanker, K.; Dalton, T.P.; Reed, J.; Li, H.; Soleimani, M.; Nebert, D.W. ZIP8, member of the solute-carrier-39 (SLC39) metal-transporter family: Characterization of transporter properties. Mol. Pharmacol. 2006, 70, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Girijashanker, K.; He, L.; Soleimani, M.; Reed, J.M.; Li, H.; Liu, Z.; Wang, B.; Dalton, T.P.; Nebert, D.W. Slc39a14 gene encodes ZIP14, a metal/bicarbonate symporter: Similarities to the ZIP8 transporter. Mol. Pharmacol. 2008, 73, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Kambe, T. Molecular and genetic features of zinc transporters in physiology and pathogenesis. Metallomics 2011, 3, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Sui, D.; Hu, J. Structural insights of ZIP4 extracellular domain critical for optimal zinc transport. Nat. Commun. 2016, 7, 11979. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, S.; Huo, H.; Salehzadeh, A.; Pocanschi, C.L.; Watts, J.C.; Wille, H.; Westaway, D.; Rogaeva, E.; St George-Hyslop, P.H.; Schmitt-Ulms, G. Family reunion—The ZIP/prion gene family. Prog. Neurobiol. 2011, 93, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Jenkitkasemwong, S.; Wang, C.Y.; Mackenzie, B.; Knutson, M.D. Physiologic implications of metal-ion transport by ZIP14 and ZIP8. Biometals 2012, 25, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Vembar, S.S.; Brodsky, J.L. One step at a time: Endoplasmic reticulum-associated degradation. Nat. Rev. Mol. Cell Biol. 2008, 9, 944–957. [Google Scholar] [CrossRef] [PubMed]
- Leach, M.R.; Cohen-Doyle, M.F.; Thomas, D.Y.; Williams, D.B. Localization of the lectin, ERp57 binding, and polypeptide binding sites of calnexin and calreticulin. J. Biol. Chem. 2002, 277, 29686–29697. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Ihara, Y.; Leach, M.R.; Cohen-Doyle, M.F.; Williams, D.B. Calreticulin functions in vitro as a molecular chaperone for both glycosylated and non-glycosylated proteins. EMBO J. 1999, 18, 6718–6729. [Google Scholar] [CrossRef] [PubMed]
- Solovyov, A.; Gilbert, H.F. Zinc-dependent dimerization of the folding catalyst, protein disulfide isomerase. Protein Sci. 2004, 13, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.D.; Wang, F.; MacDiarmid, C.W.; Clark, S.; Lyons, T.; Eide, D.J. Zinc and the Msc2 zinc transporter protein are required for endoplasmic reticulum function. J. Cell Biol. 2004, 166, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, K.; Yamazaki, T.; Ishida, Y.; Suzuki, T.; Oda, K.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Zinc transport complexes contribute to the homeostatic maintenance of secretory pathway function in vertebrate cells. J. Biol. Chem. 2006, 281, 17743–17750. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, W.; Kimura, S.; Iwanaga, T.; Furusawa, Y.; Irie, T.; Izumi, H.; Watanabe, T.; Hijikata, A.; Hara, T.; Ohara, O.; et al. Zinc Transporter SLC39A7/ZIP7 Promotes Intestinal Epithelial Self-Renewal by Resolving ER Stress. PLoS Genet. 2016, 12, e1006349. [Google Scholar] [CrossRef] [PubMed]
- Tuncay, E.; Bitirim, V.C.; Durak, A.; Carrat, G.R.J.; Taylor, K.M.; Rutter, G.A.; Turan, B. Hyperglycemia-Induced Changes in ZIP7 and ZnT7 Expression Cause Zn2+ Release from the Sarco(endo)plasmic Reticulum and Mediate ER Stress in the Heart. Diabetes 2017, 66, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Bhin, J.; Seo, J.; Kim, S.Y.; Lee, E.; Park, K.; Choi, D.H.; Takagishi, T.; Hara, T.; Hwang, D.; et al. Requirement of Zinc Transporter SLC39A7/ZIP7 for Dermal Development to Fine-Tune Endoplasmic Reticulum Function by Regulating Protein Disulfide Isomerase. J. Investig. Dermatol. 2017, 137, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Illades, D.; Chen, P.; Zogzas, C.E.; Hutchens, S.; Mercado, J.M.; Swaim, C.D.; Morrisett, R.A.; Bowman, A.B.; Aschner, M.; Mukhopadhyay, S. SLC30A10 is a cell surface-localized manganese efflux transporter, and parkinsonism-causing mutations block its intracellular trafficking and efflux activity. J. Neurosci. 2014, 34, 14079–14095. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Takeda, T.A.; Nishito, Y. Activation of zinc-requiring ectoenzymes by ZnT transporters during the secretory process: Biochemical and molecular aspects. Arch. Biochem. Biophys. 2016, 611, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ishihara, K.; Migaki, H.; Matsuura, W.; Kohda, A.; Okumura, K.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Zinc transporters, ZnT5 and ZnT7, are required for the activation of alkaline phosphatases, zinc-requiring enzymes that are glycosylphosphatidylinositol-anchored to the cytoplasmic membrane. J. Biol. Chem. 2005, 280, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Kirschke, C.P.; Gitschier, J. Functional characterization of a novel mammalian zinc transporter, ZnT6. J. Biol. Chem. 2002, 277, 26389–26395. [Google Scholar] [CrossRef] [PubMed]
- Kirschke, C.P.; Huang, L. ZnT7, a Novel Mammalian Zinc Transporter, Accumulates Zinc in the Golgi Apparatus. J. Biol. Chem. 2003, 278, 4096–4102. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T. Methods to evaluate zinc transport into and out of the secretory and endosomal-lysosomal compartments in DT40 cells. Methods Enzymol. 2014, 534, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.D.; Macdiarmid, C.W.; Eide, D.J. Heteromeric protein complexes mediate zinc transport into the secretory pathway of eukaryotic cells. J. Biol. Chem. 2005, 280, 28811–28818. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Narita, H.; Yamaguchi-Iwai, Y.; Hirose, J.; Amano, T.; Sugiura, N.; Sasaki, R.; Mori, K.; Iwanaga, T.; Nagao, M. Cloning and characterization of a novel mammalian zinc transporter, zinc transporter 5, abundantly expressed in pancreatic beta cells. J. Biol. Chem. 2002, 277, 19049–19055. [Google Scholar] [CrossRef] [PubMed]
- Kurita, H.; Okuda, R.; Yokoo, K.; Inden, M.; Hozumi, I. Protective roles of SLC30A3 against endoplasmic reticulum stress via ERK1/2 activation. Biochem. Biophys. Res. Commun. 2016, 479, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Go, S.; Kurita, H.; Yokoo, K.; Inden, M.; Kambe, T.; Hozumi, I. Protective function of SLC30A10 induced via PERK-ATF4 pathway against 1-methyl-4-phenylpyridinium. Biochem. Biophys. Res. Commun. 2017. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Morgan, H.E.; Johnson, A.; Nicholson, R.I. Structure-function analysis of HKE4, a member of the new LIV-1 subfamily of zinc transporters. Biochem. J. 2004, 377, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Kirschke, C.P.; Zhang, Y.; Yu, Y.Y. The ZIP7 gene (Slc39a7) encodes a zinc transporter involved in zinc homeostasis of the Golgi apparatus. J. Biol. Chem. 2005, 280, 15456–15463. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Civic, N.; Furuichi, T.; Shimoda, S.; Mishima, K.; Higashiyama, H.; Idaira, Y.; Asada, Y.; Kitamura, H.; Yamasaki, S.; et al. The zinc transporter SLC39A13/ZIP13 is required for connective tissue development; its involvement in BMP/TGF-beta signaling pathways. PLoS ONE 2008, 3, e3642. [Google Scholar] [CrossRef]
- Matsuura, W.; Yamazaki, T.; Yamaguchi-Iwai, Y.; Masuda, S.; Nagao, M.; Andrews, G.K.; Kambe, T. SLC39A9 (ZIP9) regulates zinc homeostasis in the secretory pathway: Characterization of the ZIP subfamily I protein in vertebrate cells. Biosci. Biotechnol. Biochem. 2009, 73, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Pang, Y.; Dong, J.; Berg, A.H. Identification and characterization of membrane androgen receptors in the ZIP9 zinc transporter subfamily: II. Role of human ZIP9 in testosterone-induced prostate and breast cancer cell apoptosis. Endocrinology 2014, 155, 4250–4265. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Walker, J.M.; Wang, F.; Park, J.G.; Palmer, A.E.; Giunta, C.; Rohrbach, M.; Steinmann, B.; Eide, D.J. Promotion of vesicular zinc efflux by ZIP13 and its implications for spondylocheiro dysplastic Ehlers-Danlos syndrome. Proc. Natl. Acad. Sci. USA 2012, 109, E3530–E3538. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, S.L.; Velasquez, V.; Croxford, T.P.; McCormick, N.H.; Lopez, V.; Macdavid, J. Mapping the zinc-transporting system in mammary cells: Molecular analysis reveals a phenotype-dependent zinc-transporting network during lactation. J. Cell. Physiol. 2012, 227, 1761–1770. [Google Scholar] [CrossRef] [PubMed]
- Hogstrand, C.; Kille, P.; Nicholson, R.I.; Taylor, K.M. Zinc transporters and cancer: A potential role for ZIP7 as a hub for tyrosine kinase activation. Trends Mol. Med. 2009, 15, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Hiscox, S.; Nicholson, R.I.; Hogstrand, C.; Kille, P. Protein Kinase CK2 Triggers Cytosolic Zinc Signaling Pathways by Phosphorylation of Zinc Channel ZIP7. Sci. Signal. 2012, 5, ra11. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Fukunaka, A.; Hagihara, M.; Watanabe, K.; Kamino, S.; Kambe, T.; Enomoto, S.; Hiromura, M. Essential Role of the Zinc Transporter ZIP9/SLC39A9 in Regulating the Activations of Akt and Erk in B-Cell Receptor Signaling Pathway in DT40 Cells. PLoS ONE 2013, 8, e58022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumanovics, A.; Poruk, K.E.; Osborn, K.A.; Ward, D.M.; Kaplan, J. YKE4 (YIL023C) encodes a bidirectional zinc transporter in the endoplasmic reticulum of Saccharomyces cerevisiae. J. Biol. Chem. 2006, 281, 22566–22574. [Google Scholar] [CrossRef] [PubMed]
- Stathakis, D.G.; Burton, D.Y.; McIvor, W.E.; Krishnakumar, S.; Wright, T.R.; O’Donnell, J.M. The catecholamines up (Catsup) protein of Drosophila melanogaster functions as a negative regulator of tyrosine hydroxylase activity. Genetics 1999, 153, 361–382. [Google Scholar] [PubMed]
- Groth, C.; Sasamura, T.; Khanna, M.R.; Whitley, M.; Fortini, M.E. Protein trafficking abnormalities in Drosophila tissues with impaired activity of the ZIP7 zinc transporter Catsup. Development 2013, 140, 3018–3027. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Morgan, H.E.; Johnson, A.; Nicholson, R.I. Structure-function analysis of a novel member of the LIV-1 subfamily of zinc transporters, ZIP14. FEBS Lett. 2005, 579, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, J.P.; Lichten, L.A.; Rivera, S.; Blanchard, R.K.; Aydemir, T.B.; Knutson, M.D.; Ganz, T.; Cousins, R.J. Interleukin-6 regulates the zinc transporter Zip14 in liver and contributes to the hypozincemia of the acute-phase response. Proc. Natl. Acad. Sci. USA 2005, 102, 6843–6848. [Google Scholar] [CrossRef] [PubMed]
- Homma, K.; Fujisawa, T.; Tsuburaya, N.; Yamaguchi, N.; Kadowaki, H.; Takeda, K.; Nishitoh, H.; Matsuzawa, A.; Naguro, I.; Ichijo, H. SOD1 as a Molecular Switch for Initiating the Homeostatic ER Stress Response under Zinc Deficiency. Mol. Cell 2013, 52, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Aydemir, T.B.; Kim, J.; Cousins, R.J. Hepatic ZIP14-mediated zinc transport is required for adaptation to endoplasmic reticulum stress. Proc. Natl. Acad. Sci. USA 2017, 114, E5805–E5814. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Ellis, R.E.; Sakaki, K.; Kaufman, R.J. Genetic Interactions Due to Constitutive and Inducible Gene Regulation Mediated by the Unfolded Protein Response in C. elegans. PLoS Genet. 2005, 1, e37. [Google Scholar] [CrossRef] [PubMed]
- Overall, C.M.; Lopez-Otin, C. Strategies for MMP inhibition in cancer: Innovations for the post-trial era. Nat. Rev. Cancer 2002, 2, 657–672. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat. Rev. Drug Discov. 2008, 7, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Reich, R.; Hoffman, A.; Veerendhar, A.; Maresca, A.; Innocenti, A.; Supuran, C.T.; Breuer, E. Carbamoylphosphonates control tumor cell proliferation and dissemination by simultaneously inhibiting carbonic anhydrase IX and matrix metalloproteinase-2. Toward nontoxic chemotherapy targeting tumor microenvironment. J. Med. Chem. 2012, 55, 7875–7882. [Google Scholar] [CrossRef] [PubMed]
- Reich, R.; Hoffman, A.; Suresh, R.R.; Shai, O.; Frant, J.; Maresca, A.; Supuran, C.T.; Breuer, E. Carbamoylphosphonates inhibit autotaxin and metastasis formation in vivo. J. Enzym. Inhib. Med. Chem. 2015, 30, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Neri, D.; Supuran, C.T. Interfering with pH regulation in tumours as a therapeutic strategy. Nat. Rev. Drug Discov. 2011, 10, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Itsumura, N.; Kibihara, Y.; Fukue, K.; Miyata, A.; Fukushima, K.; Tamagawa-Mineoka, R.; Katoh, N.; Nishito, Y.; Ishida, R.; Narita, H.; et al. Novel mutations in SLC30A2 involved in the pathogenesis of transient neonatal zinc deficiency. Pediatr. Res. 2016, 80, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, A.; Nakagawa, M.; Tsujimura, N.; Miyazaki, S.; Kizu, K.; Goto, T.; Komatsu, Y.; Matsunaga, A.; Shirakawa, H.; Narita, H.; et al. Properties of Zip4 accumulation during zinc deficiency and its usefulness to evaluate zinc status: A study of the effects of zinc deficiency during lactation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R459–R468. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [PubMed]
- Kochanczyk, T.; Drozd, A.; Krezel, A. Relationship between the architecture of zinc coordination and zinc binding affinity in proteins--insights into zinc regulation. Metallomics 2015, 7, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ishihara, K.; Migaki, H.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Two different zinc transport complexes of cation diffusion facilitator proteins localized in the secretory pathway operate to activate alkaline phosphatases in vertebrate cells. J. Biol. Chem. 2005, 280, 30956–30962. [Google Scholar] [CrossRef] [PubMed]
- Lasry, I.; Golan, Y.; Berman, B.; Amram, N.; Glaser, F.; Assaraf, Y.G. In Situ dimerization of multiple wild type and mutant zinc transporters in live cells using bimolecular fluorescence complementation. J. Biol. Chem. 2014, 289, 7275–7292. [Google Scholar] [CrossRef] [PubMed]
- Fukunaka, A.; Kurokawa, Y.; Teranishi, F.; Sekler, I.; Oda, K.; Ackland, M.L.; Faundez, V.; Hiromura, M.; Masuda, S.; Nagao, M.; et al. Tissue Nonspecific Alkaline Phosphatase Is Activated via a Two-step Mechanism by Zinc Transport Complexes in the Early Secretory Pathway. J. Biol. Chem. 2011, 286, 16363–16373. [Google Scholar] [CrossRef] [PubMed]
- McCormick, N.H.; Kelleher, S.L. ZnT4 provides zinc to zinc-dependent proteins in the trans-Golgi network critical for cell function and Zn export in mammary epithelial cells. Am. J. Physiol. Cell Physiol. 2012, 303, C291–C297. [Google Scholar] [CrossRef] [PubMed]
- Murgia, C.; Vespignani, I.; Cerase, J.; Nobili, F.; Perozzi, G. Cloning, expression, and vesicular localization of zinc transporter Dri 27/ZnT4 in intestinal tissue and cells. Am. J. Physiol. 1999, 277, G1231–G1239. [Google Scholar] [PubMed]
- Kukic, I.; Lee, J.K.; Coblentz, J.; Kelleher, S.L.; Kiselyov, K. Zinc-dependent lysosomal enlargement in TRPML1-deficient cells involves MTF-1 transcription factor and ZnT4 (Slc30a4) transporter. Biochem. J. 2013, 451, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Gitschier, J. A novel gene involved in zinc transport is deficient in the lethal milk mouse. Nat. Genet. 1997, 17, 292–297. [Google Scholar] [CrossRef] [PubMed]
- McCormick, N.H.; Lee, S.; Hennigar, S.R.; Kelleher, S.L. ZnT4 (SLC30A4)-null (“lethal milk”) mice have defects in mammary gland secretion and hallmarks of precocious involution during lactation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R33–R40. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kambe, T. The Functions of Metallothionein and ZIP and ZnT Transporters: An Overview and Perspective. Int. J. Mol. Sci. 2016, 17, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, S.; Tsuji, T.; Fujiwara, T.; Takeda, T.A.; Merriman, C.; Fukunaka, A.; Nishito, Y.; Fu, D.; Hoch, E.; Sekler, I.; et al. The PP-motif in luminal loop 2 of ZnT transporters plays a pivotal role in TNAP activation. Biochem. J. 2016, 473, 2611–2621. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Matsuda, K.; Itoh, M.; Kawaguchi, H.; Tomoike, H.; Aoyagi, T.; Nagai, R.; Hori, M.; Nakamura, Y.; Tanaka, T. Osteopenia and male-specific sudden cardiac death in mice lacking a zinc transporter gene, Znt5. Hum. Mol. Genet. 2002, 11, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Hasegawa, A.; Nakae, S.; Oboki, K.; Saito, H.; Yamasaki, S.; Hirano, T. Zinc transporter Znt5/Slc30a5 is required for the mast cell-mediated delayed-type allergic reaction but not the immediate-type reaction. J. Exp. Med. 2009, 206, 1351–1364. [Google Scholar] [CrossRef] [PubMed]
- Sheline, C.T.; Shi, C.; Takata, T.; Zhu, J.; Zhang, W.; Sheline, P.J.; Cai, A.L.; Li, L. Dietary zinc reduction, pyruvate supplementation, or zinc transporter 5 knockout attenuates beta-cell death in nonobese diabetic mice, islets, and insulinoma cells. J. Nutr. 2012, 142, 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Yu, Y.Y.; Kirschke, C.P.; Gertz, E.R.; Lloyd, K.K. Znt7 (Slc30a7)-deficient mice display reduced body zinc status and body fat accumulation. J. Biol. Chem. 2007, 282, 37053–37063. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Kirschke, C.P.; Lay, Y.A.; Levy, L.B.; Lamirande, D.E.; Zhang, P.H. Znt7-null mice are more susceptible to diet-induced glucose intolerance and insulin resistance. J. Biol. Chem. 2012, 287, 33883–33896. [Google Scholar] [CrossRef] [PubMed]
- Pound, L.D.; Sarkar, S.A.; Benninger, R.K.; Wang, Y.; Suwanichkul, A.; Shadoan, M.K.; Printz, R.L.; Oeser, J.K.; Lee, C.E.; Piston, D.W.; et al. Deletion of the mouse Slc30a8 gene encoding zinc transporter-8 results in impaired insulin secretion. Biochem. J. 2009, 421, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, T.J.; Bellomo, E.A.; Wijesekara, N.; Loder, M.K.; Baldwin, J.M.; Gyulkhandanyan, A.V.; Koshkin, V.; Tarasov, A.I.; Carzaniga, R.; Kronenberger, K.; et al. Insulin storage and glucose homeostasis in mice null for the granule zinc transporter ZnT8 and studies of the type 2 diabetes-associated variants. Diabetes 2009, 58, 2070–2083. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, K.; Ravier, M.A.; Schraenen, A.; Creemers, J.W.; Van de Plas, R.; Granvik, M.; Van Lommel, L.; Waelkens, E.; Chimienti, F.; Rutter, G.A.; et al. Insulin crystallization depends on zinc transporter ZnT8 expression, but is not required for normal glucose homeostasis in mice. Proc. Natl. Acad. Sci. USA 2009, 106, 14872–14877. [Google Scholar] [CrossRef] [PubMed]
- Wijesekara, N.; Dai, F.F.; Hardy, A.B.; Giglou, P.R.; Bhattacharjee, A.; Koshkin, V.; Chimienti, F.; Gaisano, H.Y.; Rutter, G.A.; Wheeler, M.B. Beta cell-specific Znt8 deletion in mice causes marked defects in insulin processing, crystallisation and secretion. Diabetologia 2010, 53, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, M.; Fujitani, Y.; Hara, A.; Uchida, T.; Tamura, Y.; Takeno, K.; Kawaguchi, M.; Watanabe, T.; Ogihara, T.; Fukunaka, A.; et al. The diabetes-susceptible gene SLC30A8/ZnT8 regulates hepatic insulin clearance. J. Clin. Investig. 2013, 123, 4513–4524. [Google Scholar] [CrossRef] [PubMed]
- Merriman, C.; Huang, Q.; Rutter, G.A.; Fu, D. Lipid-tuned Zinc Transport Activity of Human ZnT8 Protein Correlates with Risk for Type-2 Diabetes. J. Biol. Chem. 2016, 291, 26950–26957. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bai, S.; Sheline, C.T. hZnT8 (Slc30a8) Transgenic Mice That Overexpress the R325W Polymorph Have Reduced Islet Zn2+ and Proinsulin Levels, Increased Glucose Tolerance After a High-Fat Diet, and Altered Levels of Pancreatic Zinc Binding Proteins. Diabetes 2017, 66, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Sladek, R.; Rocheleau, G.; Rung, J.; Dina, C.; Shen, L.; Serre, D.; Boutin, P.; Vincent, D.; Belisle, A.; Hadjadj, S.; et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature 2007, 445, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Flannick, J.; Thorleifsson, G.; Beer, N.L.; Jacobs, S.B.; Grarup, N.; Burtt, N.P.; Mahajan, A.; Fuchsberger, C.; Atzmon, G.; Benediktsson, R.; et al. Loss-of-function mutations in SLC30A8 protect against type 2 diabetes. Nat. Genet. 2014, 46, 357–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabosseau, P.; Rutter, G.A. Zinc and diabetes. Arch Biochem Biophys. Arch. Biochem. Biophys. 2016, 611, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Yan, M.; Kirschke, C.P. Over-expression of ZnT7 increases insulin synthesis and secretion in pancreatic beta-cells by promoting insulin gene transcription. Exp. Cell Res. 2010, 316, 2630–2643. [Google Scholar] [CrossRef] [PubMed]
- Bellomo, E.A.; Meur, G.; Rutter, G.A. Glucose regulates free cytosolic Zn2+ concentration, Slc39 (ZiP), and metallothionein gene expression in primary pancreatic islet beta-cells. J. Biol. Chem. 2011, 286, 25778–25789. [Google Scholar] [CrossRef] [PubMed]
- Syring, K.E.; Boortz, K.A.; Oeser, J.K.; Ustione, A.; Platt, K.A.; Shadoan, M.K.; McGuinness, O.P.; Piston, D.W.; Powell, D.R.; O’Brien, R.M. Combined Deletion of Slc30a7 and Slc30a8 Unmasks a Critical Role for ZnT8 in Glucose-Stimulated Insulin Secretion. Endocrinology 2016, 157, 4534–4541. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.B.; Wenzel, H.J.; Kafer, K.E.; Schwartzkroin, P.A.; Palmiter, R.D. Elimination of zinc from synaptic vesicles in the intact mouse brain by disruption of the ZnT3 gene. Proc. Natl. Acad. Sci. USA 1999, 96, 1716–1721. [Google Scholar] [CrossRef] [PubMed]
- Sensi, S.L.; Paoletti, P.; Koh, J.Y.; Aizenman, E.; Bush, A.I.; Hershfinkel, M. The neurophysiology and pathology of brain zinc. J. Neurosci. 2011, 31, 16076–16085. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Nakamura, M.; Fujii, H.; Tamano, H. Synaptic Zn2+ homeostasis and its significance. Metallomics 2013, 5, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Vergnano, A.M.; Rebola, N.; Savtchenko, L.P.; Pinheiro, P.S.; Casado, M.; Kieffer, B.L.; Rusakov, D.A.; Mulle, C.; Paoletti, P. Zinc dynamics and action at excitatory synapses. Neuron 2014, 82, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Hirzel, K.; Muller, U.; Latal, A.T.; Hulsmann, S.; Grudzinska, J.; Seeliger, M.W.; Betz, H.; Laube, B. Hyperekplexia phenotype of glycine receptor alpha1 subunit mutant mice identifies Zn2+ as an essential endogenous modulator of glycinergic neurotransmission. Neuron 2006, 52, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, C.; Vergnano, A.M.; Filliol, D.; Ouagazzal, A.M.; Le Goff, A.; Carvalho, S.; Reiss, D.; Gaveriaux-Ruff, C.; Neyton, J.; Paoletti, P.; et al. Zinc alleviates pain through high-affinity binding to the NMDA receptor NR2A subunit. Nat. Neurosci. 2011, 14, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Adlard, P.A.; Parncutt, J.M.; Finkelstein, D.I.; Bush, A.I. Cognitive loss in zinc transporter-3 knock-out mice: A phenocopy for the synaptic and memory deficits of Alzheimer’s disease? J. Neurosci. 2010, 30, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Barnham, K.J.; Bush, A.I. Biological metals and metal-targeting compounds in major neurodegenerative diseases. Chem. Soc. Rev. 2014, 43, 6727–6749. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Noguchi, T.; Ikegami, S.; Sakurai, T.; Kakita, A.; Toyoshima, Y.; Kambe, T.; Yamada, M.; Inden, M.; Hara, H.; et al. Zinc transporters ZnT3 and ZnT6 are downregulated in the spinal cords of patients with sporadic amyotrophic lateral sclerosis. J. Neurosci. Res. 2015, 93, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Chowanadisai, W.; Lonnerdal, B.; Kelleher, S.L. Identification of a mutation in SLC30A2 (ZnT-2) in women with low milk zinc concentration that results in transient neonatal zinc deficiency. J. Biol. Chem. 2006, 281, 39699–39707. [Google Scholar] [CrossRef] [PubMed]
- Lasry, I.; Seo, Y.A.; Ityel, H.; Shalva, N.; Pode-Shakked, B.; Glaser, F.; Berman, B.; Berezovsky, I.; Goncearenco, A.; Klar, A.; et al. A Dominant Negative Heterozygous G87R Mutation in the Zinc Transporter, ZnT-2 (SLC30A2), Results in Transient Neonatal Zinc Deficiency. J. Biol. Chem. 2012, 287, 29348–29361. [Google Scholar] [CrossRef] [PubMed]
- Itsumura, N.; Inamo, Y.; Okazaki, F.; Teranishi, F.; Narita, H.; Kambe, T.; Kodama, H. Compound Heterozygous Mutations in SLC30A2/ZnT2 Results in Low Milk Zinc Concentrations: A Novel Mechanism for Zinc Deficiency in a Breast-Fed Infant. PLoS ONE 2013, 8, e64045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miletta, M.C.; Bieri, A.; Kernland, K.; Schoni, M.H.; Petkovic, V.; Fluck, C.E.; Eble, A.; Mullis, P.E. Transient Neonatal Zinc Deficiency Caused by a Heterozygous G87R Mutation in the Zinc Transporter ZnT-2 (SLC30A2) Gene in the Mother Highlighting the Importance of Zn2+ for Normal Growth and Development. Int. J. Endocrinol. 2013, 2013, 259189. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Michalczyk, A.; McKay, J.; Ford, D.; Kambe, T.; Hudek, L.; Varigios, G.; Taylor, P.E.; Ackland, M.L. Altered expression of two zinc transporters, SLC30A5 and SLC30A6, underlies a mammary gland disorder of reduced zinc secretion into milk. Genes Nutr. 2015, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.M.; Bernhardt, M.L.; Kong, B.Y.; Ahn, R.W.; Vogt, S.; Woodruff, T.K.; O’Halloran, T.V. Zinc sparks are triggered by fertilization and facilitate cell cycle resumption in mammalian eggs. ACS Chem. Biol. 2011, 6, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Que, E.L.; Bleher, R.; Duncan, F.E.; Kong, B.Y.; Gleber, S.C.; Vogt, S.; Chen, S.; Garwin, S.A.; Bayer, A.R.; Dravid, V.P.; et al. Quantitative mapping of zinc fluxes in the mammalian egg reveals the origin of fertilization-induced zinc sparks. Nat. Chem. 2015, 7, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Hasegawa, A.; Hojyo, S.; Ohashi, W.; Fukada, T.; Nishida, K.; Hirano, T. A Novel Role of the L-Type Calcium Channel alpha(1D) Subunit as a Gatekeeper for Intracellular Zinc Signaling: Zinc Wave. PLoS ONE 2012, 7, e39654. [Google Scholar] [CrossRef] [PubMed]
- Nimmanon, T.; Ziliotto, S.; Morris, S.; Flanagan, L.; Taylor, K.M. Phosphorylation of zinc channel ZIP7 drives MAPK, PI3K and mTOR growth and proliferation signalling. Metallomics 2017, 9, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Yamasaki, S.; Nishida, K.; Murakami, M.; Hirano, T. Zinc homeostasis and signaling in health and diseases: Zinc signaling. J. Biol. Inorg. Chem. 2011, 16, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Chabosseau, P.; Tuncay, E.; Meur, G.; Bellomo, E.A.; Hessels, A.; Hughes, S.; Johnson, P.R.; Bugliani, M.; Marchetti, P.; Turan, B.; et al. Mitochondrial and ER-targeted eCALWY probes reveal high levels of free Zn2+. ACS Chem. Biol. 2014, 9, 2111–2120. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Dittmer, P.J.; Park, J.G.; Jansen, K.B.; Palmer, A.E. Measuring steady-state and dynamic endoplasmic reticulum and Golgi Zn2+ with genetically encoded sensors. Proc. Natl. Acad. Sci. USA 2011, 108, 7351–7356. [Google Scholar] [CrossRef] [PubMed]
Figure 1. Subcellular localization of ZNT and ZIP transporters. ZNT transporters move cytosolic zinc into the lumen of vesicles involved in the early secretory pathway, including the endoplasmic reticulum (ER), Golgi apparatus, as well as into cytoplasmic vesicles/granules such as synaptic and secretory vesicles and insulin granules, in which specific ZNT proteins are localized. ZNT5 and ZNT6 form heterodimers to transport zinc. ZIP transporters move zinc in the opposite direction. In contrast to the specific localization of ZIP9, ZIP13, and ZIP7 in the Golgi apparatus and the ER, the subcellular location of ZNT5-ZNT6 heterodimers has not been definitively determined.
Figure 1. Subcellular localization of ZNT and ZIP transporters. ZNT transporters move cytosolic zinc into the lumen of vesicles involved in the early secretory pathway, including the endoplasmic reticulum (ER), Golgi apparatus, as well as into cytoplasmic vesicles/granules such as synaptic and secretory vesicles and insulin granules, in which specific ZNT proteins are localized. ZNT5 and ZNT6 form heterodimers to transport zinc. ZIP transporters move zinc in the opposite direction. In contrast to the specific localization of ZIP9, ZIP13, and ZIP7 in the Golgi apparatus and the ER, the subcellular location of ZNT5-ZNT6 heterodimers has not been definitively determined.

Figure 2. Phylogeny of ZNT and ZIP transporters. The neighbor-joining phylogenetic tree was constructed using ClustalW (http://clustalw.ddbj.nig.ac.jp/index.php?lang=en) protein alignment. (A) ZNT and (B) ZIP transporters. Subfamilies and subgroups are designated according to the text.
Figure 2. Phylogeny of ZNT and ZIP transporters. The neighbor-joining phylogenetic tree was constructed using ClustalW (http://clustalw.ddbj.nig.ac.jp/index.php?lang=en) protein alignment. (A) ZNT and (B) ZIP transporters. Subfamilies and subgroups are designated according to the text.

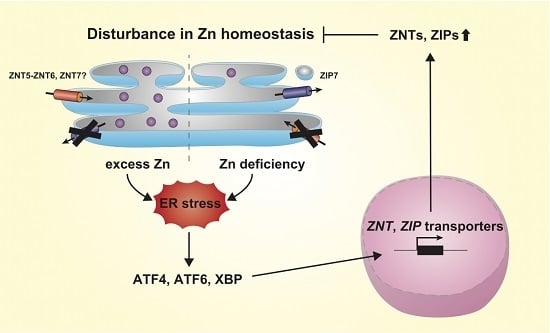
Figure 3. Model of feedback regulation for the maintenance of zinc homeostasis in the ER (in the early secretory pathway). A disturbance in zinc homeostasis, such as zinc deficiency or zinc overload, in the ER (and perhaps in the early secretory pathway) induces homeostatic ER stress. The unfolded protein response (UPR) leads to the activation of transcription factors such as ATF4, ATF6, and XBP1, and increases the transcription of several ZNT and ZIP transporter genes. These activities of ZNT and ZIP transporters then contribute to the maintenance of zinc homeostasis in the ER (and in the early secretory pathway), and thus attenuate homeostatic ER stress. Zn: zinc.
Figure 3. Model of feedback regulation for the maintenance of zinc homeostasis in the ER (in the early secretory pathway). A disturbance in zinc homeostasis, such as zinc deficiency or zinc overload, in the ER (and perhaps in the early secretory pathway) induces homeostatic ER stress. The unfolded protein response (UPR) leads to the activation of transcription factors such as ATF4, ATF6, and XBP1, and increases the transcription of several ZNT and ZIP transporter genes. These activities of ZNT and ZIP transporters then contribute to the maintenance of zinc homeostasis in the ER (and in the early secretory pathway), and thus attenuate homeostatic ER stress. Zn: zinc.

Figure 4. ZNT5-ZNT6 heterodimers and ZNT7 homodimers function to activate tissue-nonspecific ALP (TNAP) in a two-step mechanism. TNAP is specifically activated in a two-step mechanism involving ZNT5-ZNT6 heterodimers and ZNT7 homodimers as follows: first, the apo-form of TNAP is stabilized by either ZNT5-ZNT6 heterodimers or ZNT7 homodimers; second, the apo-form of TNAP is converted to the holo-form by zinc metallation. The PP-motifs in ZNT5 and ZNT7 likely play important roles in this process (see text). TNAP possesses a bimetallic core, is dimeric, and is localized to the plasma membrane via a glycophosphatidylinositol anchor. The subcellular localizations of ZNT5-ZNT6 heterodimers or ZNT7 homodimers have not been well defined. Zn: zinc.
Figure 4. ZNT5-ZNT6 heterodimers and ZNT7 homodimers function to activate tissue-nonspecific ALP (TNAP) in a two-step mechanism. TNAP is specifically activated in a two-step mechanism involving ZNT5-ZNT6 heterodimers and ZNT7 homodimers as follows: first, the apo-form of TNAP is stabilized by either ZNT5-ZNT6 heterodimers or ZNT7 homodimers; second, the apo-form of TNAP is converted to the holo-form by zinc metallation. The PP-motifs in ZNT5 and ZNT7 likely play important roles in this process (see text). TNAP possesses a bimetallic core, is dimeric, and is localized to the plasma membrane via a glycophosphatidylinositol anchor. The subcellular localizations of ZNT5-ZNT6 heterodimers or ZNT7 homodimers have not been well defined. Zn: zinc.

Table 1. Relationship between properties of zinc-requiring enzymes and ZNT transporters *.
Table 1. Relationship between properties of zinc-requiring enzymes and ZNT transporters *.
| Enzyme | Active Site | Zn Coordination Residues | ZNTs Involved in Activation | Defects Caused by Loss of ZNTs |
|---|---|---|---|---|
| ALP | Bimetallic center | Asp, His, His for Zn1His, Asp, Asp for Zn2 | ZNT5-ZNT6, ZNT7 | Loss of enzyme activityProtein destabilization |
| ATXCAIX | Bimetallic centerMononuclear Zn | Asp, His, His for Zn1,His, Asp, Asp for Zn2His, His, His | ZNT5-ZNT6, ZNT7ZNT4,ZNT5ZNT6,ZNT7 | Loss of enzyme activityDecreases in enzyme activity |
| MMP-2MMP-9 | Mononuclear Zn ** | His, His, His | ZNT5-ZNT6, ZNT7 | Loss of enzyme activityProtein destabilization *** |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kambe, T.; Matsunaga, M.; Takeda, T.-a. Understanding the Contribution of Zinc Transporters in the Function of the Early Secretory Pathway. Int. J. Mol. Sci. 2017, 18, 2179. https://doi.org/10.3390/ijms18102179
AMA Style
Kambe T, Matsunaga M, Takeda T-a. Understanding the Contribution of Zinc Transporters in the Function of the Early Secretory Pathway. International Journal of Molecular Sciences. 2017; 18(10):2179. https://doi.org/10.3390/ijms18102179
Chicago/Turabian Style
Kambe, Taiho, Mayu Matsunaga, and Taka-aki Takeda. 2017. "Understanding the Contribution of Zinc Transporters in the Function of the Early Secretory Pathway" International Journal of Molecular Sciences 18, no. 10: 2179. https://doi.org/10.3390/ijms18102179
APA Style
Kambe, T., Matsunaga, M., & Takeda, T. -a. (2017). Understanding the Contribution of Zinc Transporters in the Function of the Early Secretory Pathway. International Journal of Molecular Sciences, 18(10), 2179. https://doi.org/10.3390/ijms18102179
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.
Article Metrics
Article Access Statistics
For more information on the journal statistics, click here.
Multiple requests from the same IP address are counted as one view.
We use cookies on our website to ensure you get the best experience.
Read more about our cookies here.